
The Molecular Aspects of Disturbed Platelet Activation through ADP/P2Y12 Pathway in Multiple Sclerosis



Abstract
:1. Introduction
2. Results
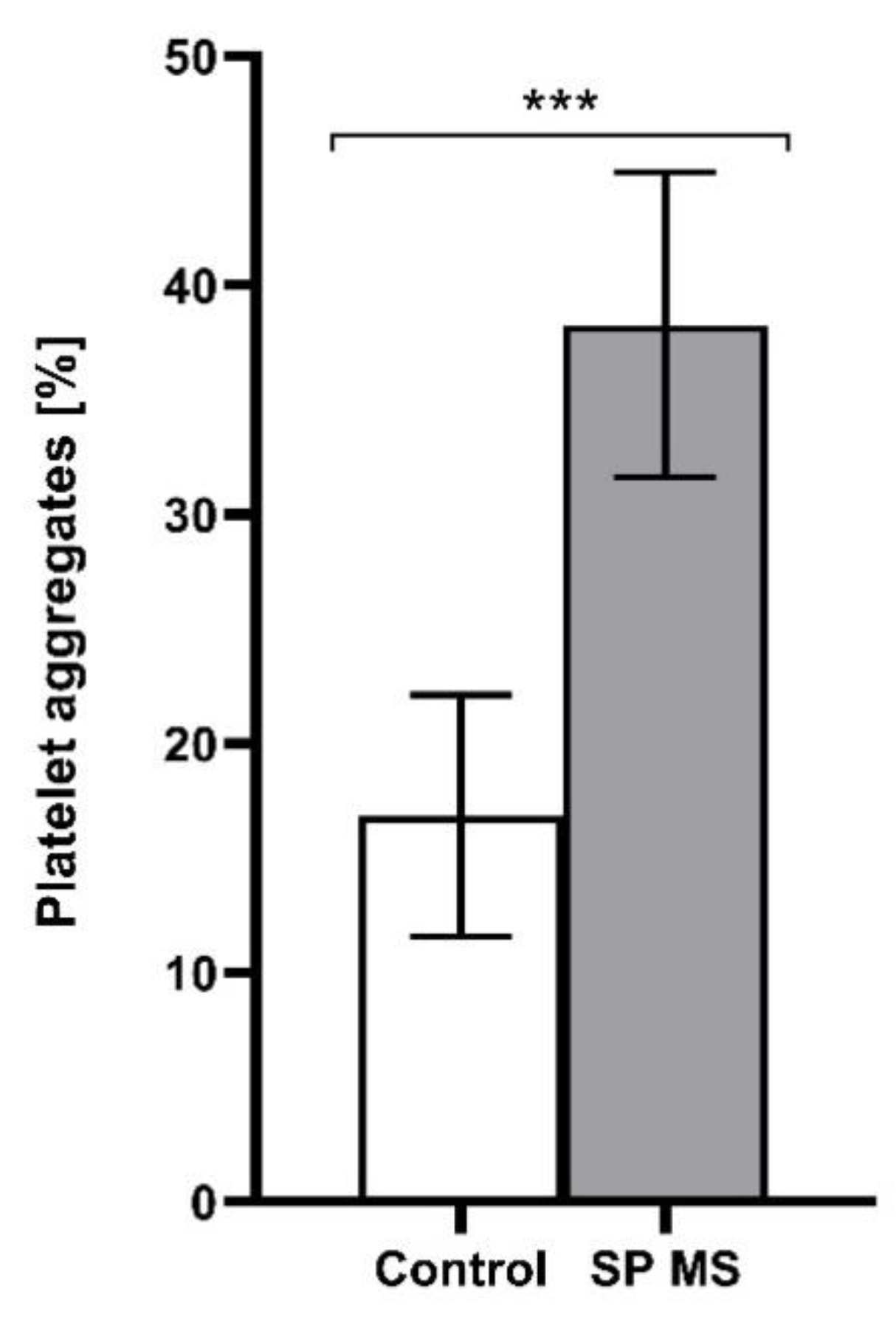
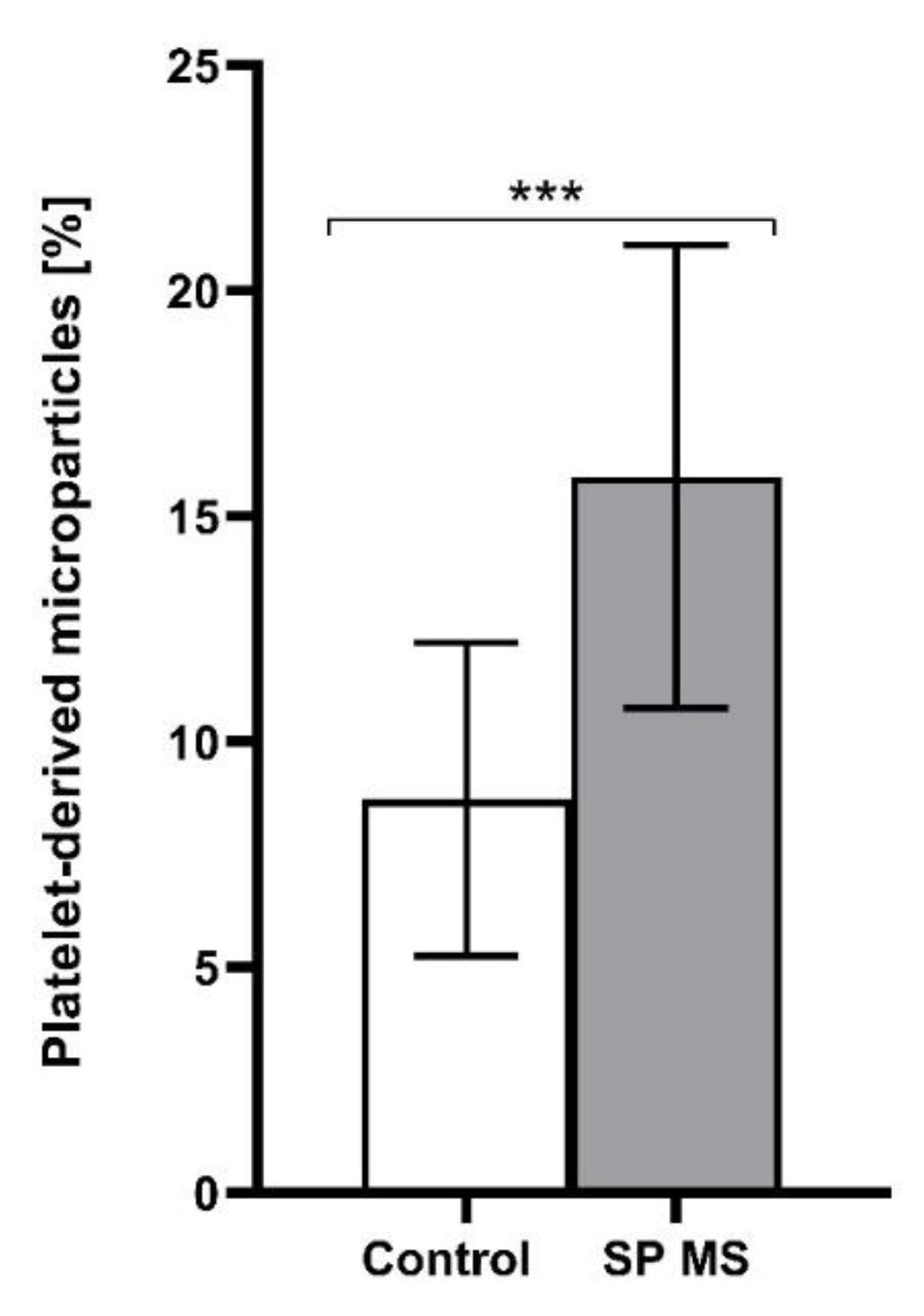
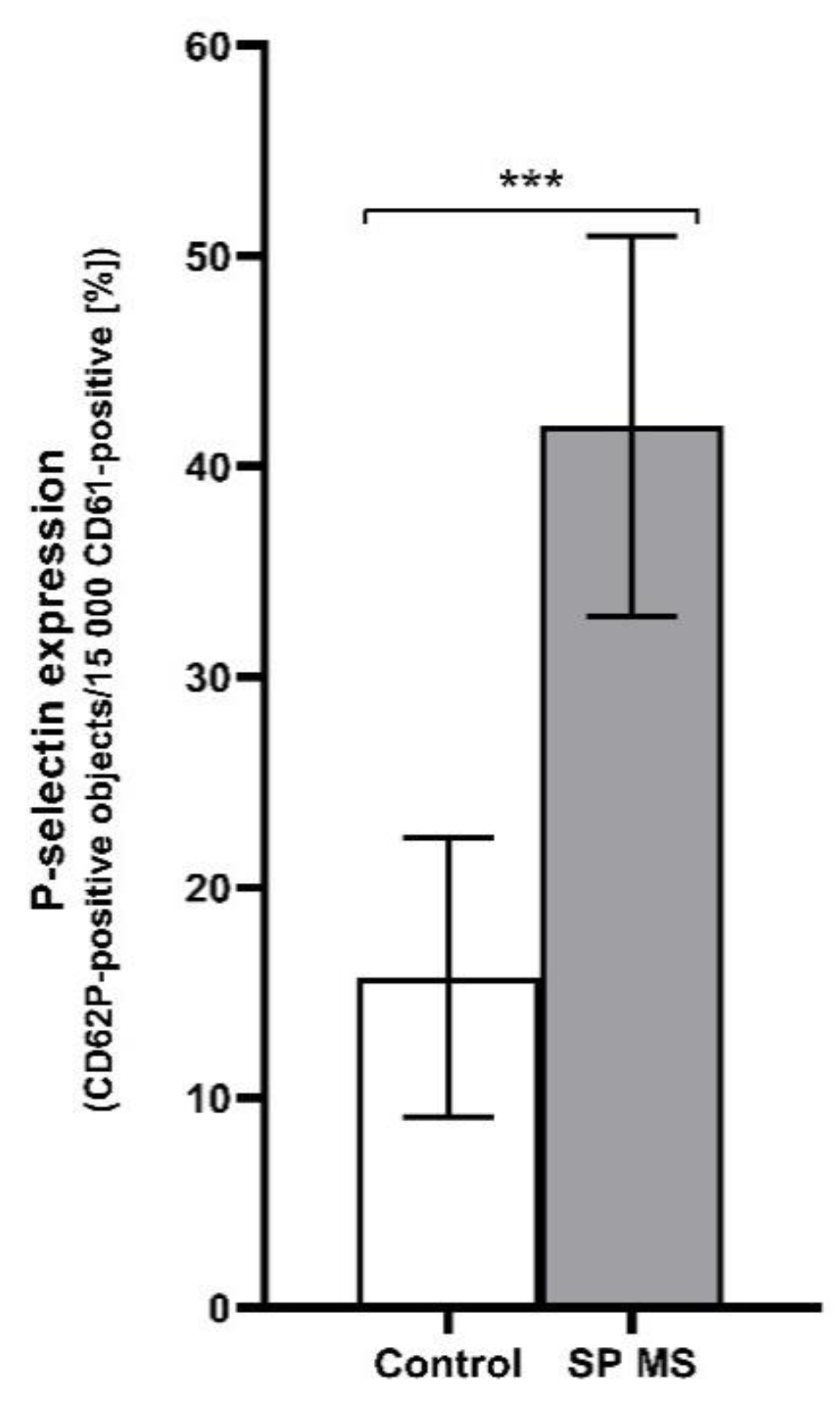
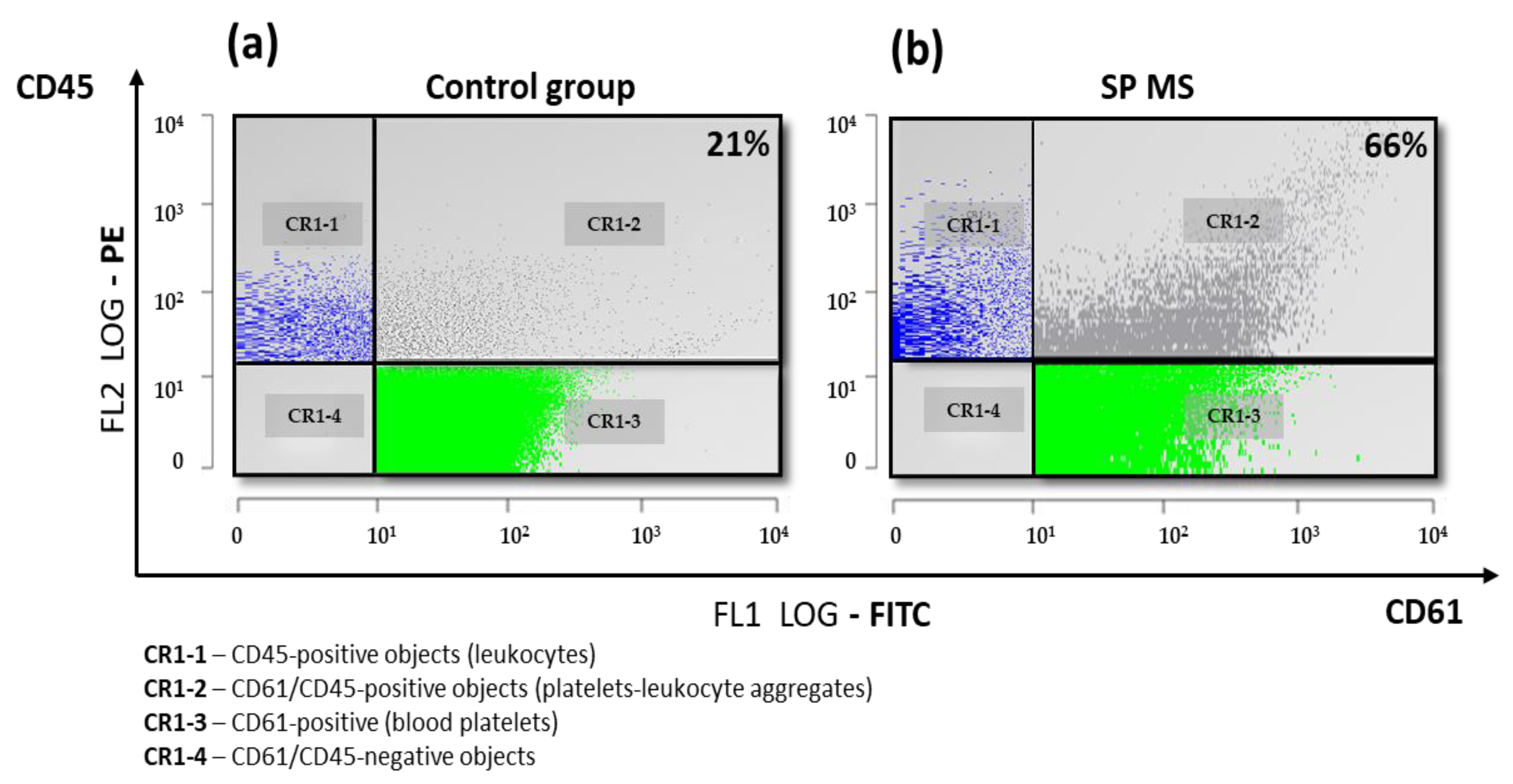
2.1. The Level of Platelets’ Markers of Activation
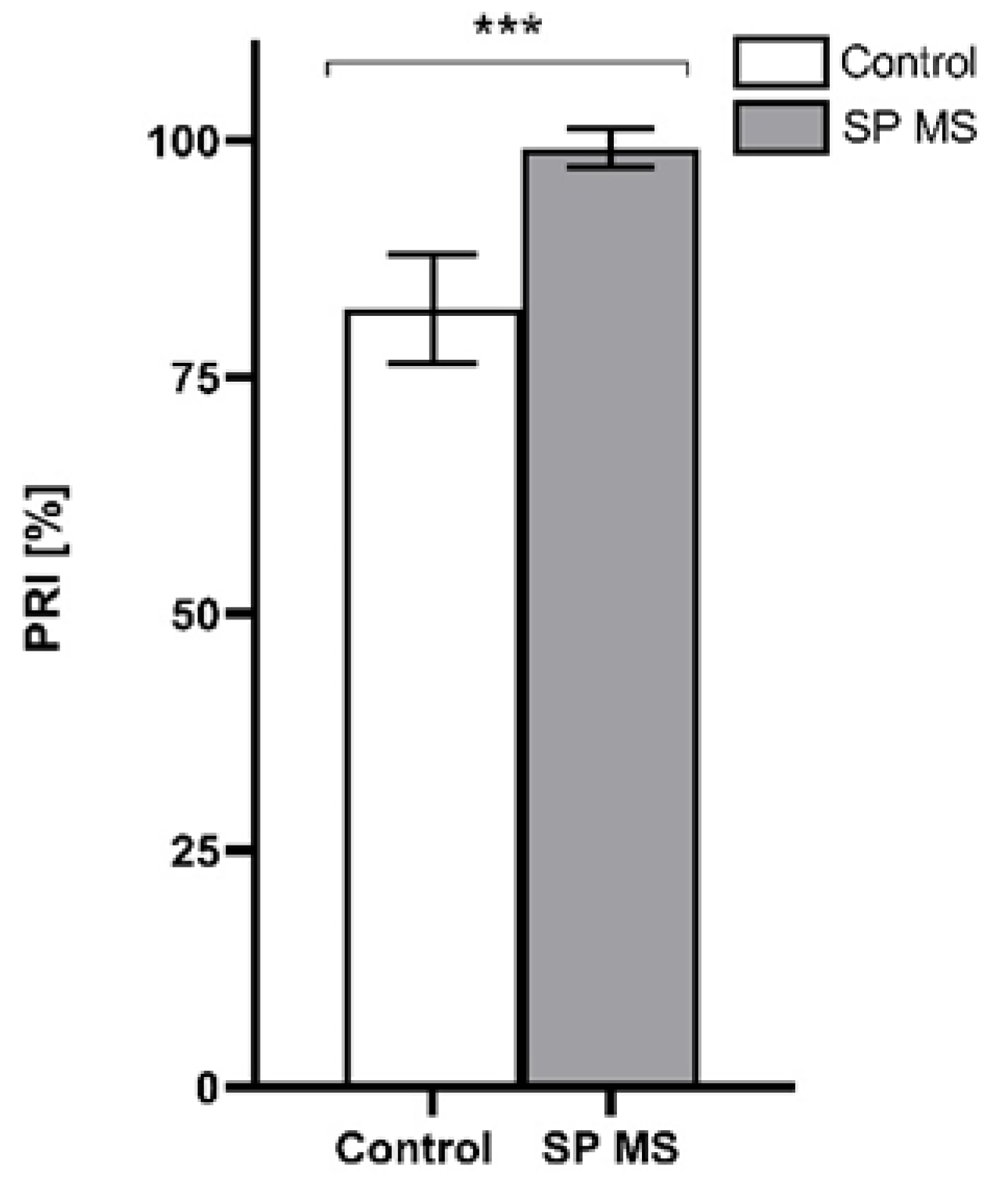
2.2. The Measurement of PRI [%]
2.3. P2Y12 Receptor Expression on Blood Platelets [%]
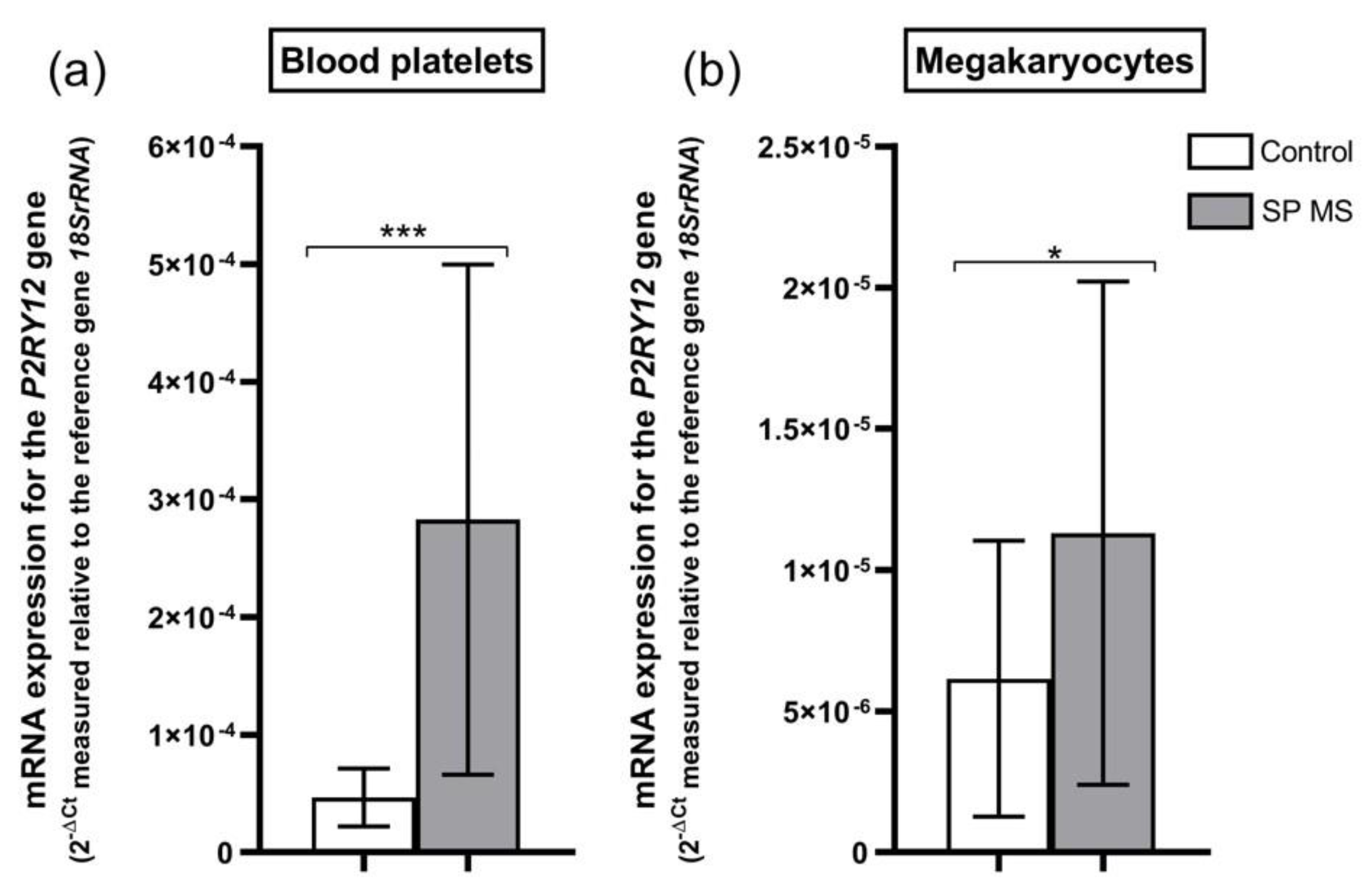
2.4. mRNA Expression for P2RY12 Gene in Blood Platelets and Megakaryocytes
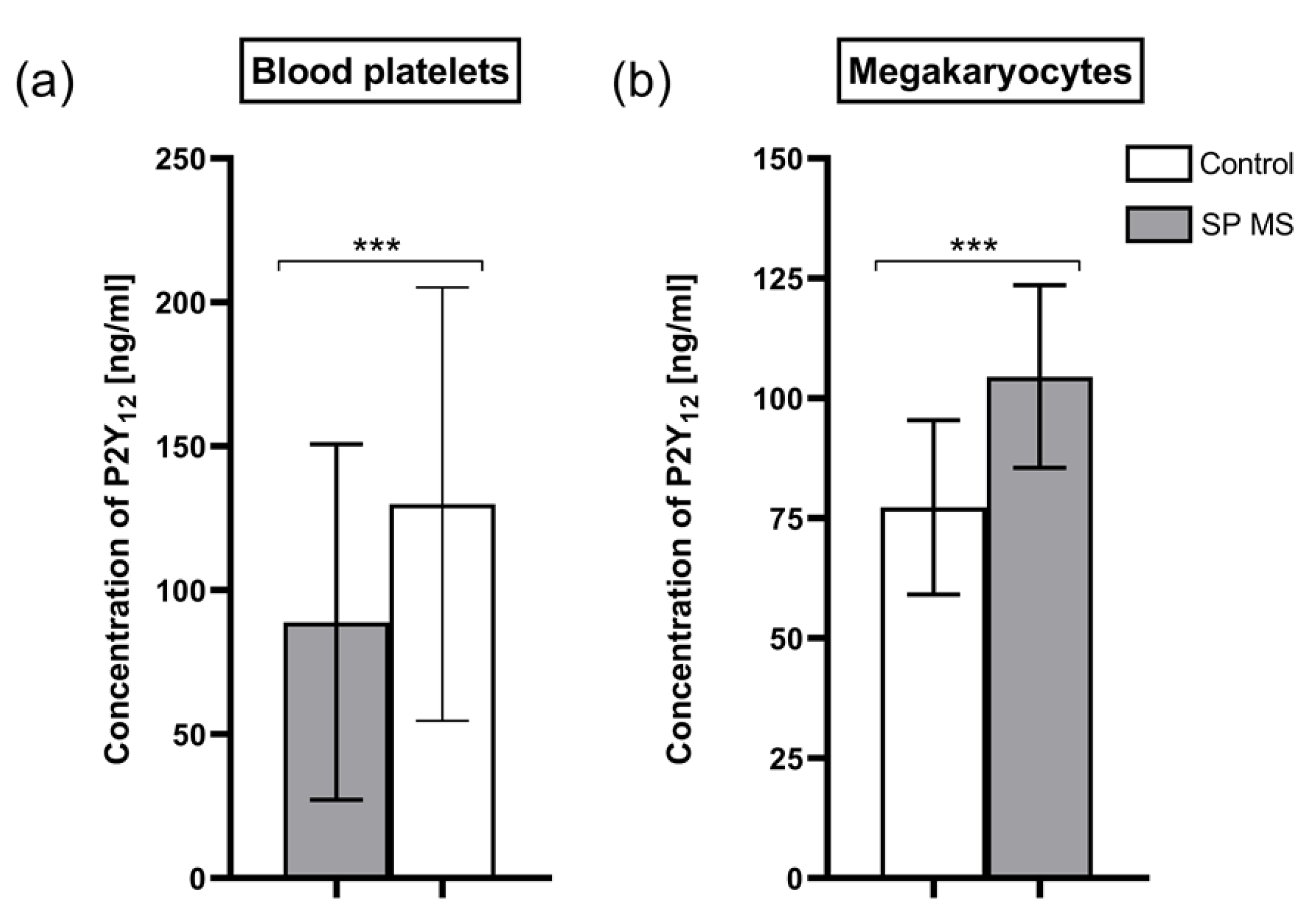
2.5. Concentration of P2Y12 Molecules in Blood Platelets and Megakaryocytes
3. Discussion
4. Materials and Methods
4.1. Cytometric Measurements
4.2. Assessment of PRI Using the VASP/P2Y12 Assay
4.3. Isolation of Blood Platelets
4.4. Isolation of Megakaryocytes
4.5. Isolation of mRNA and cDNA Synthesis
4.6. Expression of the P2RY12 Gene at the mRNA Level in Blood Platelets and Megakaryocytes
4.7. Concentration of P2Y12 Receptor in Blood Platelets and Megakaryocytes
4.8. Statistical Analysis
5. Conclusions
6. Patients
Clinical Characteristics of Patients
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rawish, E.; Nording, H.; Münte, T.; Langer, H.F. Platelets as Mediators of Neuroinflammation and Thrombosis. Front. Immunol. 2020, 11, 548631. [Google Scholar] [CrossRef]
- Li, C.; Li, J.; Li, Y.; Lang, S.; Yougbare, I.; Zhu, G.; Chen, P.; Ni, H. Crosstalk between Platelets and the Immune System: Old Systems with New Discoveries. Adv. Hematol. 2012, 2012, 384685. [Google Scholar] [CrossRef] [Green Version]
- Eisinger, F.; Patzelt, J.; Langer, H.F. The Platelet Response to Tissue Injury. Front. Med. 2018, 5, 317. [Google Scholar] [CrossRef] [Green Version]
- Tomaiuolo, M.; Brass, L.F.; Stalker, T.J. Regulation of Platelet Activation and Coagulation and Its Role in Vascular Injury and Arterial Thrombosis. Interv. Cardiol. Clin. 2017, 6, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Hu, H.; Lindqvist, M.; Wikström-Jonsson, E.; Goodall, A.H.; Hjemdahl, P. Platelet-Leukocyte Cross Talk in Whole Blood. Arter. Thromb. Vasc. Biol. 2000, 20, 2702–2708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rainger, G.E.; Chimen, M.; Harrison, M.J.; Yates, C.M.; Harrison, P.; Watson, S.; Lordkipanidzé, M.; Nash, G.B. The role of platelets in the recruitment of leukocytes during vascular disease. Platelets 2015, 26, 507–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossaint, J.; Margraf, A.; Zarbock, A. Role of Platelets in Leukocyte Recruitment and Resolution of Inflammation. Front. Immunol. 2018, 9, 2712. [Google Scholar] [CrossRef] [PubMed]
- Lisman, T. Platelet–neutrophil interactions as drivers of inflammatory and thrombotic disease. Cell Tissue Res. 2018, 371, 567–576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossaint, J.; Zarbock, A. Platelets in leucocyte recruitment and function. Cardiovasc. Res. 2015, 107, 386–395. [Google Scholar] [CrossRef] [Green Version]
- Minagar, A.; Alexander, J.S. Blood-brain barrier disruption in multiple sclerosis. Mult. Scler. J. 2003, 9, 540–549. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.-J.; Chen, W.-W.; Zhang, X. Multiple sclerosis: Pathology, diagnosis and treatments. Exp. Ther. Med. 2017, 13, 3163–3166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobson, R.; Giovannoni, G. Multiple sclerosis—A review. Eur. J. Neurol. 2019, 26, 27–40. [Google Scholar] [CrossRef] [Green Version]
