
Antimicrobial Activity of Curcumin in Nanoformulations: A Comprehensive Review

 ,
, 
Abstract
:1. Introduction
2. Free CUR
2.1. Antiviral Activity
2.2. Antibacterial Activity
2.3. Antifungal Activity
3. Curcumin in DDSs (Colloidal, Metal, and Hybrid Nanosystems)
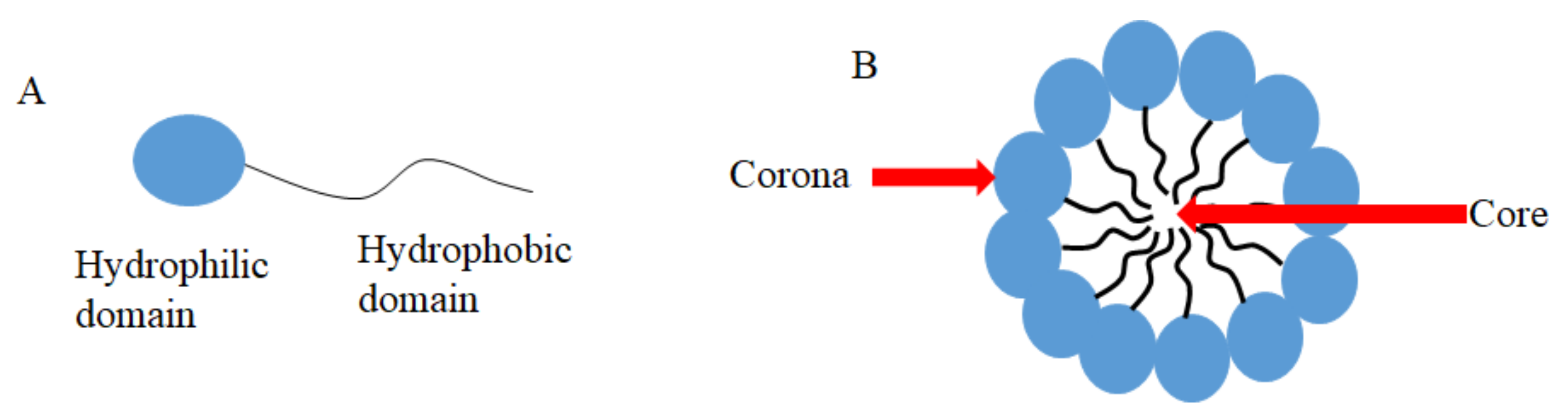
3.1. CUR in Micelles
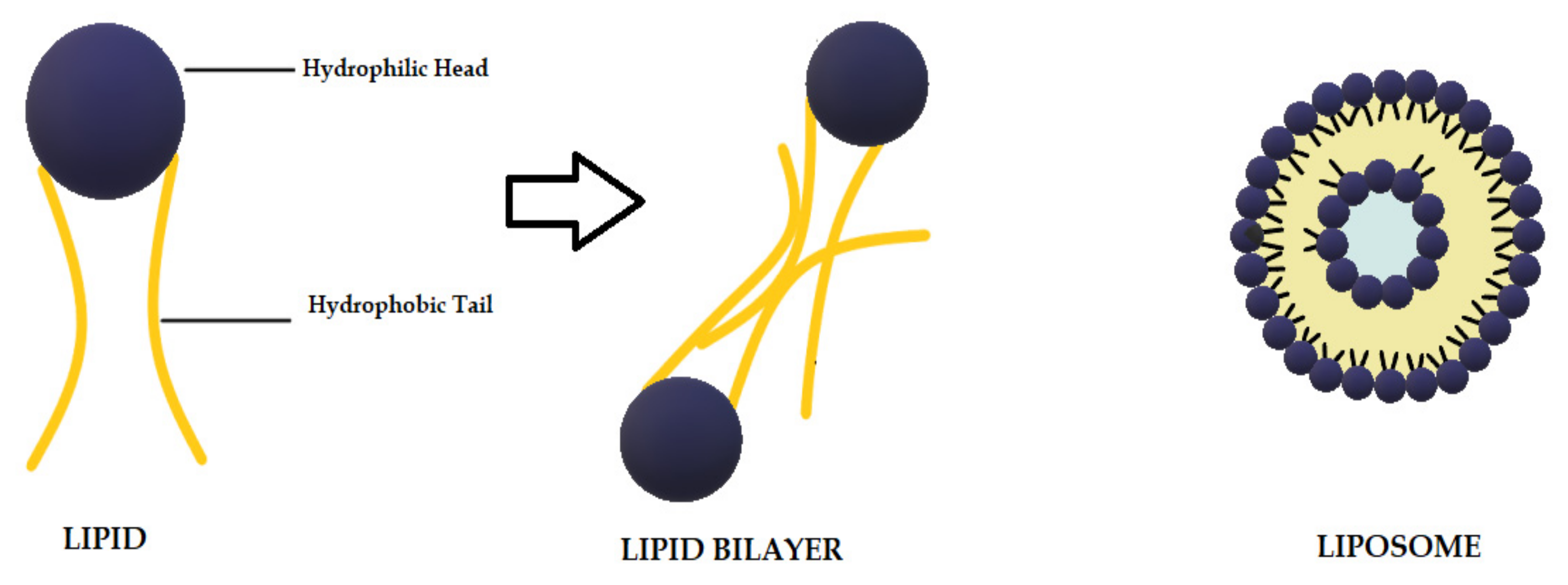
3.2. CUR in Liposomes
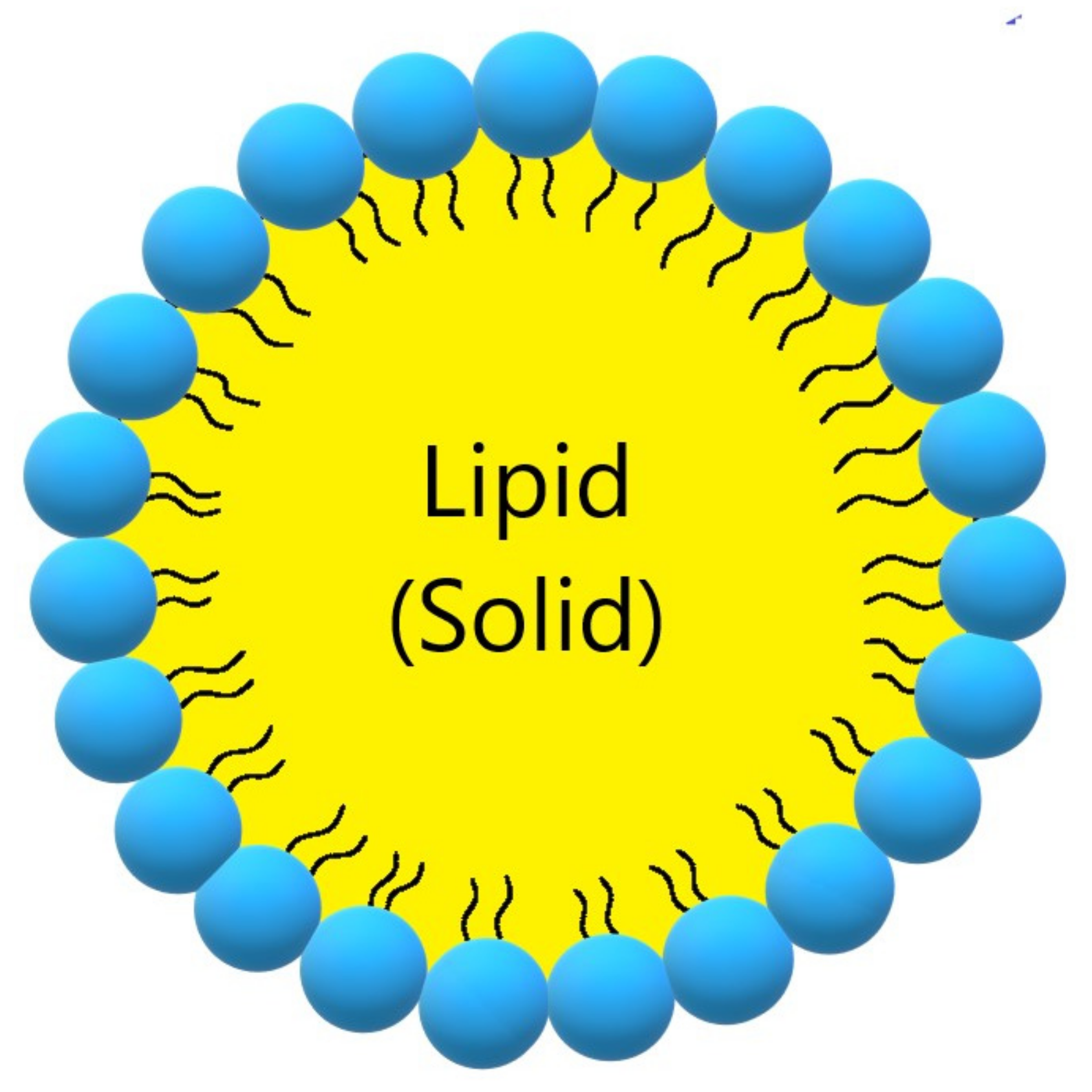
3.3. CUR in Solid Lipid Nanoparticles
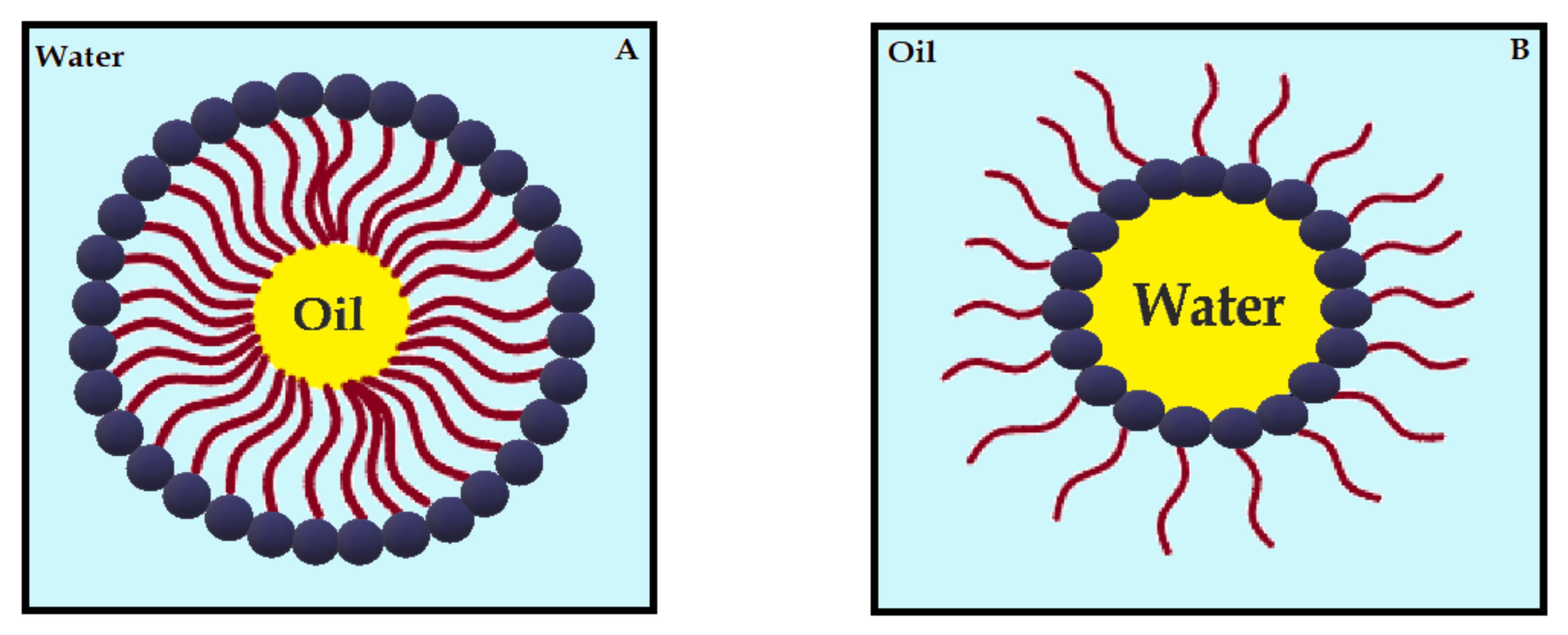
3.4. CUR in Nanoemulsions
3.5. CUR in Cyclodextrin
3.6. CUR in Chitosan
3.7. CUR in Other Polymeric DDS
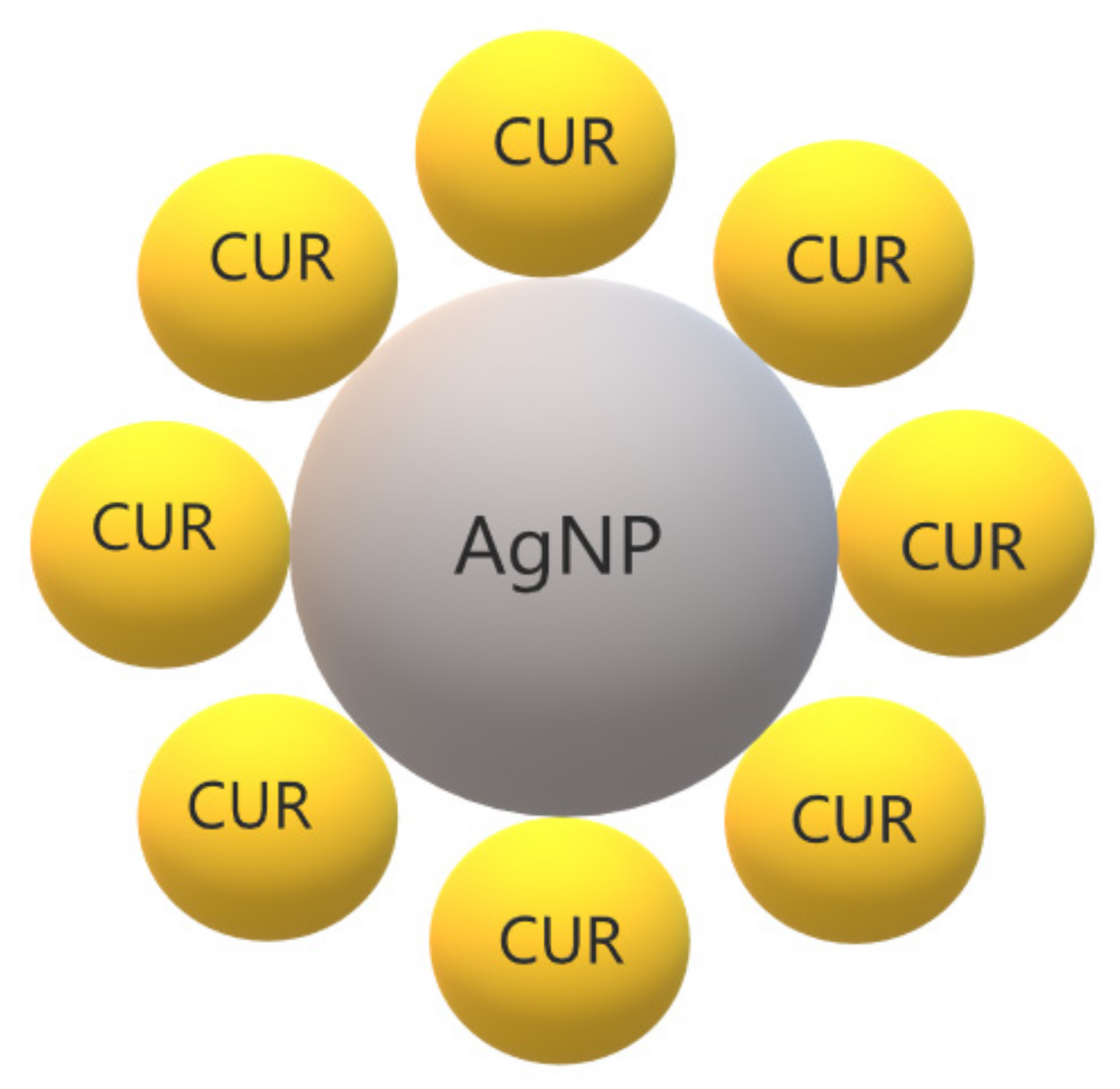
3.8. CUR with Metallic Nanoparticles
3.9. CUR in Mesoporous Particles
3.10. CUR in Graphene Nanocomposites
3.11. CUR in Quantum Dots
3.12. CUR in Films, Hydrogels, and Other Nanomaterials
4. Conclusions and Future Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Davis, C.E.; Rossati, A.; Bargiacchi, O.; Garavelli, P.L.; McMichael, A.J.; Juliusson, G. Globalization, climate change, and human health. N. Engl. J. Med. 2013, 369, 94–96. [Google Scholar] [CrossRef] [PubMed]
- Saker, L.; Lee, K.; Cannito, B.; Gilmore, A.; Campbell-Lendrum, D. Globalization and Infectious Diseases: A Review of the Linkages; WHO Special Programme on Tropical Diseases Research; WHO: Geneva, Switzerland, 2004. [Google Scholar]
- Gorji, S.; Gorji, A. COVID-19 pandemic: The possible influence of the long-term ignorance about climate change. Environ. Sci. Pollut. Res. 2021, 28, 15575–15579. [Google Scholar] [CrossRef] [PubMed]
- Tanwar, J.; Das, S.; Fatima, Z.; Hameed, S. Multidrug resistance: An emerging crisis. Interdiscip. Perspect. Infect. Dis. 2014, 2014, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farha, M.A.; Brown, E.D. Drug repurposing for antimicrobial discovery. Nat. Microbiol. 2019, 4, 565–577. [Google Scholar] [CrossRef]
- Moore, B.S.; Carter, G.T.; Brönstrup, M. Editorial: Are natural products the solution to antimicrobial resistance? Nat. Prod. Rep. 2017, 34, 685–686. [Google Scholar] [CrossRef]
- Nunes, T.S.B.S.; Rosa, L.M.; Vega-Chacón, Y.; Mima, E.G.D.O. Fungistatic action of N-Acetylcysteine on Candida albicans biofilms and its interaction with antifungal agents. Microorganisms 2020, 8, 980. [Google Scholar] [CrossRef]
- Hewlings, S.J.; Kalman, D.S. Curcumin: A review of its effects on human health. Foods 2017, 6, 92. [Google Scholar] [CrossRef]
- Gupta, S.C.; Patchva, S.; Aggarwal, B.B. Therapeutic roles of curcumin: Lessons learned from clinical trials. Aaps J. 2012, 15, 195–218. [Google Scholar] [CrossRef] [Green Version]
- Tomeh, M.A.; Hadianamrei, R.; Zhao, X. A review of curcumin and its derivatives as anticancer agents. Int. J. Mol. Sci. 2019, 20, 1033. [Google Scholar] [CrossRef] [Green Version]
- Praditya, D.; Kirchhoff, L.; Brüning, J.; Rachmawati, H.; Steinmann, J.; Steinmann, E. Anti-infective properties of the golden spice curcumin. Front. Microbiol. 2019, 10, 912. [Google Scholar] [CrossRef] [Green Version]
- Cas, M.D.; Ghidoni, R. Dietary Curcumin: Correlation between bioavailability and health potential. Nutrients 2019, 11, 2147. [Google Scholar] [CrossRef] [Green Version]
- Priyadarsini, K.I. The Chemistry of curcumin: From extraction to therapeutic agent. Molecules 2014, 19, 20091–20112. [Google Scholar] [CrossRef] [Green Version]
- Deljoo, S.; Rabiee, N.; Rabiee, M. Curcumin-hybrid nanoparticles in drug delivery system (review). Asian J. Nanosci. Mater. 2019, 2, 66–91. [Google Scholar] [CrossRef]
- Safari, J.; Zarnegar, Z. Advanced drug delivery systems: Aanotechnology of health design a review. J. Saudi Chem. Soc. 2014, 18, 85–99. [Google Scholar] [CrossRef]
- Moghadamtousi, S.Z.; Kadir, H.A.; Hassandarvish, P.; Tajik, H.; Abubakar, S.; Zandi, K. A review on antibacterial, antiviral, and antifungal activity of curcumin. Biomed Res. Int. 2014, 2014, 186864. [Google Scholar] [CrossRef]
- Da Silva, A.C.; Santos, P.D.D.F.; Silva, J.T.D.P.; Leimann, F.V.; Bracht, L.; Gonçalves, O.H. Impact of curcumin nanoformulation on its antimicrobial activity. Trends Food Sci. Technol. 2018, 72, 74–82. [Google Scholar] [CrossRef]
- Sharifi, S.; Fathi, N.; Memar, M.Y.; Khatibi, S.M.H.; Khalilov, R.; Negahdari, R.; Vahed, S.Z.; Dizaj, S.M. Anti-microbial activity of curcumin nanoformulations: New trends and future perspectives. Phytother. Res. 2020, 34, 1926–1946. [Google Scholar] [CrossRef]
- Gao, Y.; Tai, W.; Wang, N.; Li, X.; Jiang, S.; Debnath, A.K.; Du, L.; Chen, S. Identification of novel natural products as effective and broad-spectrum anti-zika virus inhibitors. Viruses 2019, 11, 1019. [Google Scholar] [CrossRef] [Green Version]
- Patwardhan, M.; Morgan, M.T.; Dia, V.; D’Souza, D.H. Heat sensitization of hepatitis A virus and Tulane virus using grape seed extract, gingerol and curcumin. Food Microbiol. 2020, 90, 103461. [Google Scholar] [CrossRef]
