
Outcomes of Deferoxamine Action on H2O2-Induced Growth Inhibition and Senescence Progression of Human Endometrial Stem Cells

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
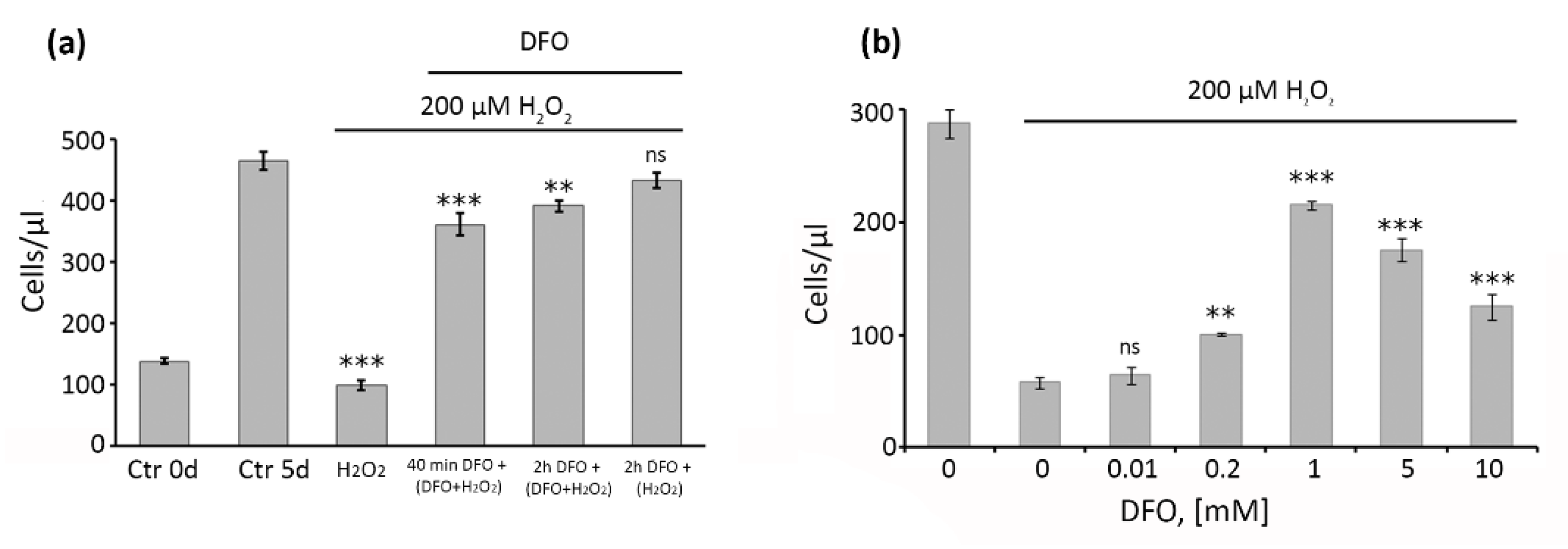
2.1. Pretreatment with 1 mM DFO for 2 h Is Optimal to Protect hMESCs from the Oxidative Stress-Induced Growth Arrest
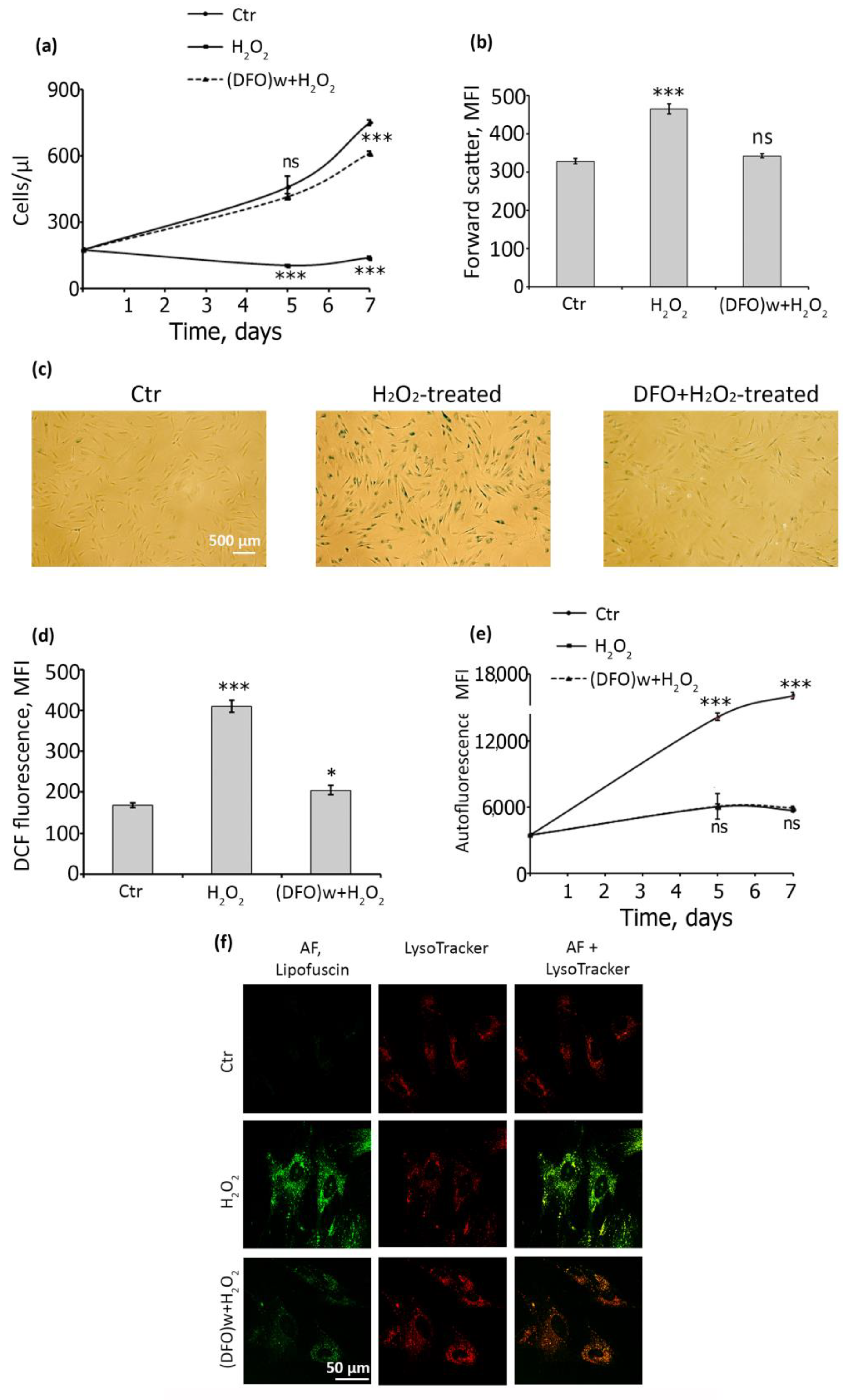
2.2. DFO Pretreatment Is Sufficient to Prevent Oxidative Stress-Induced Senescence of hMESCs
2.3. DFO-Rescued hMESCs Retain the Ability to Respond towards Oxidative Stress via Senescence Induction
2.4. DFO Protects DNA Damage by Inhibiting the Formation of Hydroxyl Radicals in H2O2-Treated hMESCs
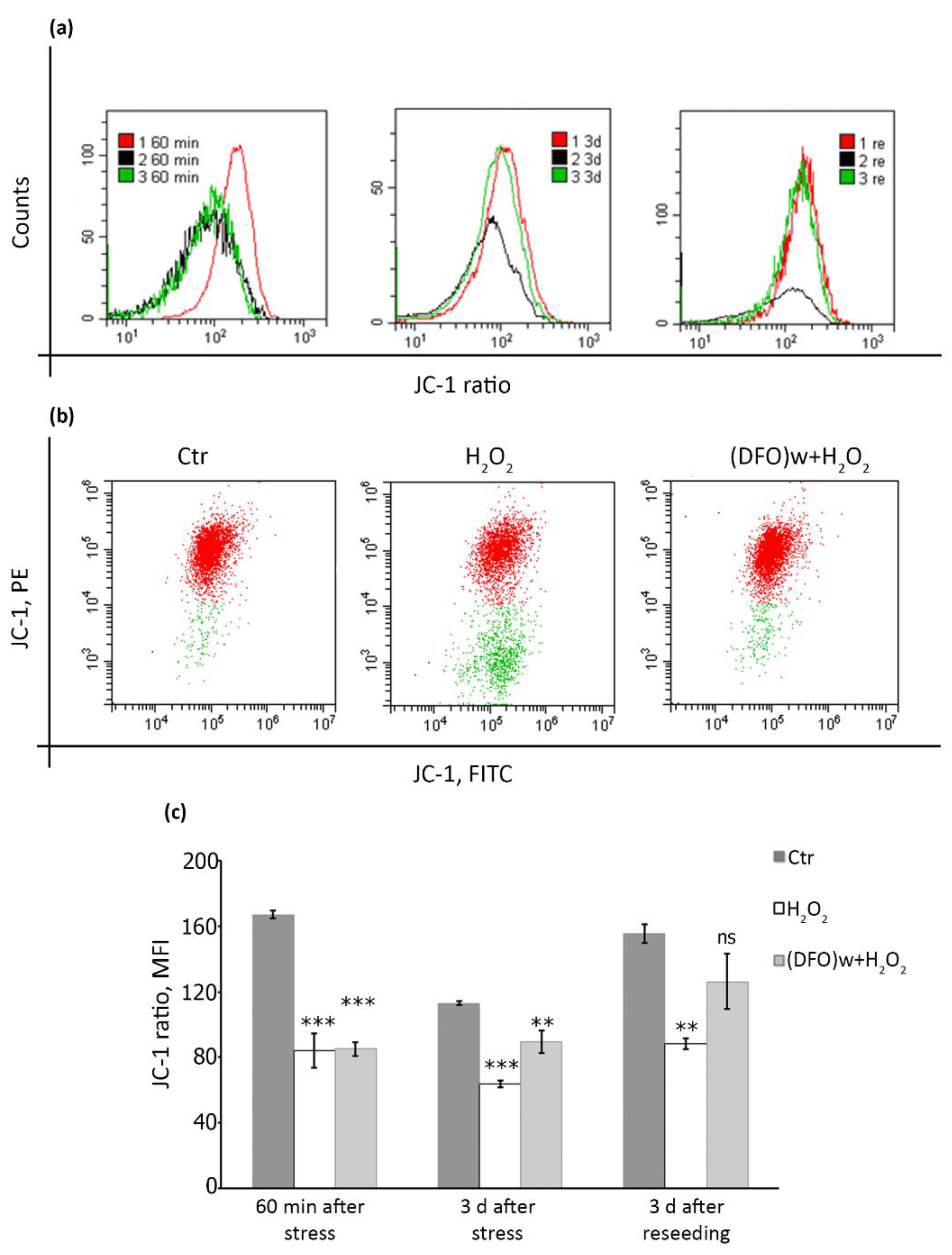
2.5. DFO Pretreatment Is Able to Restore Mitochondrial Membrane Potential during the Progression of Oxidative Stress-Induced Senescence of hMESCs
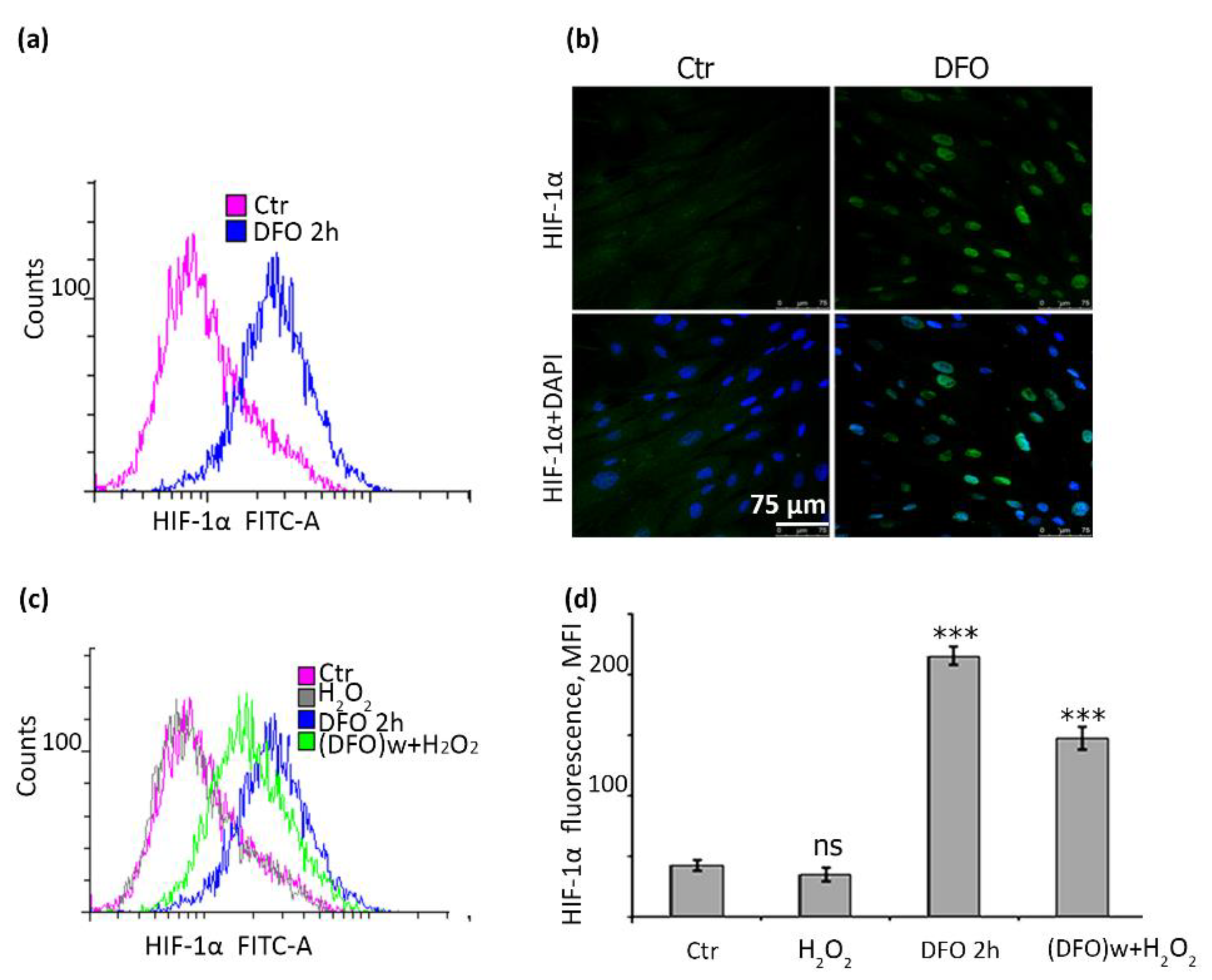
2.6. DFO Causes a Significant Accumulation of HIF-1α in hMESCs
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Experiment Design and Cell Treatments
4.3. Flow Cytometry Assay
4.3.1. Viability, Autofluorescence, Forward Scattering Assay
4.3.2. Cell Cycle Analysis
4.3.3. Qualification of Intracellular ROS, Mitochondrial Mass, Mitochondrial Membrane Potential
4.3.4. Determination of SA-β-Gal activity
4.3.5. Determination of DNA Damage, HIF-1α Accumulation and Cyclin D1
4.4. Immunofluorescent Assay
4.5. Live Cell Imaging and Confocal Microscopy Detection of Lipofuscin within Lysosomal Compartment
4.6. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fernández Vallone, V.B.; Romaniuk, M.A.; Choi, H.; Labovsky, V.; Otaegui, J.; Chasseing, N.A. Mesenchymal stem cells and their use in therapy: What has been achieved? Differentiation 2013, 85, 1–10. [Google Scholar] [CrossRef]
- Friedenstein, A.J.; Petrakova, K.V.; Kurolesova, A.I.; Frolova, G.P. Heterotopic of bone marrow. Analysis of precursor cells for osteogenic and hematopoietic tissues. Transplantation 1968, 6, 230–247. [Google Scholar] [CrossRef]
- Berebichez-Fridman, R.; Montero-Olvera, P.R. Sources and Clinical Applications of Mesenchymal Stem Cells: State-of-the-art review. Sultan Qaboos Univ. Med. J. 2018, 18, e264–e277. [Google Scholar] [CrossRef] [Green Version]
- Gargett, C.E.; Masuda, H. Adult stem cells in the endometrium. Mol. Hum. Reprod. 2010, 16, 818–834. [Google Scholar] [CrossRef]
