
Hyperlipidemic Conditions Impact Force-Induced Inflammatory Response of Human Periodontal Ligament Fibroblasts Concomitantly Challenged with P. gingivalis-LPS

 , ,
, ,
Abstract
:1. Introduction
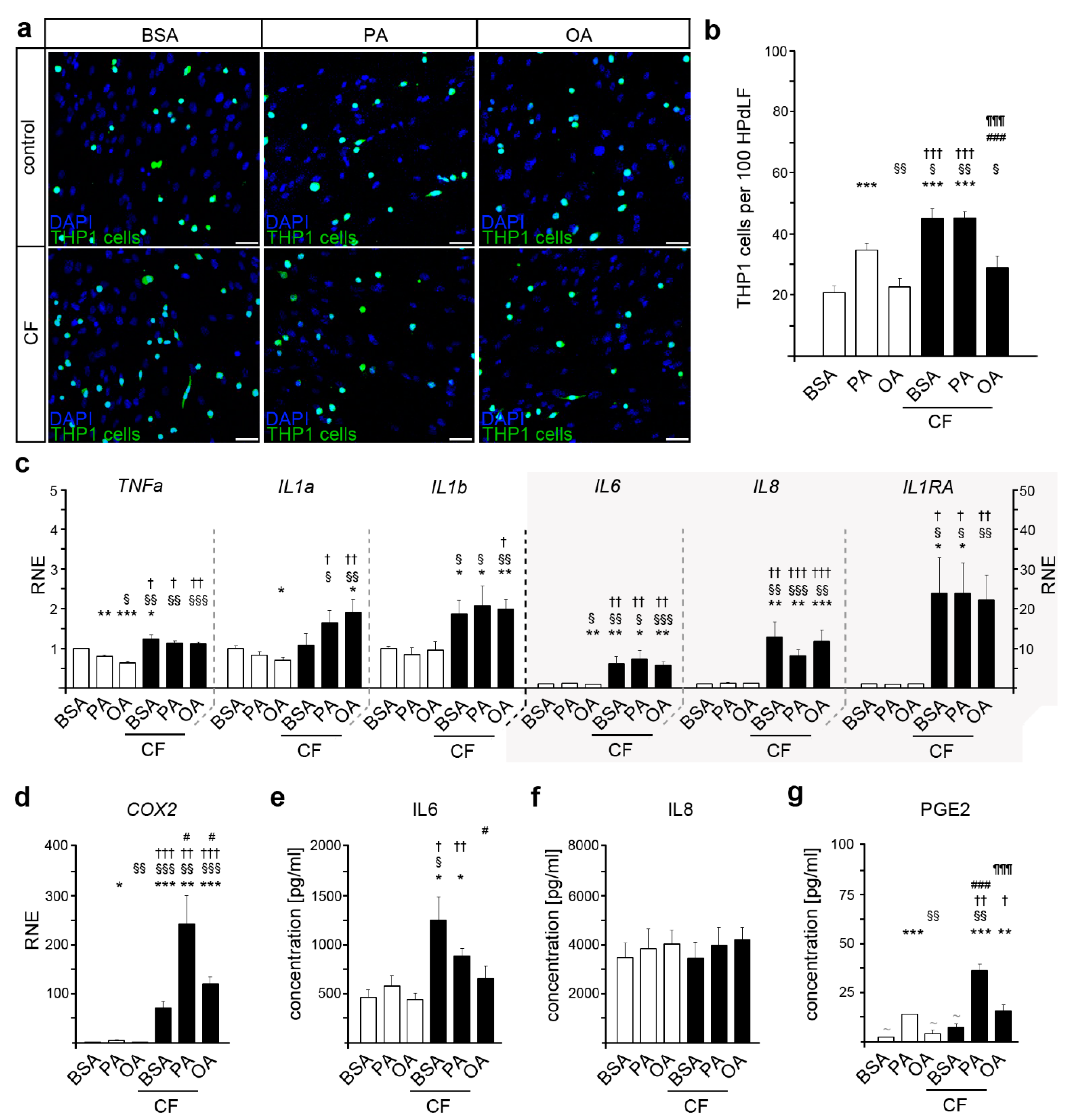
2. Results
2.1. Palmitic Acid Induces an Increased Inflammatory State in HPdLF
2.2. Oleic Acid Impacts THP1 Adhesion due to Mechanical Compression in HPdLF
2.3. COX2 Expression and PGE2 Secretion Are Altered in Mechanically Stimulated HPdLF in Relation to Fatty Acid Stimulation
2.4. Stimulation with Lipopolysaccharides from P. gingivalis Resulted in an Excessive Inflammatory Response of HPdLF Exposed to Palmitic Acid
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Fatty Acid and P. gingivalis LPS Stimulation
4.3. Mechanical Compression
4.4. THP1 Cell Adherence assay
4.5. RNA Extraction and Quantitative PCR
4.6. Enzyme-Linked Immunosorbent Assay (ELISA)
4.7. Microscopy, Image Analysis, and Statistics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Khan, S.; Barrington, G.; Bettiol, S.; Barnett, T.; Crocombe, L. Is overweight/obesity a risk factor for periodontitis in young adults and adolescents? A systematic review. Obes. Rev. Off. J. Int. Assoc. Study Obes. 2018, 19, 852–883. [Google Scholar] [CrossRef] [PubMed]
- Zorena, K.; Jachimowicz-Duda, O.; Slezak, D.; Robakowska, M.; Mrugacz, M. Adipokines and obesity. Potential link to metabolic disorders and chronic complications. Int. J. Mol. Sci. 2020, 21, 3570. [Google Scholar] [CrossRef]
- Ouchi, N.; Parker, J.L.; Lugus, J.J.; Walsh, K. Adipokines in inflammation and metabolic disease. Nat. Rev. Immunol. 2011, 11, 85–97. [Google Scholar] [CrossRef]
- Boden, G. Obesity and free fatty acids. Endocrinol. Metab. Clin. N. Am. 2008, 37, 635–646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suganami, T.; Tanimoto-Koyama, K.; Nishida, J.; Itoh, M.; Yuan, X.; Mizuarai, S.; Kotani, H.; Yamaoka, S.; Miyake, K.; Aoe, S.; et al. Role of the toll-like receptor 4/nf-kappab pathway in saturated fatty acid-induced inflammatory changes in the interaction between adipocytes and macrophages. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 84–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooke, A.A.; Connaughton, R.M.; Lyons, C.L.; McMorrow, A.M.; Roche, H.M. Fatty acids and chronic low grade inflammation associated with obesity and the metabolic syndrome. Eur. J. Pharmacol. 2016, 785, 207–214. [Google Scholar] [CrossRef]
- Ebbert, J.O.; Jensen, M.D. Fat depots, free fatty acids, and dyslipidemia. Nutrients 2013, 5, 498–508. [Google Scholar] [CrossRef] [Green Version]
- Drosatos-Tampakaki, Z.; Drosatos, K.; Siegelin, Y.; Gong, S.; Khan, S.; Van Dyke, T.; Goldberg, I.J.; Schulze, P.C.; Schulze-Spate, U. Palmitic acid and dgat1 deficiency enhance osteoclastogenesis, while oleic acid-induced triglyceride formation prevents it. J. Bone Miner. Res. Off. J. Am. Soc. Bone Miner. Res. 2014, 29, 1183–1195. [Google Scholar] [CrossRef]
- Bierman, E.L.; Dole, V.P.; Roberts, T.N. An abnormality of nonesterified fatty acid metabolism in diabetes mellitus. Diabetes 1957, 6, 475–479. [Google Scholar] [CrossRef] [PubMed]