- Li, H.; Zhong, C.; Wang, Q.; Chen, W.; Yuan, Y. Curcumin is an APE1 redox inhibitor and exhibits an antiviral activity against KSHV replication and pathogenesis. Antivir. Res. 2019, 167, 98–103. [Google Scholar] [CrossRef]
- Roshdy, W.H.; Rashed, H.A.; Kandeil, A.; Mostafa, A.; Moatasim, Y.; Kutkat, O.; Shama, N.M.A.; Gomaa, M.R.; El-Sayed, I.H.; El Guindy, N.M.; et al. EGYVIR: An immunomodulatory herbal extract with potent antiviral activity against SARS-CoV-2. PLoS ONE 2020, 15, e0241739. [Google Scholar] [CrossRef] [PubMed]
- Wen, C.-C.; Kuo, Y.-H.; Jan, J.-T.; Liang, P.-H.; Wang, S.-Y.; Liu, H.-G.; Lee, C.-K.; Chang, S.-T.; Kuo, C.-J.; Lee, S.-S.; et al. Specific plant terpenoids and lignoids possess potent antiviral activities against severe acute respiratory syndrome coronavirus. J. Med. Chem. 2007, 50, 4087–4095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryu, Y.B.; Park, S.-J.; Kim, Y.M.; Lee, J.-Y.; Seo, W.D.; Chang, J.S.; Park, K.H.; Rho, M.-C.; Lee, W.S. SARS-CoV 3CLpro inhibitory effects of quinone-methide triterpenes from Tripterygium regelii. Bioorganic Med. Chem. Lett. 2010, 20, 1873–1876. [Google Scholar] [CrossRef] [PubMed]
- Park, J.-Y.; Jeong, H.J.; Kim, J.H.; Kim, Y.M.; Park, S.-J.; Kim, D.; Park, K.H.; Lee, W.S.; Ryu, Y.B. Diarylheptanoids from Alnus japonica inhibit papain-like protease of severe acute respiratory syndrome coronavirus. Biol. Pharm. Bull. 2012, 35, 2036–2042. [Google Scholar] [CrossRef] [Green Version]
- Adeyemi, O.S.; Obeme-Imom, J.I.; Akpor, B.O.; Rotimi, D.; Batiha, G.E.-S.; Owolabi, A. Altered redox status, DNA damage and modulation of L-tryptophan metabolism contribute to antimicrobial action of curcumin. Heliyon 2020, 6, e03495. [Google Scholar] [CrossRef]
- Güran, M.; Şanlıtürk, G.; Kerküklü, N.R.; Altundag, E.M.; Yalçın, A.S. Combined effects of quercetin and curcumin on anti-inflammatory and antimicrobial parameters in vitro. Eur. J. Pharm. 2019, 859, 172486. [Google Scholar] [CrossRef]
- Freitas, M.A.; Pereira, A.H.; Pinto, J.G.; Casas, A.; Ferreira-Strixino, J. Bacterial viability after antimicrobial photodynamic therapy with curcumin on multiresistant Staphylococcus aureus. Futur. Microbiol. 2019, 14, 739–748. [Google Scholar] [CrossRef]
- Teixeira, C.G.D.S.; Sanitá, P.V.; Ribeiro, A.P.D.; Dias, L.M.; Jorge, J.H.; Pavarina, A.C. Antimicrobial photodynamic therapy effectiveness against susceptible and methicillin-resistant Staphylococcus aureus biofilms. Photodiagn. Photodyn. 2020, 30, 101760. [Google Scholar] [CrossRef]
- Akhtar, F.; Khan, A.U.; Misba, L.; Akhtar, K.; Ali, A. Antimicrobial and antibiofilm photodynamic therapy against vancomycin resistant Staphylococcus aureus (VRSA) induced infection in vitro and in vivo. Eur. J. Pharm. Biopharm. 2021, 160, 65–76. [Google Scholar] [CrossRef]
- Paolillo, F.R.; Rodrigues, P.G.S.; Bagnato, V.S.; Alves, F.; Pires, L.; Corazza, A.V. The effect of combined curcumin-mediated photodynamic therapy and artificial skin on Staphylococcus aureus-infected wounds in rats. Lasers Med. Sci. 2020, 1–8. [Google Scholar] [CrossRef]
- Li, Y.; Xu, Y.; Liao, Q.; Xie, M.; Tao, H.; Wang, H. Synergistic effect of hypocrellin B and curcumin on photodynamic inactivation of Staphylococcus aureus. Microb. Biotechnol. 2021, 14, 692–707. [Google Scholar] [CrossRef]
- Corrêa, T.Q.; Blanco, K.C.; Garcia, É.B.; Perez, S.M.L.; Chianfrone, D.J.; Morais, V.S.; Bagnato, V.S. Effects of ultraviolet light and curcumin-mediated photodynamic inactivation on microbiological food safety: A study in meat and fruit. Photodiagn. Photodyn. 2020, 30, 101678. [Google Scholar] [CrossRef]
- Le, T.D.; Phasupan, P.; Nguyen, L.T. Antimicrobial photodynamic efficacy of selected natural photosensitizers against food pathogens: Impacts and interrelationship of process parameters. Photodiagn. Photodyn. 2020, 32, 102024. [Google Scholar] [CrossRef]
- Versace, D.-L.; Moran, G.; Belqat, M.; Spangenberg, A.; Meallet-Renault, R.; Abbad-Andaloussi, S.; Brezova, V.; Malval, J.-P. Highly virulent bactericidal effects of curcumin-based μ-cages fabricated by two-photon polymerization. ACS Appl. Mater. Interfaces 2020, 12, 5050–5057. [Google Scholar] [CrossRef]
- Alves, F.; Guimarães, G.G.; Inada, N.M.; Pratavieira, S.; Bagnato, V.S.; Kurachi, C. Strategies to improve the antimicrobial efficacy of photodynamic, sonodynamic, and sonophotodynamic therapies. Lasers Surg. Med. 2021. [Google Scholar] [CrossRef]
- Azucena, R.-C.I.; Roberto, C.-L.J.; Martin, Z.-R.; Rafael, C.-Z.; Leonardo, H.-H.; Gabriela, T.-P.; Araceli, C.-R. Drug susceptibility testing and synergistic antibacterial activity of curcumin with antibiotics against Enterotoxigenic Escherichia coli. Antibiotics 2019, 8, 43. [Google Scholar] [CrossRef] [Green Version]