- Xu, Y.; Zhu, H.; Zhao, D.; Tan, J. Endometrial stem cells: Clinical application and pathological roles. Int. J. Clin. Exp. Med. 2015, 8, 22039–22044. [Google Scholar]
- Zemelko, V.I.; Grinchuk, T.M.; Domnina, A.P.; Artzibasheva, I.V.; Zenin, V.V.; Kirsanov, A.A.; Bichevaia, N.K.; Korsak, V.S.; Nikolsky, N.N. Multipotent mesenchymal stem cells of desquamated endometrium: Isolation, characterization, and application as a feeder layer for maintenance of human embryonic stem cells. Cell Tissue Biol. 2012, 6, 1–11. [Google Scholar] [CrossRef]
- Ulrich, D.; Muralitharan, R.; Gargett, C.E. Toward the use of endometrial and menstrual blood mesenchymal stem cells for cell-based therapies. Expert. Opin. Biol. Ther. 2013, 13, 1387–1400. [Google Scholar] [CrossRef]
- Geng, Y.J. Molecular mechanisms for cardiovascular stem cell apoptosis and growth in the hearts with atherosclerotic coronary disease and ischemic heart failure. Ann. N. Y. Acad. Sci. 2003, 1010, 687–697. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.A.; Shim, W.; Paik, M.J.; Lee, S.C.; Shin, J.Y.; Ahn, Y.H.; Park, K.; Kim, J.H.; Choi, S.; Lee, G. Analysis of changes in the viability and gene expression profiles of human mesenchymal stromal cells over time. Cytotherapy 2009, 11, 688–697. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.B.; Jiang, J.; Gui, C.; Hu, X.Y.; Xiang, M.X.; Wang, J.A. Angiopoietin-1 protects mesenchymal stem cells against serum deprivation and hypoxia-induced apoptosis through the PI3K/Akt pathway. Acta Pharmacol. Sin. 2008, 29, 815–822. [Google Scholar] [CrossRef] [Green Version]
- Leibacher, J.; Dauber, K.; Ehser, S.; Brixner, V.; Kollar, K.; Vogel, A.; Spohn, G.; Schäfer, R.; Seifried, E.; Henschler, R. Human mesenchymal stromal cells undergo apoptosis and fragmentation after intravenous application in immune-competent mice. Cytotherapy 2017, 19, 61–74. [Google Scholar] [CrossRef]
- Lee, S.; Choi, E.; Cha, M.J.; Hwang, K.C. Cell adhesion and long-term survival of transplanted mesenchymal stem cells: A prerequisite for cell therapy. Oxidative Med. Cell. Longev. 2015, 2015, 632902. [Google Scholar] [CrossRef] [Green Version]
