
The Divergent Immunomodulatory Effects of Short Chain Fatty Acids and Medium Chain Fatty Acids

 , ,
, , 
Abstract
:1. Introduction
2. Results
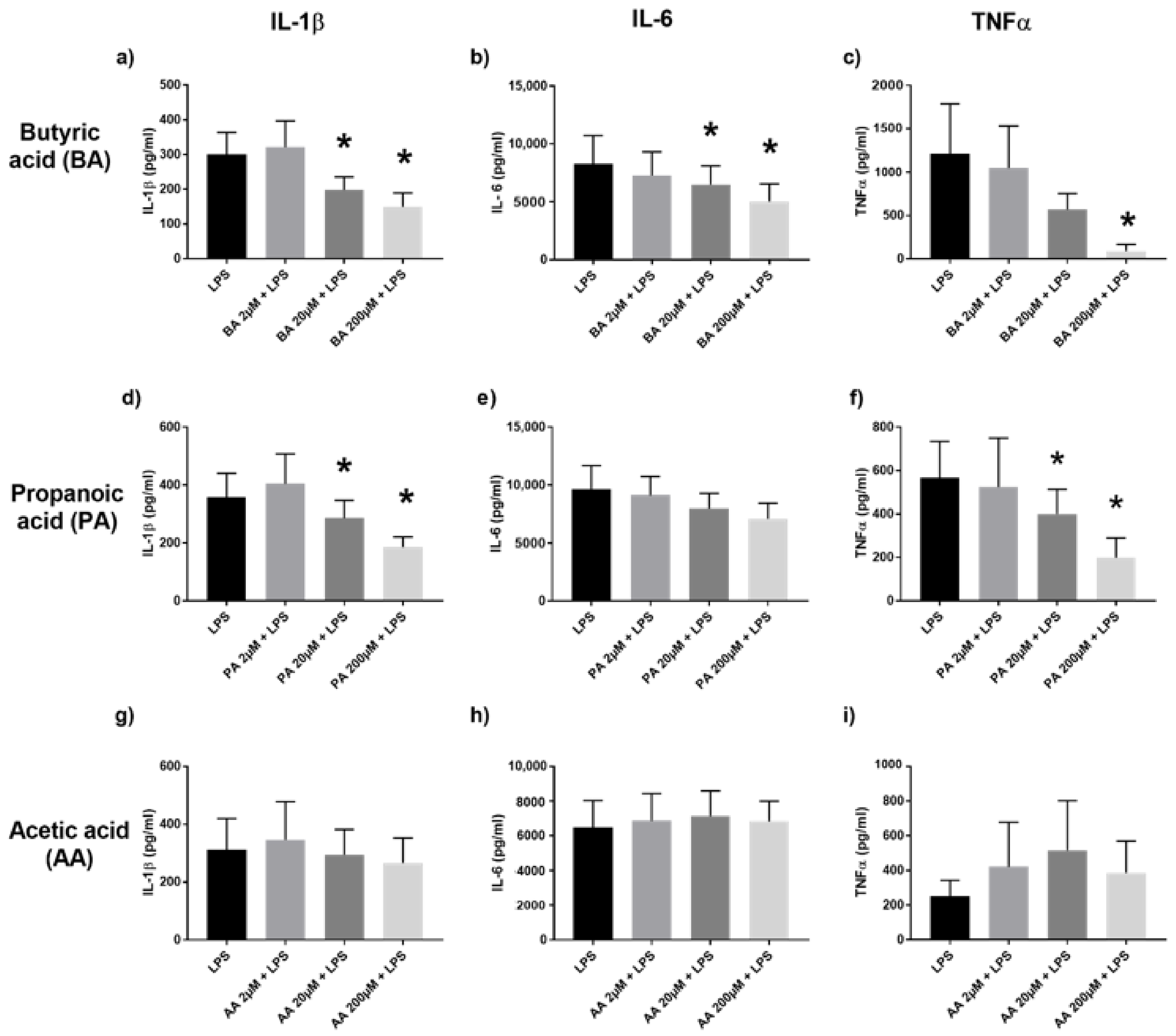
2.1. SCFA Down-Modulate the TLR4 Pathway
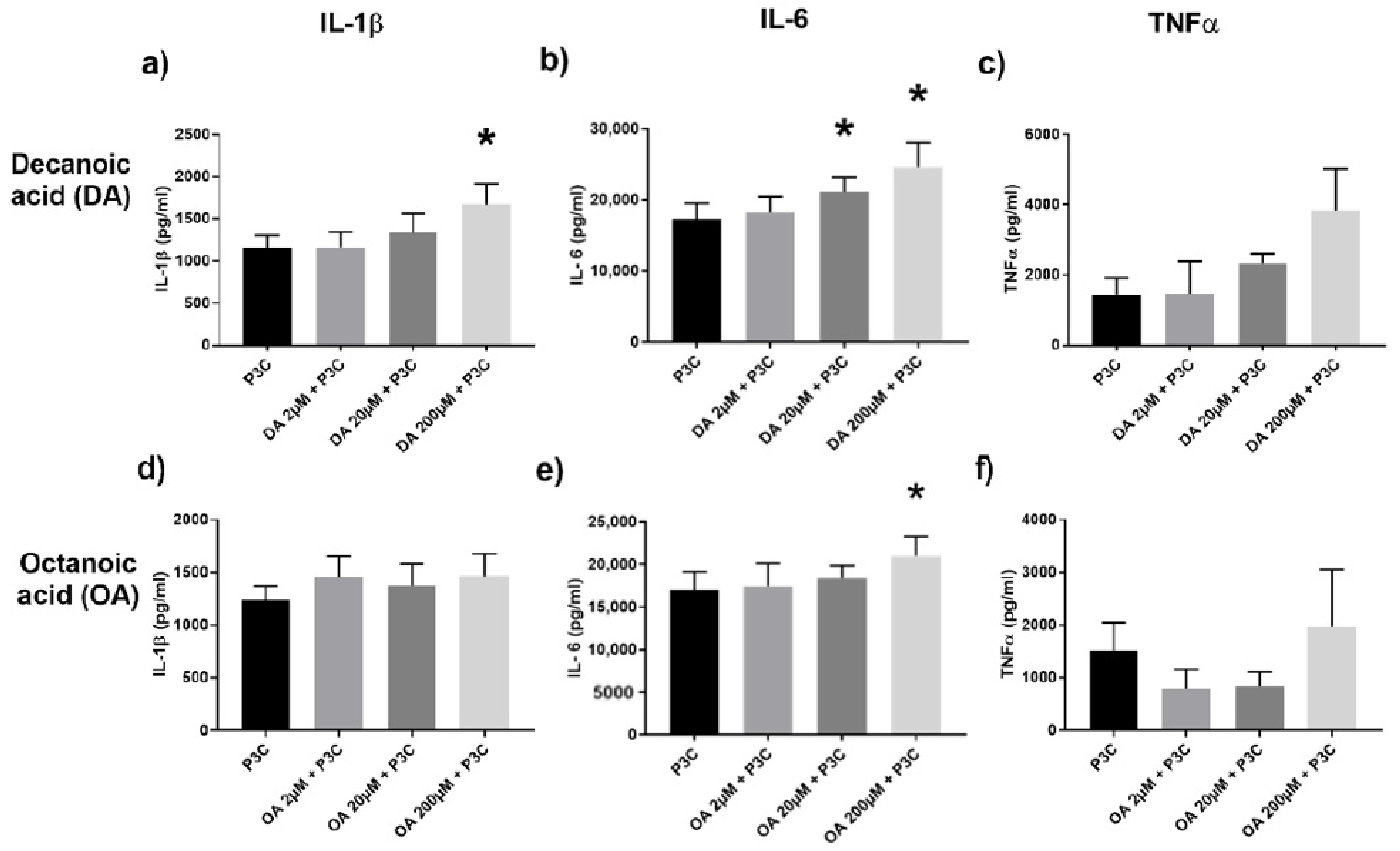
2.2. Conversely, MCFA Augment the TLR2 Pathway
2.3. Butyric Acid and Decanoic Acid Induce Variant Responses to Candida
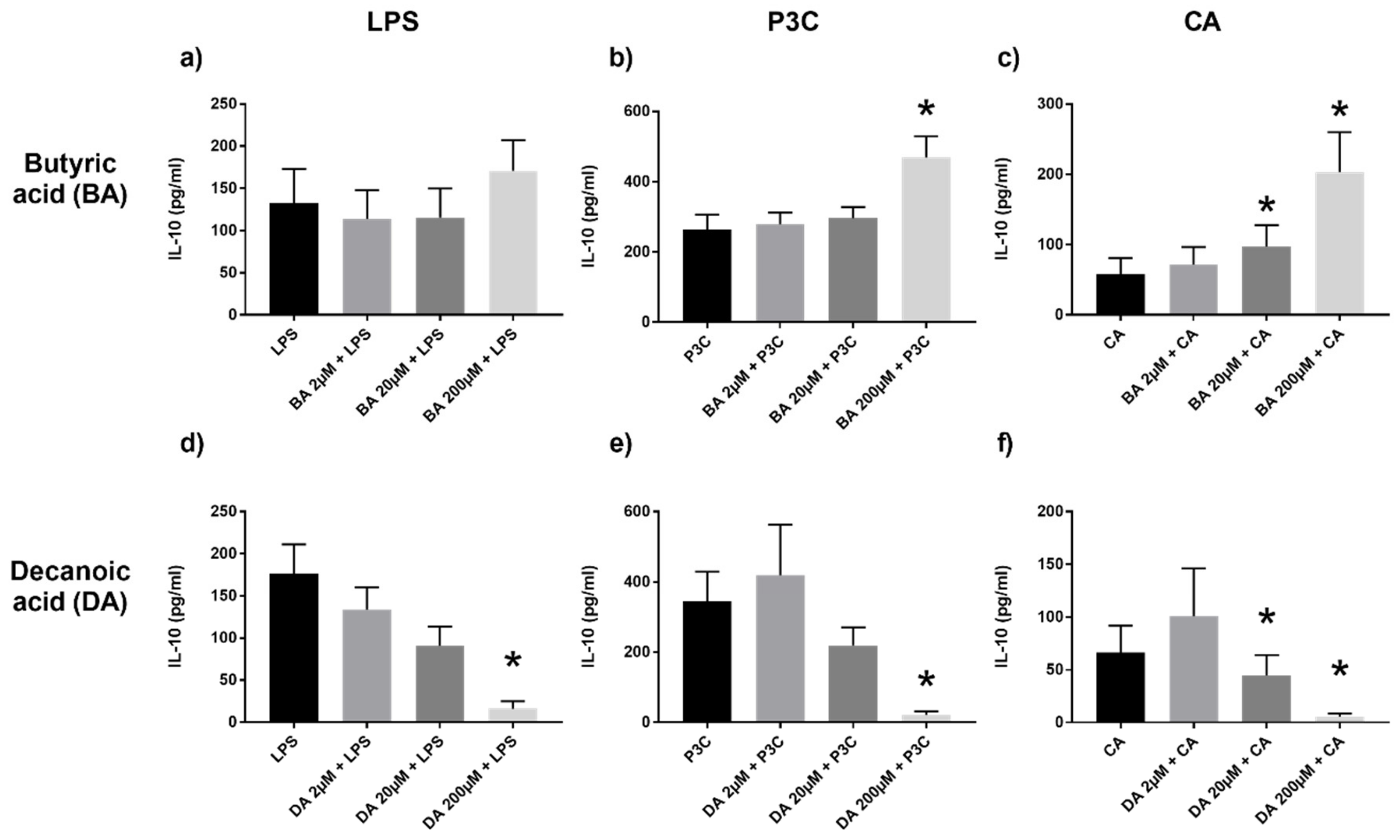
2.4. Anti-Inflammatory IL-10 Increased with SCFA and Decreased with MCFA
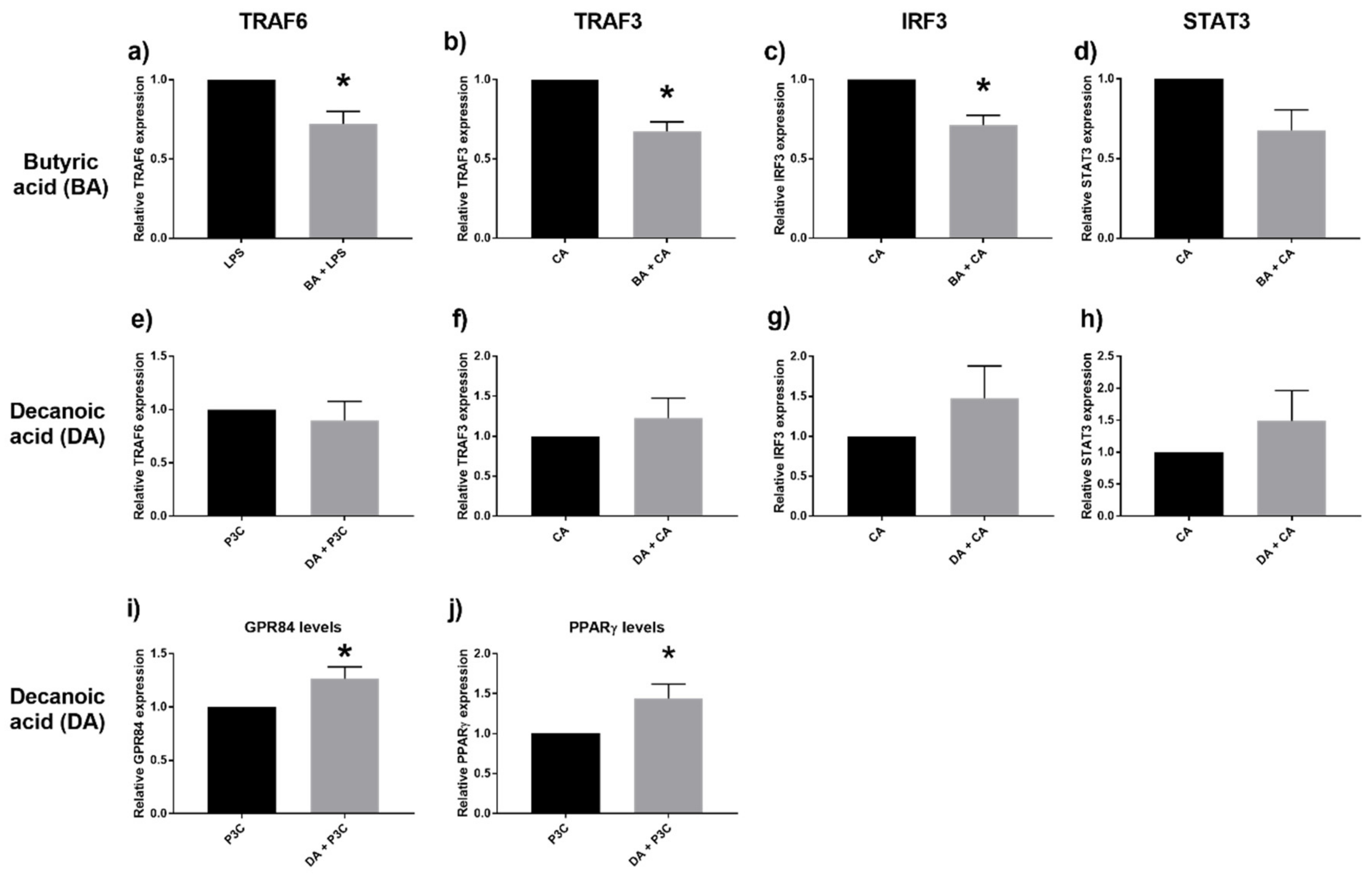
2.5. Down Regulation of TRAF3, IRF3, and TRAF6 with SCFA
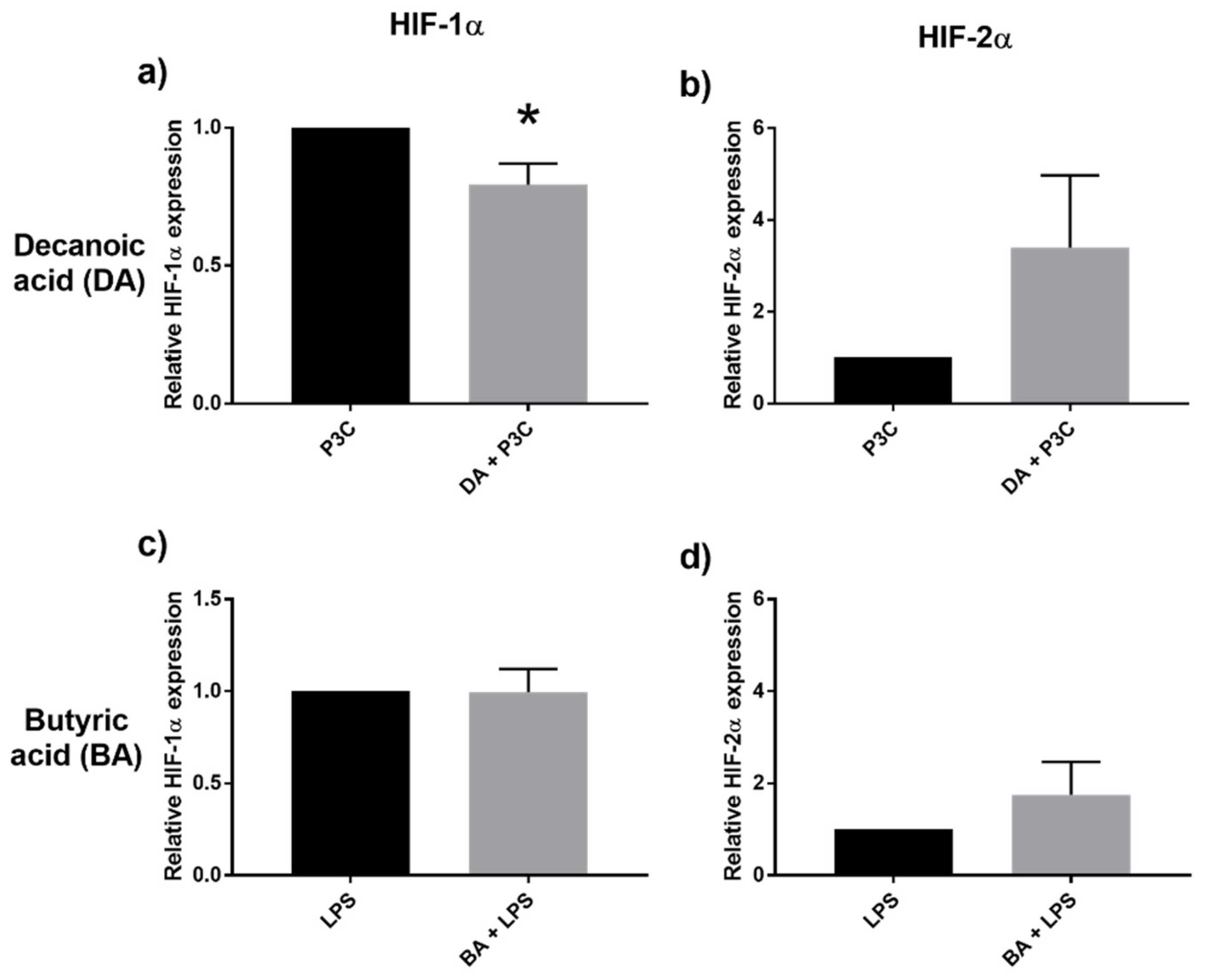
2.6. Decanoic Acid Diminished HIF-1α Transcription
3. Discussion
4. Materials and Methods
4.1. Stimuli and Reagents
4.2. PBMC Isolation and Stimulation Assays
4.3. Cytokine Analysis by ELISA
4.4. RNA Isolation and Quantitative Reverse-Transcription PCR (qPCR) Analysis
4.5. Statistical Analysis
4.6. Ethics Statement
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CA | Candida albicans |
| ELISA | Enzyme-Linked Immunosorbent Assay |
| HIF | Hypoxia-Inducible Factor |
| IL-1β | Interleukin–1 Beta |
| IL-6 | Interleukin–6 |
| LPS | Lipopolysaccharide |
| MCFA | Medium Chain Fatty Acid(s) |
| PBMC | Peripheral Blood Mononuclear Cells |
| P3C | Pam-3-Cys |
| qPCR | Quantitative Reverse-Transcription PCR |
| SCFA | Short Chain Fatty Acid(s) |
| STAT | Signal Transducer and Activator of Transcription |
| TNFα | Tumour Necrosis Factor Alpha |
| TRAF | Tumor necrosis factor receptor–associated factor |
References
- Levy, M.; Blacher, E.; Elinav, E. Microbiome, metabolites and host immunity. Curr. Opin. Microbiol. 2017, 35, 8–15. [Google Scholar] [CrossRef] [PubMed]