- Ciabattoni, G.; Porreca, E.; Di Febbo, C.; Di Iorio, A.; Paganelli, R.; Bucciarelli, T.; Pescara, L.; Del Re, L.; Giusti, C.; Falco, A.; et al. Determinants of platelet activation in Alzheimer’s disease. Neurobiol. Aging 2007, 28, 336–342. [Google Scholar] [CrossRef] [PubMed]
- Denis, H.L.; Lamontagne-Proulx, J.; St-Amour, I.; Mason, S.L.; Rowley, J.W.; Cloutier, N.; Tremblay, M.-È.; Vincent, A.T.; Gould, P.V.; Chouinard, S.; et al. Platelet abnormalities in Huntington’s disease. J. Neurol. Neurosurg. Psychiatry 2018, 90, 272–283. [Google Scholar] [CrossRef] [Green Version]
- Shrivastava, M.; Das, T.K.; Behari, M.; Pati, U.; Vivekanandhan, S. Ultrastructural Variations in Platelets and Platelet Mitochondria: A Novel Feature in Amyotrophic Lateral Sclerosis. Ultrastruct. Pathol. 2011, 35, 52–59. [Google Scholar] [CrossRef]
- A Sheremata, W.; Jy, W.; Horstman, L.L.; Ahn, Y.S.; Alexander, J.S.; Minagar, A. Evidence of platelet activation in multiple sclerosis. J. Neuroinflamm. 2008, 5, 27. [Google Scholar] [CrossRef] [Green Version]
- Bidot, C.J.; Horstman, L.L.; Jy, W.; Jimenez, J.J.; Ahn, Y.S.; Alexander, J.S.; Gonzalez-Toledo, E.; Kelley, R.E.; Minagar, A.; Bidot, C. Clinical and neuroimaging correlates of antiphospholipid antibodies in multiple sclerosis: A preliminary study. BMC Neurol. 2007, 7, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spanevello, R.M.; Mazzanti, C.M.; Maldonado, P.A.; Zanin, R.; Morsch, A.; Hannel, L.; Mazzanti, A.; Festugatto, R.; Graça, D.; Schmatz, R.; et al. Activities of enzymes that hydrolyze adenine nucleotides in platelets from rats experimentally demyelinated with ethidium bromide and treated with interferon-β. Life Sci. 2007, 80, 1109–1114. [Google Scholar] [CrossRef]
- Klineova, S.; Lublin, F.D. Clinical Course of Multiple Sclerosis. Cold Spring Harb. Perspect. Med. 2018, 8, a028928. [Google Scholar] [CrossRef]
- Roshanisefat, H.; Bahmanyar, S.; Hillert, J.; Olsson, T.; Montgomery, S. Multiple sclerosis clinical course and cardiovascular disease risk—Swedish cohort study. Eur. J. Neurol. 2014, 21, 1353–1388. [Google Scholar] [CrossRef]
- Moccia, M.; Lanzillo, R.; Palladino, R.; Maniscalco, G.T.; De Rosa, A.; Russo, C.; Massarelli, M.; Carotenuto, A.; Postiglione, E.; Caporale, O.; et al. The Framingham cardiovascular risk score in multiple sclerosis. Eur. J. Neurol. 2015, 22, 1176–1183. [Google Scholar] [CrossRef] [Green Version]
- Cerda, F.P.; Gomez, M.V.S.; Matute, C. The link of inflammation and neurodegeneration in progressive multiple sclerosis. Mult. Scler. Demyelinating Disord. 2016, 1. [Google Scholar] [CrossRef]
- Morel, A.; Rywaniak, J.; Bijak, M.; Miller, E.; Niwald, M.; Saluk, J. Flow cytometric analysis reveals the high levels of platelet activation parameters in circulation of multiple sclerosis patients. Mol. Cell. Biochem. 2017, 430, 69–80. [Google Scholar] [CrossRef] [Green Version]
- Dziedzic, A.; Morel, A.; Miller, E.; Bijak, M.; Sliwinski, T.; Synowiec, E.; Ceremuga, M.; Saluk-Bijak, J. Oxidative Damage of Blood Platelets Correlates with the Degree of Psychophysical Disability in Secondary Progressive Multiple Sclerosis. Oxidative Med. Cell. Longev. 2020, 2020, 2868014. [Google Scholar] [CrossRef]
- Dziedzic, A.; Miller, E.; Bijak, M.; Przyslo, L.; Saluk-Bijak, J. Increased Pro-Thrombotic Platelet Activity Associated with Thrombin/PAR1-Dependent Pathway Disorder in Patients with Secondary Progressive Multiple Sclerosis. Int. J. Mol. Sci. 2020, 21, 7722. [Google Scholar] [CrossRef] [PubMed]
- Shankar, H.; Garcia, A.; Prabhakar, J.; Kim, S.; Kunapuli, S.P. P2Y12 receptor-mediated potentiation of thrombin-induced thromboxane A2 generation in platelets occurs through regulation of Erk1/2 activation. J. Thromb. Haemost. 2006, 4, 638–647. [Google Scholar] [CrossRef] [PubMed]
