
Dietary Habits and Gut Microbiota in Healthy Adults: Focusing on the Right Diet. A Systematic Review

 , ,
, ,  ,
, 
Abstract
:1. Introduction
2. Results
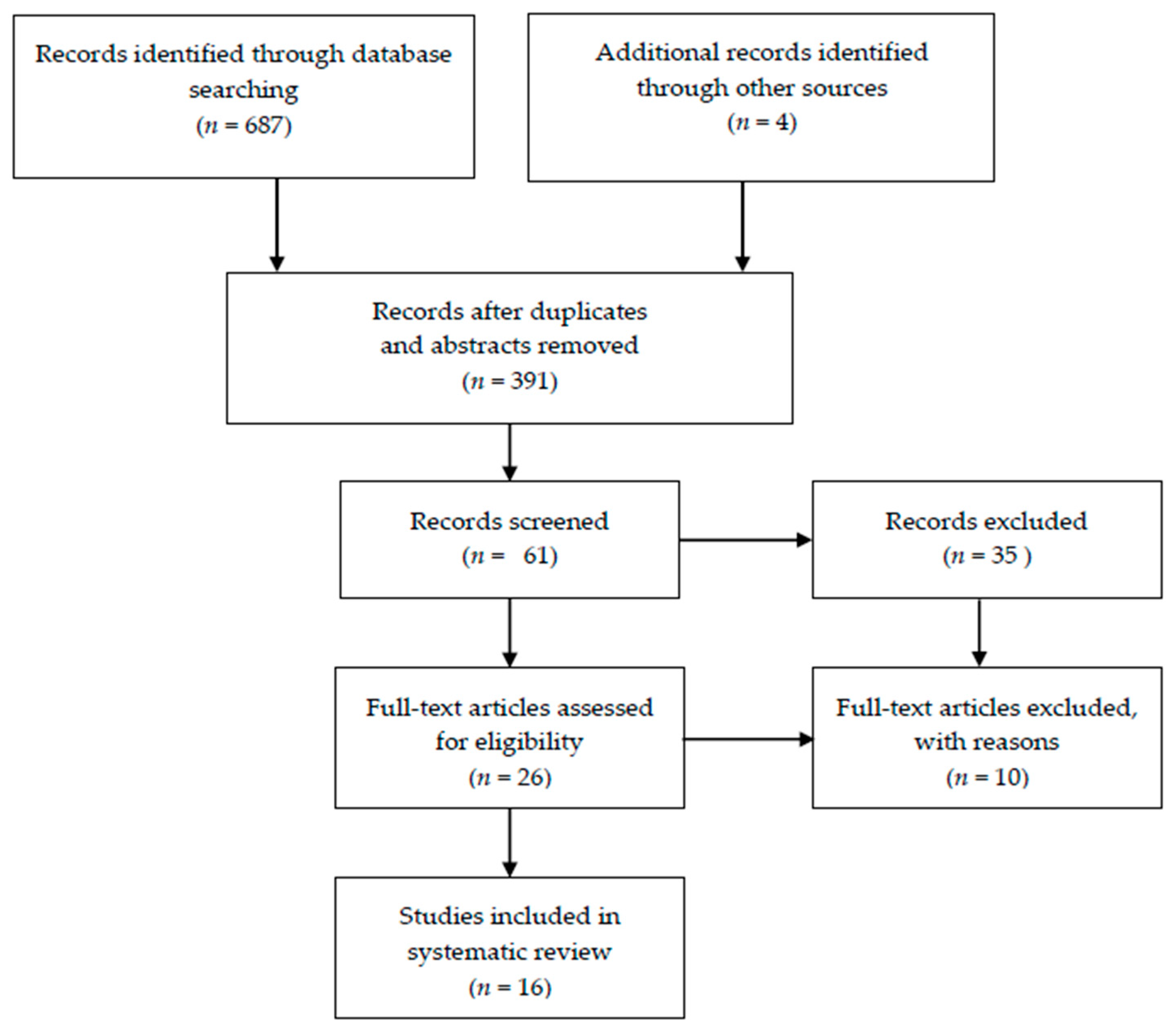
2.1. Study Characteristics
2.2. Microbiota Composition in Omnivore Population
2.3. Short-Chain Fatty Acids Production
3. Discussion
4. Materials and Methods
4.1. Search Strategy
4.2. Eligibility Criteria
4.3. Study Selection and Data Extraction
4.4. Quality Assessment of Studies
4.5. Microbiota Data Reports
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lynch, S.V.; Pedersen, O. The human intestinal microbiome in health and disease. N. Engl. J. Med. 2016, 375, 2369–2379. [Google Scholar] [CrossRef] [Green Version]
- Nieuwdorp, M.; Gilijamse, P.W.; Pai, N.; Kaplan, L.M. Role of the microbiome in energy regulation and metabolism. Gastroenterology 2014, 146, 1525–1533. [Google Scholar] [CrossRef]
- The Human Microbiome Project Consortium. Structure, function and diversity of the healthy human microbiome. Nature 2012, 486, 207–214. [Google Scholar] [CrossRef] [Green Version]
- Montalto, M.; D’onofrio, F.; Gallo, A.; Cazzato, A.; Gasbarrini, G. Intestinal microbiota and its functions. Dig. Liver Dis. Suppl. 2009, 3, 30–34. [Google Scholar] [CrossRef]
- Eckburg, P.B.; Bik, E.M.; Bernstein, C.N.; Purdom, E.; Dethlefsen, L.; Sargent, M. Diversity of the human intestinal microbial flora. Science 2005, 308, 1635–1638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, R.; Makino, H.; Cetinyurek, Y.A.; Ben-Amor, K.; Roelofs, M.; Ishikawa, E. Early-life events, including mode of delivery and type of feeding, siblings and gender, shape the developing gut microbiota. PLoS ONE 2016, 11, e0158498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rinninella, E.; Cintoni, M.; Raoul, P.; Lopetuso, L.R.; Scaldaferri, F.; Pulcini, G.; Miggiano, G.A.D.; Gasbarrini, A.; Mele, M.C. Food components and dietary habits: Keys for a healthy gut micro-biota composition. Nutrients 2019, 11, 2393. [Google Scholar] [CrossRef] [Green Version]
- Zeppa, D.; Agostini, D.; Gervasi, M.; Annibalini, G.; Amatori, S.; Ferrini, F.; Sisti, D.; Piccoli, G.; Barbieri, E.; Sestili, P.; et al. Mutual interactions among exercise, sport supplements and microbiota. Nutrients 2019, 12, 17. [Google Scholar] [CrossRef] [Green Version]
- Dorelli, B.; Gallè, F.; Vito, C.D.; Duranti, G.; Iachini, M.; Zaccarin, M.; Standoli, J.P.; Ceci, R.; Romano, F.; Liguori, G.; et al. Can physical activity influence human gut microbiota composition independently of diet? A systematic review. Nutrients 2021, 13, 1890. [Google Scholar] [CrossRef]
- Aya, V.; Flórez, A.; Perez, L.; Ramírez, J.D. Association between physical activity and changes in intestinal microbiota compo-sition: A systematic review. PLoS ONE 2021, 16, e0247039. [Google Scholar] [CrossRef]
