
Plasma Gelsolin Reinforces the Diagnostic Value of FGF-21 and GDF-15 for Mitochondrial Disorders

 , , , ,
, , , , 
 , , and
, , and 
Abstract
:1. Introduction
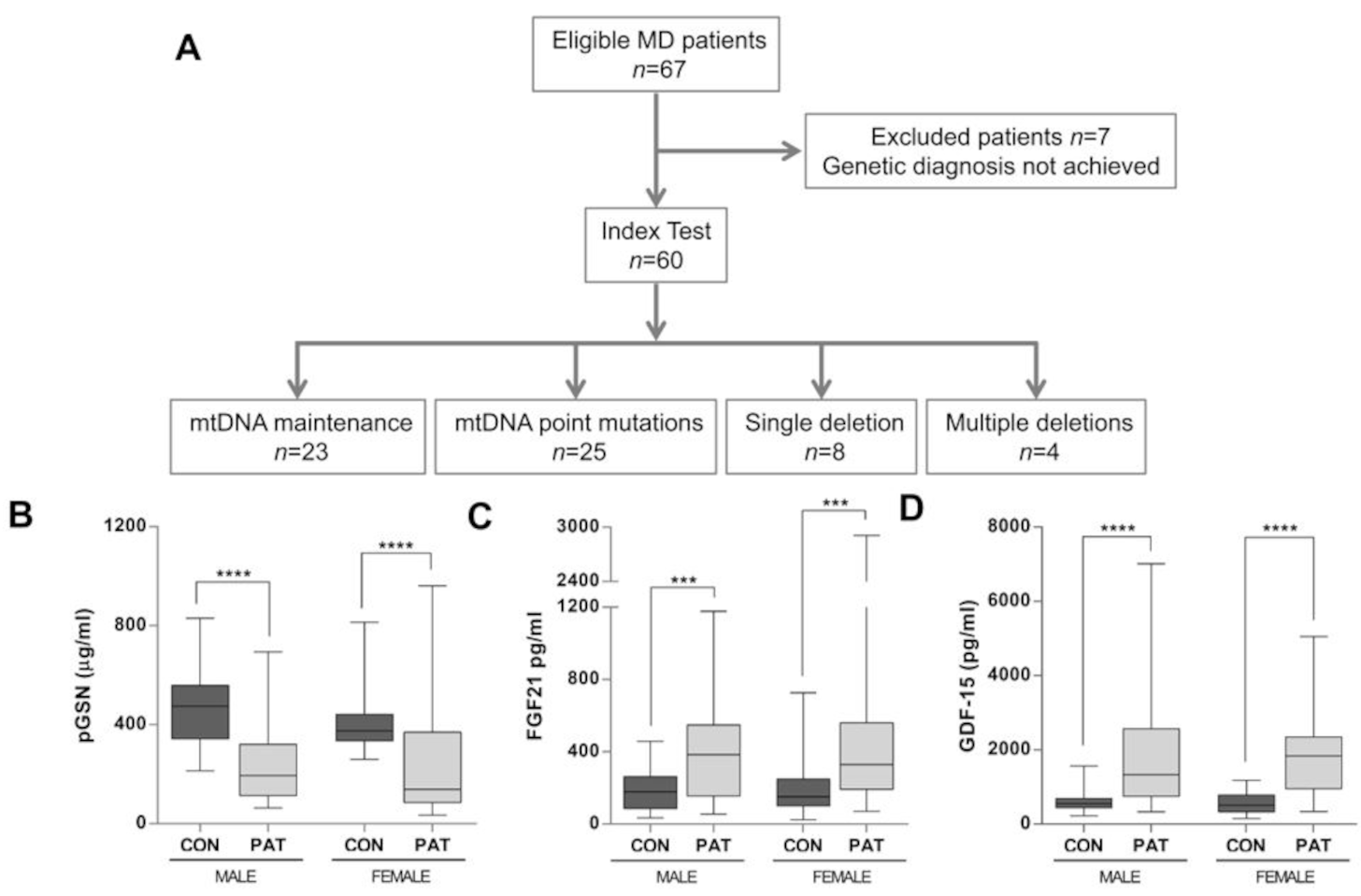
2. Results
2.1. pGSN Levels Are Significantly Decreased in MD Patients
2.2. pGSN Significantly Discriminates Young MD Patients from Healthy Controls and Non-MD Patients
2.3. pGSN Improves Detection of MD Patients Regardless of Their Clinical Phenotypes
2.4. Diagnostic Performance of pGSN, FGF-21, and GDF-15 Plasma Levels for Mitochondrial Disorders
3. Discussion
4. Materials and Methods
4.1. Participants
4.2. Southern Blot Analysis of Single and Multiple mtDNA Deletions
4.3. ELISA Assays
4.4. Statistical Data Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reid, R.A.; Moyle, J.; Mitchell, P. Synthesis of Adenosine Triphosphate by a Protonmotive Force in Rat Liver Mitochondria. Nature 1966, 212, 257–258. [Google Scholar] [CrossRef]
- Lightowlers, R.N.; Taylor, R.W.; Turnbull, D.M. Mutations causing mitochondrial disease: What is new and what challenges remain? Science 2015, 349, 1494–1499. [Google Scholar] [CrossRef]
- McFarland, R.; Taylor, R.W.; Turnbull, D.M. A neurological perspective on mitochondrial disease. Lancet Neurol. 2010, 9, 829–840. [Google Scholar] [CrossRef]
- Gorman, G.S.; Chinnery, P.F.; DiMauro, S.; Hirano, M.; Koga, Y.; McFarland, R.; Suomalainen, A.; Thorburn, D.R.; Zeviani, M.; Turnbull, D.M. Mitochondrial diseases. Nat. Rev. Dis. Primers 2016, 2, 16080. [Google Scholar] [CrossRef]
- Davis, R.L.; Liang, C.; Sue, C.M. A comparison of current serum biomarkers as diagnostic indicators of mitochondrial diseases. Neurology 2016, 86, 2010–2015. [Google Scholar] [CrossRef]
- Maresca, A.; the ER-MITO Study Group; Del Dotto, V.; Romagnoli, M.; La Morgia, C.; Di Vito, L.; Capristo, M.; Valentino, M.L.; Carelli, V. Expanding and validating the biomarkers for mitochondrial diseases. J. Mol. Med. 2020, 98, 1467–1478. [Google Scholar] [CrossRef]
- Varhaug, K.N.; Hikmat, O.; Nakkestad, H.L.; Vedeler, C.A.; Bindoff, L.A. Serum biomarkers in primary mitochondrial disorders. Brain Commun. 2021, 3, fcaa222. [Google Scholar] [CrossRef]
- BonDurant, L.D.; Ameka, M.; Naber, M.C.; Markan, K.R.; Idiga, S.O.; Acevedo, M.R.; Walsh, S.A.; Ornitz, D.M.; Potthoff, M.J. FGF21 Regulates Metabolism Through Adipose-Dependent and—Independent Mechanisms. Cell Metab. 2017, 25, 935–944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Søberg, S.; Sandholt, C.H.; Jespersen, N.Z.; Toft, U.; Madsen, A.L.; von Holstein-Rathlou, S.; Grevengoed, T.J.; Christensen, K.B.; Bredie, W.L.; Potthoff, M.J.; et al. FGF21 Is a Sugar-Induced Hormone Associated with Sweet Intake and Preference in Humans. Cell Metab. 2017, 25, 1045–1053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, R.L.; Liang, C.; Edema-Hildebrand, F.; Riley, C.; Needham, M.; Sue, C.M. Fibroblast growth factor 21 is a sensitive biomarker of mitochondrial disease. Neurology 2013, 81, 1819–1826. [Google Scholar] [CrossRef] [PubMed]
