
Effects of Trichoderma harzianum on Photosynthetic Characteristics and Fruit Quality of Tomato Plants

 , , ,
, , ,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
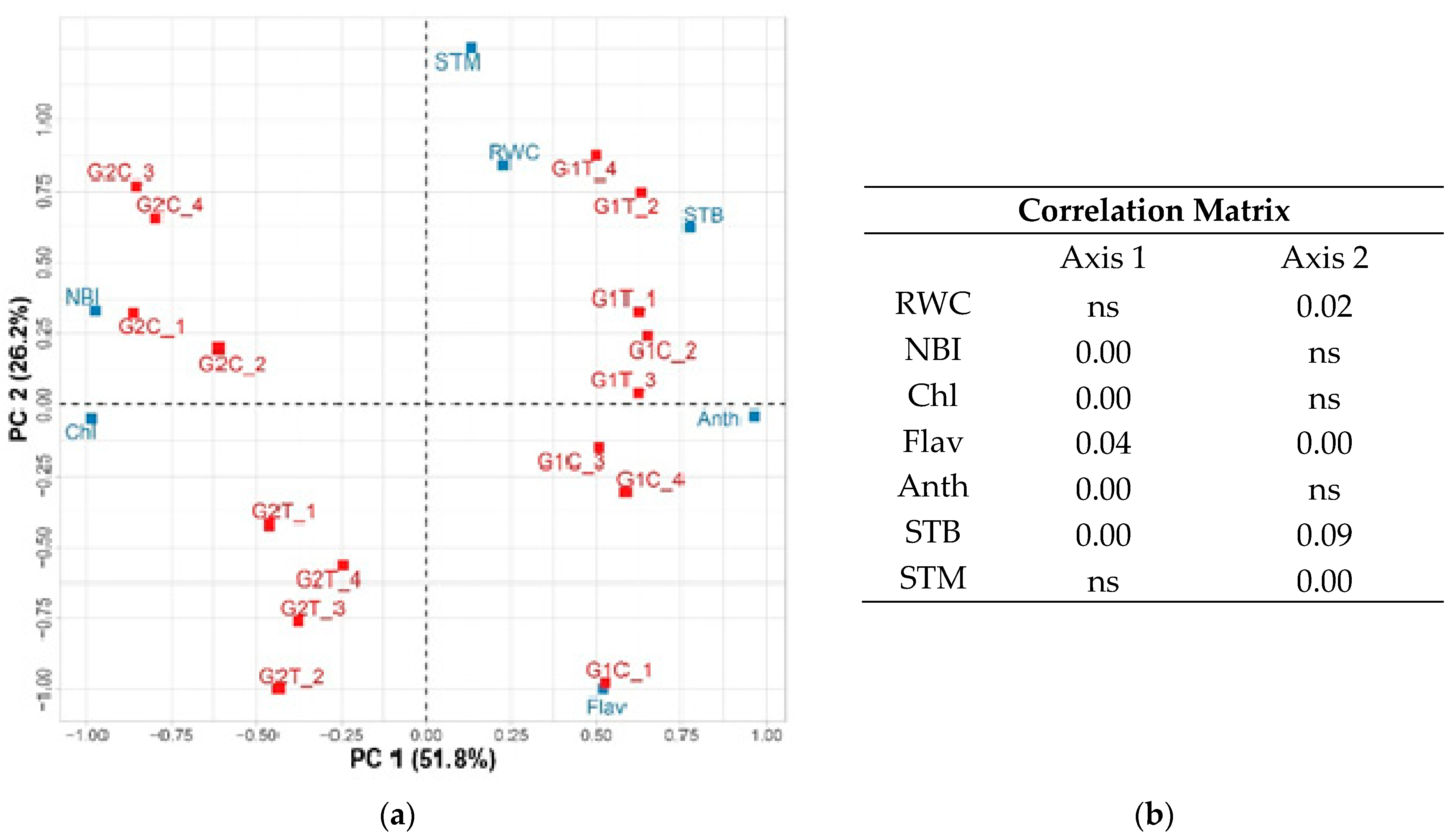
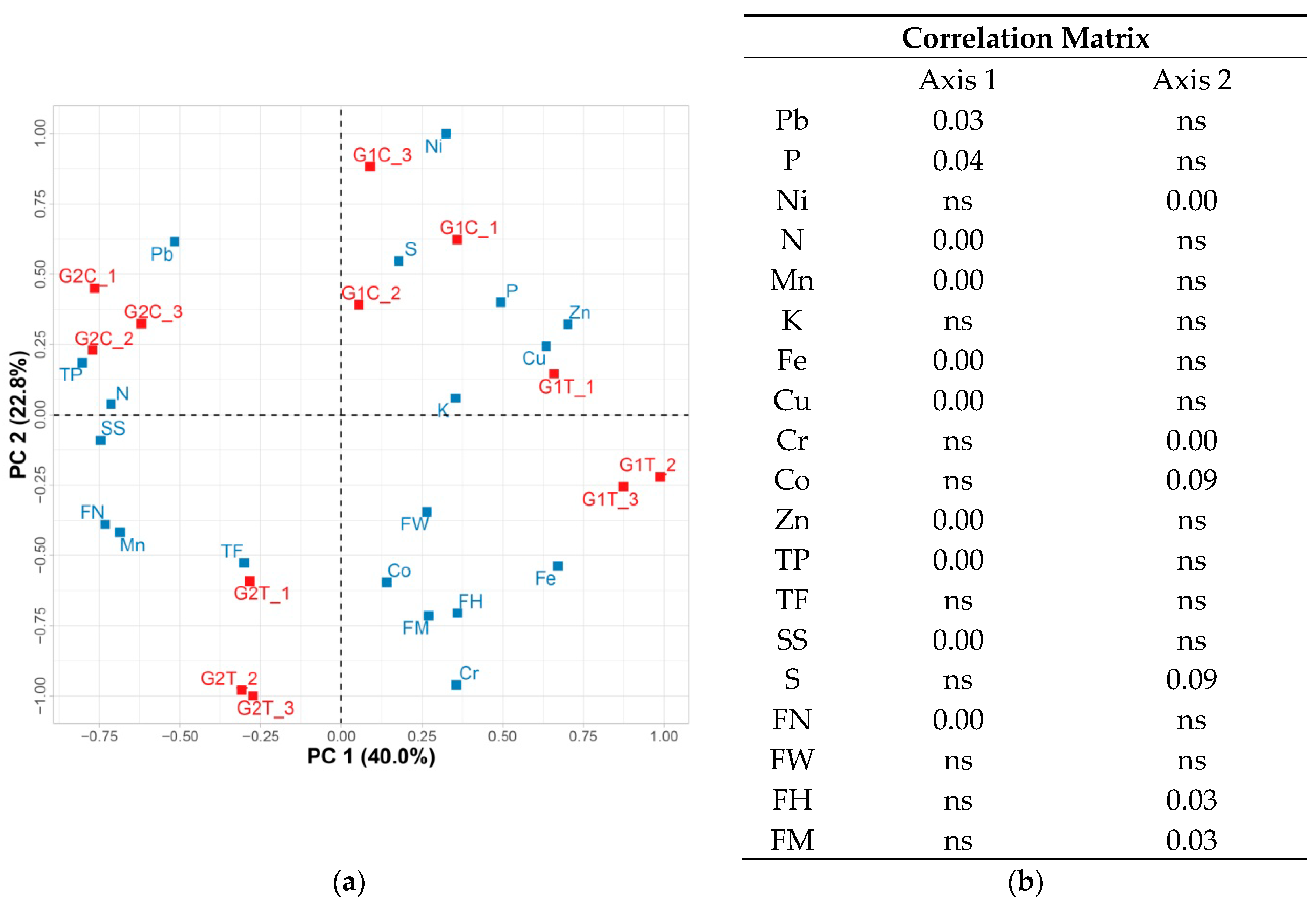
Principal Component Analysis
3. Discussion
4. Materials and Methods
4.1. Plant Material and Growth Conditions
4.2. Fungal Suspension Application
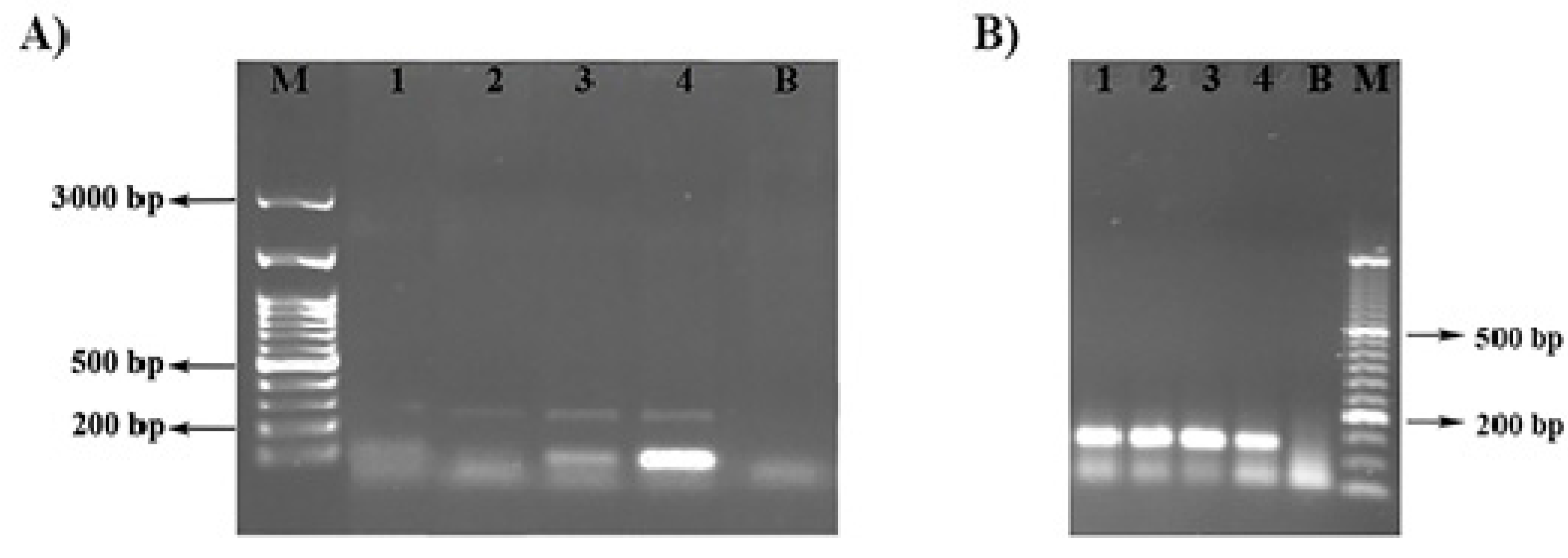
4.3. Detection of the Enzymatic Activity
4.4. RNA Extraction and Expression Analysis of Swolenin by Real-Time RT-PCR
4.5. Soil Sample Analysis
4.6. Determination of Soil water Content (SWC) and Leaf Relative Water (RWC) Content
4.7. Measurements of Epidermal Flavonols, Total Chlorophyll, Nitrogen Balance Index and Antocyanine
4.8. Extraction of Phenolics and Soluble Sugars
4.9. Determination of Total Phenolic Content (TP)
4.10. Determination of Total Flavonoid Content (TF)
4.11. Determination of Soluble Sugars Content (SS)
4.12. Determination of Starch Content (S)
4.13. Determination of Macroelements
4.14. Determination of Micronutrients and Heavy Metal Content in Soil and Tomato Fruit
4.15. Bioaccumulation Index (BI)
4.16. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ampt, E.A.; van Ruijven, J.; Raaijmakers, J.M.; Termorshuizen, A.J.; Mommer, L. Linking ecology and plant pathology to unravel the importance of soil-borne fungal pathogens in species-rich grasslands. Eur. J. Plant Pathol. 2019, 154, 141–156. [Google Scholar] [CrossRef] [Green Version]
- Frusciante, L.; Carli, P.; Ercolano, M.R.; Pernice, R.; Di Matteo, A.; Fogliano, V.; Pellegrini, N. Antioxidant nutritional quality of tomato. Mol. Nutr. Food Res. 2007, 51, 609–617. [Google Scholar] [CrossRef] [PubMed]
- Kanayama, Y. Sugar metabolism and fruit development in the tomato. Hort. J. 2017, 86, 417–425. [Google Scholar] [CrossRef] [Green Version]
- Bhowmik, D.; Kumar, K.S.; Paswan, S.; Srivastava, S. Tomato-a natural medicine and its health benefits. J. Pharmacogn. Phytochem. 2012, 1, 33–43. [Google Scholar]
- Wang, X.; Xing, Y. Evaluation of the effects of irrigation and fertilization on tomato fruit yield and quality: A principal component analysis. Sci. Rep. 2017, 7, 350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klunklin, W.; Savage, G. Effect on quality characteristics of tomatoes grown under well-watered and drought stress conditions. Foods 2017, 6, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez-Iruretagoiena, A.; Trebolazabala, J.; Martinez-Arkarazo, I.; de Diego, A.; Madariaga, J.M. Metals and metalloids in fruits of tomatoes (Solanum lycopersicum) and their cultivation soils in the Basque Country: Concentrations and accumulation trends. Food Chem. 2015, 173, 1083–1089. [Google Scholar] [CrossRef]