- Khoshlahni, N.; Sagha, M.; Mirzapour, T.; Zarif, M.N.; Mohammadzadeh-Vardin, M. Iron depletion with deferoxamine protects bone marrow-derived mesenchymal stem cells against oxidative stress-induced apoptosis. Cell Stress Chaperones 2020, 25, 1059–1069. [Google Scholar] [CrossRef]
- Burova, E.; Borodkina, A.; Shatrova, A.; Nikolsky, N. Sublethal oxidative stress induces the premature senescence of human mesenchymal stem cells derived from endometrium. Oxidative Med. Cell. Longev. 2013, 2013, 474931. [Google Scholar] [CrossRef]
- Alekseenko, L.L.; Zemelko, V.I.; Domnina, A.P.; Lyublinskaya, O.G.; Zenin, V.V.; Pugovkina, N.A.; Kozhukharova, I.V.; Borodkina, A.V.; Grinchuk, T.M.; Fridlyanskaya, I.I.; et al. Sublethal heat shock induces premature senescence rather than apoptosis in human mesenchymal stem cells. Cell Stress Chaperones 2014, 19, 355–366. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Ding, Y.; Liu, Z.; Liang, X. Senescence in mesenchymal stem cells: Functional alterations, molecular mechanisms, and rejuvenation strategies. Front. Cell Dev. Biol. 2020, 8, 258. [Google Scholar] [CrossRef]
- Domnina, A.; Ivanova, J.; Alekseenko, L.; Kozhukharova, I.; Borodkina, A.; Pugovkina, N.; Smirnova, I.; Lyublinskaya, O.; Fridlyanskaya, I.; Nikolsky, N. Three-Dimensional Compaction Switches Stress Response Programs and Enhances Therapeutic Efficacy of Endometrial Mesenchymal Stem/Stromal Cells. Front. Cell Dev. Biol. 2020, 8, 473. [Google Scholar] [CrossRef]
- Turinetto, V.; Vitale, E.; Giachino, C. Senescence in human mesenchymal stem cells: Functional changes and implications in stem cell-based therapy. Int. J. Mol. Sci. 2016, 17, 1164. [Google Scholar] [CrossRef]
- Griukova, A.; Deryabin, P.; Shatrova, A.; Burova, E.; Severino, V.; Farina, A.; Nikolsky, N.; Borodkina, A. Molecular basis of senescence transmitting in the population of human endometrial stromal cells. Aging 2019, 11, 9912–9931. [Google Scholar] [CrossRef]
- Konala, V.B.; Mamidi, M.K.; Bhonde, R.; Das, A.K.; Pochampally, R.; Pal, R. The current landscape of the mesenchymal stromal cell secretome: A new paradigm for cell-free regeneration. Cytotherapy 2016, 18, 13–24. [Google Scholar] [CrossRef] [Green Version]
- Tran, C.; Damaser, M.S. Stem cells as drug delivery methods: Application of stem cell secretome for regeneration. Adv. Drug Deliv. Rev. 2015, 82–83, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Vizoso, F.J.; Eiro, N.; Cid, S.; Schneider, J.; Perez-Fernandez, R. Mesenchymal Stem Cell Secretome: Toward Cell-Free Therapeutic Strategies in Regenerative Medicine. Int. J. Mol. Sci. 2017, 18, 1852. [Google Scholar] [CrossRef] [Green Version]