- Sanchez-Villamil, J.I.; Navarro-Garcia, F.; Castillo-Romero, A.; Gutiérrez-Gutiérrez, F.; Tapia, D.; Tapia-Pastrana, G. Curcumin blocks cytotoxicity of Enteroaggregative and Enteropathogenic Escherichia coli by blocking Pet and EspC Proteolytic release from bacterial outer membrane. Front. Cell. Infect. Microbiol. 2019, 9, 334. [Google Scholar] [CrossRef]
- Kareem, S.M.; Mahmood, S.S.; Hindi, N. Effects of Curcumin and Silymarin on the Shigella dysenteriae and Campylobacter jejuni in vitro. J. Gastrointest. Cancer 2019, 51, 824–828. [Google Scholar] [CrossRef]
- Gao, Y.; Wu, J.; Li, Z.; Zhang, X.; Lu, N.; Xue, C.; Leung, A.W.; Xu, C.; Tang, Q. Curcumin-mediated photodynamic inactivation (PDI) against DH5α contaminated in oysters and cellular toxicological evaluation of PDI-treated oysters. Photodiagn. Photodyn. 2019, 26, 244–251. [Google Scholar] [CrossRef]
- Darmani, H.; Smadi, E.A.; Bataineh, S.M. Blue light emitting diodes enhance the antivirulence effects of curcumin against Helicobacter pylori. J. Med. Microbiol. 2020, 69, 617–624. [Google Scholar] [CrossRef]
- Abdulrahman, H.; Misba, L.; Ahmad, S.; Khan, A.U. Curcumin induced photodynamic therapy mediated suppression of quorum sensing pathway of Pseudomonas aeruginosa: An approach to inhibit biofilm in vitro. Photodiagn. Photodyn. 2020, 30, 101645. [Google Scholar] [CrossRef]
- Chang, K.; Cheng, Y.; Lai, M.; Hu, A. Identification of carbonylated proteins in a bactericidal process induced by curcumin with blue light irradiation on imipenem-resistant Acinetobacter baumannii. Rapid Commun. Mass Spectrom. 2019, 34, e8548. [Google Scholar] [CrossRef]
- Sharahi, J.Y.; Ahovan, Z.A.; Maleki, D.T.; Rad, Z.R.; Rad, Z.R.; Goudarzi, M.; Shariati, A.; Bostanghadiri, N.; Abbasi, E.; Hashemi, A. In vitro antibacterial activity of curcumin-meropenem combination against extensively drug-resistant (XDR) bacteria isolated from burn wound infections. Avicenna J. Phytomed. 2020, 10, 3–10. [Google Scholar]
- Lakes, J.E.; Richards, C.I.; Flythe, M.D. Inhibition of bacteroidetes and firmicutes by select phytochemicals. Anaerobe 2020, 61, 102145. [Google Scholar] [CrossRef]
- Kumari, S.; Jayakumar, S.; Gupta, G.D.; Bihani, S.C.; Sharma, D.; Kutala, V.K.; Sandur, S.K.; Kumar, V. Antibacterial activity of new structural class of semisynthetic molecule, triphenyl-phosphonium conjugated diarylheptanoid. Free. Radic. Biol. Med. 2019, 143, 140–145. [Google Scholar] [CrossRef]
- Kumar, P.; Kandi, S.K.; Mukhopadhyay, K.; Rawat, D.S. Gagandeep Synthesis of novel monocarbonyl curcuminoids, evaluation of their efficacy against MRSA, including ex vivo infection model and their mechanistic studies. Eur. J. Med. Chem. 2020, 195, 112276. [Google Scholar] [CrossRef]
- Polaquini, C.R.; Morão, L.G.; Nazaré, A.C.; Torrezan, G.S.; Dilarri, G.; Cavalca, L.B.; Campos, D.L.; Silva, I.C.; Pereira, J.A.; Scheffers, D.-J.; et al. Antibacterial activity of 3,3′-dihydroxycurcumin (DHC) is associated with membrane perturbation. Bioorg. Chem. 2019, 90, 103031. [Google Scholar] [CrossRef]
- Cerveira, M.M.; Vianna, H.S.; Ferrer, E.M.K.; da Rosa, B.N.; de Pereira, C.M.P.; Baldissera, M.D.; Lopes, L.Q.S.; Rech, V.C.; Giongo, J.L.; Vaucher, R.D.A. Bioprospection of novel synthetic monocurcuminoids: Antioxidant, antimicrobial, and in vitro cytotoxic activities. Biomed. Pharm. 2021, 133, 111052. [Google Scholar] [CrossRef] [PubMed]
- Duracka, M.; Lukac, N.; Kacaniova, M.; Kantor, A.; Hleba, L.; Ondruska, L.; Tvrda, E. Antibiotics versus natural biomolecules: The case of in vitro induced bacteriospermia by Enterococcus faecalis in rabbit semen. Molecules 2019, 24, 4329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rocha, M.P.; Santos, M.S.; Rodrigues, P.L.F.; Araújo, T.S.D.; de Oliveira, J.M.; Rosa, L.P.; Bagnato, V.S.; da Silva, F.C. Photodynamic therapry with curcumin in the reduction of Enterococcus faecalis biofilm in bone cavity: Microbiological and spectral fluorescense analysis. Photodiagn. Photodyn. 2021, 33, 102084. [Google Scholar] [CrossRef]
- Rafeeq, S.; Shiroodi, S.; Schwarz, M.H.; Nitin, N.; Ovissipour, R. Inactivation of Aeromonas hydrophila and Vibrio parahaemolyticus by curcumin-mediated photosensitization and nanobubble-ultrasonication approaches. Foods 2020, 9, 1306. [Google Scholar] [CrossRef]
- Ganapathy, S.; Sainudeen, S.; Nair, V.S.; Zarbah, M.; Abdulla, A.M.; Najeeb, C.M. Can herbal extracts serve as antibacterial root canal irrigating solutions? antimicrobial efficacy of Tylophora indica, Curcumin longa, Phyllanthus amarus, and sodium hypochlorite on Enterococcus faecalis biofilms formed on tooth substrate: In vitro study. J. Pharm. Bioallied. Sci. 2020, 12, S423–S429. [Google Scholar] [CrossRef]
- Azizi, A.; Shohrati, P.; Goudarzi, M.; Lawaf, S.; Rahimi, A. Comparison of the effect of photodynamic therapy with curcumin and methylene Blue on Streptococcus mutans bacterial colonies. Photodiagn. Photodyn. 2019, 27, 203–209. [Google Scholar] [CrossRef]