- Suomalainen, A. Biomarkers for mitochondrial respiratory chain disorders. J. Inherit. Metab. Dis. 2011, 34, 277–282. [Google Scholar] [CrossRef]
- Lehtonen, J.M.; Forsström, S.; Bottani, E.; Viscomi, C.; Baris, O.R.; Isoniemi, H.; Höckerstedt, K.; Österlund, P.; Hurme, M.; Jylhävä, J.; et al. FGF21 is a biomarker for mitochondrial translation and mtDNA maintenance disorders. Neurology 2016, 87, 2290–2299. [Google Scholar] [CrossRef]
- Forsström, S.; Jackson, C.B.; Carroll, C.J.; Kuronen, M.; Pirinen, E.; Pradhan, S.; Marmyleva, A.; Auranen, M.; Kleine, I.-M.; Khan, N.A.; et al. Fibroblast Growth Factor 21 Drives Dynamics of Local and Systemic Stress Responses in Mitochondrial Myopathy with mtDNA Deletions. Cell Metab. 2019, 30, 1040–1054. [Google Scholar] [CrossRef]
- Su, S.-L.; Wang, W.-F.; Wu, S.-L.; Wu, H.-M.; Chang, J.-C.; Huang, C.-S.; Cheng, W.-L.; Soong, B.-W.; Lee, Y.-C.; Li, J.-Y.; et al. FGF21 in ataxia patients with spinocerebellar atrophy and mitochondrial disease. Clin. Chim. Acta 2012, 414, 225–227. [Google Scholar] [CrossRef] [PubMed]
- Koene, S.; de Laat, P.; Van Tienoven, D.H.; Vriens, D.; Brandt, A.M.; Sweep, F.; Rodenburg, R.J.; Donders, A.R.T.; Janssen, M.C.; Smeitink, J.A.M. Serum FGF21 levels in adult m.3243A>G carriers: Clinical implications. Neurology 2014, 83, 125–133. [Google Scholar] [CrossRef]
- Morovat, A.; Weerasinghe, G.; Nesbitt, V.; Hofer, M.; Agnew, T.; Quaghebeur, G.; Sergeant, K.; Fratter, C.; Guha, N.; Mirzazadeh, M.; et al. Use of FGF-21 as a Biomarker of Mitochondrial Disease in Clinical Practice. J. Clin. Med. 2017, 6, 80. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.; Xiao, Y.-C.; Zhu, H.; Xu, Q.-Y.; Qi, L.; Wang, Y.-B.; Li, X.-J.; Zheng, M.-L.; Zhong, R.-S.; Zhang, Y.; et al. Serum Fibroblast Growth Factor 21 Levels Are Correlated with the Severity of Diabetic Retinopathy. J. Diabetes Res. 2014, 2014, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Kosola, S.; Lampela, H.; Gylling, H.; Jalanko, H.; Nissinen, M.J.; Lauronen, J.; Mäkisalo, H.; Vaaralahti, K.; Miettinen, T.A.; Raivio, T.; et al. Cholesterol Metabolism Altered and FGF21 Levels High After Pediatric Liver Transplantation Despite Normal Serum Lipids. Am. J. Transplant. 2012, 12, 2815–2824. [Google Scholar] [CrossRef] [PubMed]
- Hindricks, J.; Ebert, T.; Bachmann, A.; Kralisch, S.; Lössner, U.; Kratzsch, J.; Stolzenburg, J.-U.; Dietel, A.; Beige, J.; Anders, M.; et al. Serum levels of fibroblast growth factor-21 are increased in chronic and acute renal dysfunction. Clin. Endocrinol. 2014, 80, 918–924. [Google Scholar] [CrossRef]