- Rauch, B.H.; Rosenkranz, A.C.; Ermler, S.; Böhm, A.; Driessen, J.; Fischer, J.W.; Sugidachi, A.; Jakubowski, J.A.; Schrör, K. Regulation of Functionally Active P2Y12 ADP Receptors by Thrombin in Human Smooth Muscle Cells and the Presence of P2Y12 in Carotid Artery Lesions*. Arter. Thromb. Vasc. Biol. 2010, 30, 2434–2442. [Google Scholar] [CrossRef] [Green Version]
- Woulfe, D.; Yang, J.; Brass, L. ADP and platelets: The end of the beginning. J. Clin. Investig. 2001, 107, 1503–1505. [Google Scholar] [CrossRef]
- Bagai, A.; Wang, T.Y.; Goodman, S.G.; Fisher, H.N.; Welsh, R.C.; Dery, J.-P.; Zhang, X.; Zhu, Y.E.; Cheema, A.N.; Dehghani, P.; et al. Longitudinal treatment patterns with ADP receptor inhibitors after myocardial infarction: Insights from the Canadian Observational AntiPlatelet sTudy. Int. J. Cardiol. 2017, 228, 459–464. [Google Scholar] [CrossRef]
- Offermanns, S. Activation of Platelet Function Through G Protein–Coupled Receptors. Circ. Res. 2006, 99, 1293–1304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harizi, H.; Corcuff, J.-B.; Gualde, N. Arachidonic-acid-derived eicosanoids: Roles in biology and immunopathology. Trends Mol. Med. 2008, 14, 461–469. [Google Scholar] [CrossRef]
- Yagami, T.; Koma, H.; Yamamoto, Y. Pathophysiological Roles of Cyclooxygenases and Prostaglandins in the Central Nervous System. Mol. Neurobiol. 2016, 53, 4754–4771. [Google Scholar] [CrossRef] [PubMed]
- Marjan, M.N.; Hamzeh, M.T.; Rahman, E.; Sadeq, V. A computational prospect to aspirin side effects: Aspirin and COX-1 interaction analysis based on non-synonymous SNPs. Comput. Biol. Chem. 2014, 51, 57–62. [Google Scholar] [CrossRef]
- Zeiler, T.; Gritzka, D.; Karger, R.; Kretschmer, V. The effect of ASA on platelet activation during apheresis and on in-vitro properties of stored platelet concentrates. Transfusion 2004, 44, 1300–1305. [Google Scholar] [CrossRef] [PubMed]
- Patrono, C.; Rocca, B. The future of antiplatelet therapy in cardiovascular disease. Annu. Rev. Med. 2010, 61, 49–61. [Google Scholar] [CrossRef] [Green Version]
- Cowley, M.J.; Kuritzky, L. Developments in antiplatelet therapy for acute coronary syndromes and considerations for long-term management. Curr. Med. Res. Opin. 2009, 25, 1477–1490. [Google Scholar] [CrossRef]
- Tang, X.-F.; Fan, J.-Y.; Meng, J.; Jin, C.; Yuan, J.-Q.; Yang, Y.-J. Impact of new oral or intravenous P2Y12 inhibitors and clopidogrel on major ischemic and bleeding events in patients with coronary artery disease: A meta-analysis of randomized trials. Atherosclerosis 2014, 233, 568–578. [Google Scholar] [CrossRef]
- Horstman, L.L. Platelet microparticles: A wide-angle perspective. Crit. Rev. Oncol. 1999, 30, 111–142. [Google Scholar] [CrossRef]
- Ohlmann, P.; Lecchi, A.; El-Tayeb, A.; Müller, C.E.; Cattaneo, M.; Gachet, C. The platelet P2Y12 receptor under normal and pathological conditions. Assessment with the radiolabeled selective antagonist [3H]PSB-0413. Purinergic Signal. 2012, 9, 59–66. [Google Scholar] [CrossRef] [Green Version]
- Gachet, C. P2Y12 receptors in platelets and other hematopoietic and non-hematopoietic cells. Purinergic Signal. 2012, 8, 609–619. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, S.; Ding, Z. Role of P2Y12 Receptor in Thrombosis. Adv. Exp. Med. Biol. 2016, 906, 307–324. [Google Scholar] [CrossRef]
- Storey, R.; Sanderson, H.M.; White, A.E.; May, J.A.; Cameron, K.E.; Heptinstall, S. The central role of the P 2T receptor in amplification of human platelet activation, aggregation, secretion and procoagulant activity. Br. J. Haematol. 2000, 110, 925–934. [Google Scholar] [CrossRef] [PubMed]