- Bian, X.; Chi, L.; Gao, B.; Tu, P.; Ru, H.; Lu, K. Gut microbiome response to sucralose and its potential role in inducing liver inflammation in mice. Front. Physiol. 2017, 8, 487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chassaing, B.; Koren, O.; Goodrich, J.K.; Poole, A.C.; Srinivasan, S.; Ley, R.E. Dietary emulsifiers impact the mouse gut microbiota promoting colitis and metabolic syndrome. Nature 2015, 519, 92–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nettleton, J.E.; Reimer, R.A.; Shearer, J. Reshaping the gut microbiota: Impact of low calorie sweeteners and the link to insulin resistance? Physiol. Behav. 2016, 164, 488–493. [Google Scholar] [CrossRef] [PubMed]
- Valdes, A.M.; Walter, J.; Segal, E.; Spector, T.D. Role of the gut microbiota in nutrition and health. BMJ 2018, 361, k2179. [Google Scholar] [CrossRef] [Green Version]
- Widmer, R.J.; Flammer, A.J.; Lerman, L.O.; Lerman, A. The Mediterranean diet, its components, and cardiovascular disease. Am. J. Med. 2015, 128, 229–238. [Google Scholar] [CrossRef] [Green Version]
- Chambers, E.S.; Byrne, C.S.; Morrison, D.J.; Murphy, K.G.; Preston, T.; Tedford, C.; Garcia-Perez, I.; Fountana, S.; Serrano-Contreras, J.I.; Holmes, E.; et al. Dietary supplementation with inulin-propionate ester or inulin improves insulin sensitivity in adults with overweight and obesity with distinct effects on the gut microbiota, plasma metabolome and systemic inflammatory responses: A randomised cross-over trial. Gut 2019, 68, 1430–1438. [Google Scholar] [CrossRef]
- Costantini, L.; Molinari, R.; Farinon, B.; Merendino, N. Impact of omega-3 fatty acids on the gut microbiota. Int. J. Mol. Sci. 2017, 18, 2645. [Google Scholar] [CrossRef] [Green Version]
- Gentile, C.L.; Weir, T.L. The gut microbiota at the intersection of diet and human health. Science 2018, 362, 776–780. [Google Scholar] [CrossRef] [Green Version]
- Stefani, S.; Ngatidjan, S.; Paotiana, M.; Sitompul, K.A.; Abdullah, M.; Sulistianingsih, D.P.; Shankar, A.H.; Agustina, R. Dietary quality of predominantly traditional diets is associated with blood glucose profiles, but not with total fecal Bifidobacterium in Indonesian women. PLoS ONE 2018, 13, e0208815. [Google Scholar] [CrossRef]
- Ferrocino, I.; Cagno, R.D.; Angelis, M.D.; Turroni, S.; Vannini, L.; Bancalari, E.; Rantsiou, K.; Cardinali, G.; Neviani, E.; Cocolin, L. Fecal microbiota in healthy subjects following omnivore, vegetarian and vegan diets: Culturable populations and rRNA DGGE profiling. PLoS ONE 2015, 10, e0128669. [Google Scholar] [CrossRef] [Green Version]
- Filippis, F.D.; Pellegrini, N.; Vannini, L.; Jeffery, I.B.; Storia, A.L.; Laghi, L.; Serrazanetti, D.I.; Cagno, R.D.; Ferrocino, I.; Lazzi, C.; et al. High-level adherence to a Mediterranean diet beneficially impacts the gut microbiota and associated metabolome. Gut 2016, 65, 1812–1821. [Google Scholar] [CrossRef] [PubMed]
- Federici, E.; Prete, R.; Lazzi, C.; Pellegrini, N.; Moretti, M.; Corsetti, A.; Cenci, G. Bacterial composition, genotoxicity, and cytotoxicity of fecal samples from individuals consuming omnivorous or vegetarian diets. Front. Microbiol. 2017, 8, 300. [Google Scholar] [CrossRef] [Green Version]