- Emmerson, P.J.; Duffin, K.L.; Chintharlapalli, S.; Wu, X. GDF15 and Growth Control. Front. Physiol. 2018, 9, 1712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalko, S.G.; Paco, S.; Jou, C.; Rodríguez, M.A.; Meznaric, M.; Rogac, M.; Jekovec-Vrhovsek, M.; Sciacco, M.; Moggio, M.; Fagiolari, G.; et al. Transcriptomic profiling of TK2 deficient human skeletal muscle suggests a role for the p53 signalling pathway and identifies growth and differentiation factor-15 as a potential novel biomarker for mitochondrial myopathies. BMC Genom. 2014, 15, 91. [Google Scholar] [CrossRef] [Green Version]
- Koene, S.; De Laat, P.; Van Tienoven, D.H.; Weijers, G.; Vriens, D.; Sweep, F.C.G.J.; Timmermans, J.; Kapusta, L.; Janssen, M.C.H.; Smeitink, J.A.M.; et al. Serum GDF15 Levels Correlate to Mitochondrial Disease Severity and Myocardial Strain, but Not to Disease Progression in Adult m.3243A>G Carriers. In JIMD Reports; Springer: Berlin/Heidelberg, Germany, 2015; Volume 24, pp. 69–81. [Google Scholar]
- Yatsuga, S.; Fujita, Y.; Ishii, A.; Fukumoto, Y.; Arahata, H.; Kakuma, T.; Kojima, T.; Ito, M.; Tanaka, M.; Saiki, R.; et al. Growth differentiation factor 15 as a useful biomarker for mitochondrial disorders. Ann. Neurol. 2015, 78, 814–823. [Google Scholar] [CrossRef] [Green Version]
- Lehtonen, J.M.; Auranen, M.; Darin, N.; Sofou, K.; Bindoff, L.; Hikmat, O.; Uusimaa, J.; Vieira, P.; Tulinius, M.; Lönnqvist, T.; et al. Diagnostic value of serum biomarkers FGF21 and GDF15 compared to muscle sample in mitochondrial disease. J. Inherit. Metab. Dis. 2021, 44, 469–480. [Google Scholar] [CrossRef]
- Fujita, Y.; Ito, M.; Kojima, T.; Yatsuga, S.; Koga, Y.; Tanaka, M. GDF15 is a novel biomarker to evaluate efficacy of pyruvate therapy for mitochondrial diseases. Mitochondrion 2015, 20, 34–42. [Google Scholar] [CrossRef]
- Montero, R.; Yubero, D.; Villarroya, J.; Henares, D.; Jou, C.; Rodríguez, M.A.; Ramos, F.; Nascimento, A.; Ortez, C.I.; Campistol, J.; et al. GDF-15 Is Elevated in Children with Mitochondrial Diseases and Is Induced by Mitochondrial Dysfunction. PLoS ONE 2016, 11, e0148709. [Google Scholar] [CrossRef] [Green Version]
- Poulsen, N.S.; Madsen, K.L.; Hornsyld, T.M.; Eisum, A.-S.V.; Fornander, F.; Buch, A.E.; Stemmerik, M.G.; Ruiz-Ruiz, C.; Krag, T.O.; Vissing, J. Growth and differentiation factor 15 as a biomarker for mitochondrial myopathy. Mitochondrion 2020, 50, 35–41. [Google Scholar] [CrossRef]
- Adela, R.; Banerjee, S.K. GDF-15 as a Target and Biomarker for Diabetes and Cardiovascular Diseases: A Translational Prospective. J. Diabetes Res. 2015, 2015, 490842. [Google Scholar] [CrossRef]
- Nohara, S.; Ishii, A.; Yamamoto, F.; Yanagiha, K.; Moriyama, T.; Tozaka, N.; Miyake, Z.; Yatsuga, S.; Koga, Y.; Hosaka, T.; et al. GDF-15, a mitochondrial disease biomarker, is associated with the severity of multiple sclerosis. J. Neurol. Sci. 2019, 405, 116429. [Google Scholar] [CrossRef]