- O’Keefe, S.J.D. Diet, microorganisms and their metabolites, and colon cancer. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 691–706. [Google Scholar] [CrossRef] [PubMed]
- Dalile, B.; Van Oudenhove, L.; Vervliet, B.; Verbeke, K. The role of short-chain fatty acids in microbiota–gut–brain communication. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 461–478. [Google Scholar] [CrossRef]
- Parada Venegas, D.; De la Fuente, M.K.; Landskron, G.; González, M.J.; Quera, R.; Dijkstra, G.; Harmsen, H.J.M.; Faber, K.N.; Hermoso, M.A. Short Chain Fatty Acids (SCFAs)-mediated gut epithelial and immune regulation and its relevance for inflammatory bowel diseases. Front. Immunol. 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Takeuchi, H.; Sekine, S.; Kojima, K.; Aoyama, T. The application of medium-chain fatty acids: Edible oil with a suppressing effect on body fat accumulation. Asia Pac. J. Clin. Nutr. 2008, 17 (Suppl. 1), 320–323. [Google Scholar] [PubMed]
- Andoh, A.; Takaya, H.; Araki, Y.; Tsujikawa, T.; Fujiyama, Y.; Bamba, T. Medium- and long-chain fatty acids differentially modulate interleukin-8 secretion in human fetal intestinal epithelial cells. J. Nutr. 2000, 130, 2636–2640. [Google Scholar] [CrossRef] [PubMed]
- Lyudinina, A.Y.; Ivankova, G.E.; Bojko, E.R. Priority use of medium-chain fatty acids during high-intensity exercise in cross-country skiers. J. Int. Soc. Sports Nutr. 2018, 15, 57. [Google Scholar] [CrossRef] [Green Version]
- Sonnay, S.; Chakrabarti, A.; Thevenet, J.; Wiederkehr, A.; Christinat, N.; Masoodi, M. Differential Metabolism of medium-chain fatty acids in differentiated human-induced pluripotent stem cell-derived astrocytes. Front. Physiol. 2019, 10. [Google Scholar] [CrossRef]
- Li, L.; Wang, B.; Yu, P.; Wen, X.; Gong, D.; Zeng, Z. Medium and long chain fatty acids differentially modulate apoptosis and release of inflammatory cytokines in human liver cells. J. Food Sci. 2016, 81, H1546–H1552. [Google Scholar] [CrossRef]