- Centeno, D.C.; Osorio, S.; Nunes-Nesi, A.; Bertolo, A.L.F.; Carneiro, R.T.; Araújo, W.L.; Steinhauser, M.-C.; Michalska, J.; Rohrmann, J.; Geigenberger, P.; et al. Malate Plays a Crucial Role in Starch Metabolism, Ripening, and Soluble Solid Content of Tomato Fruit and Affects Postharvest Softening. Plant Cell 2011, 23, 162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Shi, J.; Xie, Q.; Jiang, Y.; Yu, N.; Wang, E. Nutrient exchange and regulation in arbuscular mycorrhizal symbiosis. Mol. Plant 2017, 10, 1147–1158. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.Y.; Haque, M.M.; Molla, A.H.; Rahman, M.M.; Alam, M.Z. Antioxidant compounds and minerals in tomatoes by Trichoderma-enriched biofertilizer and their relationship with the soil environments. J. Integr. Agric. 2017, 16, 691–703. [Google Scholar] [CrossRef]
- de Santiago, A.; Quintero, J.M.; Avilés, M.; Delgado, A. Effect of Trichoderma asperellum strain T34 on iron, copper, manganese, and zinc uptake by wheat grown on a calcareous medium. Plant Soil 2011, 342, 97–104. [Google Scholar] [CrossRef]
- Li, Y.T.; Hwang, S.G.; Huang, Y.M.; Huang, C.H. Effects of Trichoderma asperellum on nutrient uptake and Fusarium wilt of tomato. Crop Prot. 2018, 110, 275–282. [Google Scholar] [CrossRef]
- Carillo, P.; Woo, S.L.; Comite, E.; El-Nakhel, C.; Rouphael, Y.; Fusco, G.M.; Borzacchiello, A.; Lanzuise, S.; Vinale, F. Application of Trichoderma harzianum, 6-pentyl-α-pyrone and plant biopolymer formulations modulate plant metabolism and fruit quality of plum tomatoes. Plants 2020, 9, 771. [Google Scholar] [CrossRef] [PubMed]
- Şesan, T.E.; Oancea, A.O.; Ştefan, L.M.; Mănoiu, V.S.; Ghiurea, M.; Răut, I.; Pomohaci, C.M. Effects of foliar treatment with a Trichoderma plant biostimulant consortium on Passiflora caerulea L. yield and quality. Microorganisms 2020, 8, 123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fiorini, L.; Guglielminetti, L.; Mariotti, L.; Curadi, M.; Picciarelli, P.; Scartazza, A.; Sarrocco, S.; Vannacci, G. Trichoderma harzianum T6776 modulates a complex metabolic network to stimulate tomato cv. Micro-Tom growth. Plant Soil 2016, 400, 351–366. [Google Scholar] [CrossRef]
- Harman, G.E.; Doni, F.; Khadka, R.B.; Uphoff, N. Endophytic strains of Trichoderma increase plants’ photosynthetic capability. J. Appl. 2021, 130, 529–546. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Cisneros, M.F.; Ornelas-Paz, J.J.; Olivas-Orozco, G.I.; Acosta-Muñiz, C.H.; Sepúlveda-Ahumada, D.R.; Pérez-Corral, D.A.; Rios-Velasco, C.; Salas-Marina, M.A.; Fernández-Pavía, S.P. Efect of Trichoderma spp. and phytopathogenic fungi on plant growth and tomato fruit quality. Rev. Mex. Fitopatol. 2018, 36, 444–456. [Google Scholar]
- Viterbo, A.D.A.; Chet, I. TasHyd1, a new hydrophobin gene from the biocontrol agent Trichoderma asperellum, is involved in plant root colonization. Mol. Plant Pathol. 2006, 7, 249–258. [Google Scholar] [CrossRef]
- Samolski, I.; Rinco’ n, A.M.; Pinzo’ n, L.M.; Viterbo, A.; Monte, E. The qid74 gene from Trichoderma harzianum has a role in root architecture and plant biofertilization. Microbiology 2012, 158, 129–138. [Google Scholar] [CrossRef]
