
Effect of a Cytoprotective Dose of Dehydroleucodine, Xanthatin, and 3-Benzyloxymethyl-5H-furan-2-one on Gastric Mucosal Lesions Induced by Mast Cell Activation


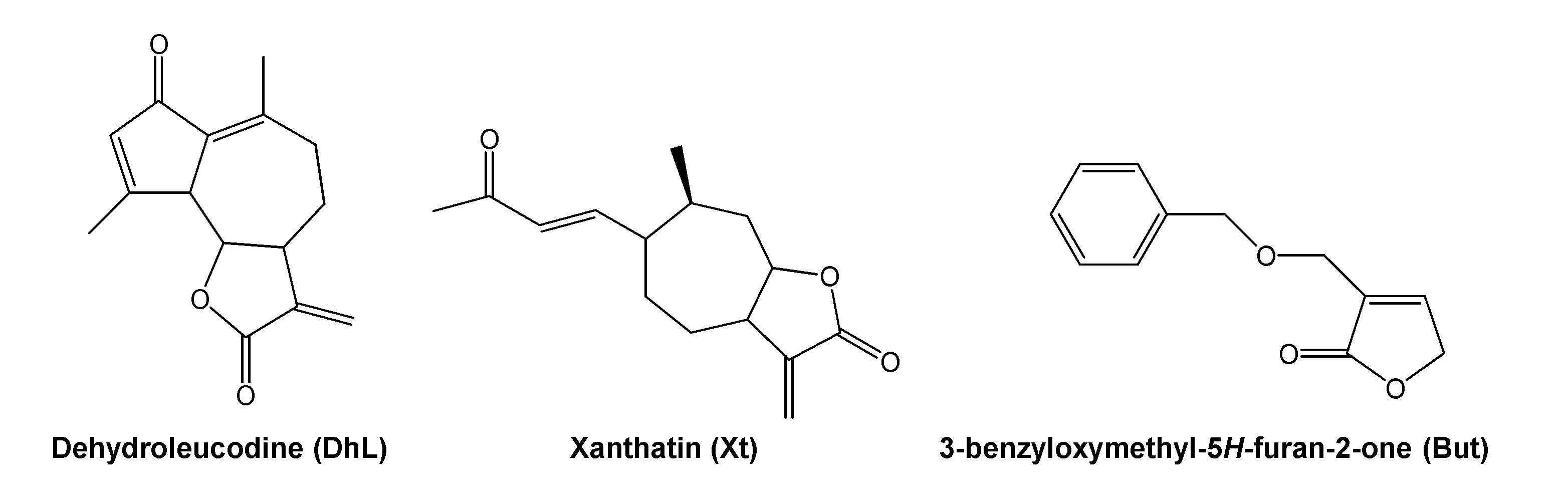{kind=link}
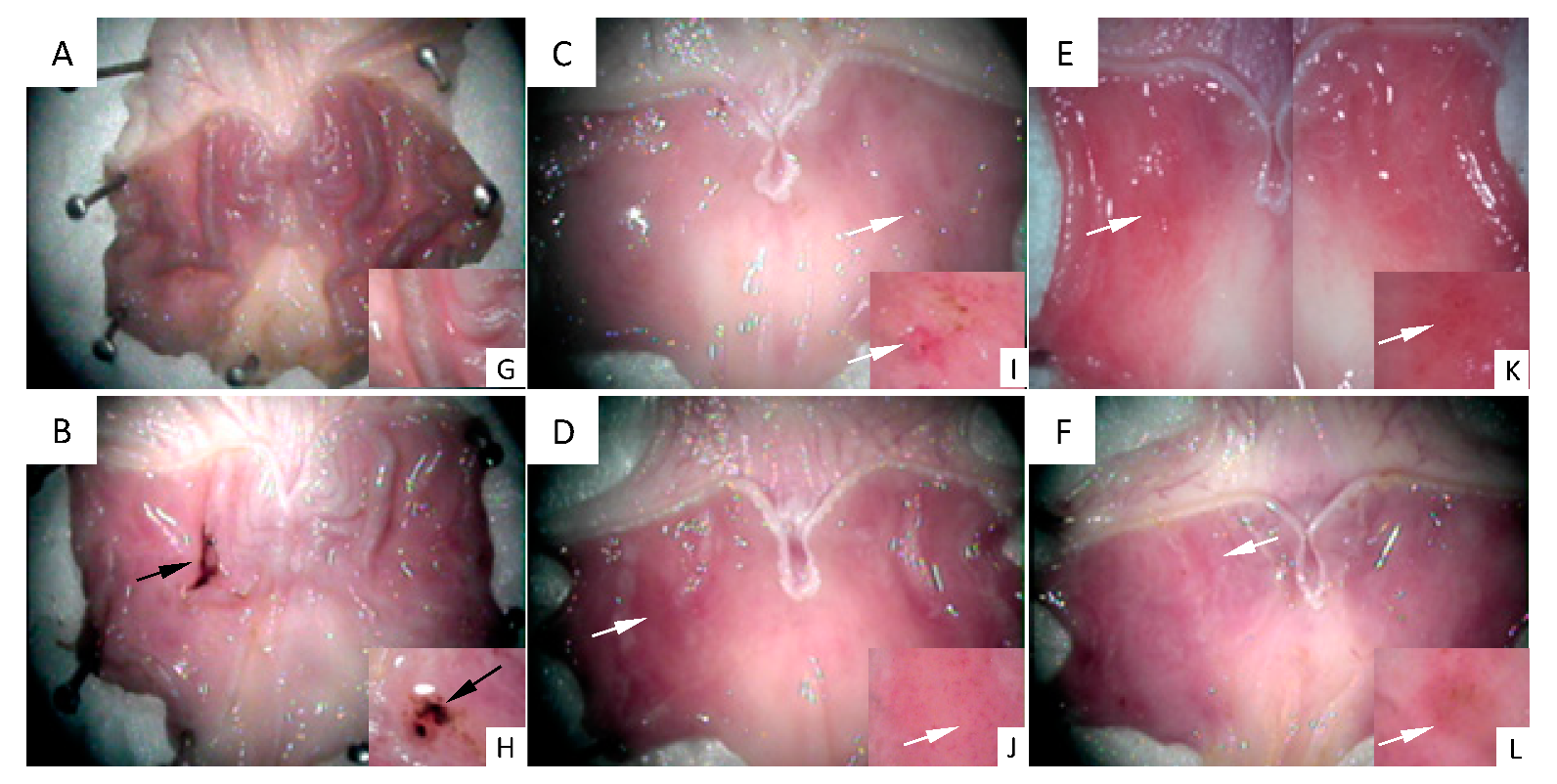{kind=link}
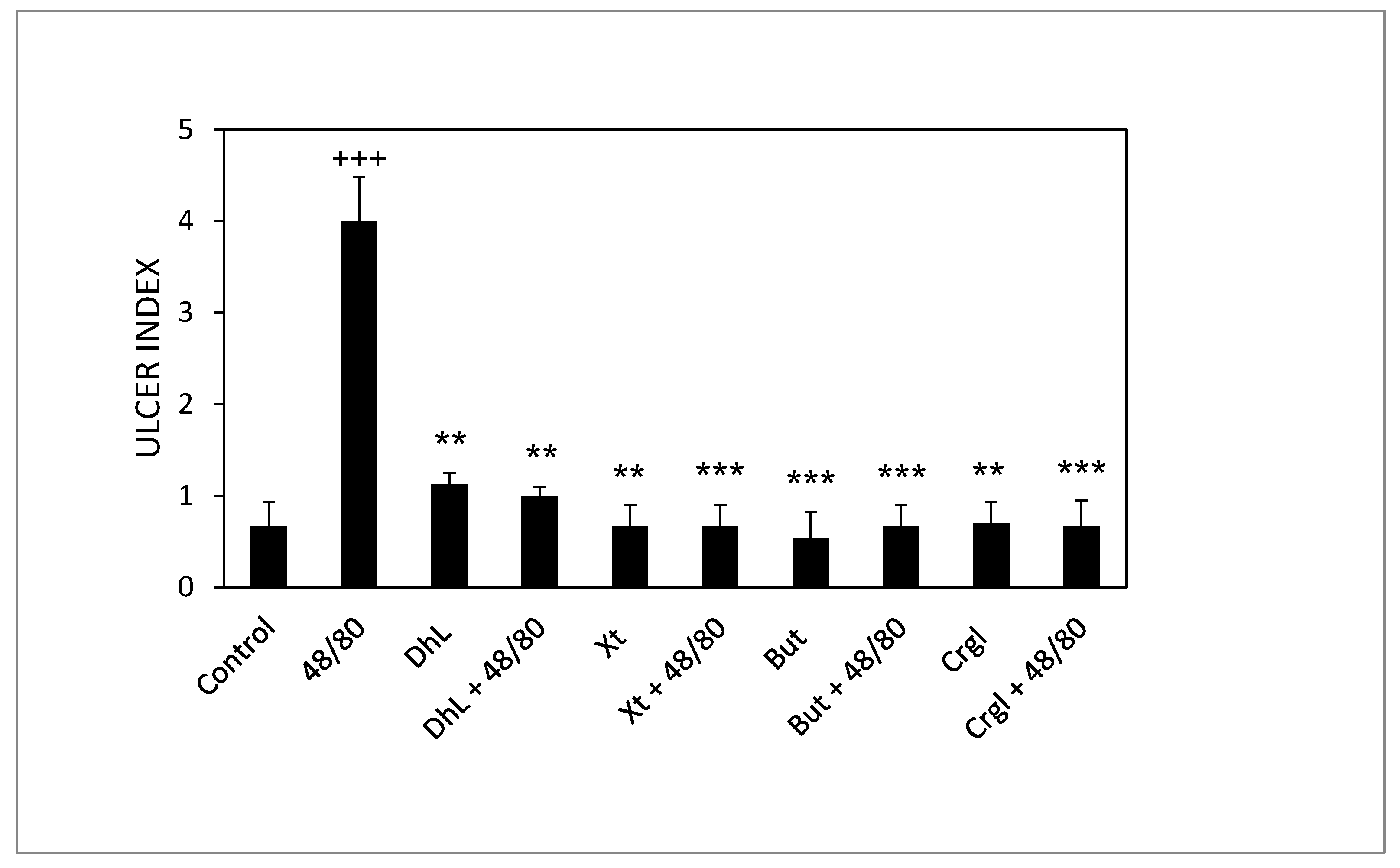{kind=link}
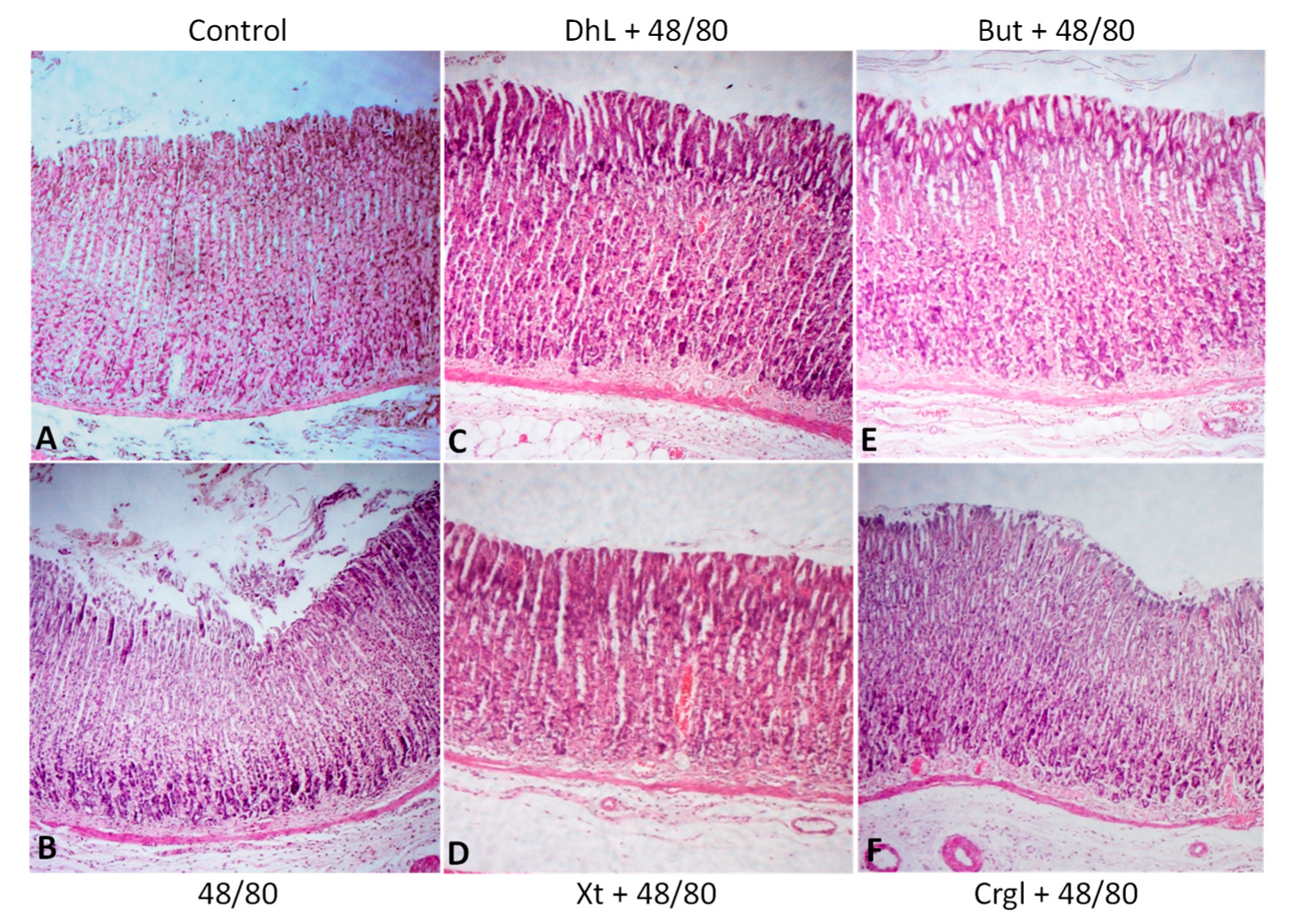{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Macroscopic Evaluation of Stomachs and Ulcer Index Values
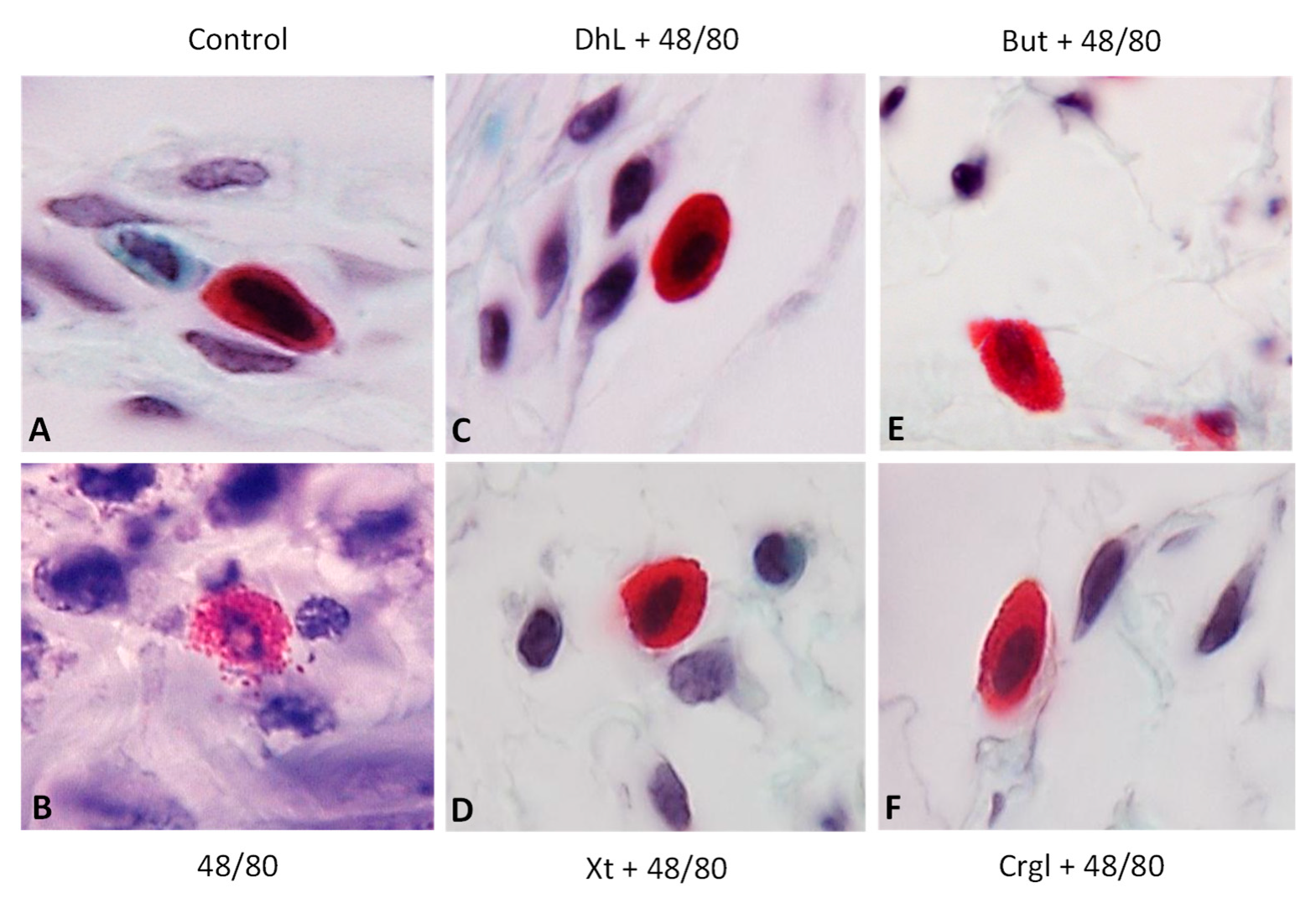
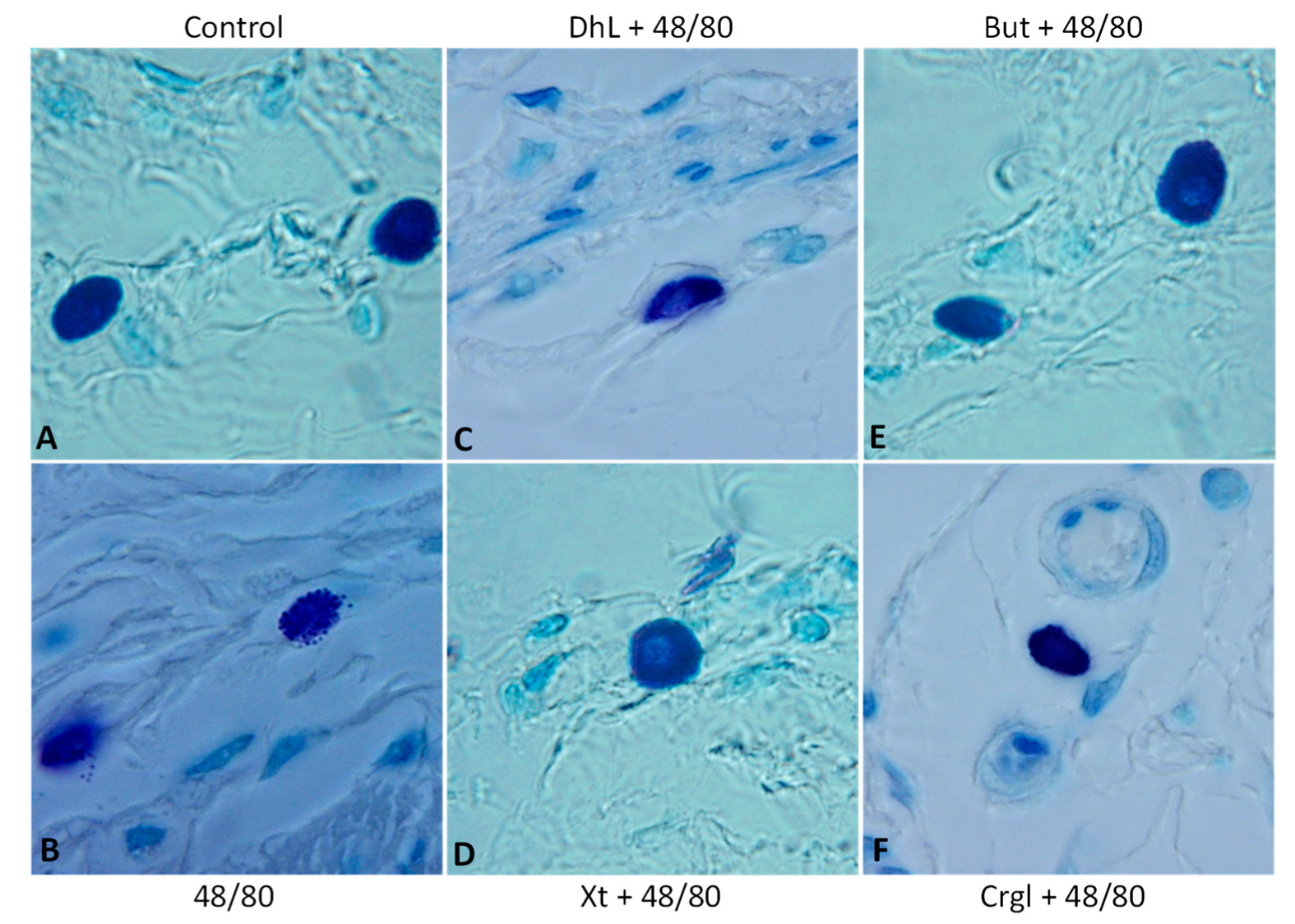
2.2. Histological Analysis
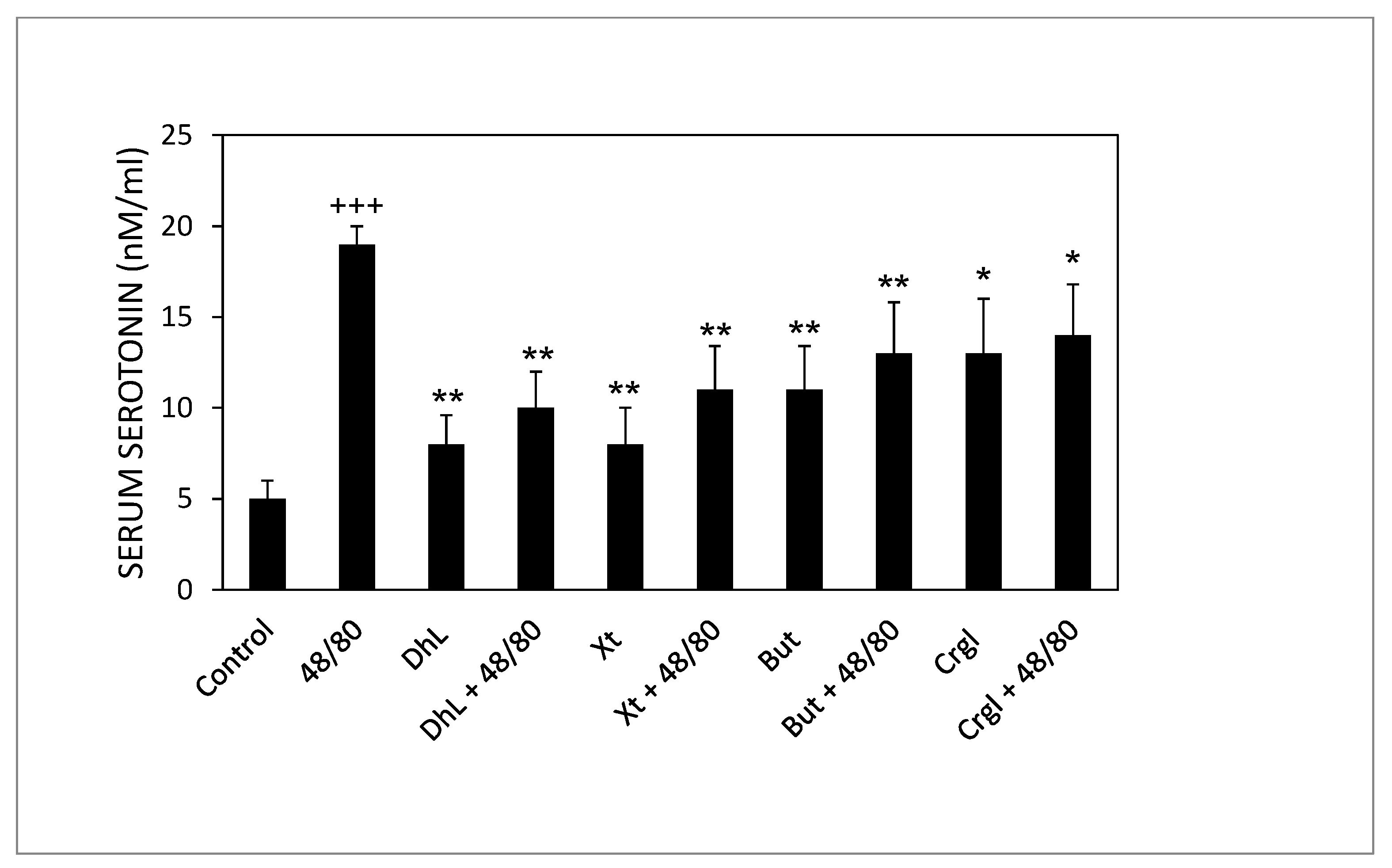
2.3. Serum Serotonin Concentrations
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Animals
4.3. Isolation and Purification of Dehydroleucodine
4.4. Isolation and Purification of Xanthatin
4.5. Preparation of 3-Benzyloxymethyl-5H-furan-2-one
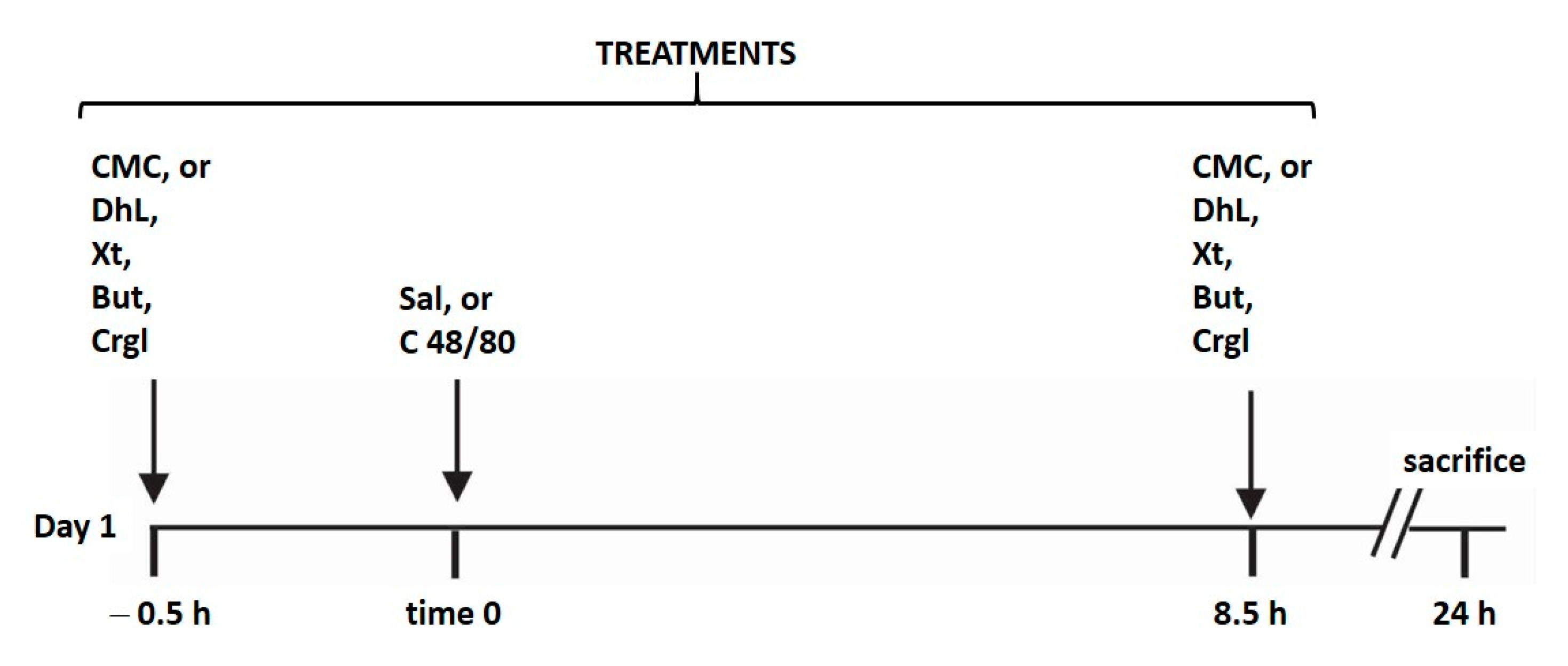
4.6. General Protocol and Induction of Gastric Ulcer by Mast Cell Activation
4.7. Scoring System
4.8. Light Microscopy
4.9. Determination of Serum Serotonin
4.10. Statistics
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Ardalani, H.; Hadipanah, A.; Sahebkar, A. Medicinal Plants in the Treatment of Peptic Ulcer Disease: A Review. Mini Rev. Med. Chem. 2019, 20, 662–702. [Google Scholar] [CrossRef]
- Szabo, S. “Gastric cytoprotection” is still relevant. J. Gastroenterol. Hepatol. 2014, 29, 124–132. [Google Scholar] [CrossRef] [Green Version]