- Persson, H.L.; Yu, Z.; Tirosh, O.; Eaton, J.W.; Brunk, U.T. Prevention of oxidant-induced cell death by lysosomotropic iron chelators. Free Radic. Biol. Med. 2003, 34, 1295–1305. [Google Scholar] [CrossRef]
- Kurz, T.; Leake, A.; Von Zglinicki, T.; Brunk, U.T. Relocalized redox-active lysosomal iron is an important mediator of oxidative-stress-induced DNA damage. Biochem. J. 2004, 378, 1039–1045. [Google Scholar] [CrossRef] [Green Version]
- Lloyd, J.B.; Cable, H.; Rice-Evans, C. Evidence that desferrioxamine cannot enter cells by passive diffusion. Biochem. Pharmacol. 1991, 41, 1361–1363. [Google Scholar] [CrossRef]
- Borodkina, A.; Shatrova, A.; Abushik, P.; Nikolsky, N.; Burova, E. Interaction between ROS dependent DNA damage, mitochondria and p38 MAPK underlies senescence of human adult stem cells. Aging 2014, 6, 481–495. [Google Scholar] [CrossRef] [Green Version]
- Doulias, P.T.; Christoforidis, S.; Brunk, U.T.; Galaris, D. Endosomal and lysosomal effects of desferrioxamine: Protection of HeLa cells from hydrogen peroxide-induced DNA damage and induction of cell-cycle arrest. Free Radic. Biol. Med. 2003, 35, 719–728. [Google Scholar] [CrossRef]
- Kurz, T.; Gustafsson, B.; Brunk, U.T. Intralysosomal iron chelation protects against oxidative stress-induced cellular damage. FEBS J. 2006, 273, 3106–3117. [Google Scholar] [CrossRef]
- Barbouti, A.; Doulias, P.T.; Zhu, B.Z.; Frei, B.; Galaris, D. Intracellular iron, but not copper, plays a critical role in hydrogen peroxide-induced DNA damage. Free Radic. Biol. Med. 2001, 31, 490–498. [Google Scholar] [CrossRef]
- Tenopoulou, M.; Doulias, P.T.; Barbouti, A.; Brunk, U.; Galaris, D. Role of compartmentalized redox-active iron in hydrogen peroxide-induced DNA damage and apoptosis. Biochem. J. 2005, 387, 703–710. [Google Scholar] [CrossRef]
- Terman, A.; Dalen, H.; Eaton, J.W.; Neuzil, J.; Brunk, U.T. Aging of cardiac myocytes in culture: Oxidative stress, lipofuscin accumulation, and mitochondrial turnover. Ann. N. Y. Acad. Sci. 2004, 1019, 70–77. [Google Scholar] [CrossRef]