- Ramírez-Valdespino, C.A.; Casas-Flores, S.; Olmedo-Monfil, V. Trichoderma as a model to study effector-like molecules. Front. Biol. 2019, 10, 1030. [Google Scholar] [CrossRef] [PubMed]
- Brotman, Y.; Briff, E.; Viterbo, A.; Chet, I. Role of swollenin, an expansin-like protein from Trichoderma, in plant root colonization. Plant Physiol. 2008, 147, 779–789. [Google Scholar] [CrossRef] [Green Version]
- Brotman, Y.; Lisec, J.; Méret, M.; Chet, I.; Willmitzer, L.; Viterbo, A. Transcript and metabolite analysis of the Trichoderma-induced systemic resistance response to Pseudomonas syringae in Arabidopsis thaliana. Microbiology 2012, 158, 139–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vukelić, I.; Racić, G.; Bojović, M.; Ćurčić, N.; Mrkajić, D.; Jovanović, D.; Panković, D. Effect of Trichoderma harzianum on morpho-physiological parametars and metal uptake of tomato plants. Matica Srpska J. Nat. Sci. 2020, 139, 61–71. [Google Scholar] [CrossRef]
- Vinale, F.; Sivasithamparam, K.; Ghisalberti, E.L.; Woo, S.L.; Nigro, M.; Marra, R.; Manganiello, G. Trichoderma secondary metabolites active on plants and fungal pathogens. Open Mycol. J. 2014, 8, 127–139. [Google Scholar] [CrossRef] [Green Version]
- Mehetre, S.T.; Mukherjee, P.K. Trichoderma improves nutrient use efficiency in crop plants. In Nutrient Use Efficiency: From Basics to Advances, 1st ed.; Rakshit, A., Bahadur, H.B., Singh, A.S., Eds.; Springer: New Delhi, India, 2015; pp. 173–180. [Google Scholar]
- Woo, S.L.; Ruocco, M.; Vinale, F.; Nigro, M.; Marra, R.; Lombardi, N.; Lorito, M. Trichoderma-based products and their widespread use in agriculture. Open Mycol. J. 2014, 8, 71–126. [Google Scholar] [CrossRef] [Green Version]
- Pascale, A.; Vinale, F.; Manganiello, G.; Nigro, M.; Lanzuise, S.; Ruocco, M.; Lorito, M. Trichoderma and its secondary metabolites improve yield and quality of grapes. Crop Prot. 2017, 92, 176–181. [Google Scholar] [CrossRef] [Green Version]
- Brotman, Y.; Landau, U.; Cuadros-Inostroza, Á.; Takayuki, T.; Fernie, A.R.; Chet, I.; Viterbo, A.; Willmitzer, L. Trichoderma-plant root colonization: Escaping early plant defense responses and activation of the antioxidant machinery for saline stress tolerance. PLoS Pathog. 2013, 9, e1003221. [Google Scholar] [CrossRef]
- Kapri, A.; Tewari, L. Phosphate solubilization potential and phosphatase activity of rhizospheric Trichoderma spp. Braz. J. Microbiol. 2010, 41, 787–795. [Google Scholar] [CrossRef] [Green Version]
- Harman, G.E.; Lorito, M.; Di Pietro, A.; Hayes, C.K. N-acetyl-β-glucosaminidase isolated from Trichoderma harzianum. U.S. Patent 5,474,926, 12 December 1995. [Google Scholar]
- Harman, G.E.; Howell, C.R.; Viterbo, A.; Chet, I.; Lorito, M. Trichoderma species—Opportunistic, avirulent plant symbionts. Nat. Rev. Microbiol. 2004, 2, 43–56. [Google Scholar] [CrossRef]
- Tucci, M.; Ruocco, M.; De Masi, L.; De Palma, M.; Lorito, M. The beneficial effect of Trichoderma spp. on tomato is modulated by the plant genotype. Mol. Plant Pathol. 2012, 12, 341–354. [Google Scholar] [CrossRef]