- Soares, J.M.; Silva, K.O.O.; Inada, N.M.; Bagnato, V.S.; Blanco, K.C. Optimization for microbial incorporation and efficiency of photodynamic therapy using variation on curcumin formulation. Photodiagn. Photodyn. 2020, 29, 101652. [Google Scholar] [CrossRef]
- Méndez, D.A.C.; Gutierres, E.; Lamarque, G.C.C.; Rizzato, V.L.; Buzalaf, M.A.R.; Machado, M.A.A.M.; Cruvinel, T. The effectiveness of curcumin-mediated antimicrobial photodynamic therapy depends on pre-irradiation and biofilm growth times. Photodiagn. Photodyn. 2019, 27, 474–480. [Google Scholar] [CrossRef]
- Li, X.; Yin, L.; Ramage, G.; Li, B.; Tao, Y.; Zhi, Q.; Lin, H.; Zhou, Y. Assessing the impact of curcumin on dual-species biofilms formed by Streptococcus mutans and Candida albicans. Microbiology 2019, 8, e937. [Google Scholar] [CrossRef] [Green Version]
- Mahdizade-Ari, M.; Pourhajibagher, M.; Bahador, A. Changes of microbial cell survival, metabolic activity, efflux capacity, and quorum sensing ability of Aggregatibacter actinomycetemcomitans due to antimicrobial photodynamic therapy-induced bystander effects. Photodiagn. Photodyn. 2019, 26, 287–294. [Google Scholar] [CrossRef]
- Pan, H.; Wang, D.; Zhang, F. In vitro antimicrobial effect of curcumin-based photodynamic therapy on Porphyromonas gingivalis and Aggregatibacter actinomycetemcomitans. Photodiagn. Photodyn. 2020, 32, 102055. [Google Scholar] [CrossRef]
- Böcher, S.; Wenzler, J.-S.; Falk, W.; Braun, A. Comparison of different laser-based photochemical systems for periodontal treatment. Photodiagn. Photodyn. 2019, 27, 433–439. [Google Scholar] [CrossRef]
- Zago, L.H.D.P.; De Annunzio, S.R.; De Oliveira, K.T.; Barbugli, P.A.; Valdes, B.R.; Feres, M.; Fontana, C.R. Antimicrobial photodynamic therapy against metronidazole-resistant dental plaque bactéria. J. Photochem. Photobiol. B Biol. 2020, 209, 111903. [Google Scholar] [CrossRef]
- Sha, A.M.; Garib, B.T.; Sha, A. Antibacterial effect of curcumin against clinically isolated Porphyromonas gingivalis and connective tissue reactions to curcumin gel in the subcutaneous tissue of rats. Biomed. Res. Int. 2019, 2019, 6810936. [Google Scholar] [CrossRef] [Green Version]
- Ivanaga, C.A.; Miessi, D.M.J.; Nuernberg, M.A.A.; Claudio, M.M.; Garcia, V.G.; Theodoro, L.H. Antimicrobial photodynamic therapy (aPDT) with curcumin and LED, as an enhancement to scaling and root planing in the treatment of residual pockets in diabetic patients: A randomized and controlled split-mouth clinical trial. Photodiagn. Photodyn. 2019, 27, 388–395. [Google Scholar] [CrossRef]
- Da Silva, F.C.; Rosa, L.P.; Santos, G.P.D.O.; Inada, N.M.; Blanco, K.C.; Araújo, T.S.D.; Bagnato, V.S. Total mouth photodynamic therapy mediated by blue led and curcumin in individuals with AIDS. Expert Rev. Anti-Infect. 2020, 18, 689–696. [Google Scholar] [CrossRef]
- Narayanan, V.S.; Muddaiah, S.; Shashidara, R.; Sudheendra, U.S.; Deepthi, N.C.; Samaranayake, L. Variable antifungal activity of curcumin against planktonic and biofilm phase of different candida species. Indian J. Dent. Res. 2020, 31, 145–148. [Google Scholar] [CrossRef]
- Tan, Y.; Leonhard, M.; Moser, D.; Ma, S.; Schneider-Stickler, B. Antibiofilm efficacy of curcumin in combination with 2-aminobenzimidazole against single- and mixed-species biofilms of Candida albicans and Staphylococcus aureus. Colloids Surf. B Biointerfaces 2019, 174, 28–34. [Google Scholar] [CrossRef]
- Da Silva, F.C.; Rodrigues, P.L.F.; Araújo, T.S.D.; Santos, M.S.; de Oliveira, J.M.; Rosa, L.P.; Santos, G.P.D.O.; de Araújo, B.P.; Bagnato, V.S. Fluorescence spectroscopy of Candida albicans biofilms in bone cavities treated with photodynamic therapy using blue LED (450 nm) and curcumin. Photodiagn. Photodyn. 2019, 26, 366–370. [Google Scholar] [CrossRef]
- Ma, J.; Shi, H.; Sun, H.; Li, J.; Bai, Y. Antifungal effect of photodynamic therapy mediated by curcumin on Candida albicans biofilms in vitro. Photodiagn. Photodyn. 2019, 27, 280–287. [Google Scholar] [CrossRef]
- Jordão, C.C.; de Sousa, T.V.; Klein, M.I.; Dias, L.M.; Pavarina, A.C.; Carmello, J.C. Antimicrobial photodynamic therapy reduces gene expression of Candida albicans in biofilms. Photodiagn. Photodyn. 2020, 31, 101825. [Google Scholar] [CrossRef]
- Merigo, E.; Chevalier, M.; Conti, S.; Ciociola, T.; Fornaini, C.; Manfredi, M.; Vescovi, P.; Doglio, A. Antimicrobial effect on Candida albicans biofilm by application of different wavelengths and dyes and the synthetic killer decapeptide KP. Laser Ther. 2019, 28, 180–186. [Google Scholar] [CrossRef]
- Vega-Chacón, Y.; de Albuquerque, M.C.; Pavarina, A.C.; Goldman, G.H.; Mima, E.G.D.O. Verapamil inhibits efflux pumps in Candida albicans, exhibits synergism with fluconazole, and increases survival of Galleria mellonella. Virulence 2021, 12, 231–243. [Google Scholar] [CrossRef]