- Bertolo, A.; Baur, M.; Guerrero, J.; Pötzel, T.; Stoyanov, J. Autofluorescence is a reliable in vitro marker of cellular senescence in human mesenchymal stromal cells. Sci. Rep. 2019, 9, 2074. [Google Scholar] [CrossRef] [Green Version]
- Leontieva, O.V.; Demidenko, Z.N.; Blagosklonny, M.V. MEK drives cyclin D1 hyperelevation during geroconversion. Cell Death Differ. 2013, 20, 1241–1249. [Google Scholar] [CrossRef] [Green Version]
- Campisi, J. Cellular senescence as a tumor-suppressor mechanism. Trends Cell Biol. 2001, 11, S27–S31. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.M.; Bartholomew, J.C.; Campisi, J.; Acosta, M.; Reagan, J.D.; Ames, B.N. Molecular analysis of H2O2-induced senescent-like growth arrest in normal human fibroblasts: p53 and Rb control G1 arrest but not cell replication. Biochem. J. 1998, 332, 43–50. [Google Scholar] [CrossRef] [Green Version]
- Sanmartín-Suárez, C.; Soto-Otero, R.; Sánchez-Sellero, I.; Méndez-Álvarez, E. Antioxidant properties of dimethyl sulfoxide and its viability as a solvent in the evaluation of neuroprotective antioxidants. J. Pharmacol. Toxicol. Methods 2011, 63, 209–215. [Google Scholar] [CrossRef]
- Shmulevich, R.; Krizhanovsky, V. Cell Senescence, DNA Damage, and Metabolism. Antioxid. Redox Signal. 2021, 34, 324–334. [Google Scholar] [CrossRef] [PubMed]
- Sivandzade, F.; Bhalerao, A.; Cucullo, L. Analysis of the mitochondrial membrane potential using the cationic JC-1 dye as a sensitive fluorescent probe. Bio Protoc. 2019, 9, e3128. [Google Scholar] [CrossRef] [PubMed]
- Tada-Oikawa, S.; Oikawa, S.; Kawanishi, M.; Yamada, M.; Kawanishi, S. Generation of hydrogen peroxide precedes loss of mitochondrial membrane potential during DNA alkylation-induced apoptosis. FEBS Lett. 1999, 442, 65–69. [Google Scholar] [CrossRef] [Green Version]
- Esfahani, M.; Karimi, F.; Afshar, S.; Niknazar, S.; Sohrabi, S.; Najafi, R. Prolyl hydroxylase inhibitors act as agents to enhance the efficiency of cell therapy. Expert. Opin. Biol. Ther. 2015, 15, 1739–1755. [Google Scholar] [CrossRef] [PubMed]
- Welford, S.M.; Bedogni, B.; Gradin, K.; Poellinger, L.; Broome Powell, M.; Giaccia, A.J. HIF1alpha delays premature senescence through the activation of MIF. Genes Dev. 2006, 20, 3366–3371. [Google Scholar] [CrossRef] [Green Version]