- Losasso, C.; Eckert, E.M.; Mastrorilli, E.; Villiger, J.; Mancin, M.; Patuzzi, I.; Cesare, A.D.; Cibin, V.; Barrucci, F.; Pernthaler, J.; et al. Assessing the influence of vegan, vegetarian and omnivore oriented westernized dietary styles on human gut microbiota: A cross sectional study. Front. Microbiol. 2018, 9, 317. [Google Scholar] [CrossRef]
- Luisi, M.L.E.; Lucarini, L.; Biffi, B.; Rafanelli, E.; Pietramellara, G.; Durante, M.; Vidali, S.; Provensi, G.; Madiai, S.; Gheri, C.F.; et al. Effect of Mediterranean diet enriched in high quality extra virgin olive oil on oxidative stress. Inflammation and gut microbiota in obese and normal weight adult subjects. Front. Pharmacol. 2019, 10, 1366. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, T.S.; Rampelli, S.; Jeffery, I.B.; Santoro, A.; Neto, M.; Capri, M.; Giampieri, E.; Jennings, A.; Candela, M.; Turroni, S.; et al. Mediterranean diet intervention alters the gut microbiome in older people reducing frailty and improving health status: The NU-AGE 1-year dietary intervention across five European countries. Gut 2020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sofi, F.; Dinu, M.; Pagliai, G.; Cesari, F.; Marcucci, R.; Casini, A. Mediterranean versus vegetarian diet for cardiovascular disease prevention (the CARDIVEG study): Study protocol for a randomized controlled trial. Trials 2016, 17, 233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trefflich, I.; Jabakhanji, A.; Menzel, J.; Blaut, M.; Michalsen, A.; Lampen, A.; Abraham, K.; Weikert. Is a vegan or a vegetarian diet associated with the microbiota composition in the gut? Results of a new cross-sectional study and systematic review. Crit. Rev. Food Sci. Nutr. 2019, 21, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Díaz, I.; Fernández-Navarro, T.; Sánchez, B.; Margolles, A.; González, S. Mediterranean diet and faecal microbiota: A transversal study. Food Funct. 2016, 7, 2347–2356. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Díaz, I.; Fernández-Navarro, T.; Salazar, N.; Bartolomé, B.; Moreno-Arribas, M.V.; Andres-Galiana, E.J.D.; Fernández-Martínez, J.L.; Reyes-Gavilán, C.G.D.L.; Gueimonde, M.; González, S. Adherence to a Mediterranean diet influences the fecal metabolic profile of microbial-derived phenolics in a Spanish cohort of middle-age and older people. J. Agric. Food Chem. 2017, 65, 586–595. [Google Scholar] [CrossRef]
- Franco-de-Moraes, A.C.; Almeida-Pititto, B.D.; Fernandes, G.D.R.; Gomes, E.P.; Pereira, A.D.C.; Ferreira, S.R.G. Worse inflammatory profile in omnivores than in vegetarians associates with the gut microbiota composition. Diabetol. Metab. Syndr. 2017, 9, 62. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.D.; Compher, C.; Chen, E.Z.; Smith, S.A.; Shah, R.D.; Bittinger, K.; Chehoud, C.; Albenberg, L.G.; Nessel, L.; Gilroy, E.; et al. Comparative metabolomics in vegans and omnivores reveal constraints on diet-dependent gut microbiota metabolite production. Gut 2016, 65, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Wan, Y.; Yin, K.; Wei, Y.; Wang, B.; Yu, X.; Ni, Y.; Zheng, J.; Huang, T.; Song, M.; et al. Lower circulating branched-chain amino acid concentrations among vegetarians are associated with changes in gut microbial composition and function. Mol. Nutr. Food Res. 2019, 63, e1900612. [Google Scholar] [CrossRef] [PubMed]