- Tsygankova, P.G.; Itkis, Y.S.; Krylova, T.D.; Kurkina, M.; Bychkov, I.O.; Ilyushkina, A.A.; Zabnenkova, V.; Mikhaylova, S.V.; Pechatnikova, N.L.; Sheremet, N.L.; et al. Plasma FGF-21 and GDF-15 are elevated in different inherited metabolic diseases and are not diagnostic for mitochondrial disorders. J. Inherit. Metab. Dis. 2019, 42, 918–933. [Google Scholar] [CrossRef]
- Garrido-Pérez, N.; Vela-Sebastián, A.; López-Gallardo, E.; Emperador, S.; Iglesias, E.; Meade, P.; Jiménez-Mallebrera, C.; Montoya, J.; Bayona-Bafaluy, M.P.; Ruiz-Pesini, E. Oxidative Phosphorylation Dysfunction Modifies the Cell Secretome. Int. J. Mol. Sci. 2020, 21, 3374. [Google Scholar] [CrossRef]
- Yin, H.L.; Stossel, T.P. Control of cytoplasmic actin gel–sol transformation by gelsolin, a calcium-dependent regulatory protein. Nature 1979, 281, 583–586. [Google Scholar] [CrossRef] [PubMed]
- Marín-Buera, L.; García-Bartolomé, A.; Morán, M.; López-Bernardo, E.; Cadenas, S.; Hidalgo, B.; Sánchez, R.; Seneca, S.; Arenas, J.; Martín, M.A.; et al. Differential proteomic profiling unveils new molecular mechanisms associated with mitochondrial complex III deficiency. J. Proteom. 2015, 113, 38–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwiatkowski, D.J.; Stossel, T.P.; Orkin, S.H.; Mole, J.E.; Coltens, H.R.; Yin, H.L. Plasma and cytoplasmic gelsolins are encoded by a single gene and contain a duplicated actin-binding domain. Nature 1986, 323, 455–458. [Google Scholar] [CrossRef] [PubMed]
- Kwiatkowski, D.J.; Mehl, R.; Yin, H.L. Genomic organization and biosynthesis of secreted and cytoplasmic forms of gelsolin. J. Cell Biol. 1988, 106, 375–384. [Google Scholar] [CrossRef] [Green Version]
- Yin, H.L.; Kwiatkowski, D.J.; Mole, J.E.; Cole, F.S. Structure and biosynthesis of cytoplasmic and secreted variants of gelsolin. J. Biol. Chem. 1984, 259, 5271–5276. [Google Scholar] [CrossRef]
- Kwiatkowski, D.J.; Mehl, R.; Izumo, S.; Nadal-Ginard, B.; Yin, H.L. Muscle is the major source of plasma gelsolin. J. Biol. Chem. 1988, 263, 8239–8243. [Google Scholar] [CrossRef]
- Kusano, H.; Shimizu, S.; Koya, R.C.; Fujita, H.; Kamada, S.; Kuzumaki, N.; Tsujimoto, Y. Human gelsolin prevents apoptosis by inhibiting apoptotic mitochondrial changes via closing VDAC. Oncogene 2000, 19, 4807–4814. [Google Scholar] [CrossRef] [Green Version]
- Antequera, D.; Vargas, T.; Ugalde, C.; Spuch, C.; Molina, J.A.; Ferrer, I.; Bermejo-Pareja, F.; Carro, E. Cytoplasmic gelsolin increases mitochondrial activity and reduces Aβ burden in a mouse model of Alzheimer’s disease. Neurobiol. Dis. 2009, 36, 42–50. [Google Scholar] [CrossRef]
- García-Bartolomé, A.; Peñas, A.; Marín-Buera, L.; Jarne, T.L.; Pérez-Pérez, R.; Morán, M.; Arenas, J.; Martín, M.A.; Ugalde, C. Respiratory chain enzyme deficiency induces mitochondrial location of actin-binding gelsolin to modulate the oligomerization of VDAC complexes and cell survival. Hum. Mol. Genet. 2017, 26, 2493–2506. [Google Scholar] [CrossRef]