- Miethke, M.; Marahiel, M.A. Siderophore-based iron acquisition and pathogen control. Microbiol. Mol. Biol. Rev. 2007, 71, 413–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franchini, M.; Gandini, G.; de Gironcoli, M.; Vassanelli, A.; Borgna-Pignatti, C.; Aprili, G. Safety and efficacy of subcutaneous bolus injection of deferoxamine in adult patients with iron overload. Blood 2000, 95, 2776–2779. [Google Scholar] [CrossRef]
- Najafi, R.; Sharifi, A.M. Deferoxamine preconditioning potentiates mesenchymal stem cell homing in vitro and in streptozotocin-diabetic rats. Expert. Opin. Biol. Ther. 2013, 13, 959–972. [Google Scholar] [CrossRef]
- Peyvandi, A.A.; Abbaszadeh, H.A.; Roozbahany, N.A.; Pourbakht, A.; Khoshsirat, S.; Niri, H.H.; Peyvandi, H.; Niknazar, S. Deferoxamine promotes mesenchymal stem cell homing in noise-induced injured cochlea through PI3K/AKT pathway. Cell Prolif. 2018, 51, e12434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nouri, F.; Salehinejad, P.; Nematollahi-Mahani, S.N.; Kamarul, T.; Zarrindast, M.R.; Sharifi, A.M. Deferoxamine preconditioning of neural-like cells derived from human Wharton’s Jelly mesenchymal stem cells as a strategy to promote their tolerance and therapeutic potential: An in vitro study. Cell. Mol. Neurobiol. 2016, 36, 689–700. [Google Scholar] [CrossRef]
- Chen, Q.; Ames, B.N. Senescence-like growth arrest induced by hydrogen peroxide in human diploid fibroblast F65 cells. Proc. Natl. Acad. Sci. USA 1994, 91, 4130–4134. [Google Scholar] [CrossRef] [Green Version]
- Yoon, Y.S.; Cho, H.; Lee, J.H.; Yoon, G. Mitochondrial dysfunction via disruption of complex II activity during iron chelation-induced senescence-like growth arrest of Chang cells. Ann. N. Y. Acad. Sci. 2004, 1011, 123–132. [Google Scholar] [CrossRef]
- Yoon, G.; Kim, H.J.; Yoon, Y.S.; Cho, H.; Lim, I.K.; Lee, J.H. Iron chelation-induced senescence-like growth arrest in hepatocyte cell lines: Association of transforming growth factor beta1 (TGF-beta1)-mediated p27Kip1 expression. Biochem. J. 2002, 366, 613–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coleman, P.R.; Chang, G.; Hutas, G.; Grimshaw, M.; Vadas, M.A.; Gamble, J.R. Age-associated stresses induce an anti-inflammatory senescent phenotype in endothelial cells. Aging 2013, 5, 913–924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, H.L.; Zhong, Q.; Qin, Y.L.; Bu, Q.Q.; Han, X.A.; Jia, H.T.; Liu, H.W. Hypoxia-mimetic agents inhibit proliferation and alter the morphology of human umbilical cord-derived mesenchymal stem cells. BMC Cell Biol. 2011, 12, 32. [Google Scholar] [CrossRef] [Green Version]