- Yedidia, I.; Srivastva, A.K.; Kapulnik, Y.; Chet, I. Effect of Trichoderma harzianum on microelement concentrations and increased growth of cucumber plants. Plant Soil. 2001, 235, 235–242. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Schansker, G.; Brestic, M.; Bussotti, F.; Calatayud, A.; Ferroni, L.; Goltsev, V.; Guidi, L.; Jajoo, A.; Li, P.; et al. Frequently asked questions about chlorophyll fluorescence, the sequel. Photosynth. Res. 2017, 132, 13–66. [Google Scholar] [CrossRef] [Green Version]
- Cerovic, Z.G.; Masdoumier, G.; Ghozlen, N.B.; Latouche, G. A new optical leaf-clip meter for simultaneous non-destructive assessment of leaf chlorophyll and epidermal flavonoids. Physiol. Plant. 2012, 146, 251–260. [Google Scholar] [CrossRef] [PubMed]
- Cerovic, Z.G.; Ounis, A.; Cartelat, A.; Latouche, G.; Goulas, Y.; Meyer, S.; Moya, I. The use of chlorophyll fluorescence excitation spectra for the non-destructive in situ assessment of UV-absorbing compounds in leaves. Plant Cell Environ. 2002, 25, 1663–1676. [Google Scholar] [CrossRef]
- Goulas, Y.; Cerovic, Z.G.; Cartelat, A.; Moya, I. Dualex: A new instrument for field measurements of epidermal ultraviolet absorbance by chlorophyll fluorescence. Appl. Opt. 2004, 43, 4488–4496. [Google Scholar] [CrossRef]
- Ibrahim, M.H.; Jaafar, H.Z.E. Dualex 4 reading values as indicator of total flavonoid and chlorophyll content of three varieties of Labisia pumila Benth.(Kacip Fatimah) seedlings under greenhouse condition. Trans. Malays. Soc. Plant Physiol. 2012, 20, 102–105. [Google Scholar]
- Agati, G.; Cerovic, Z.G.; Dalla Marta, A.; Di Stefano, V.; Pinelli, P.; Traversi, M.L.; Orlandini, S. Optically-assessed preformed flavonoids and susceptibility of grapevine to Plasmopara viticola under different light regimes. Funct. Plant Biol. 2008, 35, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Agati, G.; Tuccio, L.; Kusznierewicz, B.; Chmiel, T.; Bartoszek, A.; Kowalski, A.; Kaniszewski, S.; Grzegorzewska, M.; Kosson, R.; Kaniszewski, S. Nondestructive optical sensing of flavonols and chlorophyll in white head cabbage (Brassica oleracea L. var. capitata subvar. alba) grown under different nitrogen regimens. J. Agric. Food Chem. 2016, 64, 85–94. [Google Scholar] [CrossRef]
- Cartelat, A.; Cerovic, Z.G.; Goulas, Y.; Meyer, S.; Lelarge, C.; Prioul, J.L.; Barbottin, A.; Jeuffroy, M.H.; Gate, P.; Agati, G.; et al. Optically assessed contents of leaf polyphenolics and chlorophyll as indicators of nitrogen deficiency in wheat (Triticum aestivum L.). Field Crops Res. 2005, 91, 35–49. [Google Scholar] [CrossRef]
- Guller, N.S.; Pehlivan, N.; Karaoglu, S.A.; Guzel, S.; Bozdeveci, A. Trichoderma atroviride ID20G inoculation ameliorates drought stress-induced damages by improving antioxidant defence in maize seedlings. Acta Physiol. Plant. 2016, 38, 19. [Google Scholar] [CrossRef]