- Taskinen, M.R.; Bogardus, C.; Kennedy, A.; Howard, B.V. Multiple disturbances of free fatty acid metabolism in noninsulin-dependent diabetes. Effect of oral hypoglycemic therapy. J. Clin. Investig. 1985, 76, 637–644. [Google Scholar] [CrossRef] [Green Version]
- Prisby, R.D.; Swift, J.M.; Bloomfield, S.A.; Hogan, H.A.; Delp, M.D. Altered bone mass, geometry and mechanical properties during the development and progression of type 2 diabetes in the zucker diabetic fatty rat. J. Endocrinol. 2008, 199, 379–388. [Google Scholar] [CrossRef] [Green Version]
- Soares, E.A.; Nakagaki, W.R.; Garcia, J.A.; Camilli, J.A. Effect of hyperlipidemia on femoral biomechanics and morphology in low-density lipoprotein receptor gene knockout mice. J. Bone Miner. Metab. 2012, 30, 419–425. [Google Scholar] [CrossRef] [PubMed]
- Cistola, D.P.; Small, D.M. Fatty acid distribution in systems modeling the normal and diabetic human circulation. A 13c nuclear magnetic resonance study. J. Clin. Investig. 1991, 87, 1431–1441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kissebah, A.H.; Alfarsi, S.; Adams, P.W.; Wynn, V. Role of insulin resistance in adipose tissue and liver in the pathogenesis of endogenous hypertriglyceridaemia in man. Diabetologia 1976, 12, 563–571. [Google Scholar] [CrossRef]
- Ferreri, C.; Masi, A.; Sansone, A.; Giacometti, G.; Larocca, A.V.; Menounou, G.; Scanferlato, R.; Tortorella, S.; Rota, D.; Conti, M.; et al. Fatty acids in membranes as homeostatic, metabolic and nutritional biomarkers: Recent advancements in analytics and diagnostics. Diagnostics 2016, 7, 1. [Google Scholar] [CrossRef]
- Sokolova, M.; Vinge, L.E.; Alfsnes, K.; Olsen, M.B.; Eide, L.; Kaasboll, O.J.; Attramadal, H.; Torp, M.K.; Fosshaug, L.E.; Rashidi, A.; et al. Palmitate promotes inflammatory responses and cellular senescence in cardiac fibroblasts. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2017, 1862, 234–245. [Google Scholar] [CrossRef]
- Pillon, N.J.; Azizi, P.M.; Li, Y.E.; Liu, J.; Wang, C.; Chan, K.L.; Hopperton, K.E.; Bazinet, R.P.; Heit, B.; Bilan, P.J.; et al. Palmitate-induced inflammatory pathways in human adipose microvascular endothelial cells promote monocyte adhesion and impair insulin transcytosis. Am. J. Physiol. Endocrinol. Metab. 2015, 309, E35–E44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nemecz, M.; Constantin, A.; Dumitrescu, M.; Alexandru, N.; Filippi, A.; Tanko, G.; Georgescu, A. The distinct effects of palmitic and oleic acid on pancreatic beta cell function: The elucidation of associated mechanisms and effector molecules. Front. Pharmacol. 2018, 9, 1554. [Google Scholar] [CrossRef] [Green Version]
- Gillet, C.; Spruyt, D.; Rigutto, S.; Dalla Valle, A.; Berlier, J.; Louis, C.; Debier, C.; Gaspard, N.; Malaisse, W.J.; Gangji, V.; et al. Oleate abrogates palmitate-induced lipotoxicity and proinflammatory response in human bone marrow-derived mesenchymal stem cells and osteoblastic cells. Endocrinology 2015, 156, 4081–4093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Souza, C.O.; Valenzuela, C.A.; Baker, E.J.; Miles, E.A.; Rosa Neto, J.C.; Calder, P.C. Palmitoleic acid has stronger anti-inflammatory potential in human endothelial cells compared to oleic and palmitic acids. Mol. Nutr. Food Res. 2018, 62, e1800322. [Google Scholar] [CrossRef]