- Wan, Y.; Wang, F.; Yuan, J.; Li, J.; Jiang, D.; Zhang, J.; Li, H.; Wang, R.; Tang, J.; Huang, T.; et al. Effects of dietary fat on gut microbiota and faecal metabolites, and their relationship with cardiometabolic risk factors: A 6-month randomised controlled-feeding trial. Gut 2019, 68, 1417–1429. [Google Scholar] [CrossRef] [Green Version]
- Higgins, J.P.T.; Green, S.; Altman, D.G. Assessing Risk of Bias in Included Studies Published Online. In Cochrane Handbook for Systematic Reviews of Interventions: Cochrane Book Series; Wiley: Hoboken, NJ, USA, 2008; Chapter 8. [Google Scholar]
- Mitsou, E.K.; Kakali, A.; Antonopoulou, S.; Mountzouris, K.C.; Yannakoulia, M.; Panagiotakos, D.B.; Kyriacou, A. Adherence to the Mediterranean diet is associated with the gut microbiota pattern and gastrointestinal characteristics in an adult population. Br. J. Nutr. 2017, 117, 1645–1655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pagliai, G.; Russo, E.; Niccolai, E.; Dinu, M.; Pilato, V.D.; Magrini, A.; Bartolucci, G.; Baldi, S.; Menicatti, M.; Giusti, B.; et al. Influence of a 3-month low-calorie Mediterranean diet compared to the vegetarian diet on human gut microbiota and SCFA: The CARDIVEG Study. Eur. J. Nutr. 2019. [Google Scholar] [CrossRef]
- Santoro, A.; Pini, E.; Scurti, M. Combating inflammaging through a Mediterranean whole diet approach: The NU-AGE project’s conceptual framework and design. Mech. Ageing Dev. 2014, 136, 3–13. [Google Scholar] [CrossRef]
- Wan, Y.; Wang, F.; Yuan, J. Effects of macronutrient distribution on weight and related cardiometabolic profile in healthy non-obese chinese: A 6-month, randomized controlled-feeding trial. EBioMedicine 2017, 22, 200–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palleja, A.; Mikkelsen, K.H.; Forslund, S.H. Recovery of gut microbiota of healthy adults following antibiotic exposure. Nat. Microbiol. 2018, 3, 1255–1265. [Google Scholar] [CrossRef]
- Shanahan, F.; Van Sinderen, D.; O’Toole, P.W.; Stanton, C. Feeding the microbiota: Transducer of nutrient signals for the host. Gut 2017, 66, 1709–1717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ercolini, D.; Fogliano, V. Food design to feed the human gut microbiota. J. Agric. Food Chem. 2018, 66, 3754–3758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerasimidis, K.; Bryden, K.; Chen, X.; Papachristou, E.; Verney, A.; Roig, M.; Hansen, R.; Nichols, B.; Papadopoulou, R.; Parrett, A. The impact of food additives, artificial sweeteners and domestic hygiene products on the human gut microbiome and its fibre fermentation capacity. Eur. J. Nutr. 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roca-Saavedra, P.; Mendez-Vilabrille, V.; Miranda, J.M.; Nebot, C.; Cardelle-Cobas, A.; Franco, C.M.; Cepeda, A. Food additives, contaminants and other minor components: Effects on human gut microbiota—A review. J. Physiol. Biochem. 2017, 74, 69–83. [Google Scholar] [CrossRef]