- Choi, S.R.; Lee, S.A.; Kim, Y.J.; Ok, C.Y.; Lee, H.J.; Hahm, K.B. Role of heat shock proteins in gastric inflammation and ulcer healing. J. Physiol. Pharmacol. 2009, 60, 5–17. [Google Scholar] [PubMed]
- Narayanan, M.; Reddy, K.M.; Marsicano, E. Peptic Ulcer Disease and Helicobacter pylori infection. Sci. Med. 2018, 115, 219–224. [Google Scholar]
- Kuna, L.; Jakab, J.; Smolic, R.; Raguz-Lucic, N.; Vcev, A.; Smolic, M. Peptic Ulcer Disease: A Brief Review of Conventional Therapy and Herbal Treatment Options. J. Clin. Med. 2019, 8, 179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sverdén, E.; Agréus, L.; Dunn, J.M.; Lagergren, J. Peptic ulcer disease. BMJ 2019, 367, 1–8. [Google Scholar] [CrossRef]
- Malfertheiner, P.; Schulz, C. Peptic Ulcer: Chapter Closed? Dig. Dis. 2020, 38, 112–116. [Google Scholar] [CrossRef]
- Keita, Å.V.; Söderholm, J.D. Mucosal permeability and mast cells as targets for functional gastrointestinal disorders. Curr. Opin. Pharmacol. 2018, 43, 66–71. [Google Scholar] [CrossRef]
- Ramsay, D.B.; Stephen, S.; Borum, M.; Voltaggio, L.; Doman, D.B. Mast cells in gastrointestinal disease. Gastroenterol. Hepatol. 2010, 6, 772–777. [Google Scholar]
- Hodges, K.; Kennedy, L.; Meng, F.; Alpini, G.; Francis, H. Mast cells, disease and gastrointestinal cancer: A comprehensive review of recent findings. Bone 2008, 23, 1–7. [Google Scholar]