- Tan, J.; McKenzie, C.; Potamitis, M.; Thorburn, A.N.; Mackay, C.R.; Macia, L. Chapter three-The role of short-Chain fatty acids in health and disease. In Advances in Immunology; Alt, F.W., Ed.; Academic Press: Cambridge, MA, USA, 2014; Volume 121, pp. 91–119. [Google Scholar]
- Segain, J.-P.; de la Blétière, D.R.; Bourreille, A.; Leray, V.; Gervois, N.; Rosales, C.; Ferrier, L.; Bonnet, C.; Blottière, H.M.; Galmiche, J.-P. Butyrate inhibits inflammatory responses through NFκB inhibition: Implications for Crohn’s disease. Gut 2000, 47, 397–403. [Google Scholar] [CrossRef] [Green Version]
- Thangaraju, M.; Cresci, G.A.; Liu, K.; Ananth, S.; Gnanaprakasam, J.P.; Browning, D.D.; Mellinger, J.D.; Smith, S.B.; Digby, G.J.; Lambert, N.A.; et al. GPR109A is a G-protein–Coupled receptor for the bacterial fermentation product butyrate and functions as a tumor suppressor in colon. Cancer Res. 2009, 69, 2826–2832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, M.A.; Jackson, J.; Stanton, M.; Rojas-Triana, A.; Bober, L.; Laverty, M.; Yang, X.; Zhu, F.; Liu, J.; Wang, S.; et al. Short-chain fatty acids act as antiinflammatory mediators by regulating prostaglandin E(2) and cytokines. World J. Gastroenterol. 2009, 15, 5549–5557. [Google Scholar] [CrossRef] [PubMed]
- Bloemen, J.G.; Venema, K.; van de Poll, M.C.; Olde Damink, S.W.; Buurman, W.A.; Dejong, C.H. Short chain fatty acids exchange across the gut and liver in humans measured at surgery. Clin. Nutr. 2009, 28, 657–661. [Google Scholar] [CrossRef] [PubMed]
- Peters, S.G.; Pomare, E.W.; Fisher, C.A. Portal and peripheral blood short chain fatty acid concentrations after caecal lactulose instillation at surgery. Gut 1992, 33, 1249. [Google Scholar] [CrossRef] [Green Version]