- Nagargoje, A.A.; Akolkar, S.V.; Siddiqui, M.M.; Subhedar, D.D.; Sangshetti, J.N.; Khedkar, V.M.; Shingate, B.B. Quinoline based monocarbonyl curcumin analogs as potential antifungal and antioxidant agents: Synthesis, bioevaluation and molecular docking study. Chem. Biodivers. 2019, 17, 1900624. [Google Scholar] [CrossRef] [PubMed]
- Jennings, M.; Parks, R. Curcumin as an antiviral agent. Viruses 2020, 12, 1242. [Google Scholar] [CrossRef] [PubMed]
- Mathew, D.; Hsu, W.-L. Antiviral potential of curcumin. J. Funct. Foods 2018, 40, 692–699. [Google Scholar] [CrossRef]
- Teli, D.M.; Shah, M.B.; Chhabria, M.T. In silico screening of natural compounds as potential inhibitors of SARS-CoV-2 main protease and spike RBD: Targets for COVID-19. Front. Mol. Biosci. 2021, 7, 599079. [Google Scholar] [CrossRef]
- Jena, A.B.; Kanungo, N.; Nayak, V.; Chainy, G.B.N.; Dandapat, J. Catechin and curcumin interact with S protein of SARS-CoV2 and ACE2 of human cell membrane: Insights from computational studies. Sci. Rep. 2021, 11, 1–14. [Google Scholar] [CrossRef]
- Kumar, M.; Sodhi, K.K.; Singh, D.K. Addressing the potential role of curcumin in the prevention of COVID-19 by targeting the Nsp9 replicase protein through molecular docking. Arch. Microbiol. 2021, 203, 1691–1696. [Google Scholar] [CrossRef]
- Patel, A.; Rajendran, M.; Shah, A.; Patel, H.; Pakala, S.B.; Karyala, P. Virtual screening of curcumin and its analogs against the spike surface glycoprotein of SARS-CoV-2 and SARS-CoV. J. Biomol. Struct. Dyn. 2021, 5, 1–9. [Google Scholar] [CrossRef]
- Allam, L.; Ghrifi, F.; Mohammed, H.; El Hafidi, N.; El Jaoudi, R.; El Harti, J.; Lmimouni, B.; Belyamani, L.; Ibrahimi, A. Targeting the GRP78-dependant SARS-CoV-2 cell entry by peptides and small molecules. Bioinform. Biol. Insights 2020, 14, 1177932220965505. [Google Scholar] [CrossRef]
- Ibrahim, M.A.; Abdelrahman, A.H.; Hussien, T.A.; Badr, E.A.; Mohamed, T.A.; El-Seedi, H.R.; Pare, P.W.; Efferth, T.; Hegazy, M.-E.F. In silico drug discovery of major metabolites from spices as SARS-CoV-2 main protease inhibitors. Comput. Biol. Med. 2020, 126, 104046. [Google Scholar] [CrossRef]
- Kumar, S.; Kashyap, P.; Chowdhury, S.; Kumar, S.; Panwar, A.; Kumar, A. Identification of phytochemicals as potential therapeutic agents that binds to Nsp15 protein target of coronavirus (SARS-CoV-2) that are capable of inhibiting virus replication. Phytomedicine 2021, 85, 153317. [Google Scholar] [CrossRef]
- Maurya, V.K.; Kumar, S.; Prasad, A.K.; Bhatt, M.L.B.; Saxena, S.K. Structure-based drug designing for potential antiviral activity of selected natural products from Ayurveda against SARS-CoV-2 spike glycoprotein and its cellular receptor. Virusdisease 2020, 31, 179–193. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, X.; Omarini, A.B.; Li, B. Virtual screening for functional foods against the main protease of SARS-CoV-2. J. Food Biochem. 2020, 44, 13481. [Google Scholar] [CrossRef]
- Barh, D.; Tiwari, S.; Weener, M.E.; Azevedo, V.; Góes-Neto, A.; Gromiha, M.M.; Ghosh, P. Multi-omics-based identification of SARS-CoV-2 infection biology and candidate drugs against COVID-19. Comput. Biol. Med. 2020, 126, 104051. [Google Scholar] [CrossRef]
- Zheng, D.; Huang, C.; Huang, H.; Zhao, Y.; Khan, M.R.U.; Zhao, H.; Huang, L. Antibacterial mechanism of curcumin: A review. Chem. Biodivers. 2020, 17, 2000171. [Google Scholar] [CrossRef]
- Teow, S.-Y.; Liew, K.; Ali, S.A.; Khoo, A.S.-B.; Peh, S.-C. Antibacterial action of curcumin against Staphylococcus aureus: A brief review. J. Trop. Med. 2016, 2016, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Santezi, C.; Reina, B.D.; Dovigo, L.N. Curcumin-mediated photodynamic therapy for the treatment of oral infections—a review. Photodiagn. Photodyn. 2018, 21, 409–415. [Google Scholar] [CrossRef] [Green Version]
- Alves, F.; Pavarina, A.C.; Mima, E.G.D.O.; McHale, A.P.; Callan, J.F. Antimicrobial sonodynamic and photodynamic therapies against Candida albicans. Biofouling 2018, 34, 357–367. [Google Scholar] [CrossRef] [Green Version]
- Forouzanfar, F.; Forouzanfar, A.; Sathyapalan, T.; Orafai, H.M.; Sahebkar, A. Curcumin for the management of periodontal diseases: A review. Curr. Pharm. Des. 2020, 26, 4277–4284. [Google Scholar] [CrossRef]
- Cheraghipour, K.; Ezatpour, B.; Masoori, L.; Marzban, A.; Sepahvand, A.; Rouzbahani, A.K.; Moridnia, A.; Khanizadeh, S.; Mahmoudvand, H. Anti-Candida activity of curcumin: A systematic review. Curr. Drug Discov. Technol. 2021, 18, 379–390. [Google Scholar] [CrossRef]
- Kataoka, K.; Harada, A.; Nagasaki, Y. Block copolymer micelles for drug delivery: Design, characterization and biological significance. Adv. Drug Deliv. Rev. 2001, 47, 113–131. [Google Scholar] [CrossRef]