- Yu, Z.; Wang, Z.; Zhang, Y.; Wang, Y.; Liu, Z. Biocontrol and growth-promoting effect of Trichoderma asperellum TaspHu1 isolate from Juglans mandshurica rhizosphere soil. Microbiol. Res. 2021, 242, 126596. [Google Scholar] [CrossRef]
- Liu, B.; Ji, S.; Zhang, H.; Wang, Y.; Liu, Z. Isolation of Trichoderma in the rhizosphere soil of Syringa oblata from Harbin and their biocontrol and growth promotion function. Microbiol. Res. 2020, 235, 126445. [Google Scholar] [CrossRef] [PubMed]
- Alexandru, M.; Lazăr, D.; Ene, M.; Sesan, T.E. Influence of some Trichoderma species on photosynthesis intensity and pigments in tomatoes. Rom. Biotech. Lett. 2013, 18, 8499–8510. [Google Scholar]
- Hassan, S.; Mathesius, U. The role of flavonoids in root-rhizosphere signalling: Opportunities and challenges for improving plant-microbe interactions. J. Exp. Bot. 2012, 63, 3429–3444. [Google Scholar] [CrossRef] [Green Version]
- Mayo-Prieto, S.; Marra, R.; Vinale, F.; Rodríguez-González, Á.; Woo, S.L.; Lorito, M.; Casquero, P.A. Effect of Trichoderma velutinum and Rhizoctonia solani on the metabolome of bean plants (Phaseolus vulgaris L.). Int. J. Mol. Sci. 2019, 20, 549. [Google Scholar] [CrossRef] [Green Version]
- Coppola, M.; Diretto, G.; Digilio, M.C.; Woo, S.L.; Giuliano, G.; Molisso, D.; Rao, R. Transcriptome and metabolome reprogramming in tomato plants by Trichoderma harzianum strain T22 primes and enhances defence responses against aphids. Front. physiol. 2019, 10, 745. [Google Scholar] [CrossRef]
- Bhandari, S.R.; Cho, M.C.; Lee, J.G. Genotypic variation in carotenoid, ascorbic acid, total phenolic, and flavonoid contents, and antioxidant activity in selected tomato breeding lines. Hortic. Environ. Biotechnol. 2016, 57, 440–452. [Google Scholar] [CrossRef]
- Singh, S.P.; Singh, H.B.; Singh, D.K. Effect of Trichoderma harzianum on mineral component and antioxidant activity of tomato fruits. Vegetos 2013, 26, 237–244. [Google Scholar] [CrossRef]
- Ortega-García, J.G.; Montes-Belmont, R.; Rodríguez-Monroy, M.; Ramírez-Trujillo, J.A.; Suárez-Rodríguez, R.; Sepúlveda-Jiménez, G. Effect of Trichoderma asperellum applications and mineral fertilization on growth promotion and the content of phenolic compounds and flavonoids in onions. Sci. Hortic. 2015, 195, 8–16. [Google Scholar] [CrossRef]
- NawRocka, J.; Szczech, M.; MałolepSza, U. Trichoderma atroviride enhances phenolic synthesis and cucumber protection against Rhizoctonia solani. Plant Prot. Sci. 2017, 54, 17–23. [Google Scholar]
- Lombardi, N.; Salzano, A.M.; Troise, A.D.; Scaloni, A.; Vitaglione, P.; Vinale, F.; Marra, R.; Caira, S.; Lorito, M.; d’Errico, G.; et al. Effect of Trichoderma Bioactive Metabolite Treatments on the Production, Quality, and Protein Profile of Strawberry Fruits. J. Agric. Food Chem. 2020, 68, 7246–7258. [Google Scholar] [CrossRef]
- Martinez-Valverde, I.; Periago, M.J.; Provan, G.; Chesson, A. Phenolic compounds, lycopene and antioxidant activity in commercial varieties of tomato (Lycopersicum esculentum). J. Sci. Food. Agric. 2002, 82, 323–330. [Google Scholar] [CrossRef]