- Suez, J.; Korem, T.; Zeevi, D.; Zilberman-Schapira, G.; Thaiss, C.A.; Maza, O.; Israeli, D.; Zmora, N.; Gilad, S.; Weinberger, A.; et al. Artificial sweeteners induce glucose intolerance by altering the gut microbiota. Nature 2014, 514, 181–186. [Google Scholar] [CrossRef]
- Halmos, E.P.; Mack, A.; Gibson, P.R. Review article: Emulsifiers in the food supply and implications for gastrointestinal disease. Aliment. Pharmacol. Ther. 2019, 49, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Trichopoulou, A.; Martínez-González, M.A.; Tong, T.Y.; Forouhi, N.G.; Khandelwal, S.; Prabhakaran, D.; Mozaffarian, D.; Lorgeril, M.D. Definitions and potential health benefits of the Mediterranean diet: Views from experts around the world. BMC Med. 2014, 12, 112. [Google Scholar] [CrossRef] [Green Version]
- Schuppler, M.; Mertens, F.; Schön, G.; Göbelet, U.B. Molecular characterization of nocardioform actinomycetes in activated sludge by 16S rRNA analysis. Microbiology 1995, 141, 513–521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blaut, M.; Collins, M.D.; Welling, G.W.; Doré, J.; Van Loo, J.; Voset, W.D. Molecular biological methods for studying the gut microbiota: The EU human gut flora project. Br. J. Nutr. 2002, 87, S203–S211. [Google Scholar] [CrossRef] [Green Version]
- Arnold, J.W.; Roach, J.; Azcarate-Peril, M.A. Emerging technologies for gut microbiome research. Trends Microbiol. 2016, 24, 887–901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allaband, C.; McDonald, D.; Vázquez-Baeza, Y.; Minich, J.J.; Tripathi, A.; Brenner, D.A.; Loomba, R.; Smarr, L.; Sandborn, L.J.; Schnabl, B.; et al. Microbiome 101: Studying, analyzing, and interpreting gut microbiome data for clinicians. Clin. Gastroenterol. Hepatol. 2019, 17, 218–230. [Google Scholar] [CrossRef] [PubMed]
- Marietta, E.; Rishi, A.; Taneja, V. Immunogenetic control of the intestinal microbiota. Immunology 2015, 145, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Christ, A.; Lauterbach, M.; Latz, E. Western diet and the immune system: An inflammatory connection. Immunity 2019, 51, 794–811. [Google Scholar] [CrossRef]
- Zheng, D.; Liwinski, T.; Elinav, E. Interaction between microbiota and immunity in health and disease. Cell Res. 2020, 30, 492–506. [Google Scholar] [CrossRef] [PubMed]
- Devkota, S.; Wang, Y.; Musch, M.W.; Leone, V.; Fehlner-Peach, H.; Nadimpalli, A.; Antonopoulos, D.A.; Jabri, B.; Chang, E.B. Dietary-fat-induced taurocholic acid promotes pathobiont expansion and colitis in Il10−/− mice. Nature 2012, 487, 104–108. [Google Scholar] [CrossRef] [Green Version]
- Cheng, L.; Jin, H.; Qiang, Y.; Wu, S.; Yan, C.; Han, M.; Xiao, T.; Yan, N.; An, H.; Zhou, X.; et al. High fat diet exacerbates dextran sulfate sodium induced colitis through disturbing mucosal dendritic cell homeostasis. Int. Immunopharmacol. 2016, 40, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Moher, D.; Altman, D.G.; Liberati, A.; Tetzlaff, J. PRISMA statement. Epidemiology 2011, 22, 128. [Google Scholar] [CrossRef] [Green Version]