- Bischoff, S.C. Mast cells in gastrointestinal disorders. Eur. J. Pharmacol. 2016, 778, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Andre, F.; Gillon, J.; Andre, C.; Fournier, S. Role of mast mediators in pathogenesis of anaphylactic gastric ulcer. Digestion 1983, 28, 108–113. [Google Scholar] [CrossRef]
- Sammarco, G.; Gallo, G.; Vescio, G.; Picciariello, A.; De Paola, G.; Trompetto, M.; Currò, G.; Ammendola, M. Mast Cells, microRNAs and Others: The Role of Translational Research on Colorectal Cancer in the Forthcoming Era of Precision Medicine. J. Clin. Med. 2020, 9, 2852. [Google Scholar] [CrossRef]
- Cildir, G.; Yip, K.H.; Pant, H.; Tergaonkar, V.; Lopez, A.F.; Tumes, D.J. Understanding mast cell heterogeneity at single cell resolution. Trends Immunol. 2021, 1–13. [Google Scholar]
- Dellon, E.S.; Chen, X.; Miller, C.R.; Fritchie, K.J.; Rubinas, T.C.; Woosley, J.T.; Shaheen, N. Tryptase staining of mast cells may differentiate eosinophilic esophagitis from gastroesophageal reflux disease. Physiol. Behav. 2019, 176, 139–148. [Google Scholar] [CrossRef] [Green Version]
- Caruso, R.A.; Parisi, A.; Crisafulli, C.; Bonanno, A.; Lucianò, R.; Branca, G.; Scardigno, M.; Fedele, F. Intraepithelial infiltration by mast cells in human Helicobacter pylori active gastritis. Ultrastruct. Pathol. 2011, 35, 251–255. [Google Scholar] [CrossRef] [PubMed]