- Tamai, R.; Kiyoura, Y. Heat-killed Candida albicans augments synthetic bacterial component-induced proinflammatory cytokine production. Folia Microbiol. 2019, 64, 555–566. [Google Scholar] [CrossRef]
- Gantner, B.N.; Simmons, R.M.; Canavera, S.J.; Akira, S.; Underhill, D.M. Collaborative induction of inflammatory responses by dectin-1 and Toll-like receptor 2. J. Exp. Med. 2003, 197, 1107–1117. [Google Scholar] [CrossRef] [Green Version]
- Gow, N.A.; van de Veerdonk, F.L.; Brown, A.J.; Netea, M.G. Candida albicans morphogenesis and host defence: Discriminating invasion from colonization. Nat. Rev. Microbiol. 2011, 10, 112–122. [Google Scholar] [CrossRef] [Green Version]
- Sam, Q.H.; Chang, M.W.; Chai, L.Y.A. The fungal mycobiome and its interaction with gut bacteria in the host. Int. J. Mol. Sci. 2017, 18, 330. [Google Scholar] [CrossRef] [Green Version]
- Netea, M.G.; Brown, G.D.; Kullberg, B.J.; Gow, N.A. An integrated model of the recognition of Candida albicans by the innate immune system. Nat. Rev. Microbiol. 2008, 6, 67–78. [Google Scholar] [CrossRef]
- Jung, J.; Zeng, H.; Horng, T. Metabolism as a guiding force for immunity. Nat. Cell Biol. 2019, 21, 85–93. [Google Scholar] [CrossRef]
- Cuervo, A.; Salazar, N.; Ruas-Madiedo, P.; Gueimonde, M.; González, S. Fiber from a regular diet is directly associated with fecal short-chain fatty acid concentrations in the elderly. Nutr. Res. 2013, 33, 811–816. [Google Scholar] [CrossRef]
- Shimizu, H.; Masujima, Y.; Ushiroda, C.; Mizushima, R.; Taira, S.; Ohue-Kitano, R.; Kimura, I. Dietary short-chain fatty acid intake improves the hepatic metabolic condition via FFAR3. Sci. Rep. 2019, 9, 16574. [Google Scholar] [CrossRef] [Green Version]