- Carrillo, C.; Cavia Mdel, M.; Alonso-Torre, S. Role of oleic acid in immune system; mechanism of action; a review. Nutr. Hosp. 2012, 27, 978–990. [Google Scholar] [PubMed] [Green Version]
- de Lima-Salgado, T.M.; Alba-Loureiro, T.C.; do Nascimento, C.S.; Nunes, M.T.; Curi, R. Molecular mechanisms by which saturated fatty acids modulate tnf-alpha expression in mouse macrophage lineage. Cell Biochem. Biophys. 2011, 59, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Al-Shudiefat, A.A.; Sharma, A.K.; Bagchi, A.K.; Dhingra, S.; Singal, P.K. Oleic acid mitigates tnf-alpha-induced oxidative stress in rat cardiomyocytes. Mol. Cell. Biochem. 2013, 372, 75–82. [Google Scholar] [CrossRef]
- Korbecki, J.; Bajdak-Rusinek, K. The effect of palmitic acid on inflammatory response in macrophages: An overview of molecular mechanisms. Inflamm. Res. 2019, 68, 915–932. [Google Scholar] [CrossRef] [Green Version]
- Zhou, H.; Urso, C.J.; Jadeja, V. Saturated fatty acids in obesity-associated inflammation. J. Inflamm. Res. 2020, 13, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Sergi, D.; Morris, A.C.; Kahn, D.E.; McLean, F.H.; Hay, E.A.; Kubitz, P.; MacKenzie, A.; Martinoli, M.G.; Drew, J.E.; Williams, L.M. Palmitic acid triggers inflammatory responses in n42 cultured hypothalamic cells partially via ceramide synthesis but not via tlr4. Nutr. Neurosci. 2020, 23, 321–334. [Google Scholar] [CrossRef] [PubMed]
- Novgorodov, S.A.; Wu, B.X.; Gudz, T.I.; Bielawski, J.; Ovchinnikova, T.V.; Hannun, Y.A.; Obeid, L.M. Novel pathway of ceramide production in mitochondria: Thioesterase and neutral ceramidase produce ceramide from sphingosine and acyl-coa. J. Biol. Chem. 2011, 286, 25352–25362. [Google Scholar] [CrossRef] [Green Version]
- Hannun, Y.A. Functions of ceramide in coordinating cellular responses to stress. Science 1996, 274, 1855–1859. [Google Scholar] [CrossRef] [PubMed]
- Symmank, J.; Chorus, M.; Appel, S.; Marciniak, J.; Knaup, I.; Bastian, A.; Hennig, C.L.; Doding, A.; Schulze-Spate, U.; Jacobs, C.; et al. Distinguish fatty acids impact survival, differentiation and cellular function of periodontal ligament fibroblasts. Sci. Rep. 2020, 10, 15706. [Google Scholar] [CrossRef] [PubMed]
- Ravaut, G.; Legiot, A.; Bergeron, K.F.; Mounier, C. Monounsaturated fatty acids in obesity-related inflammation. Int. J. Mol. Sci. 2020, 22, 330. [Google Scholar] [CrossRef]
- Romana-Souza, B.; Saguie, B.O.; Pereira de Almeida Nogueira, N.; Paes, M.; Dos Santos Valenca, S.; Atella, G.C.; Monte-Alto-Costa, A. Oleic acid and hydroxytyrosol present in olive oil promote ros and inflammatory response in normal cultures of murine dermal fibroblasts through the nf-kappab and nrf2 pathways. Food Res. Int. 2020, 131, 108984. [Google Scholar] [CrossRef] [PubMed]
- Malodobra-Mazur, M.; Cierzniak, A.; Dobosz, T. Oleic acid influences the adipogenesis of 3t3-l1 cells via DNA methylation and may predispose to obesity and obesity-related disorders. Lipids Health Dis. 2019, 18, 230. [Google Scholar] [CrossRef] [Green Version]
- Cecoro, G.; Annunziata, M.; Iuorio, M.T.; Nastri, L.; Guida, L. Periodontitis, low-grade inflammation and systemic health: A scoping review. Medicina 2020, 56, 272. [Google Scholar] [CrossRef] [PubMed]
- Kinane, D.F.; Stathopoulou, P.G.; Papapanou, P.N. Periodontal diseases. Nat. Reviews. Dis. Primers 2017, 3, 17038. [Google Scholar] [CrossRef]