- García-Bartolomé, A.; Peñas, A.; Illescas, M.; Bermejo, V.; López-Calcerrada, S.; Pérez-Pérez, R.; Marín-Buera, L.; Domínguez-González, C.; Arenas, J.; Martín, M.A.; et al. Altered Expression Ratio of Actin-Binding Gelsolin Isoforms Is a Novel Hallmark of Mitochondrial OXPHOS Dysfunction. Cells 2020, 9, 1922. [Google Scholar] [CrossRef]
- Feldt, J.; Schicht, M.; Garreis, F.; Welss, J.; Schneider, U.W.; Paulsen, F. Structure, regulation and related diseases of the actin-binding protein gelsolin. Expert Rev. Mol. Med. 2018, 20, e7. [Google Scholar] [CrossRef]
- Peddada, N.; Sagar, A.; Ashish; Garg, R. Plasma gelsolin: A general prognostic marker of health. Med. Hypotheses 2012, 78, 203–210. [Google Scholar] [CrossRef]
- Suomalainen, A.; Elo, J.M.; Pietiläinen, K.; Hakonen, A.H.; Sevastianova, K.; Korpela, M.; Isohanni, P.; Marjavaara, S.K.; Tyni, T.; Kiuru-Enari, S.; et al. FGF-21 as a biomarker for muscle-manifesting mitochondrial respiratory chain deficiencies: A diagnostic study. Lancet Neurol. 2011, 10, 806–818. [Google Scholar] [CrossRef]
- Formichi, P.; Cardone, N.; Taglia, I.; Cardaioli, E.; Salvatore, S.; Gerfo, A.L.; Simoncini, C.; Montano, V.; Siciliano, G.; Mancuso, M.; et al. Fibroblast growth factor 21 and grow differentiation factor 15 are sensitive biomarkers of mitochondrial diseases due to mitochondrial transfer-RNA mutations and mitochondrial DNA deletions. Neurol. Sci. 2020, 41, 3653–3662. [Google Scholar] [CrossRef]
- Osborn, T.M.; Verdrengh, M.; Stossel, T.P.; Tarkowski, A.; Bokarewa, M. Decreased levels of the gelsolin plasma isoform in patients with rheumatoid arthritis. Arthritis Res. Ther. 2008, 10, R117. [Google Scholar] [CrossRef] [Green Version]
- Güntert, A.; Campbell, J.; Saleem, M.; O’Brien, D.P.; Thompson, A.J.; Byers, H.L.; Ward, M.A.; Lovestone, S. Plasma Gelsolin is Decreased and Correlates with Rate of Decline in Alzheimer’s Disease. J. Alzheimers Dis. 2010, 21, 585–596. [Google Scholar] [CrossRef] [Green Version]
- Suhler, E.; Lin, W.; Yin, H.L.; Lee, W.M. Decreased plasma gelsolin concentrations in acute liver failure, myocardial infarction, septic shock, and myonecrosis. Crit. Care Med. 1997, 25, 594–598. [Google Scholar] [CrossRef]
- Huang, L.-F.; Yao, Y.-M.; Li, J.-F.; Dong, N.; Liu, C.; Yu, Y.; He, L.-X.; Sheng, Z.-Y. Reduction of Plasma Gelsolin Levels Correlates with Development of Multiple Organ Dysfunction Syndrome and Fatal Outcome in Burn Patients. PLoS ONE 2011, 6, e25748. [Google Scholar] [CrossRef] [Green Version]
- Lee, P.-S.; Drager, L.R.; Stossel, T.P.; Moore, F.D.; Rogers, S.O. Relationship of Plasma Gelsolin Levels to Outcomes in Critically Ill Surgical Patients. Ann. Surg. 2006, 243, 399–403. [Google Scholar] [CrossRef]