- Jafarzadeh, A.; Nemati, M.; Jafarzadeh, S. The important role played by chemokines influence the clinical outcome of Helicobacter pylori infection. Life Sci. 2019, 231, 116688. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, S.; Krishnan, B.; Ota, H.; Segura, A.M.; Hattori, T.; Graham, D.Y.; Genta, R.M. Mast cell involvement in gastritis with or without Helicobacter pylori infection. Gastroenterology 1997, 113, 746–754. [Google Scholar] [CrossRef]
- Kobayashi, T.; Yabana, T. Mechanisms of direct gastric mucosal damage by NSAIDs. Nihon Rinsho 2007, 65, 1803–1806. [Google Scholar]
- Konturek, P.; Brzozowski, T.; Konturek, S. Stress and the gut: Pathophysiology, clinical consequences, diagnostic approach and treatment options. J. Physiol. Pharmacol. 2011, 62, 591–599. [Google Scholar]
- Penissi, A.B.; Fogal, T.H.; Guzmán, J.A.; Piezzi, R.S. Gastroduodenal mucosal protection induced by dehydroleucodine: Mucus secretion and role of monoamines. Dig. Dis. Sci. 1998, 43, 791–798. [Google Scholar] [CrossRef]
- Penissi, A.B.; Piezzi, R.S. Effect of dehydroleucodine on mucus production: A quantitative study. Dig. Dis. Sci. 1999, 44, 708–712. [Google Scholar] [CrossRef]
- Favier, L.S.; María, A.O.M.; Wendel, G.H.; Borkowski, E.J.; Giordano, O.S.; Pelzer, L.; Tonn, C.E. Anti-ulcerogenic activity of xanthanolide sesquiterpenes from Xanthium cavanillesii in rats. J. Ethnopharmacol. 2005, 100, 260–267. [Google Scholar] [CrossRef]
- Ceñal, J.P.; Carreras, C.R.; Tonn, C.E.; Padrón, J.I.; Ramírez, M.A.; Díaz, D.D.; García-Tellado, F.; Martín, V.S. Acid-mediated highly regioselective oxidation of substituted furans: A simple and direct entry to substituted butenolides. Synlett 2005, 1575–1578. [Google Scholar]