- Deo, P.N.; Deshmukh, R. Oral microbiome: Unveiling the fundamentals. J. Oral Maxillofac. Pathol. 2019, 23, 122–128. [Google Scholar]
- Nakayama, M.; Ohara, N. Molecular mechanisms of porphyromonas gingivalis-host cell interaction on periodontal diseases. Jpn. Dent. Sci. Rev. 2017, 53, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Mulhall, H.; Huck, O.; Amar, S. Porphyromonas gingivalis, a long-range pathogen: Systemic impact and therapeutic implications. Microorganisms 2020, 8, 869. [Google Scholar] [CrossRef] [PubMed]
- Amar, S.; Zhou, Q.; Shaik-Dasthagirisaheb, Y.; Leeman, S. Diet-induced obesity in mice causes changes in immune responses and bone loss manifested by bacterial challenge. Proc. Natl. Acad. Sci. USA 2007, 104, 20466–20471. [Google Scholar] [CrossRef] [Green Version]
- Cani, P.D.; Amar, J.; Iglesias, M.A.; Poggi, M.; Knauf, C.; Bastelica, D.; Neyrinck, A.M.; Fava, F.; Tuohy, K.M.; Chabo, C.; et al. Metabolic endotoxemia initiates obesity and insulin resistance. Diabetes 2007, 56, 1761–1772. [Google Scholar] [CrossRef] [Green Version]
- Le Sage, F.; Meilhac, O.; Gonthier, M.P. Porphyromonas gingivalis lipopolysaccharide induces pro-inflammatory adipokine secretion and oxidative stress by regulating toll-like receptor-mediated signaling pathways and redox enzymes in adipocytes. Mol. Cell. Endocrinol. 2017, 446, 102–110. [Google Scholar] [CrossRef]
- Kato, T.; Yamazaki, K.; Nakajima, M.; Date, Y.; Kikuchi, J.; Hase, K.; Ohno, H.; Yamazaki, K. Oral administration of porphyromonas gingivalis alters the gut microbiome and serum metabolome. mSphere 2018, 3, e00460-18. [Google Scholar] [CrossRef] [Green Version]
- Arimatsu, K.; Yamada, H.; Miyazawa, H.; Minagawa, T.; Nakajima, M.; Ryder, M.I.; Gotoh, K.; Motooka, D.; Nakamura, S.; Iida, T.; et al. Oral pathobiont induces systemic inflammation and metabolic changes associated with alteration of gut microbiota. Sci. Rep. 2014, 4, 4828. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, M.; Fukasawa, S. Is inflammation a friend or foe for orthodontic treatment?: Inflammation in orthodontically induced inflammatory root resorption and accelerating tooth movement. Int. J. Mol. Sci. 2021, 22, 2388. [Google Scholar] [CrossRef]
- Yamaguchi, M. Rank/rankl/opg during orthodontic tooth movement. Orthod. Craniofacial Res. 2009, 12, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Jonsson, D.; Nebel, D.; Bratthall, G.; Nilsson, B.O. The human periodontal ligament cell: A fibroblast-like cell acting as an immune cell. J. Periodontal Res. 2011, 46, 153–157. [Google Scholar] [CrossRef] [PubMed]
- Weltman, B.; Vig, K.W.; Fields, H.W.; Shanker, S.; Kaizar, E.E. Root resorption associated with orthodontic tooth movement: A systematic review. Am. J. Orthod. Dentofac. Orthop. 2010, 137, 462–476, discussion 412A. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wishney, M. Potential risks of orthodontic therapy: A critical review and conceptual framework. Aust. Dent. J. 2017, 62 (Suppl. 1), 86–96. [Google Scholar] [CrossRef] [Green Version]