- Mounzer, K.C.; Moncure, M.; Smith, Y.R.; DiNubile, M.J. Relationship of Admission Plasma Gelsolin Levels to Clinical Outcomes in Patients after Major Trauma. Am. J. Respir. Crit. Care Med. 1999, 160, 1673–1681. [Google Scholar] [CrossRef]
- Wang, H.; Cheng, B.; Chen, Q.; Wu, S.; Lv, C.; Xie, G.; Jin, Y.; Fang, X. Time course of plasma gelsolin concentrations during severe sepsis in critically ill surgical patients. Crit. Care 2008, 12, R106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Z.; Lee, A.; Nouwens, A.; Henderson, R.D.; McCombe, P.A. Mass spectrometry analysis of plasma from amyotrophic lateral sclerosis and control subjects. Amyotroph. Lateral Scler. Front. Degener. 2018, 19, 362–376. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.P.; Chen, R.L.; Preston, J.E. The influence of ageing in the cerebrospinal fluid concentrations of proteins that are derived from the choroid plexus, brain, and plasma. Exp. Gerontol. 2012, 47, 323–328. [Google Scholar] [CrossRef] [PubMed]
- Pierredon, S.; Ribaux, P.; Tille, J.-C.; Petignat, P.; Cohen, M. Comparative secretome of ovarian serous carcinoma: Gelsolin in the spotlight. Oncol. Lett. 2017, 13, 4965–4973. [Google Scholar] [CrossRef] [Green Version]
- Morava, E.; van den Heuvel, L.; Hol, F.; De Vries, M.C.; Hogeveen, M.; Rodenburg, R.J.; Smeitink, J.A.M. Mitochondrial disease criteria: Diagnostic applications in children. Neurology 2006, 67, 1823–1826. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patient | MD (n = 60) | Non-MD (n = 41) | Control (n = 56) | p-Value | |
|---|---|---|---|---|---|
| Age, mean (SD) | 46.8 (15.8) | 48.0 (18.1) | 50.1 (19.5) | 0.61 | |
| Gender | F | 38 (63%) | 21 (51%) | 31 (55%) | 0.45 |
| M | 22 (37%) | 20 (49%) | 25 (45%) | ||
| pGSN, median [p25, p75] | 143.0 [102.1; 335.7] | 240.0 [120.4; 375.5] | 399.0 [338.0; 515.3] | <0.001 | |
| FGF21, median [p25, p75] | 353.1 [198.9; 588.0] | 100.5 [34.2; 219.0] | 154.4 [98.1; 254.9] | <0.001 | |
| GDF15, median [p25, p75] | 1777.0 [800.9; 2292.0] | 550.6 [361.9; 1061.6] | 546.4 [376.7; 761.1] | <0.001 |
| Study Cohorts | Area Under the ROC Curve Values | 1 Global p-Value | 2p-Value (3 Adjusted p-Value after Bonferroni’s Correction) | |||||
|---|---|---|---|---|---|---|---|---|
| pGSN | FGF21 | GDF15 | pGSN vs. FGF-21 | pGSN vs. GDF-15 | FGF-21 vs. GDF-15 | |||
| All subjects | MD vs. CON | 0.83 (0.75;0.92) | 0.77 (0.68; 0.86) | 0.87 (0.82; 0.94) | 0.1320 | 0.2969 (0.5939) | 0.3919 (0.7838) | 0.0443 (0.0886) |
| MD vs. Non-MD | 0.58 (0.47; 0.70) | 0.77 (0.67; 0.87) | 0.76 (0.66; 0.87) | 0.0457 | 0.0227 (0.0455) | 0.0282 (0.0565) | 0.9308 (0.9999) | |
| Non-MD vs. CON | 0.75 (0.64; 0.86) | 0.61 (0.49; 0.74) | 0.57 (0.45; 0.69) | 0.0553 | 0.0981 (0.1961) | 0.0306 (0.0612) | 0.6577 (0.999) | |
| Subjects > 50 yrs old | MD vs. CON | 0.84 (0.72; 0.96) | 0.74 (0.60; 0.87) | 0.87 (0.76; 0.98) | 0.1921 | 0.2038 (0.4076) | 0.6642 (0.9999) | 0.0871 (0.1742) |