- Yoon, Y.S.; Byun, H.O.; Cho, H.; Kim, B.K.; Yoon, G. Complex II defect via down-regulation of iron-sulfur subunit induces mitochondrial dysfunction and cell cycle delay in iron chelation-induced senescence-associated growth arrest. J. Biol. Chem. 2003, 278, 51577–51586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Passos, J.F.; Nelson, G.; Wang, C.; Richter, T.; Simillion, C.; Proctor, C.J.; Miwa, S.; Olijslagers, S.; Hallinan, J.; Wipat, A.; et al. Feedback between p21 and reactive oxygen production is necessary for cell senescence. Mol. Syst. Biol. 2010, 6, 347. [Google Scholar] [CrossRef]
- Babbs, C.F.; Steiner, M.G. Detection and quantitation of hydroxyl radical using dimethyl sulfoxide as molecular probe. Methods Enzymol. 1990, 186, 137–147. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.; Sun, L.; Chen, X.; Zhang, D. Oxidative stress, mitochondrial damage and neurodegenerative diseases. Neural Regen. Res. 2013, 8, 2003–2014. [Google Scholar] [CrossRef] [PubMed]
- Kaeberlein, M.; Kapahi, P. The hypoxic response and aging. Cell Cycle 2009, 8, 2324. [Google Scholar] [CrossRef] [PubMed]
- Leontieva, O.V.; Natarajan, V.; Demidenko, Z.N.; Burdelya, L.G.; Gudkov, A.V.; Blagosklonny, M.V. Hypoxia suppresses conversion from proliferative arrest to cellular senescence. Proc. Natl. Acad. Sci. USA 2012, 109, 13314–13318. [Google Scholar] [CrossRef] [Green Version]
- Jin, Y.; Kato, T.; Furu, M.; Nasu, A.; Kajita, Y.; Mitsui, H.; Ueda, M.; Aoyama, T.; Nakayama, T.; Nakamura, T.; et al. Mesenchymal stem cells cultured under hypoxia escape from senescence via down-regulation of p16 and extracellular signal regulated kinase. Biochem. Biophys. Res. Commun. 2010, 391, 1471–1476. [Google Scholar] [CrossRef]
- Lee, J.H.; Yoon, Y.M.; Lee, S.H. Hypoxic preconditioning promotes the bioactivities of mesenchymal stem cells via the HIF-1α-GRP78-Akt axis. Int. J. Mol. Sci. 2017, 18, 1320. [Google Scholar] [CrossRef]