- Wells, G.A.; Shea, B.; O’Connell, D.; Peterson, J.; Welch, V.; Losos, M.; Tugwell, P. The Newcastle-Ottawa Scale (NOS) for Assessing the Quality of Nonrandomised Studies in Meta-Analyses; Ottawa Hospital Research Institute: Ottawa, ON, Canada, 2001. [Google Scholar]
- Ding, N.; Zhang, X.; Zhang, X.D.; Jing, J.; Liu, S.S.; Mu, Y.P.; Peng, L.L.; Yan, Y.J.; Xiao, G.M.; Bi, X.Y.; et al. Impairment of spermatogenesis and sperm motility by the high-fat diet-induced dysbiosis of gut microbes. Gut 2020. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Author, Year, Reference | Study Design | Study Population (Males, Mean or Median Age) | BMI (kg/m2) Mean or Median Value | Diet Intervention | Region | Dietary Assessment | Microbiota Assessment and Metabolomic Analysis | Outcome 1 Significant Difference in Microbiota Composition | Outcome 2 Short-Chain Fatty Acid (SCFAs) Production Variation |
|---|---|---|---|---|---|---|---|---|---|
| Ferrocino I, 2015 [20] | Cross- sectional | 153, M 76, 18–55 ys | >18 | 51 vegetarians, 51 vegans and 51 omnivores ° >12 months * | Italy | Self-reported | Real-time quantitative PCR and rRNA-DGGE | Phylum: Bacteroidetes Genus: Bacteroides Species: B. fragilis | NR |
| De Filippis F, 2016 [21] | Cross- sectional | 153 M 64, 18–55 ys | >18 | 51 vegetarians, 51 vegans and 51 omnivores ° >12 months ¹,* | Italy | Dietary score based on tertiles. The Healthy Food Diversity index (HFD) | 16S rRNA sequencing + metabolomic analysis using gas-chromatography mass spectrometry-solid-phase microextraction | Phylum: Firmicutes Genus: Ruminococcus Genus: Streptococcus | Faecal SCFA profile increasing with MD and plant-based diets. (Prevotella was the only Bacteroidetes having positive correlations with SCFA |
| Wu GD, 2016 [31] | Cross- sectional | 31, NR | NR | 15 vegans and 16 omnivores for ≥6 months | USA | Food frequency questionnaire (FFQ) | 16S rRNA sequencing +metabolomic analysis of SCFAs faces with nuclear magnetic resonance (NMR) spectra of faecal water | No significant difference at genus level | No significant effect on the levels of faecal SCFAs despite plant-based diet increase |
| Gùtierrez-Diàz I, 2016 [28] | Cross- sectional | 31, M 8, 42.1 ± 10.9 ys | NR | 31 subjects following Med Diet ° ≥ 6 months | Spain | Mediterranean Diet Score | 16S rRNA sequencing + metabolomic analysis faceal SCFAs detected by gas chromatographymass spectrometry (MS) | -Phylum: Bacteroidetes, Genus: Prevotella -Lower levels of Phylum Firmicutes and Genus Ruminococcus | Higher concentration of faecal propionate and butyrate |
| Gùtierrez-Diàz I, 2017 [29] | Cross- sectional | 74, M 20, 50 ≥ 65 ys | 25–30 | 74 subjects following Mediterranean diet ° | Spain | Med Diet Score | 16S rRNA sequencing and UPLC-ESI-MS/MS method for phenolic metabolytes analysis | -Phylum: Firmicutes Genus: Clostridium Species: Cl. XVIa -Phylum: Firmicutes Genus: Faecalibacterium Species: F. prausnitzii | NR |