| MD vs. Non-MD | 0.49 (0.31; 0.67) | 0.76 (0.61; 0.91) | 0.72 (0.56; 0.88) | 0.0763 | 0.0245 (0.0489) | 0.0760 (0.1521) | 0.6968 (0.9999) | |
| Non-MD vs. CON | 0.79 (0.63; 0.94) | 0.61 (0.43; 0.79) | 0.71 (0.56; 0.87) | 0.3327 | 0.1385 (0.2770) | 0.5132 (0.9999) | 0.4062 (0.8123) | |
| Subjects ≤ 50 yrs old | MD vs. CON | 0.83 (0.70; 0.94) | 0.82 (0.73; 0.93) | 0.93 (0.88; 0.99) | 0.0979 | 0.8768 (0.9999) | 0.0943 (0.1887) | 0.0953 (0.9999) |
| MD vs. Non-MD | 0.67 (0.51; 0.80) | 0.78 (0.63; 0.92) | 0.85 (0.72; 0.98) | 0.2012 | 0.2764 (0.5529) | 0.0738 (0.1476) | 0.4764 (0.9528) | |
| Non-MD vs. CON | 0.69 (0.53; 0.86) | 0.61 (0.43; 0.79) | 0.56 (0.39; 0.74) | 0.4969 | 0.4289 (0.8579) | 0.2805 (0.5610) | 0.7348 (0.9999) | |
| Study CohortsS | Cut-Off Values | Sensitivity% (95% CI) | Specificity% (95% CI) | PPV% (95% CI) | NPV% (95% CI) | PLR (95% CI) | NLR (95% CI) | Efficiency % |
|---|---|---|---|---|---|---|---|---|
| All subjects | MD vs. Control | |||||||
| pGSN ≤ 252 | 66.10 (54.02; 78.18) | 98.21 (94.75; 100) | 97.50 (92.66; 100) | 73.33 (63.33; 83.34) | 36.92 (5.24; 259.79) | 0.34 (0.24; 0.49) | 81.74 | |
| FGF-21 ≥ 300 | 61.67 (49.36; 73.97) | 83.93 (74.31; 93.55) | 80.43 (68.97; 91.90) | 67.14 (56.14; 78.15) | 3.83 (2.04; 7.21) | 0.45 (0.32; 0.64) | 72.41 | |
| GDF-15 ≥ 975 | 71.93 (60.26; 83.59) | 92.86 (86.11; 99.60) | 91.11 (82.80; 99.43) | 76.47 (66.39; 86.55) | 10.07 (3.86; 26.26) | 0.30 (0.19; 0.46) | 82.30 | |
| Model (pGSN + GDF-15) ≥ 0.43 | 89.29 (81.18; 97.39) | 92.86 (86.11; 99.60) | 92.59 (85.61; 99.58) | 89.66 (81.82; 97.49) | 12.50 (4.84; 32.29) | 0.11 (0.05; 0.24) | 91.07 | |
| MD vs. non-MD | ||||||||
| FGF-21 ≥ 184 | 76.67 (65.96; 87.37) | 73.17 (59.61; 86.73) | 80.70 (70.46; 90.95) | 68.18 (54.42; 81.94) | 2.85 (1.69; 4.82) | 0.31 (0.19; 0.52) | 75.25 | |
| GDF-15 ≥ 1072 | 70.18 (58.30; 82.05) | 75.61 (62.46; 88.75) | 80.00 (68.91; 91.09) | 64.58 (51.05; 78.11) | 2.87 (1.63; 5.06) | 0.39 (0.25; 0.60) | 72.45 | |
| Subjects > 50 yrs old | MD vs. Control | |||||||
| pGSN ≤ 320 | 69.57 (50.76; 88.37) | 88.24 (77.41; 99.07) | 80.00 (62.47; 97.53) | 81.08 (68.46; 93.70) | 5.91 (2.26; 15.44) | 0.34 (0.18; 0.64) | 80.70 | |
| FGF-21 ≥ 300 | 73.91 (55.97; 91.86) | 73.53 (58.70; 88.36) | 65.38 (47.10; 83.67) | 80.65 (66.74; 94.55) | 2.79 (1.51; 5.14) | 0.35 (0.17; 0.72) | 73.68 | |
| GDF-15 ≥ 1227 | 80.95 (64.16; 97.75) | 94.12 (86.21; 100) | 89.47 (75.67; 100) | 88.89 (78.62; 99.15) | 13.76 (3.53; 53.66) | 0.20 (0.08; 0.49) | 89.09 | |
| MD vs. Non-MD | ||||||||
| FGF-21 ≥ 300 | 73.91 (55.97; 91.86) | 78.95 (60.62; 97.28) | 80.95 (64.16; 97.75) | 71.43 (52.11; 90.75) | 3.51 (1.42; 8.67) | 0.33 (0.15; 0.68) | 76.19 | |
| GDF-15 ≥ 1707 | 80.95 (64.16; 97.75) | 47.37 (24.92; 69.82) | 62.96 (44.75; 81.18) | 69.23 (44.14; 94.32) | 1.53 (0.95; 2.47) | 0.40 (0.14; 1.09) | 65.00 | |
| Non-MD vs. Control | ||||||||