- Penissi, A.B.; Vera, M.E.; Mariani, M.L.; Rudolph, M.I.; Ceñal, J.P.; de Rosas, J.C.; Fogal, T.H.; Tonn, C.E.; Favier, L.S.; Giordano, O.S.; et al. Novel anti-ulcer α,β-unsaturated lactones inhibit compound 48/80-induced mast cell degranulation. Eur. J. Pharmacol. 2009, 612, 122–130. [Google Scholar] [CrossRef]
- Vera, M.E.; Persia, F.A.; Mariani, M.L.; Rudolph, M.I.; Fogal, T.H.; Ceñal, J.P.; Favier, L.S.; Tonn, C.E.; Penissi, A.B. Activation of human leukemic mast cell line LAD2 is modulated by dehydroleucodine and xanthatin. Leuk. Lymphoma 2012, 53, 1795–1803. [Google Scholar] [CrossRef]
- Coll, R.C.; Vargas, P.M.; Mariani, M.L.; Penissi, A.B. Natural α,β-unsaturated lactones inhibit neuropeptide-induced mast cell activation in an in vitro model of neurogenic inflammation. Inflamm. Res. 2020, 69, 1039–1051. [Google Scholar] [CrossRef] [PubMed]
- Yasuhiro, T.; Konaka, A.; Kato, S.; Takeuchi, K. Gastric mucosal damage induced by compound 48/80: Roles of serotonin and nitric oxide. J. Gastroenterol. Hepatol. 1998, 13, 1099–1106. [Google Scholar] [CrossRef]
- Marazzi-Uberti, E.; Turba, C. Experimental Gastric Ulcer Caused by Histamine in Guinea-Pigs. Med. Exp. 1961, 292, 284–292. [Google Scholar] [CrossRef]