- Akbar, M.U.; Zia, K.M.; Nazir, A.; Iqbal, J.; Ejaz, S.A.; Akash, M.S.H. Pluronic-based mixed polymeric micelles enhance the therapeutic potential of curcumin. Aaps Pharmscitech 2018, 19, 2719–2739. [Google Scholar] [CrossRef]
- Huang, F.; Gao, Y.; Zhang, Y.; Cheng, T.; Ou, H.; Yang, L.; Liu, J.; Shi, L.; Liu, J. Silver-decorated polymeric micelles combined with curcumin for enhanced antibacterial activity. ACS Appl. Mater. Interfaces 2017, 9, 16880–16889. [Google Scholar] [CrossRef]
- Takrami, S.R.; Ranji, N.; Sadeghizadeh, M. Antibacterial effects of curcumin encapsulated in nanoparticles on clinical isolates of Pseudomonas aeruginosa through downregulation of efflux pumps. Mol. Biol. Rep. 2019, 46, 2395–2404. [Google Scholar] [CrossRef]
- Teng, F.; Deng, P.; Song, Z.; Zhou, F.; Feng, R. Enhanced effect in combination of curcumin- and ketoconazole-loaded methoxy poly (ethylene glycol)-poly (ε-caprolactone) micelles. Biomed. Pharm. 2017, 88, 43–51. [Google Scholar] [CrossRef]
- Pourhajibaghera, M.; Esboeibc, B.; Hodjata, M.; Bahadord, A. Sonodynamic excitation of nanomicelle curcumin for eradication of Streptococcus mutans under sonodynamic antimicrobial chemotherapy: Enhanced anti-caries activity of nanomicelle curcumin. Photodiagn. Photodyn. 2020, 30, 101780. [Google Scholar] [CrossRef]
- Rupel, K.; Zupin, L.; Brich, S.; Mardirossian, M.; Ottaviani, G.; Gobbo, M.; Di Lenarda, R.; Pricl, S.; Crovella, S.; Zacchigna, S.; et al. Antimicrobial activity of amphiphilic nanomicelles loaded with curcumin against Pseudomonas aeruginosa alone and activated by blue laser light. J. Biophotonics 2021, 14, 202000350. [Google Scholar] [CrossRef]
- Dias, V.H.C.; Malacrida, A.M.; dos Santos, A.R.; Batista, A.F.P.; Campanerut-Sá, P.A.Z.; Braga, G.; Bona, E.; Caetano, W.; Mikcha, J.M.G. pH interferes in photoinhibitory activity of curcumin nanoencapsulated with pluronic® P123 against Staphylococcus aureus. Photodiagn. Photodyn. 2021, 33, 102085. [Google Scholar] [CrossRef]
- Zhu, Y.; Luo, Q.; Zhang, H.; Cai, Q.; Li, X.; Shen, Z.; Zhu, W. A shear-thinning electrostatic hydrogel with antibacterial activity by nanoengineering of polyelectrolytes. Biomater. Sci. 2020, 8, 1394–1404. [Google Scholar] [CrossRef] [PubMed]
- Barros, C.H.N.; Hiebner, D.W.; Fulaz, S.; Vitale, S.; Quinn, L.; Casey, E. Synthesis and self-assembly of curcumin-modified amphiphilic polymeric micelles with antibacterial activity. J. Nanobiotechnol. 2021, 19, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Valizadeh, H.; Abdolmohammadi-Vahid, S.; Danshina, S.; Gencer, M.Z.; Ammari, A.; Sadeghi, A.; Roshangar, L.; Aslani, S.; Esmaeilzadeh, A.; Ghaebi, M.; et al. Nano-curcumin therapy, a promising method in modulating inflammatory cytokines in COVID-19 patients. Int. Immunopharmacol. 2020, 89, 107088. [Google Scholar] [CrossRef] [PubMed]
- Aditya, N.; Chimote, G.; Gunalan, K.; Banerjee, R.; Patankar, S.; Madhusudhan, B. Curcuminoids-loaded liposomes in combination with arteether protects against Plasmodium berghei infection in mice. Exp. Parasitol. 2012, 131, 292–299. [Google Scholar] [CrossRef]
- Shome, S.; Das Talukdar, A.; Choudhury, M.D.; Bhattacharya, M.K.; Upadhyaya, H. Curcumin as potential therapeutic natural product: A nanobiotechnological perspective. J. Pharm. Pharm. 2016, 68, 1481–1500. [Google Scholar] [CrossRef]
- Chen, Y.; Lu, Y.; Lee, R.J.; Xiang, G. Nano encapsulated curcumin: And its potential for biomedical applications. Int. J. Nanomed. 2020, 15, 3099–3120. [Google Scholar] [CrossRef]
- Ding, T.; Li, T.; Wang, Z.; Li, J. Curcumin liposomes interfere with quorum sensing system of Aeromonas sobria and in silico analysis. Sci. Rep. 2017, 7, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Mittal, A.; Kumar, N.; Chauhan, N.S. Curcumin encapsulated PEGylated nanoliposomes: A potential anti-infective therapeutic agent. Indian J. Microbiol. 2019, 59, 336–343. [Google Scholar] [CrossRef]
- Battista, S.; Maggi, M.A.; Bellio, P.; Galantini, L.; D’Archivio, A.A.; Celenza, G.; Colaiezzi, R.; Giansanti, L. Curcuminoids-loaded liposomes: Influence of lipid composition on their physicochemical properties and efficacy as delivery systems. Colloids Surf. A Phys. Eng. Asp. 2020, 597, 124759. [Google Scholar] [CrossRef]
- Gao, M.; Long, X.; Du, J.; Teng, M.; Zhang, W.; Wang, Y.; Wang, X.; Wang, Z.; Zhang, P.; Li, J. Enhanced curcumin solubility and antibacterial activity by encapsulation in PLGA oily core nanocapsules. Food Funct. 2020, 11, 448–455. [Google Scholar] [CrossRef]
- Bhatia, E.; Sharma, S.; Jadhav, K.; Banerjee, R. Combinatorial liposomes of berberine and curcumin inhibit biofilm formation and intracellular methicillin resistant Staphylococcus aureus infections and associated inflammation. J. Mater. Chem. B 2021, 9, 864–875. [Google Scholar] [CrossRef]