- Dorsam, R.T.; Kunapuli, S.P. Central role of the P2Y12 receptor in platelet activation. J. Clin. Investig. 2004, 113, 340–345. [Google Scholar] [CrossRef] [PubMed]
- Wallentin, L. P2Y12 inhibitors: Differences in properties and mechanisms of action and potential consequences for clinical use. Eur. Heart J. 2009, 30, 1964–1977. [Google Scholar] [CrossRef] [Green Version]
- André, P.; Delaney, S.M.; LaRocca, T.; Vincent, D.; DeGuzman, F.; Jurek, M.; Koller, B.; Phillips, D.R.; Conley, P.B. P2Y12 regulates platelet adhesion/activation, thrombus growth, and thrombus stability in injured arteries. J. Clin. Investig. 2003, 112, 398–406. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.-C.; Wu, S.-Y.; Liao, C.-Y.; Teng, C.-M.; Wu, Y.-C.; Kuo, S.-C. The roles and mechanisms of PAR4 and P2Y12/phosphatidylinositol 3-kinase pathway in maintaining thrombin-induced platelet aggregation. Br. J. Pharmacol. 2010, 161, 643–658. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Foster, C.; Lecchi, A.; Quinton, T.M.; Prosser, D.M.; Jin, J.; Cattaneo, M.; Kunapuli, S.P. Protease-activated receptors 1 and 4 do not stimulate Gi signaling pathways in the absence of secreted ADP and cause human platelet aggregation independently of Gisignaling. Blood 2002, 99, 3629–3636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, T.H.; Li, J.G.; Dores, M.R.; Trejo, J. Protease-activated receptor-4 and purinergic receptor P2Y12 dimerize, co-internalize, and activate Akt signaling via endosomal recruitment of β-arrestin. J. Biol. Chem. 2017, 292, 13867–13878. [Google Scholar] [CrossRef] [Green Version]
- Khan, A.; Li, D.; Ibrahim, S.; Smyth, E.; Woulfe, D.S. The Physical Association of the P2Y12 Receptor with PAR4 Regulates Arrestin-Mediated Akt Activation. Mol. Pharmacol. 2014, 86, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Persson, R.; Lee, S.; Yood, M.U.; Wagner, M.; Minton, N.; Niemcryk, S.; Lindholm, A.; Evans, A.; Jick, S. Incident cardiovascular disease in patients diagnosed with multiple sclerosis: A multi-database study. Mult. Scler. Relat. Disord. 2020, 37, 101423. [Google Scholar] [CrossRef]
- Mincu, R.I.; Magda, L.S.; Florescu, M.; Velcea, A.; Mihaila, S.; Mihalcea, D.; Popescu, B.O.; Chiru, A.; Tiu, C.; Cinteza, M.; et al. Cardiovascular Dysfunction in Multiple Sclerosis. MAEDICA J. Clin. Med. 2015, 10, 364–370. [Google Scholar]
- Morel, A.; Bijak, M.; Miller, E.; Rywaniak, J.; Miller, S.; Saluk, J. Relationship between the Increased Haemostatic Properties of Blood Platelets and Oxidative Stress Level in Multiple Sclerosis Patients with the Secondary Progressive Stage. Oxid. Med. Cell. Longev. 2015, 2015, 240918. [Google Scholar] [CrossRef] [Green Version]
- Morel, A.; Miller, E.; Bijak, M.; Saluk, J. The increased level of COX-dependent arachidonic acid metabolism in blood platelets from secondary progressive multiple sclerosis patients. Mol. Cell. Biochem. 2016, 420, 85–94. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Wu, L.; Chen, J.; Dong, L.; Chen, C.; Wen, Z.; Hu, J.; Fleming, I.; Wang, D.W. Metabolism pathways of arachidonic acids: Mechanisms and potential therapeutic targets. Signal Transduct. Target. Ther. 2021, 6, 1–30. [Google Scholar] [CrossRef]
- Giacomazzi, A.; Degan, M.; Calabria, S.; Meneguzzi, A.; Minuz, P. Antiplatelet Agents Inhibit the Generation of Platelet-Derived Microparticles. Front. Pharmacol. 2016, 7, 314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takano, K.; Asazuma, N.; Satoh, K.; Yatomi, Y.; Ozaki, Y. Collagen-induced generation of platelet-derived microparticles in whole blood is dependent on ADP released from red blood cells and calcium ions. Platelets 2004, 15, 223–229. [Google Scholar] [CrossRef]