- Silva-Beltrán, N.P.; Ruiz-Cruz, S.; Cira-Chávez, L.A.; Estrada-Alvarado, M.I.; Ornelas-Paz, J.D.J.; López-Mata, M.A.; Del-Toro-Sánchez, C.L.; Ayala-Zavala, J.F.; Márquez-Ríos, E. Total phenolic, flavonoid, tomatine, and tomatidine contents and antioxidant and antimicrobial activities of extracts of tomato plant. Int. J. Anal. Chem. 2015, 2015, 284071. [Google Scholar] [CrossRef] [Green Version]
- Matsukura, C. Sugar accumulation in tomato fruit and its modification using molecular breeding techniques. In Functional Genomics and Biotechnology in Solanaceae and Cucurbitaceae Crops, 1st ed.; Ezura, H., Ariizumi, T., Garcia-Mas, J., Rose, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2016; pp. 141–154. [Google Scholar]
- Borrero, C.; Trillas, M.I.; Delgado, A.; Avilés, M. Effect of ammonium/nitrate ratio in nutrient solution on control of Fusarium wilt of tomato by Trichoderma asperellum T34. Plant Pathol. 2012, 61, 132–139. [Google Scholar] [CrossRef]
- Li, R.X.; Cai, F.; Pang, G.; Shen, Q.R.; Li, R.; Chen, W. Solubilisation of phosphate and 415 micronutrients by Trichoderma harzianum and its relationship with the promotion of 416 tomato plant growth. PLoS ONE 2015, 10, e0130081. [Google Scholar]
- Vargas, J.T.; Rodríguez-Monroy, M.; Meyer, M.L.; Montes-Belmont, R.; Sepúlveda-Jiménez, G. Trichoderma asperellum ameliorates phytotoxic effects of copper in onion (Allium cepa L.). Environ. Exp. Bot. 2017, 136, 85–93. [Google Scholar] [CrossRef]
- Kacprzak, M.J.; Rosikon, K.; Fijalkowski, K.; Grobelak, A. The effect of Trichoderma on heavy metal mobility and uptake by Miscanthus giganteus, Salix sp.; Phalarisarundinacea, and Panicum virgatum. Appl. Environ. Soil Sci. 2014, 2014, 506142. [Google Scholar] [CrossRef] [Green Version]
- Beheshtizadeh, H.; Rezaie, A.; Rezaie, A.; Ghandi, A. Principal component analysis and determination of the selection criteria in bread wheat (Triticum aestivum L.) genotypes. Int. J. Agric. Crop Sci. 2013, 5, 2024. [Google Scholar]
- Zivanovic, B.; Vidovic, M.; Komic, S.M.; Jovanovic, L.; Kolarz, P.; Morina, F.; Jovanovic, S.V. Contents of phenolics and carotenoids in tomato grown under polytunnels with different UV-transmission rates. Turk. J. Agric. For. 2017, 41, 113–120. [Google Scholar] [CrossRef]
- Grierson, D.; Kader, A.A. Fruit Ripening and Quality. In The Tomato Crop, 1st ed.; Atherton, J., Rudich, J., Eds.; Springer: Dordrecht, The Netherlands, 1986; pp. 241–280. [Google Scholar]
- Racić, G.; Körmöczi, P.; Kredics, L.; Raičević, V.; Mutavdžić, B.; Vrvić, M.M.; Panković, D. Effect of the edaphic factors and metal content in soil on the diversity of Trichoderma spp. Environ. Sci. Pollut. Res. 2017, 24, 3375–3386. [Google Scholar] [CrossRef] [Green Version]