- Garlet, T.P.; Coelho, U.; Silva, J.S.; Garlet, G.P. Cytokine expression pattern in compression and tension sides of the periodontal ligament during orthodontic tooth movement in humans. Eur. J. Oral Sci. 2007, 115, 355–362. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Jacox, L.A.; Little, S.H.; Ko, C.C. Orthodontic tooth movement: The biology and clinical implications. Kaohsiung J. Med Sci. 2018, 34, 207–214. [Google Scholar] [CrossRef] [Green Version]
- von Bremen, J.; Wagner, J.; Ruf, S. Correlation between body mass index and orthodontic treatment outcome. Angle Orthod. 2013, 83, 371–375. [Google Scholar] [CrossRef] [Green Version]
- Saloom, H.F.; Papageorgiou, S.N.; Carpenter, G.H.; Cobourne, M.T. Impact of obesity on orthodontic tooth movement in adolescents: A prospective clinical cohort study. J. Dent. Res. 2017, 96, 547–554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, B.; Liu, D.; Zhang, C.; Zhang, T.; Wang, X.; Yang, R.; Liu, Y.; He, D.; Zhou, Y. Obesity attenuates force-induced tooth movement in mice with the elevation of leptin level: A preliminary translational study. Am. J. Transl. Res. 2018, 10, 4107–4118. [Google Scholar]
- von Bremen, J.; Lorenz, N.; Ruf, S. Impact of body mass index on oral health during orthodontic treatment: An explorative pilot study. Eur. J. Orthod. 2016, 38, 386–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, J.J. Effects of obesity on bone metabolism. J. Orthop. Surg. Res. 2011, 6, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirschneck, C.; Fanghanel, J.; Wahlmann, U.; Wolf, M.; Roldan, J.C.; Proff, P. Interactive effects of periodontitis and orthodontic tooth movement on dental root resorption, tooth movement velocity and alveolar bone loss in a rat model. Ann. Anat. Anat. Anz. Off. Organ Anat. Ges. 2017, 210, 32–43. [Google Scholar] [CrossRef]
- Okamoto, A.; Ohnishi, T.; Bandow, K.; Kakimoto, K.; Chiba, N.; Maeda, A.; Fukunaga, T.; Miyawaki, S.; Matsuguchi, T. Reduction of orthodontic tooth movement by experimentally induced periodontal inflammation in mice. Eur. J. Oral Sci. 2009, 117, 238–247. [Google Scholar] [CrossRef] [PubMed]
- Bosshart, H.; Heinzelmann, M. Thp-1 cells as a model for human monocytes. Ann. Transl. Med. 2016, 4, 438. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Li, M.; Tan, L.; Huang, S.; Zhao, L.; Tang, T.; Liu, J.; Zhao, Z. Analysis of time-course gene expression profiles of a periodontal ligament tissue model under compression. Arch. Oral Biol. 2013, 58, 511–522. [Google Scholar] [CrossRef]
- Chomarat, P.; Banchereau, J.; Davoust, J.; Palucka, A.K. Il-6 switches the differentiation of monocytes from dendritic cells to macrophages. Nat. Immunol. 2000, 1, 510–514. [Google Scholar] [CrossRef] [PubMed]
- Zaslona, Z.; Serezani, C.H.; Okunishi, K.; Aronoff, D.M.; Peters-Golden, M. Prostaglandin e2 restrains macrophage maturation via e prostanoid receptor 2/protein kinase a signaling. Blood 2012, 119, 2358–2367. [Google Scholar] [CrossRef]
- Ramadan, D.E.; Hariyani, N.; Indrawati, R.; Ridwan, R.D.; Diyatri, I. Cytokines and chemokines in periodontitis. Eur. J. Dent. 2020, 14, 483–495. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Hu, S.; Cui, Y.; Sun, M.K.; Xie, F.; Zhang, Q.; Jin, J. Saturated fatty acids up-regulate cox-2 expression in prostate epithelial cells via toll-like receptor 4/nf-kappab signaling. Inflammation 2014, 37, 467–477. [Google Scholar] [CrossRef]