- Koretou, O. Relationship between the staining property of mast cell granule with alcian blue-safranin O and toluidine blue O, and the content of mast cell protease I in the granule of rat peritoneal mast cell. Acta Histochem. Cytochem. 1988, 21, 25–32. [Google Scholar] [CrossRef] [Green Version]
- Abebaw, M.; Mishra, B.; Gelayee, D.A. Evaluation of anti-ulcer activity of the leaf extract of Osyris quadripartita Decne. (Santalaceae) in rats. J. Exp. Pharmacol. 2017, 9, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, A.P.; Bajpai, A.; Chandra, S. A Comprehensive Review on the Screening Models for the Pharmacological Assessment of Antiulcer Drugs. Curr. Clin. Pharmacol. 2019, 14, 175–196. [Google Scholar] [CrossRef]
- Zhang, J.; Fu, Z.L.; Chu, Z.X.; Song, B.W. Gastroprotective Activity of the Total Flavones from Abelmoschus manihot (L.) Medic Flowers. Evid. Based Complement. Altern. Med. 2020, 2020, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferjan, I.; Čarman-Kržan, M.; Erjavec, F. Comparison of histamine and serotonin release from rat peritoneal mast cells induced by nerve growth factor and compound 48/80. Inflamm. Res. 1997, 46, 23–24. [Google Scholar] [CrossRef]
- Arisawa, T.; Nakazawa, S.; Asai, J.; Tsukamoto, Y. Histological evaluation of mast cells in rat gastric mucosal lesions induced by compound 48/80. Digestion 1989, 43, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Ohta, Y.; Kobayashi, T.; Inui, K..; Yoshino, J.; Nakazawa, S. Protective effect of teprenone against acute gastric mucosal lesions induced by compound 48/80, a mast cell degranulator, in rats. J. Pharmacol. Sci. 2003, 93, 337–346. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, T.; Ohta, Y.; Yoshino, J. Preventive effect of ebselen on acute gastric mucosal lesion development in rats treated with compound 48/80. Eur. J. Pharmacol. 2001, 414, 271–279. [Google Scholar] [CrossRef]
- Kobayashi, T.; Ohta, Y.; Inui, K.; Yoshino, J.; Nakazawa, S. Protective effect of omeprazole against acute gastric mucosal lesions induced by compound 48/80, a mast cell degranulator, in rats. Pharmacol. Res. 2002, 46, 75–84. [Google Scholar] [CrossRef]
- Akagi, M.; Matsui, N.; Mochizuki, S.; Tasaka, K. Inhibitory effect of egualen sodium: A new stable derivative of azulene on histamine release from mast cell-like cells in the stomach. Pharmacology 2001, 63, 203–209. [Google Scholar] [CrossRef]
- Ohta, Y.; Kobayashi, T.; Hayashi, T.; Inui, K.; Yoshino, J. Saikokeishito extract prevents progression of acute gastric mucosal lesions induced by compound 48/80, a mast cell degranulator, in rats. J. Tradit. Med. 2006, 23, 101–111. [Google Scholar]
- Ohta, Y.; Kamiya, Y.; Imai, Y.; Arisawa, T.; Nakano, H. Plaunotol prevents the progression of acute gastric mucosal lesions induced by compound 48/80, a mast cell degranulator, in rats. Pharmacology 2005, 74, 182–192. [Google Scholar] [CrossRef] [PubMed]