- Frank, D.N.; Amand, A.L.S.; Feldman, R.A.; Boedeker, E.C.; Harpaz, N.; Pace, N.R. Molecular-phylogenetic characterization of microbial community imbalances in human inflammatory bowel diseases. Proc. Natl. Acad. Sci. USA 2007, 104, 13780–13785. [Google Scholar] [CrossRef] [Green Version]
- Treem, W.R.; Ahsan, N.; Shoup, M.; Hyams, J.S. Fecal short-chain fatty acids in children with inflammatory bowel disease. J. Pediatr. Gastroenterol. Nutr. 1994, 18, 159–164. [Google Scholar] [CrossRef]
- Ferwerda, B.; McCall, M.B.B.; Alonso, S.; Giamarellos-Bourboulis, E.J.; Mouktaroudi, M.; Izagirre, N.; Syafruddin, D.; Kibiki, G.; Cristea, T.; Hijmans, A.; et al. TLR4 polymorphisms, infectious diseases, and evolutionary pressure during migration of modern humans. Proc. Natl. Acad. Sci. USA 2007, 104, 16645–16650. [Google Scholar] [CrossRef] [Green Version]
- Jakus, P.B.; Kalman, N.; Antus, C.; Radnai, B.; Tucsek, Z.; Gallyas, F., Jr.; Sumegi, B.; Veres, B. TRAF6 is functional in inhibition of TLR4-mediated NF-κB activation by resveratrol. J. Nutr. Biochem. 2013, 24, 819–823. [Google Scholar] [CrossRef]
- Yamamoto, M.; Sato, S.; Hemmi, H.; Hoshino, K.; Kaisho, T.; Sanjo, H.; Takeuchi, O.; Sugiyama, M.; Okabe, M.; Takeda, K.; et al. Role of adaptor TRIF in the MyD88-Independent toll-like receptor signaling pathway. Science 2003, 301, 640–643. [Google Scholar] [CrossRef]
- Li, J.; Lee, D.; Madrenas, J. Evolving bacterial envelopes and plasticity of TLR2-dependent responses: Basic research and translational opportunities. Front. Immunol. 2013, 4. [Google Scholar] [CrossRef] [Green Version]
- Nastasi, C.; Candela, M.; Bonefeld, C.M.; Geisler, C.; Hansen, M.; Krejsgaard, T.; Biagi, E.; Andersen, M.H.; Brigidi, P.; Ødum, N.; et al. The effect of short-chain fatty acids on human monocyte-derived dendritic cells. Sci. Rep. 2015, 5, 16148. [Google Scholar] [CrossRef]