- Coll, T.; Palomer, X.; Blanco-Vaca, F.; Escola-Gil, J.C.; Sanchez, R.M.; Laguna, J.C.; Vazquez-Carrera, M. Cyclooxygenase 2 inhibition exacerbates palmitate-induced inflammation and insulin resistance in skeletal muscle cells. Endocrinology 2010, 151, 537–548. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.K.; White, E.S.; Wettlaufer, S.H.; Grifka, H.; Hogaboam, C.M.; Thannickal, V.J.; Horowitz, J.C.; Peters-Golden, M. Prostaglandin e(2) induces fibroblast apoptosis by modulating multiple survival pathways. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2009, 23, 4317–4326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martien, S.; Pluquet, O.; Vercamer, C.; Malaquin, N.; Martin, N.; Gosselin, K.; Pourtier, A.; Abbadie, C. Cellular senescence involves an intracrine prostaglandin e2 pathway in human fibroblasts. Biochim. Et Biophys. Acta 2013, 1831, 1217–1227. [Google Scholar] [CrossRef]
- Paik, J.S.; Cho, W.K.; Oh, E.H.; Lee, S.B.; Yang, S.W. Palmitate induced secretion of il-6 and mcp-1 in orbital fibroblasts derived from patients with thyroid-associated ophthalmopathy. Mol. Vis. 2012, 18, 1467–1477. [Google Scholar]
- Seifi, M.; Eslami, B.; Saffar, A.S. The effect of prostaglandin e2 and calcium gluconate on orthodontic tooth movement and root resorption in rats. Eur. J. Orthod. 2003, 25, 199–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saito, M.; Saito, S.; Ngan, P.W.; Shanfeld, J.; Davidovitch, Z. Interleukin 1 beta and prostaglandin e are involved in the response of periodontal cells to mechanical stress in vivo and in vitro. Am. J. Orthod. Dentofac. Orthop. 1991, 99, 226–240. [Google Scholar] [CrossRef]
- Cheng, Z.; Abayasekara, D.R.; Elmes, M.; Kirkup, S.; Wathes, D.C. Effect of oleic acid supplementation on prostaglandin production in maternal endometrial and fetal allantochorion cells isolated from late gestation ewes. Placenta 2015, 36, 1011–1017. [Google Scholar] [CrossRef] [Green Version]
- Hageman, J.R.; McCulloch, K.; Hunt, C.E.; Cobb, M.; Quade, B.; Crussi, F.; Pachman, L. Oleic acid lung injury increases plasma prostaglandin levels. Prostaglandins Leukot. Essent. Fat. Acids 1989, 35, 157–164. [Google Scholar] [CrossRef]
- Yamasaki, K.; Shibata, Y.; Fukuhara, T. The effect of prostaglandins on experimental tooth movement in monkeys (macaca fuscata). J. Dent. Res. 1982, 61, 1444–1446. [Google Scholar] [CrossRef]
- Caglaroglu, M.; Erdem, A. Histopathologic investigation of the effects of prostaglandin e2 administered by different methods on tooth movement and bone metabolism. Korean J. Orthod. 2012, 42, 118–128. [Google Scholar] [CrossRef] [Green Version]
- Chao, C.F.; Shih, C.; Wang, T.M.; Lo, T.H. Effects of prostaglandin e2 on alveolar bone resorption during orthodontic tooth movement. Acta Anat. 1988, 132, 304–309. [Google Scholar] [CrossRef]
- Mano, M.; Arakawa, T.; Mano, H.; Nakagawa, M.; Kaneda, T.; Kaneko, H.; Yamada, T.; Miyata, K.; Kiyomura, H.; Kumegawa, M.; et al. Prostaglandin e2 directly inhibits bone-resorbing activity of isolated mature osteoclasts mainly through the ep4 receptor. Calcif. Tissue Int. 2000, 67, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Take, I.; Kobayashi, Y.; Yamamoto, Y.; Tsuboi, H.; Ochi, T.; Uematsu, S.; Okafuji, N.; Kurihara, S.; Udagawa, N.; Takahashi, N. Prostaglandin e2 strongly inhibits human osteoclast formation. Endocrinology 2005, 146, 5204–5214. [Google Scholar] [CrossRef] [Green Version]