- Buckland, R.; Sugidachi, A.; A Jakubowski, J.; Storey, R.; Judge, H.M. Relationship between degree of P2Y12 receptor blockade and inhibition of P2Y12-mediated platelet function. Thromb. Haemost. 2010, 103, 1210–1217. [Google Scholar] [CrossRef] [PubMed]
- Behan, M.W.H.; Fox, S.C.; Heptinstall, S.; Storey, R. Inhibitory effects of P2Y12receptor antagonists on TRAP-induced platelet aggregation, procoagulant activity, microparticle formation and intracellular calcium responses in patients with acute coronary syndromes. Platelets 2005, 16, 73–80. [Google Scholar] [CrossRef]
- Simak, J.; Gelderman, M.P.; Yu, H.; Wright, V.; Baird, A.E. Circulating endothelial microparticles in acute ischemic stroke: A link to severity, lesion volume and outcome. J. Thromb. Haemost. 2006, 4, 1296–1302. [Google Scholar] [CrossRef]
- Wang, J.; Huang, Y.-J.; Wang, Y.; Xu, M.-G.; Wang, L.; Wang, S.; Tao, J. Increased Circulating CD31+/CD42− Microparticles Are Associated With Impaired Systemic Artery Elasticity in Healthy Subjects. Am. J. Hypertens. 2007, 20, 957–964. [Google Scholar] [CrossRef] [Green Version]
- Lacroix, R.; Dubois, C.; Leroyer, A.; Sabatier, F.; Dignat-George, F. Revisited role of microparticles in arterial and venous thrombosis. J. Thromb. Haemost. 2013, 11, 24–35. [Google Scholar] [CrossRef]
- Italiano, J.E.; Mairuhu, A.T.A.; Flaumenhaft, R. Clinical relevance of microparticles from platelets and megakaryocytes. Curr. Opin. Hematol. 2010, 17, 578–584. [Google Scholar] [CrossRef] [Green Version]
- Gros, A.; Ollivier, V.; Ho-Tin-Noã, B. Platelets in Inflammation: Regulation of Leukocyte Activities and Vascular Repair. Front. Immunol. 2015, 5, 678. [Google Scholar] [CrossRef]
- Manne, B.K.; Xiang, S.C.; Rondina, M.T. Platelet secretion in inflammatory and infectious diseases. Platelets 2017, 28, 155–164. [Google Scholar] [CrossRef]
- Wagner, D.D.; Burger, P.C. Platelets in Inflammation and Thrombosis. Arter. Thromb. Vasc. Biol. 2003, 23, 2131–2137. [Google Scholar] [CrossRef]
- Gawaz, M.; Langer, H.; May, A.E. Platelets in inflammation and atherogenesis. J. Clin. Investig. 2005, 115, 3378–3384. [Google Scholar] [CrossRef] [Green Version]
- Golebiewska, E.M.; Poole, A.W. Platelet secretion: From haemostasis to wound healing and beyond. Blood Rev. 2015, 29, 153–162. [Google Scholar] [CrossRef] [Green Version]
- Sharda, A.; Flaumenhaft, R. The life cycle of platelet granules. F1000Research 2018, 7, 236. [Google Scholar] [CrossRef]
- Furie, B.; Furie, B.C. Role of platelet P-selectin and microparticle PSGL-1 in thrombus formation. Trends Mol. Med. 2004, 10, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Kuenz, B.; Lutterotti, A.; Khalil, M.; Ehling, R.; Gneiss, C.; Deisenhammer, F.; Reindl, M.; Berger, T. Plasma levels of soluble adhesion molecules sPECAM-1, sP-selectin and sE-selectin are associated with relapsing-remitting disease course of multiple sclerosis. J. Neuroimmunol. 2005, 167, 143–149. [Google Scholar] [CrossRef]
- Lam, F.W.; Vijayan, K.V.; Rumbaut, R.E. Platelets and Their Interactions with Other Immune Cells. Compr. Physiol. 2015, 5, 1265–1280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dixon, D.A.; Tolley, N.D.; Bemis-Standoli, K.; Martinez, M.L.; Weyrich, A.S.; Morrow, J.D.; Prescott, S.M.; Zimmerman, G.A. Expression of COX-2 in platelet-monocyte interactions occurs via combinatorial regulation involving adhesion and cytokine signaling. J. Clin. Investig. 2006, 116, 2727–2738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jonnalagadda, D.; Izu, L.T.; Whiteheart, S. Platelet secretion is kinetically heterogeneous in an agonist-responsive manner. Blood 2012, 120, 5209–5216. [Google Scholar] [CrossRef] [PubMed]