- Alique, M.; Sánchez-López, E.; Bodega, G.; Giannarelli, C.; Carracedo, J.; Ramírez, R. Hypoxia-Inducible Factor-1α: The Master Regulator of Endothelial Cell Senescence in Vascular Aging. Cells 2020, 9, 195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, M.; Song, L.P.; Jiang, Y.; Liu, W.; Yu, Y.; Chen, G.Q. Hypoxia-mimetic agents desferrioxamine and cobalt chloride induce leukemic cell apoptosis through different hypoxia-inducible factor-1alpha independent mechanisms. Apoptosis 2006, 11, 67–77. [Google Scholar] [CrossRef] [PubMed]
- Fujisawa, K.; Takami, T.; Okada, S.; Hara, K.; Matsumoto, T.; Yamamoto, N.; Yamasaki, T.; Sakaida, I. Analysis of metabolomic changes in mesenchymal stem cells on treatment with desferrioxamine as a hypoxia mimetic compared with hypoxic conditions. Stem Cells 2018, 36, 1226–1236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Cui, Y.; Shi, M.; Zhang, Q.; Wang, Q.; Chen, X. Deferoxamine promotes MDA-MB-231 cell migration and invasion through increased ROS-dependent HIF-1α accumulation. Cell Physiol. Biochem. 2014, 33, 1036–1046. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shatrova, A.N.; Burova, E.B.; Kharchenko, M.V.; Smirnova, I.S.; Lyublinskaya, O.G.; Nikolsky, N.N.; Borodkina, A.V. Outcomes of Deferoxamine Action on H2O2-Induced Growth Inhibition and Senescence Progression of Human Endometrial Stem Cells. Int. J. Mol. Sci. 2021, 22, 6035. https://doi.org/10.3390/ijms22116035
Shatrova AN, Burova EB, Kharchenko MV, Smirnova IS, Lyublinskaya OG, Nikolsky NN, Borodkina AV. Outcomes of Deferoxamine Action on H2O2-Induced Growth Inhibition and Senescence Progression of Human Endometrial Stem Cells. International Journal of Molecular Sciences. 2021; 22(11):6035. https://doi.org/10.3390/ijms22116035
Chicago/Turabian StyleShatrova, Alla N., Elena B. Burova, Marianna V. Kharchenko, Irina S. Smirnova, Olga G. Lyublinskaya, Nikolay N. Nikolsky, and Aleksandra V. Borodkina. 2021. "Outcomes of Deferoxamine Action on H2O2-Induced Growth Inhibition and Senescence Progression of Human Endometrial Stem Cells" International Journal of Molecular Sciences 22, no. 11: 6035. https://doi.org/10.3390/ijms22116035
APA StyleShatrova, A. N., Burova, E. B., Kharchenko, M. V., Smirnova, I. S., Lyublinskaya, O. G., Nikolsky, N. N., & Borodkina, A. V. (2021). Outcomes of Deferoxamine Action on H2O2-Induced Growth Inhibition and Senescence Progression of Human Endometrial Stem Cells. International Journal of Molecular Sciences, 22(11), 6035. https://doi.org/10.3390/ijms22116035