- Vernot, J.P. Senescence-associated pro-inflammatory cytokines and tumor cell plasticity. Front. Mol. Biosci. 2020, 7, 63. [Google Scholar] [CrossRef]
- Galluzzi, L.; Vitale, I.; Aaronson, S.A.; Abrams, J.M.; Adam, D.; Agostinis, P.; Alnemri, E.S.; Altucci, L.; Amelio, I.; Andrews, D.W.; et al. Molecular mechanisms of cell death: Recommendations of the nomenclature committee on cell death 2018. Cell Death Differ. 2018, 25, 486–541. [Google Scholar] [CrossRef] [PubMed]
- Janjic Rankovic, M.; Docheva, D.; Wichelhaus, A.; Baumert, U. Effect of static compressive force on in vitro cultured pdl fibroblasts: Monitoring of viability and gene expression over 6 days. Clin. Oral Investig. 2020, 24, 2497–2511. [Google Scholar] [CrossRef]
- Zhou, B.R.; Zhang, J.A.; Zhang, Q.; Permatasari, F.; Xu, Y.; Wu, D.; Yin, Z.Q.; Luo, D. Palmitic acid induces production of proinflammatory cytokines interleukin-6, interleukin-1beta, and tumor necrosis factor-alpha via a nf-kappab-dependent mechanism in hacat keratinocytes. Mediat. Inflamm. 2013, 2013, 530429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bunn, R.C.; Cockrell, G.E.; Ou, Y.; Thrailkill, K.M.; Lumpkin, C.K., Jr.; Fowlkes, J.L. Palmitate and insulin synergistically induce il-6 expression in human monocytes. Cardiovasc. Diabetol. 2010, 9, 73. [Google Scholar] [CrossRef] [Green Version]
- Volpe, C.M.; Abreu, L.F.; Gomes, P.S.; Gonzaga, R.M.; Veloso, C.A.; Nogueira-Machado, J.A. The production of nitric oxide, il-6, and tnf-alpha in palmitate-stimulated pbmncs is enhanced through hyperglycemia in diabetes. Oxidative Med. Cell. Longev. 2014, 2014, 479587. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, H.G.; Vinolo, M.A.; Magdalon, J.; Vitzel, K.; Nachbar, R.T.; Pessoa, A.F.; dos Santos, M.F.; Hatanaka, E.; Calder, P.C.; Curi, R. Oral administration of oleic or linoleic acid accelerates the inflammatory phase of wound healing. J. Investig. Dermatol. 2012, 132, 208–215. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, T.; Kita, M.; Oseko, F.; Nakamura, T.; Imanishi, J.; Kanamura, N. Cytokine production in human periodontal ligament cells stimulated with porphyromonas gingivalis. J. Periodontal Res. 2006, 41, 554–559. [Google Scholar] [CrossRef]
- Liu, J.; Tang, X.; Li, C.; Pan, C.; Li, Q.; Geng, F.; Pan, Y. Porphyromonas gingivalis promotes the cell cycle and inflammatory cytokine production in periodontal ligament fibroblasts. Arch. Oral Biol. 2015, 60, 1153–1161. [Google Scholar] [CrossRef] [PubMed]
- Fadel, H.T.; Pliaki, A.; Gronowitz, E.; Marild, S.; Ramberg, P.; Dahlen, G.; Yucel-Lindberg, T.; Heijl, L.; Birkhed, D. Clinical and biological indicators of dental caries and periodontal disease in adolescents with or without obesity. Clin. Oral Investig. 2014, 18, 359–368. [Google Scholar] [CrossRef]
- Shikama, Y.; Kudo, Y.; Ishimaru, N.; Funaki, M. Possible involvement of palmitate in pathogenesis of periodontitis. J. Cell. Physiol. 2015, 230, 2981–2989. [Google Scholar] [CrossRef] [Green Version]