- Leiter, O.; Walker, T.L. Platelets in Neurodegenerative Conditions—Friend or Foe? Front. Immunol. 2020, 11, 747. [Google Scholar] [CrossRef]
- Curvers, J.; De Wildt-Eggen, J.; Heeremans, J.; Scharenberg, J.; de Korte, D.; Van Der Meer, P.F. Flow cytometric measurement of CD62P (P-selectin) expression on platelets: A multicenter optimization and standardization effort. Transfusion 2008, 48, 1439–1446. [Google Scholar] [CrossRef]
- Habets, K.L.; Trouw, L.A.; Levarht, E.N.; Korporaal, S.J.; Habets, P.A.; De Groot, P.; Huizinga, T.W.; Toes, R.E. Anti-citrullinated protein antibodies contribute to platelet activation in rheumatoid arthritis. Arthritis Res. Ther. 2015, 17, 209. [Google Scholar] [CrossRef] [Green Version]
- Emoretti, R.; Epansiot, J.; Ebettati, D.; Estrazielle, N.; Ghersi-Egea, J.-F.; Edamante, G.; Efleiss, B.; Etitomanlio, L.; Egressens, P. Blood-brain barrier dysfunction in disorders of the developing brain. Front. Neurosci. 2015, 9, 40. [Google Scholar] [CrossRef] [Green Version]
- Shin, J.-H.; Kwon, H.-W.; Cho, H.-J.; Rhee, M.H.; Park, H.-J. Vasodilator-stimulated phosphoprotein-phosphorylation by ginsenoside Ro inhibits fibrinogen binding to αIIb/β3 in thrombin-induced human platelets. J. Ginseng Res. 2016, 40, 359–365. [Google Scholar] [CrossRef] [Green Version]
- Rondina, M.T.; Weyrich, A.S. Regulation of the genetic code in megakaryocytes and platelets. J. Thromb. Haemost. 2015, 13, S26–S32. [Google Scholar] [CrossRef]
- Rowley, J.W.; Schwertz, H.; Weyrich, A.S. Platelet mRNA. Curr. Opin. Hematol. 2012, 19, 385–391. [Google Scholar] [CrossRef]
- Patel, S.R.; Hartwig, J.H.; Italiano, J.E. The biogenesis of platelets from megakaryocyte proplatelets. J. Clin. Investig. 2005, 115, 3348–3354. [Google Scholar] [CrossRef] [Green Version]
- Mallouk, N.; Varvat, J.; Berger, A.; Epinat, M.; Accassat, S.; Garcin, A.; Montmartin, A.; Li, G.; Garnier, P.; Mismetti, P.; et al. Assessment of a flow cytometry technique for studying signaling pathways in platelets: Monitoring of VASP phosphorylation in clinical samples. Pr. Lab. Med. 2018, 11, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, U.R.; Geiger, J.; Walter, U.; Eigenthaler, M. Flow cytometry analysis of intracellular VASP phosphorylation for the assessment of activating and inhibitory signal transduction pathways in human platelets—Definition and detection of ticlopidine/clopidogrel effects. Thromb. Haemost. 1999, 82, 1145–1152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walkowiak, B.; Michalak, E.; Koziolkiewicz, W.; Cierniewski, C.S. Rapid photometric method for estimation of platelet count in blood plasma or platelet suspension. Thromb. Res. 1989, 56, 763–766. [Google Scholar] [CrossRef]
- Polman, C.H.; Reingold, S.C.; Banwell, B.; Clanet, M.; Cohen, J.A.; Filippi, M.; Fujihara, K.; Havrdova, E.K.; Hutchinson, M.; Kappos, L.; et al. Diagnostic criteria for multiple sclerosis: 2010 Revisions to the McDonald criteria. Ann. Neurol. 2011, 69, 292–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lublin, F.D.; Reingold, S.C. Defining the clinical course of multiple sclerosis: Results of an international survey. National Multiple Sclerosis Society (USA) Advisory Committee on Clinical Trials of New Agents in Multiple Sclerosis. Neurology 1996, 46, 907–911. [Google Scholar] [CrossRef] [PubMed]