- Meissner, F.; Scheltema, R.A.; Mollenkopf, H.J.; Mann, M. Direct proteomic quantification of the secretome of activated immune cells. Science 2013, 340, 475–478. [Google Scholar] [CrossRef]
- Suzuki, M.; Takaishi, S.; Nagasaki, M.; Onozawa, Y.; Iino, I.; Maeda, H.; Komai, T.; Oda, T. Medium-chain fatty acid-sensing receptor, GPR84, is a proinflammatory receptor. J. Biol. Chem. 2013, 288, 10684–10691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.-Y.; Della-Fera, M.A.; Rayalam, S.; Park, H.J.; Ambati, S.; Hausman, D.B.; Hartzell, D.L.; Baile, C.A. Regulation of adipogenesis by medium-chain fatty acids in the absence of hormonal cocktail. J. Nutr. Biochem. 2009, 20, 537–543. [Google Scholar] [CrossRef] [PubMed]
- Downes, N.L.; Laham-Karam, N.; Kaikkonen, M.U.; Ylä-Herttuala, S. Differential but complementary HIF1α and HIF2α transcriptional regulation. Mol. Ther. 2018, 26, 1735–1745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelly, C.J.; Zheng, L.; Campbell, E.L.; Saeedi, B.; Scholz, C.C.; Bayless, A.J.; Wilson, K.E.; Glover, L.E.; Kominsky, D.J.; Magnuson, A.; et al. Crosstalk between microbiota-derived short-chain fatty acids and intestinal epithelial HIF augments tissue barrier function. Cell Host Microbe 2015, 17, 662–671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rankin, E.B.; Rha, J.; Selak, M.A.; Unger, T.L.; Keith, B.; Liu, Q.; Haase, V.H. Hypoxia-inducible factor 2 regulates hepatic lipid metabolism. Mol. Cell Biol. 2009, 29, 4527–4538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, J.H.J.; Ravikumar, S.; Wang, Y.-M.; Thamboo, T.P.; Ong, L.; Chen, J.; Goh, J.G.; Tay, S.H.; Chengchen, L.; Win, M.S.; et al. Bimodal influence of Vitamin D in host response to systemic candida infection—Vitamin D dose matters. J. Infect. Dis. 2015, 212, 635–644. [Google Scholar] [CrossRef] [Green Version]