- Racić, G.; Vukelić, I.; Prokić, L.; Ćurčić, N.; Zorić, M.; Jovanović, L.; Panković, D. The influence of Trichoderma brevicompactum treatment and drought on physiological parameters, abscisic acid content and signalling pathway marker gene expression in leaves and roots of tomato. Ann. Appl. Biol. 2018, 173, 213–221. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of Folin-Ciocalteu reagent. Methods Enzymol. 1998, 299, 52–178. [Google Scholar]
- Dewanto, V.; Wu, X.; Adom, K.K.; Liu, R.H. Thermal Processing Enhances the Nutritional Value of Tomatoes by Increasing Total Antioxidant Activity. J. Agric. Food Chem. 2002, 50, 3010–3014. [Google Scholar] [CrossRef]
- Yemm, E.W.; Willis, A. The estimation of carbohydrates in plant extracts by anthrone. Biochem. J. 1954, 57, 508. [Google Scholar] [CrossRef] [Green Version]
- Kostić, A.Ž.; Gašić, U.M.; Pešić, M.B.; Stanojević, S.P.; Barać, M.B.; Mačukanović-Jocić, M.P.; Avramov, S.N.; Tešić, Ž.L. Phytochemical analysis and total antioxidant capacity of rhizome, above-ground vegetative parts and flower of three iris species. Chem. Biodivers. 2019, 16, e1800565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rose, R.; Rose, C.L.; Omi, S.K.; Forry, K.R.; Durall, D.M.; Bigg, W.L. Starch determination by perchloric acid vs enzymes: Evaluating the accuracy and precision of six colorimetric methods. J. Agric. Food Chem. 1991, 39, 2–11. [Google Scholar] [CrossRef]
- Jones, J.B. Laboratory Guide for Conducting Soil Tests and Plant Analysis, 1st ed.; CRC Press: Boca Raton, FL, USA, 2001; pp. 109–121. [Google Scholar]
- Thioulouse, J.; Dray, S. ade4TkGUI: ade4 Tcl/Tk Graphical User Interface. R Package Version 0.2-5. 2009. Available online: http://CRAN.R-project.org/package=ade4TkGUI (accessed on 20 June 2021).




{kind=link}
{kind=link}
{kind=link}
| A | B | C | D | E | F | G | H | I | J | K | L | M | N | O | P | Q | R | S | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| T. harzianum SZMC 22660 | 1 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 3 | 0 | 0 | 0 | 0 | 0 | 3 | 0 | 0 |
| SWC (%) | RWC (%) | Chl | Flav | NBI | Anth | |
|---|---|---|---|---|---|---|
| G1C | 24.50 ± 2.4 ab | 49.32 ± 3.7 a | 29.1 ± 1.8 c | 0.67 ± 0.03 a | 43.15 ± 1.2 c | 0.11 ± 0.005 a |
| G1T | 27.33 ± 3.0 a | 48.73 ± 4.7 a | 26.4 ± 1.2 c | 0.59 ± 0.02 b | 48.23 ± 3.8 c | 0.12 ± 0.008 a |
| G2C | 30.75 ± 0.8 a | 50.18 ± 4.3 a | 41.7 ± 1.1 a | 0.52 ± 0.03 c | 77.5 ± 1.5 a | 0.08 ± 0.008 b |
| G2T | 30.95 ± 1.1 a | 54.67 ± 2.6 a | 36.53 ± 1.2 b | 0.65 ± 0.03 ab | 61.25 ± 4.6 b | 0.09 ± 0.008 b |
| STB (cm) | SDM (cm) | FW (cm × 10−2) | FH (cm × 10−2) | FN | FM (g) | |
|---|---|---|---|---|---|---|
| G1C | 16.36 ± 0.76 a | 10.6 ± 1.11 a | 60.03 ± 2.44 a | 57.8 ± 3.87 a | 12 ± 2 b | 130 ± 9.29 a |
| G1T | 16.7 ± 1.31 a | 12.36 ± 1.01 a | 61.96 ± 2.05 a | 61.3 ± 2.87 a | 11 ± 1 b | 143 ± 3.78 a |
| G2C | 13.87 ± 1.45 ab | 11.6 ± 1.05 a | 59.5 ± 4.37 a | 57.1 ± 1.42 a | 21 ± 3 a | 133 ± 4.57 a |
| G2T | 11.4 ± 0.87 b | 10.3 ± 0.81 a | 61.3 ± 3.61 a | 60.6 ± 1.57 a | 20 ± 1 a | 141 ± 11.35 a |
| TP (GEA mg/100 gDW) | TF (mg/100 gDW) | S (mg/g DW) | SS (mg/g DW) | N (%) | P (ppm) | K (ppm) | |
|---|---|---|---|---|---|---|---|