- Muluke, M.; Gold, T.; Kiefhaber, K.; Al-Sahli, A.; Celenti, R.; Jiang, H.; Cremers, S.; Van Dyke, T.; Schulze-Spate, U. Diet-induced obesity and its differential impact on periodontal bone loss. J. Dent. Res. 2016, 95, 223–229. [Google Scholar] [CrossRef] [Green Version]
- Kirschneck, C.; Batschkus, S.; Proff, P.; Kostler, J.; Spanier, G.; Schroder, A. Valid gene expression normalization by rt-qpcr in studies on hpdl fibroblasts with focus on orthodontic tooth movement and periodontitis. Sci. Rep. 2017, 7, 14751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative pcr and the 2(-delta delta c(t)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Gene | Gene Symbol | NCBI Gene ID | Primer Sequence | Length |
|---|---|---|---|---|
| C-X-C motif chemokine ligand 8 | IL8 | 3576 | fw TTGGCAGCCTTCCTGATTTCTrew GGTCCACTCTCAATCACTCTCA | 149 bp |
| Interleukin 1 alpha | IL1α | 3552 | fw GACTGCCCAAGATGAAGACCArev CCAAGCACACCCAGTAGTCT | 185 bp |
| Interleukin 1 beta | IL1β | 3553 | fw CGAATCTCCGACCACCACTArev AGCCTCGTTATCCCATGTGT | 186 bp |
| Interleukin-1 receptor antagonist | IL1RN(IL1RA) | 3557 | fw GATGTGCCTGTCCTGTGTCArev ACTCAAAACTGGTGGTGGGG | 146 bp |
| Interleukin 6 | IL6 | 3569 | fw CATCCTCGACGGCATCTCAGrew TCACCAGGCAAGTCTCCTCA | 164 bp |
| Prostaglandin-endoperoxide synthase 2 | PTGS2(COX2) | 5743 | fw GATGATTGCCCGACTCCCTTrew GGCCCTCGCTTATGATCTGT | 185 bp |
| Ribosomal protein L22 | RPL22 | 6146 | fw TGATTGCACCCACCCTGTAGrev GGTTCCCAGCTTTTCCGTTC | 98 bp |
| TATA-box binding protein | TBP | 6908 | fw CGGCTGTTTAACTTCGCTTCCrev TGGGTTATCTTCACACGCCAAG | 86 bp |
| Tumor necrosis factor | TNFα | 7124 | fw CACGCTCTTCTGCCTGCTGrev AGGCTTGTCACTCGGGGTT | 130 bp |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Symmank, J.; Appel, S.; Bastian, J.A.; Knaup, I.; Marciniak, J.; Hennig, C.-L.; Döding, A.; Schulze-Späte, U.; Jacobs, C.; Wolf, M. Hyperlipidemic Conditions Impact Force-Induced Inflammatory Response of Human Periodontal Ligament Fibroblasts Concomitantly Challenged with P. gingivalis-LPS. Int. J. Mol. Sci. 2021, 22, 6069. https://doi.org/10.3390/ijms22116069
Symmank J, Appel S, Bastian JA, Knaup I, Marciniak J, Hennig C-L, Döding A, Schulze-Späte U, Jacobs C, Wolf M. Hyperlipidemic Conditions Impact Force-Induced Inflammatory Response of Human Periodontal Ligament Fibroblasts Concomitantly Challenged with P. gingivalis-LPS. International Journal of Molecular Sciences. 2021; 22(11):6069. https://doi.org/10.3390/ijms22116069
Chicago/Turabian StyleSymmank, Judit, Sophie Appel, Jana Asisa Bastian, Isabel Knaup, Jana Marciniak, Christoph-Ludwig Hennig, Annika Döding, Ulrike Schulze-Späte, Collin Jacobs, and Michael Wolf. 2021. "Hyperlipidemic Conditions Impact Force-Induced Inflammatory Response of Human Periodontal Ligament Fibroblasts Concomitantly Challenged with P. gingivalis-LPS" International Journal of Molecular Sciences 22, no. 11: 6069. https://doi.org/10.3390/ijms22116069
APA StyleSymmank, J., Appel, S., Bastian, J. A., Knaup, I., Marciniak, J., Hennig, C. -L., Döding, A., Schulze-Späte, U., Jacobs, C., & Wolf, M. (2021). Hyperlipidemic Conditions Impact Force-Induced Inflammatory Response of Human Periodontal Ligament Fibroblasts Concomitantly Challenged with P. gingivalis-LPS. International Journal of Molecular Sciences, 22(11), 6069. https://doi.org/10.3390/ijms22116069