- Goh, J.G.; Ravikumar, S.; Win, M.S.; Cao, Q.; Tan, A.L.; Lim, J.H.J.; Leong, W.; Herbrecht, R.; Troke, P.F.; Kullberg, B.J.; et al. Neutrophils differentially attenuate immune response to Aspergillus infection through complement receptor 3 and induction of myeloperoxidase. Cell. Microbiol. 2018, 20, e12798. [Google Scholar] [CrossRef]
- Redonnet, A.; Bonilla, S.; Noël-Suberville, C.; Pallet, V.; Dabadie, H.; Gin, H.; Higueret, P. Relationship between peroxisome proliferator-activated receptor gamma and retinoic acid receptor alpha gene expression in obese human adipose tissue. Int. J. Obes. 2002, 26, 920–927. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Wu, X.; Simonavicius, N.; Tian, H.; Ling, L. Medium-chain fatty acids as ligands for orphan G protein-coupled receptor GPR84. J. Biol. Chem. 2006, 281, 34457–34464. [Google Scholar] [CrossRef] [Green Version]
- Krotova, K.; Patel, J.M.; Block, E.R.; Zharikov, S. Hypoxic upregulation of arginase II in human lung endothelial cells. Am. J. Physiol. Cell Physiol. 2010, 299, C1541–C1548. [Google Scholar] [CrossRef]
- Ebersole, J.L.; Novak, M.J.; Orraca, L.; Martinez-Gonzalez, J.; Kirakodu, S.; Chen, K.C.; Stromberg, A.; Gonzalez, O.A. Hypoxia-inducible transcription factors, HIF1A and HIF2A, increase in aging mucosal tissues. Immunology 2018, 154, 452–464. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer (Human) | Sequence (5′-3′) | References |
|---|---|---|
| B2M_F | ATG AGT ATG CCT GCC GTG TG | [37,38] |
| B2M_R | CCA AAT GCG GCA TCT TCA AAC | [37,38] |
| STAT3_F | CAG CAG CTT GAC ACA CGG TA | [37] |
| STAT3_R | AAA CAC CAA AGT GGC ATG TGA | [37] |
| TRAF3_F | TCT TGA GGA AAG ACC TGC GAG | [38] |