- Kurtzke, J.F. Rating neurologic impairment in multiple sclerosis: An expanded disability status scale (EDSS). Neurology 1983, 33, 1444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beck, A.T.; Steer, R.A.; Ball, R.; Ranieri, W.F. Comparison of Beck Depression Inventories-IA and-II in Psychiatric Outpatients. J. Pers. Assess. 1996, 67, 588–597. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Agonist | Platelet Activity Markers | Spearman’s Rank Correlation | Benjamini-Hochberg FDR (Q = 5%) | Holm-Sidak (α = 0.05) | Bonferroni-Sidak (α = 0.05) | |
|---|---|---|---|---|---|---|
| Rho (95% CI) | p-Value | Adjusted p-Value 1 # | Adjusted p-Value 2 # | Adjusted p-Value 3 # | ||
| mRNA expression for the P2RY12 gene in control blood platelets vs | ||||||
| ADP | PAGs [%] | 0.286 (0.017–0.516) | 0.0328 | 0.0376 | 0.1485 | 0.5509 |
| PMPs [%] | 0.337 (0.073–0.556) | 0.0112 | 0.0202 | 0.1165 | 0.2369 | |
| P-selectin [%] | 0.355 (0.094–0.570) | 0.0073 | 0.0143 | 0.0911 | 0.1613 | |
| TRAP-6 | PAGs [%] | 0.535 (0.207–0.756) | 0.0023 | 0.0076 | 0.0450 | 0.0538 |
| PMPs [%] | 0.426 (0.067–0.688) | 0.0188 | 0.0279 | 0.1485 | 0.3659 | |
| P-selectin [%] | 0.478 (0.132–0.721) | 0.0074 | 0.0143 | 0.0911 | 0.1633 | |
| mRNA expression for the P2RY12 gene in SP MS blood platelets vs | ||||||
| ADP | PAGs [%] | 0.512 (0.249–0.705) | 0.0003 | 0.0038 | 0.0069 | 0.0072 |
| PMPs [%] | 0.441 (0.161–0.655) | 0.0024 | 0.0076 | 0.0450 | 0.0560 | |
| P-selectin [%] | 0.327 (0.028–0.572) | 0.0282 | 0.0338 | 0.1485 | 0.4967 | |
| TRAP-6 | PAGs [%] | 0.419 (0.079–0.673) | 0.0150 | 0.0252 | 0.1403 | 0.3042 |
| PMPs [%] | 0.392 (0.045–0.654) | 0.0243 | 0.0306 | 0.1485 | 0.4459 | |
| P-selectin [%] | 0.369 (0.013–0.642) | 0.0375 | 0.0411 | 0.1485 | 0.6004 | |
| mRNA expression for the P2RY12 gene in control megakaryocytes vs | ||||||
| ADP | PAGs [%] | 0.348 (0.001–0.621) | 0.0436 | 0.0458 | 0.1485 | 0.6570 |
| PMPs [%] | 0.394 (0.0536–0.652) | 0.0213 | 0.0284 | 0.1485 | 0.4035 | |
| P-selectin [%] | 0.469 (0.146–0.702) | 0.0051 | 0.0129 | 0.0738 | 0.1155 | |
| TRAP-6 | PAGs [%] | 0.533 (0.203–0.754) | 0.0024 | 0.0076 | 0.0450 | 0.0560 |
| PMPs [%] | 0.545 (0.206–0.755) | 0.0023 | 0.0076 | 0.0450 | 0.0538 | |
| P-selectin [%] | 0.540 (0.213–0.759) | 0.0021 | 0.0076 | 0.0432 | 0.0492 | |
| mRNA expression for the P2RY12 gene in SP MS megakaryocytes vs | ||||||
| ADP | PAGs [%] | 0.445 (0.075–0.707) | 0.0177 | 0.0279 | 0.1485 | 0.3486 |
| PMPs [%] | 0.577 (0.249–0.786) | 0.0013 | 0.0076 | 0.0282 | 0.0307 | |
| P-selectin [%] | 0.553 (0.216–0.773) | 0.0002 | 0.0038 | 0.0048 | 0.0048 | |
| TRAP-6 | PAGs [%] | 0.530 (0.185–0.759) | 0.0037 | 0.0104 | 0.0576 | 0.0851 |
| PMPs [%] | 0.499 (0.144–0.741) | 0.0068 | 0.0143 | 0.0911 | 0.1511 | |
| P-selectin [%] | 0.433 (0.059-0.699) | 0.0214 | 0.0284 | 0.1485 | 0.4050 | |
| Concentration of P2Y12 in control blood platelets vs | ||||||
| ADP | PAGs [%] | 0.418 (0.082–0.668) | 0.0139 | 0.0350 | 0.1894 | 0.2853 |
| PMPs [%] | 0.429 (0.097–0.676) | 0.0112 | 0.0314 | 0.1870 | 0.2369 | |
| P-selectin [%] | 0.373 (0.029–0.637) | 0.0299 | 0.0424 | 0.2354 | 0.5174 | |
| TRAP-6 | PAGs [%] | 0.379 (0.011–0.657) | 0.0385 | 0.0424 | 0.2354 | 0.6103 |
| PMPs [%] | 0.486 (0.146–0.727) | 0.0060 | 0.0314 | 0.1240 | 0.1345 | |
| P-selectin [%] | 0.513 (0.176–0.741) | 0.0038 | 0.0314 | 0.0838 | 0.0873 | |
| Concentration of P2Y12 in SP MS blood platelets vs | ||||||
| ADP | PAGs [%] | 0.312 (0.008–0.564) | 0.0387 | 0.0424 | 0.2354 | 0.6122 |
| PMPs [%] | 0.479 (0.204–0.684) | 0.0010 | 0.0252 | 0.0237 | 0.0237 | |
| P-selectin [%] | 0.317 (0.013–0.567) | 0.0361 | 0.0424 | 0.2354 | 0.5862 | |
| TRAP-6 | PAGs [%] | 0.358 (0.000–0.627) | 0.0441 | 0.0463 | 0.2354 | 0.6612 |
| PMPs [%] | 0.367 (0.016–0.634) | 0.0358 | 0.0424 | 0.2354 | 0.5831 | |