- Ohta, Y.; Kobayashi, T.; Imai, Y.; Inui, K.; Yoshino, J.; Nakazawa, S. Effect of oral vitamin E administration on acute gastric mucosal lesion progression in rats treated with compound 48/80, a mast cell degranulator. Biol. Pharm. Bull. 2006, 29, 675–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vera, M.E.; Mariani, M.L.; Aguilera, C.; Penissi, A.B. Effect of a Cytoprotective Dose of Dehydroleucodine, Xanthatin, and 3-Benzyloxymethyl-5H-furan-2-one on Gastric Mucosal Lesions Induced by Mast Cell Activation. Int. J. Mol. Sci. 2021, 22, 5983. https://doi.org/10.3390/ijms22115983
Vera ME, Mariani ML, Aguilera C, Penissi AB. Effect of a Cytoprotective Dose of Dehydroleucodine, Xanthatin, and 3-Benzyloxymethyl-5H-furan-2-one on Gastric Mucosal Lesions Induced by Mast Cell Activation. International Journal of Molecular Sciences. 2021; 22(11):5983. https://doi.org/10.3390/ijms22115983
Chicago/Turabian StyleVera, Mariano Ezequiel, María Laura Mariani, Cristina Aguilera, and Alicia Beatriz Penissi. 2021. "Effect of a Cytoprotective Dose of Dehydroleucodine, Xanthatin, and 3-Benzyloxymethyl-5H-furan-2-one on Gastric Mucosal Lesions Induced by Mast Cell Activation" International Journal of Molecular Sciences 22, no. 11: 5983. https://doi.org/10.3390/ijms22115983
APA StyleVera, M. E., Mariani, M. L., Aguilera, C., & Penissi, A. B. (2021). Effect of a Cytoprotective Dose of Dehydroleucodine, Xanthatin, and 3-Benzyloxymethyl-5H-furan-2-one on Gastric Mucosal Lesions Induced by Mast Cell Activation. International Journal of Molecular Sciences, 22(11), 5983. https://doi.org/10.3390/ijms22115983