| G1C | 1632.41 ± 68.74 b | 455.56 ± 47.13 b | 30.61 ± 3.81 ab | 225.18 ± 14.96 bc | 1.28 ± 0.03 c | 0.79 ± 0.02 a | 2.85 ± 0.28 a |
| G1T | 1408.29 ± 45.21 c | 333.26 ± 9.51 c | 37.65 ± 2.01 a | 192 ± 31.57 c | 1.27 ± 0.02 c | 0.77 ± 0.015 a | 2.98 ± 0.14 a |
| G2C | 2001.77 ± 52.61 a | 368.14 ± 29.02 c | 36.52 ± 4.62 a | 337.07 ± 34.51 a | 2.21 ± 0.18 a | 0.71 ± 0.045 a | 2.78 ± 0.28 a |
| G2T | 1710.63 ± 33.31 b | 548.41 ± 30.66 a | 22.74 ± 4.09 b | 284.09 ± 4.26 ab | 1.59 ± 0.08 b | 0.74 ± 0.035 a | 2.80 ± 0.00 a |
| Mn | Fe | Cu | Zn | Ni | Cr | Pb | Co | |
|---|---|---|---|---|---|---|---|---|
| Fruit (a) | ||||||||
| G1C | 7.5 ± 0.1 c | 34.6 ± 1.0 b | 9.33 ± 0.15 a | 12.61 ± 0.61 a | 1.32 ± 0.05 a | 0.56 ± 0.005 b | 0.032 ± 0.01 a | 0.05 ± 0.01 a |
| G1T | 7.1 ± 0.05 d | 42.79 ± 0.11 a | 10.79 ± 1.55 ab | 12.79 ± 0.45 a | 1.08 ± 0.0 b | 0.89 ± 0.09 a | 0.0176 ± 0.03 b | 0.069 ± 0.02 a |
| G2C | 7.8 ± 0.1 b | 22.52 ± 1.00 c | 8.85 ± 0.02 ab | 11.16 ± 0.04 b | 0.995 ± 0.005 b | 0.48 ± 0.09 b | 0.030 ± 0.004 a | 0.05 ± 0.0005 a |
| G2T | 8.05 ± 0.1 a | 38.77 ± 2.81 a | 8.11 ± 0.36 b | 11.45 ± 0.79 ab | 0.49 ± 0.1 c | 0.99 ± 0.005 a | 0.0257 ± 0.045 a | 0.051 ± 0 a |
| Soil (b) | ||||||||
| 496.2 ± 3.9 | 13,685 ± 176.77 | 43.55 ± 1.21 | 39.13 ± 1.20 | 23.69 ± 0.59 | 23.07 ± 0.99 | 13.77 ± 0.94 | 9.043 ± 0.01 | |
| BI (c) | ||||||||
| G1C | 1.51 | 0.25 | 21.42 | 32.23 | 5.57 | 2.43 | 0.23 | 0.55 |
| G1T | 1.43 | 0.32 | 24.78 | 32.69 | 4.56 | 3.86 | 0.12 | 0.76 |
| G2C | 1.57 | 0.17 | 20.32 | 28.52 | 4.20 | 2.08 | 0.21 | 0.55 |
| G2T | 1.62 | 0.29 | 18.62 | 29.26 | 2.07 | 4.29 | 0.19 | 0.56 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vukelić, I.D.; Prokić, L.T.; Racić, G.M.; Pešić, M.B.; Bojović, M.M.; Sierka, E.M.; Kalaji, H.M.; Panković, D.M. Effects of Trichoderma harzianum on Photosynthetic Characteristics and Fruit Quality of Tomato Plants. Int. J. Mol. Sci. 2021, 22, 6961. https://doi.org/10.3390/ijms22136961
Vukelić ID, Prokić LT, Racić GM, Pešić MB, Bojović MM, Sierka EM, Kalaji HM, Panković DM. Effects of Trichoderma harzianum on Photosynthetic Characteristics and Fruit Quality of Tomato Plants. International Journal of Molecular Sciences. 2021; 22(13):6961. https://doi.org/10.3390/ijms22136961
Chicago/Turabian StyleVukelić, Igor D., Ljiljana T. Prokić, Gordana M. Racić, Mirjana B. Pešić, Mirjana M. Bojović, Edyta M. Sierka, Hazem M. Kalaji, and Dejana M. Panković. 2021. "Effects of Trichoderma harzianum on Photosynthetic Characteristics and Fruit Quality of Tomato Plants" International Journal of Molecular Sciences 22, no. 13: 6961. https://doi.org/10.3390/ijms22136961
APA StyleVukelić, I. D., Prokić, L. T., Racić, G. M., Pešić, M. B., Bojović, M. M., Sierka, E. M., Kalaji, H. M., & Panković, D. M. (2021). Effects of Trichoderma harzianum on Photosynthetic Characteristics and Fruit Quality of Tomato Plants. International Journal of Molecular Sciences, 22(13), 6961. https://doi.org/10.3390/ijms22136961