| Federici E, 2017 [22] | Cross- sectional | 29, M 14, 39 ± 10 ys, 33 ± 7 ys and 41 ± 9 ys, respectively | 20.7 ± 2.2, 22.3 ± 2.2 and 22.6 ± 1.7, respectively | 12 vegetarians, 10 vegans and 7 omnivores for >12 months | Italy | 7-day weighed food diary | Faecal microbial counts | -Phylum: Actinobacteria Genus: Corynebacteria -Phylum: Firmicutes Genus Staphylococcus | NR |
| Franco-de-Moraes AC, 2017 [30] | Cross- sectional | 268, M 123, respectively 49.6 ± 8.5 ys, 49.6 ± 8.6 ys and 49.1 ± 8.2 ys | <40 | 66 strict vegetarians, 102 lacto-ovo-vegetarians, and 100 omnivores for the last 12 months | Brasil | NR | 16s RNA gene analysis | -Phylum: Protecobacteria Genus: Succinivibrio - Phylum: Protecobacteria Genus: Halomonas | NR |
| Mitso E.K, 2017 [35] | Cross- sectional | 100, M 48, 41.27 ± 13.33 ys | 27.29 ± 4.48 | 100 following Mediterranean diet ° (3 tertiles of adherence: low tertile, medium, tertile and high tertile) | Greece | Food Frequency Questionnaire (FFQ) and MedDiet Score | 16s rRNA sequencing and metabolomic analysis were performed with capillary gas chromatography for faecal SCFAs | Phylum: Bacteroidetes Genus: Bacteroides + Increase of C. albicans | Med Diet positively linked to total SCFA |
| Losasso C, 2018 [23] | Cross- sectional | 101, M 33, 42.5 ± 13.0 ys | 23.8 ± 4.4 | Vegans 26, vegetarians 32, and omnivores 43 for >12 months | Italy | Food frequency questionnaire (FFQ) and 24 h dietary recall | 16s rRNA sequencing | No difference for bacterial community composition | NR |
| Stefani, S 2018 [19] | Cross- sectional | 240 healthy women, 38.0 (31.0–44.0) ys | 24.9 ± 49.5 | Two groups of 120 women of West Sumatera and West Java provinces following predominantly animal- or plant-based traditional diets | Indonesia | 2-day-repeated 24-h food recalls | DNA extraction and quantification of Bifidobacterium DNA using the Bifidobacterium sp. standard primer and using Real-Time PCR System | No significant alteration of genus Bifidobacterium | NR |
| Pagliai G, 2019 [26,36] | Randomized cross-over | 23, M 7, 58.6 ± 9.8 ys | 31.06 ± 0.67 and 30.10 ± 0.61 for the two groups | 23 omnivorous ° entrolled: 11 following low-calorie Mediterran Diet and 12 Vegan Diet for three months and then crossed | Italy | Dietary randomization | 16s rRNA sequencing and gas chromatography–mass spectrometry system for SCFAs | Phylum: Bacteroidetes Genus: Enterorhabdus, Lachnoclostridium, and Parabacteroides | Mean variation of each SCFA Increase of 10% of propionic acid |
| Trefflich I, 2019 [27] | Cross- sectional | 72, 36 M, 37.5 (32.5–44.0) and 38.5 (32.0–46.0) ys, respectively | 22.9 (± 3.2) and 24.0 (±2.1) | 36 vegan and 36 omnivorous participants following diet for >12 months | Germany | NR | 16S rRNA (rRNA) gene sequencing | Modest differences Not significant between vegans and omnivores at phylum, family, genus, and species level. | NR |
| Wang F, 2019 [32] | Cross- sectional | 36, 28.1 ys | NR | 36 adults following a vegan (12), a lacto-ovo vegetarian (12), or an omnivorous diet (12) for > 6 months | China | Nutrition System of Traditional Chinese Medicine Combining with Western Medicine, version 11.0 | 16s rRNA gene analysis | Phylum: Bacteroidetes Genus: Bacteroides | NR |