| P-selectin [%] | 0.393 (0.041–0.658) | 0.0259 | 0.0424 | 0.2354 | 0.4673 | |
| Concentration of P2Y12 in control megakaryocytes vs | ||||||
| ADP | PAGs [%] | 0.496 (0.139–0.739) | 0.0073 | 0.0314 | 0.1426 | 0.1613 |
| PMPs [%] | 0.425 (0.050–0.695) | 0.0241 | 0.0424 | 0.2354 | 0.4432 | |
| P-selectin [%] | 0.475 (0.112–0.726) | 0.0107 | 0.0314 | 0.1870 | 0.2275 | |
| TRAP-6 | PAGs [%] | 0.406 (0.027–0.683) | 0.0320 | 0.0424 | 0.2354 | 0.5418 |
| PMPs [%] | 0.438 (0.066–0.703) | 0.0197 | 0.0384 | 0.2333 | 0.3797 | |
| P-selectin [%] | 0.439 (0.066–0.703) | 0.0198 | 0.0384 | 0.2333 | 0.3812 | |
| Concentration of P2Y12 in SP MS megakaryocytes vs | ||||||
| ADP | PAGs [%] | 0.476 (0.115–0.727) | 0.0103 | 0.0314 | 0.1870 | 0.2200 |
| PMPs [%] | 0.472 (0.108–0.724) | 0.0112 | 0.0314 | 0.1870 | 0.2369 | |
| P-selectin [%] | 0.473 (0.110–0.725) | 0.0110 | 0.0314 | 0.1870 | 0.2331 | |
| TRAP-6 | PAGs [%] | 0.413 (0.036–0.687) | 0.0288 | 0.0424 | 0.2354 | 0.5041 |
| PMPs [%] | 0.400 (0.021–0.679) | 0.0349 | 0.0424 | 0.2354 | 0.5737 | |
| P-selectin [%] | 0.441 (0.069–0.705) | 0.0188 | 0.0384 | 0.2333 | 0.3659 | |
| Clinical Characteristics | Reference Range | Control Group (n = 55) | SP MS (n = 55) |
|---|---|---|---|
| Age (years) | - | 48.7 ± 12.2 | 48.6 ± 12.5 |
| Female (%) | - | 58 | 60 |
| BMI (kg/m2) | 18.5–24.9 | 22.31 ± 5.12 | 21.1 ± 9.7 |
| EDSS (0–10) | 0–10 | NA | 5.5 ± 1.8 |
| BDI (≤9) | 9–47 | NA | 9.6 ± 4.6 |
| Mean disease duration (years) | - | NA | 12.3 ± 9.5 |
| TG (mg/dL) | 35–150 | 131.45 ± 79.57 | 73 ± 61.27 |
| TChol (mg/dL) | 115–190 | 189 ± 44.20 | 201 ± 399.1 |
| HDL (mg/dL) | >45 | 60.87 ± 16.71 | 72 ± 20.4 |
| LDL (mg/dL) | <70 | 110.80 ± 38.73 | 90 ± 41.5 |
| LEU (×103/μL) | 4–10 | 6.46 ± 1.53 | 5.15 ± 2.4 |
| RBC (×106/μL) | 3.80–5.40 | 4.81 ± 0.39 | 4.41 ± 0.7 |
| PLT (×103/μL) | 150–400 | 258.89 ± 68.10 | 321.95 ± 42.99 |
| GLU (mg/dL) | 70–99 | 96.69 ± 26.07 | 84.1 ± 12 |
| CR (mg/dL) | 0.50–0.90 | 85.22 ± 16.80 | 72.4 ± 13.4 |
| TSH (mIU/mL) | 0.27–4.20 | 1.71 ± 0.80 | 1.6 ± 0.9 |
| AST (IU/L) | <32 | 21.09 ± 16.09 | 26.7 ± 14.07 |
| ALT (U/L) | <33 | 21.15 ± 13.06 | 28 ± 12.4 |
| PT (sec) | 12–16 | 12.28 ± 2.29 | 13.5 ± 2.01 |
| INR (sec) | 0.91–1.31 | 1.01 ± 0.05 | 1.5 ± 0.04 |
| APTT (sec) | 26–40 | 29.97 ± 2.30 | 31.2 ± 1.45 |
| FG (mg/dL) | 150–450 | 316.94 ± 58.92 | 320.25 ± 45.87 |
| ESR (mm/h) | 3–12 | 9.35 ± 8.90 | 8.45 ± 1.54 |
| CRP (mg/L) | <5 | 3.15 ± 0.99 | 12.5 ± 5.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dziedzic, A.; Miller, E.; Saluk-Bijak, J.; Niwald, M.; Bijak, M. The Molecular Aspects of Disturbed Platelet Activation through ADP/P2Y12 Pathway in Multiple Sclerosis. Int. J. Mol. Sci. 2021, 22, 6572. https://doi.org/10.3390/ijms22126572
Dziedzic A, Miller E, Saluk-Bijak J, Niwald M, Bijak M. The Molecular Aspects of Disturbed Platelet Activation through ADP/P2Y12 Pathway in Multiple Sclerosis. International Journal of Molecular Sciences. 2021; 22(12):6572. https://doi.org/10.3390/ijms22126572
Chicago/Turabian StyleDziedzic, Angela, Elzbieta Miller, Joanna Saluk-Bijak, Marta Niwald, and Michal Bijak. 2021. "The Molecular Aspects of Disturbed Platelet Activation through ADP/P2Y12 Pathway in Multiple Sclerosis" International Journal of Molecular Sciences 22, no. 12: 6572. https://doi.org/10.3390/ijms22126572
APA StyleDziedzic, A., Miller, E., Saluk-Bijak, J., Niwald, M., & Bijak, M. (2021). The Molecular Aspects of Disturbed Platelet Activation through ADP/P2Y12 Pathway in Multiple Sclerosis. International Journal of Molecular Sciences, 22(12), 6572. https://doi.org/10.3390/ijms22126572