- Hazzah, H.A.; Farid, R.M.; Nasra, M.M.; Hazzah, W.A.; El-Massik, M.A.; Abdallah, O.Y. Gelucire-based nanoparticles for curcumin targeting to oral mucosa: Preparation, characterization, and antimicrobial activity Assessment. J. Pharm. Sci. 2015, 104, 3913–3924. [Google Scholar] [CrossRef]
- Isacchi, B.; Bergonzi, M.C.; Grazioso, M.; Righeschi, C.; Pietretti, A.; Severini, C.; Bilia, A.R. Artemisinin and artemisinin plus curcumin liposomal formulations: Enhanced antimalarial efficacy against Plasmodium berghei-infected mice. Eur. J. Pharm. Biopharm. 2012, 80, 528–534. [Google Scholar] [CrossRef]
- Coma-Cros, E.M.; Biosca, A.; Lantero, E.; Manca, M.L.; Caddeo, C.; Gutiérrez, L.; Ramírez, M.; Borgheti-Cardoso, L.N.; Manconi, M.; Fernàndez-Busquets, X. Antimalarial activity of orally administered curcumin incorporated in Eudragit®-containing liposomes. Int. J. Mol. Sci. 2018, 19, 1361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, M.; Su, X.; Ding, B.; He, X.; Liu, X.; Yu, A.; Lou, H.; Zhai, G. Advances in nanotechnology-based delivery systems for curcumin. Nanomedicine 2012, 7, 1085–1100. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Eral, H.B.; Hatton, T.A.; Doyle, P.S. Nanoemulsions: Formation, properties and applications. Soft Matter 2016, 12, 2826–2841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flora, G.; Gupta, D.; Tiwari, A. Nanocurcumin: A promising therapeutic advancement over native curcumin. Crit. Rev. Drug Carr. Syst. 2013, 30, 331–368. [Google Scholar] [CrossRef]
- Mirani, A.; Kundaikar, H.; Velhal, S.; Patel, V.; Bandivdekar, A.; Degani, M.; Patravale, V. Tetrahydrocurcumin-loaded vaginal nanomicrobicide for prophylaxis of HIV/AIDS: In silico study, formulation development, and in vitro evaluation. Drug Deliv. Transl. Res. 2019, 9, 828–847. [Google Scholar] [CrossRef]
- Bonfim, C.M.D.; Monteleoni, L.F.; Calmon, M.D.F.; Cândido, N.M.; Provazzi, P.J.S.; Lino, V.D.S.; Rabachini, T.; Sichero, L.; Villa, L.L.; Quintana, S.M.; et al. Antiviral activity of curcumin-nanoemulsion associated with photodynamic therapy in vulvar cell lines transducing different variants of HPV-16. Artif. Cells Nanomed. Biotechnol. 2020, 48, 515–524. [Google Scholar] [CrossRef] [Green Version]
- Nabila, N.; Suada, N.K.; Denis, D.; Yohan, B.; Adi, A.C.; Veterini, A.S.; Anindya, A.L.; Sasmono, R.T.; Rachmawati, H. Antiviral action of curcumin encapsulated in nanoemulsion against four serotypes of dengue virus. Pharm. Nanotechnol. 2020, 8, 54–62. [Google Scholar] [CrossRef]
- Kaur, A.; Saxena, Y.; Bansal, R.; Gupta, S.; Tyagi, A.; Sharma, R.K.; Ali, J.; Panda, A.K.; Gabrani, R.; Dang, S. Intravaginal delivery of polyphenon 60 and curcumin nanoemulsion gel. Aaps Pharmscitech 2017, 18, 2188–2202. [Google Scholar] [CrossRef]
- Ngwabebhoh, F.A.; Erdagi, S.I.; Yildiz, U. Pickering emulsions stabilized nanocellulosic-based nanoparticles for coumarin and curcumin nanoencapsulations: In vitro release, anticancer and antimicrobial activities. Carbohydr. Polym. 2018, 201, 317–328. [Google Scholar] [CrossRef]
- Khan, M.; Ali, M.; Shah, W.; Shah, A.; Yasinzai, M.M. Curcumin-loaded self-emulsifying drug delivery system (cu-SEDDS): A promising approach for the control of primary pathogen and secondary bacterial infections in cutaneous leishmaniasis. Appl. Microbiol. Biotechnol. 2019, 103, 7481–7490. [Google Scholar] [CrossRef]
- Hashim, A.F.; Hamed, S.; Hamid, H.A.A.; Abd-Elsalam, K.A.; Golonka, I.; Musiał, W.; El-Sherbiny, I.M. Antioxidant and antibacterial activities of omega-3 rich oils/curcumin nanoemulsions loaded in chitosan and alginate-based microbeads. Int. J. Biol. Macromol. 2019, 140, 682–696. [Google Scholar] [CrossRef]
- Chen, K.; Qian, Y.; Wu, S.; Qiu, X.; Yang, D.; Lei, L. Neutral fabrication of UV-blocking and antioxidation lignin-stabilized high internal phase emulsion encapsulates for high efficient antibacterium of natural curcumin. Food Funct. 2019, 10, 3543–3555. [Google Scholar] [CrossRef]
- Lewińska, A.; Jaromin, A.; Jezierska, J. Role of architecture of N-oxide surfactants in the design of nanoemulsions for Candida skin infection. Colloids Surf. B Biointerfaces 2020, 187, 110639. [Google Scholar] [CrossRef]
- Lakkakula, J.R.; Krause, R.W.M. A vision for cyclodextrin nanoparticles in drug delivery systems and pharmaceutical applications. Nanomedicine 2014, 9, 877–894. [Google Scholar] [CrossRef]
- Macaev, F.; Boldescu, V.; Geronikaki, A.; Sucman, N. Recent advances in the use of cyclodextrins in antifungal formulations. Curr. Top. Med. Chem. 2013, 13, 2677–2683. [Google Scholar] [CrossRef]
- Kurita, T.; Makino, Y. Novel curcumin oral delivery systems. Anticancer Res. 2013, 33, 2807–2821. [Google Scholar]
- Del Valle, E. Cyclodextrins and their uses: A review. Process. Biochem. 2004, 39, 1033–1046. [Google Scholar] [CrossRef]
- Atwood, J.L. Comprehensive Supramolecular Chemistry; Pergamon: New York, NY, USA, 1996; Volume 3. [Google Scholar]