| Luisi MLE, 2019 [24] | Cross- sectional | 36, M 17, 41.4 ± 14.42 and 52.1 ± 13.04 years, respectively for cases and controls | Cases ≥25 Controls 18.5 and 24.9 | 36 following typical MD ° and cases receiving a low-calorie MD for 3 months; both cases and controls utilized 40 g/die of HQ-EVOO as the only cooking and dressing fat | Italy | NR | dsDNA extracted from all the samples | No significant modification measurable | NR |
| Wan Y, 2019 [33,38] | Observer- blinded, RCT | 217, M 114, Respectively, 23.3 (3.4), 23.6 (4.0) ys and 23.4 (4.1) ys | 21.7 (2.6) | Lower-fat diet (73), moderate-fat diet (73) and higher-fat diet (71) for 6 months | China | Daily diary | 16s RRNA sequencing and mass spectrometry system for SCFAs | -Lower-fat diet associated with increased Phyla: Firmicutes Genus: Blautia and Faecalibacterium -Higher-fat associated with increased Genus: Alistipes and Bacteroides (phylum: Bacteroidetes). | Total SCFAs significantly decreased in the higher-fat diet group in comparison with the other groups. |
| Ghosh TS, 2020 [25,37] | Randomized, multicentre, single-blind, controlled trial | 612, M 286, Median age 65-75 ys | Median value 26.8 (18.8- 44.6) and 26.9 (18.5–46) | 289 controls and 323 in MedDiet ° for 12 months | UK, France, Netherlands, Italy, and Poland | Adherence scores to the MedDiet, based on the NU-AGE Food Based Dietary Guidelines (FBDG) | DNA and 16S rRNA gene sequencing | “Diet Positive OTUs”: Phylum: Firmicutes Genus: Faecalibacterium Species: faecalibacterium prausnitzii -Phylum: Firmicutes Genus: Roseburia Species: Roseburia hominis -Phylum: Firmicutes Genus: Eubacterium -Phylum: Bacteroidetes Genus: Bacteroides Species: Bacteroides thetaiotaomicron -Phylum: Bacteroidetes Genus: Prevotella Species: Prevotella copri -Phylum: Firmicutes Genus: Anaerostipes Species: Anaerostipes hadrus | NR |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gibiino, G.; De Siena, M.; Sbrancia, M.; Binda, C.; Sambri, V.; Gasbarrini, A.; Fabbri, C. Dietary Habits and Gut Microbiota in Healthy Adults: Focusing on the Right Diet. A Systematic Review. Int. J. Mol. Sci. 2021, 22, 6728. https://doi.org/10.3390/ijms22136728
Gibiino G, De Siena M, Sbrancia M, Binda C, Sambri V, Gasbarrini A, Fabbri C. Dietary Habits and Gut Microbiota in Healthy Adults: Focusing on the Right Diet. A Systematic Review. International Journal of Molecular Sciences. 2021; 22(13):6728. https://doi.org/10.3390/ijms22136728
Chicago/Turabian StyleGibiino, Giulia, Martina De Siena, Monica Sbrancia, Cecilia Binda, Vittorio Sambri, Antonio Gasbarrini, and Carlo Fabbri. 2021. "Dietary Habits and Gut Microbiota in Healthy Adults: Focusing on the Right Diet. A Systematic Review" International Journal of Molecular Sciences 22, no. 13: 6728. https://doi.org/10.3390/ijms22136728
APA StyleGibiino, G., De Siena, M., Sbrancia, M., Binda, C., Sambri, V., Gasbarrini, A., & Fabbri, C. (2021). Dietary Habits and Gut Microbiota in Healthy Adults: Focusing on the Right Diet. A Systematic Review. International Journal of Molecular Sciences, 22(13), 6728. https://doi.org/10.3390/ijms22136728