| TRAF3_R | GCG ATC ATC GGA ACC TGA CT | [38] |
| TRAF6_F | TTG CCA TGA AAA GAT GCA GAG G | [38] |
| TRAF6_R | AGC CTG GGC CAA CAT TCT C | [38] |
| IRF3_F | AGA GGC TCG TGA TGG TCA AG | [38] |
| IRF3_R | AGG TCC ACA GTA TTC TCC AGG | [38] |
| PPARγ_F | CACAAGAACAGATCCAGTGGTTGCAG | [39] |
| PPARγ_R | AATAATAAGGTGGAGATGCAGGCTCC | [39] |
| GPR84_F | TTCAGCCCTTCTCTGTGGACA | [40] |
| GPR84_R | TGCAGAAGGTGGCACCG | [40] |
| HIF1α_F | TGCTCATCAGTTGCCACTTC | [41] |
| HIF1α_R | TCCTCACACGCAAATAGCTG | [41] |
| HIF2A_F | GAAGCGACAGCTGGAGTATG | [42] |
| HIF2A_R | TGAGGTTCTTCATCCGTTTCC | [42] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sam, Q.H.; Ling, H.; Yew, W.S.; Tan, Z.; Ravikumar, S.; Chang, M.W.; Chai, L.Y.A. The Divergent Immunomodulatory Effects of Short Chain Fatty Acids and Medium Chain Fatty Acids. Int. J. Mol. Sci. 2021, 22, 6453. https://doi.org/10.3390/ijms22126453
Sam QH, Ling H, Yew WS, Tan Z, Ravikumar S, Chang MW, Chai LYA. The Divergent Immunomodulatory Effects of Short Chain Fatty Acids and Medium Chain Fatty Acids. International Journal of Molecular Sciences. 2021; 22(12):6453. https://doi.org/10.3390/ijms22126453
Chicago/Turabian StyleSam, Qi Hui, Hua Ling, Wen Shan Yew, Zhaohong Tan, Sharada Ravikumar, Matthew Wook Chang, and Louis Yi Ann Chai. 2021. "The Divergent Immunomodulatory Effects of Short Chain Fatty Acids and Medium Chain Fatty Acids" International Journal of Molecular Sciences 22, no. 12: 6453. https://doi.org/10.3390/ijms22126453
APA StyleSam, Q. H., Ling, H., Yew, W. S., Tan, Z., Ravikumar, S., Chang, M. W., & Chai, L. Y. A. (2021). The Divergent Immunomodulatory Effects of Short Chain Fatty Acids and Medium Chain Fatty Acids. International Journal of Molecular Sciences, 22(12), 6453. https://doi.org/10.3390/ijms22126453