| pGSN ≤ 240 | 63.16 (41.47; 84.45) | 97.06 (91.38; 100) | 92.31 (77.82; 100) | 82.50 (70.72; 94.28) | 21.48 (3.02; 152.70) | 0.37 (0.21; 0.68) | 84.91 | |
| Subjects ≤ 50 yrs old | MD vs. Control | |||||||
| pGSN ≤ 252 | 73.53 (58.70; 88.36) | 100 (100; 100) | 100 (100; 100) | 70.97 (54.99; 86.95) | na | 0.26 (0.15; 0.46) | 83.93 | |
| FGF-21 ≥ 225 | 65.71 (49.99; 81.44) | 90.91 (78.90; 100) | 92.00 (81.37; 100) | 62.50 (45.73; 79.27) | 7.22 (1.88; 27.68) | 0.37 (0.23; 0.60) | 75.44 | |
| GDF-15 ≥ 634 | 85.29 (73.39; 97.20) | 90.91 (78.90; 100) | 93.55 (84.90; 100) | 80.00 (64.32; 95.68) | 9.38 (2.48; 35.43) | 0.16 (0.07; 0.36) | 87.50 | |
| MD vs. Non-MD | ||||||||
| pGSN ≤ 310 | 79.41 (65.82; 93.00) | 52.38 (31.02; 73.74) | 72.97 (58.66; 87.28) | 61.11 (38.59; 83.63) | 1.66 (1.03; 2.69) | 0.39 (0.18; 0.85) | 69.09 | |
| FGF-21 ≥ 166 | 74.29 (59.81; 88.77) | 76.19 (57.97; 94.41) | 83.87 (70.92; 96.82) | 64.00 (45.18; 82.82) | 3.12 (1.41; 6.87) | 0.33 (0.18; 0.62) | 75.00 | |
| GDF-15 ≥ 636 | 85.29 (73.39; 97.20) | 80.95 (64.16; 97.75) | 87.88 (76.74; 99.01) | 77.27 (59.76; 94.78) | 4.47 (1.83; 10.93) | 0.18 (0.07; 0.41) | 83.64 | |
| Non-MD vs. Control | ||||||||
| pGSN ≤ 245 | 73.53 (58.70; 88.36) | 100 (100; 100) | 100 (100; 100) | 70.97 (54.99; 86.95) | na | 0.26 (0.15; 0.46) | 83.93 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peñas, A.; Fernández-De la Torre, M.; Laine-Menéndez, S.; Lora, D.; Illescas, M.; García-Bartolomé, A.; Morales-Conejo, M.; Arenas, J.; Martín, M.A.; Morán, M.; et al. Plasma Gelsolin Reinforces the Diagnostic Value of FGF-21 and GDF-15 for Mitochondrial Disorders. Int. J. Mol. Sci. 2021, 22, 6396. https://doi.org/10.3390/ijms22126396
Peñas A, Fernández-De la Torre M, Laine-Menéndez S, Lora D, Illescas M, García-Bartolomé A, Morales-Conejo M, Arenas J, Martín MA, Morán M, et al. Plasma Gelsolin Reinforces the Diagnostic Value of FGF-21 and GDF-15 for Mitochondrial Disorders. International Journal of Molecular Sciences. 2021; 22(12):6396. https://doi.org/10.3390/ijms22126396
Chicago/Turabian StylePeñas, Ana, Miguel Fernández-De la Torre, Sara Laine-Menéndez, David Lora, María Illescas, Alberto García-Bartolomé, Montserrat Morales-Conejo, Joaquín Arenas, Miguel A. Martín, María Morán, and et al. 2021. "Plasma Gelsolin Reinforces the Diagnostic Value of FGF-21 and GDF-15 for Mitochondrial Disorders" International Journal of Molecular Sciences 22, no. 12: 6396. https://doi.org/10.3390/ijms22126396
APA StylePeñas, A., Fernández-De la Torre, M., Laine-Menéndez, S., Lora, D., Illescas, M., García-Bartolomé, A., Morales-Conejo, M., Arenas, J., Martín, M. A., Morán, M., Domínguez-González, C., & Ugalde, C. (2021). Plasma Gelsolin Reinforces the Diagnostic Value of FGF-21 and GDF-15 for Mitochondrial Disorders. International Journal of Molecular Sciences, 22(12), 6396. https://doi.org/10.3390/ijms22126396