- Saokham, P.; Muankaew, C.; Jansook, P.; Loftsson, T. Solubility of cyclodextrins and drug/cyclodextrin complexes. Molecules 2018, 23, 1161. [Google Scholar] [CrossRef] [Green Version]
- Davis, M.E.; Brewster, M.E. Cyclodextrin-based pharmaceutics: Past, present and future. Nat. Rev. Drug Discov. 2004, 3, 1023–1035. [Google Scholar] [CrossRef]
- Smith, A.J.; Oertle, J.; Prato, D. Multiple actions of curcumin including anticancer, anti-inflammatory, antimicrobial and enhancement via cyclodextrin. J. Cancer Ther. 2015, 06, 257–272. [Google Scholar] [CrossRef] [Green Version]
- Hegge, A.B.; Nielsen, T.T.; Larsen, K.L.; Bruzell, E.; Tønnesen, H.H. Impact of curcumin supersaturation in antibacterial photodynamic therapy—effect of cyclodextrin type and amount: Studies on curcumin and curcuminoides XLV. J. Pharm. Sci. 2012, 101, 1524–1537. [Google Scholar] [CrossRef]
- Hegge, A.B.; Vukicevic, M.; Bruzell, E.; Kristensen, S.; Tønnesen, H. Solid dispersions for preparation of phototoxic supersaturated solutions for antimicrobial photodynamic therapy (aPDT). Eur. J. Pharm. Biopharm. 2013, 83, 95–105. [Google Scholar] [CrossRef]
- Wikene, K.; Hegge, A.B.; Bruzell, E.; Tønnesen, H.H. Formulation and characterization of lyophilized curcumin solid dispersions for antimicrobial photodynamic therapy (aPDT): Studies on curcumin and curcuminoids LII. Drug Dev. Ind. Pharm. 2014, 41, 969–977. [Google Scholar] [CrossRef] [Green Version]
- Shlar, I.; Droby, S.; Rodov, V. Antimicrobial coatings on polyethylene terephthalate based on curcumin/cyclodextrin complex embedded in a multilayer polyelectrolyte architecture. Colloids Surf. B Biointerfaces 2018, 164, 379–387. [Google Scholar] [CrossRef]
- Gupta, A.; Keddie, D.; Kannappan, V.; Gibson, H.; Khalil, I.; Kowalczuk, M.; Martin, C.; Shuai, X.; Radecka, I. Production and characterisation of bacterial cellulose hydrogels loaded with curcumin encapsulated in cyclodextrins as wound dressings. Eur. Polym. J. 2019, 118, 437–450. [Google Scholar] [CrossRef]
- Reddy, D.N.K.; Kumar, R.; Wang, S.-P.; Huang, F.-Y. Curcumin-C3 complexed with α-, β-cyclodextrin exhibits antibacterial and antioxidant properties suitable for cancer treatments. Curr. Drug Metab. 2020, 20, 988–1001. [Google Scholar] [CrossRef]
- Alizadeh, N.; Malakzadeh, S. Antioxidant, antibacterial and anti-cancer activities of β-and γ-CDs/curcumin loaded in chitosan nanoparticles. Int. J. Biol. Macromol. 2020, 147, 778–791. [Google Scholar] [CrossRef]
- Brasch, J.; Beck-Jendroschek, V.; Walther, G.; Rubbel, D. Clinical isolates of Trichophyton rubrum are completely inhibited by photochemical treatment with a γ-cyclodextrin formulation of curcuminoids. Mycoses 2020, 63, 369–375. [Google Scholar] [CrossRef] [PubMed]
- Stasiłowicz, A.; Tykarska, E.; Lewandowska, K.; Kozak, M.; Miklaszewski, A.; Kobus-Cisowska, J.; Szymanowska, D.; Plech, T.; Jenczyk, J.; Cielecka-Piontek, J. Hydroxypropyl-β-cyclodextrin as an effective carrier of curcumin-piperine nutraceutical system with improved enzyme inhibition properties. J. Enzym. Inhib. Med. Chem. 2020, 35, 1811–1821. [Google Scholar] [CrossRef] [PubMed]
- Shlar, I.; Droby, S.; Choudhary, R.; Rodov, V. The mode of antimicrobial action of curcumin depends on the delivery system: Monolithic nanoparticles vs. supramolecular inclusion complex. Rsc Adv. 2017, 7, 42559–42569. [Google Scholar] [CrossRef] [Green Version]
- Szejtli, J. Past, present and futute of cyclodextrin research. Pure Appl. Chem. 2004, 76, 1825–1845. [Google Scholar] [CrossRef] [Green Version]
- Saheb, M.; Fereydouni, N.; Nemati, S.; Barreto, G.E.; Johnston, T.P.; Sahebkar, A. Chitosan-based delivery systems for curcumin: A review of pharmacodynamic and pharmacokinetic aspects. J. Cell. Physiol. 2019, 234, 12325–12340. [Google Scholar] [CrossRef]
- Pellá, M.C.; Lima-Tenório, M.K.; Tenório-Neto, E.; Guilherme, M.R.; Muniz, E.C.; Rubira, A.F. Chitosan-based hydrogels: From preparation to biomedical applications. Carbohydr. Polym. 2018, 196, 233–245. [Google Scholar] [CrossRef]
- Rinaudo, M. Chitin and chitosan: Properties and applications. Prog. Polym. Sci. 2006, 31, 603–632. [Google Scholar] [CrossRef]
- Chung, Y.-C.; Su, Y.-P.; Chen, C.-C.; Jia, G.; Wang, H.-L.; Wu, J.C.G.; Lin, J.-G. Relationship between antibacterial activity of chitosan and surface characteristics of cell wall. Acta Pharm. Sin. 2004, 25, 932–936. [Google Scholar]
- Krausz, A.E.; Adler, B.L.; Cabral, V.; Navati, M.; Doerner, J.; Charafeddine, R.; Chandra, D.; Liang, H.; Gunther, L.; Clendaniel, A.; et al. Curcumin-encapsulated nanoparticles as innovative antimicrobial and wound healing agent. Nanomed. Nanotechnol. Biol. Med. 2015, 11, 195–206. [Google Scholar] [CrossRef] [Green Version]
- Saranya, T.; Raja, B.; Biswas, R.; Jayakumar, R.; Sathianarayanan, S. Synthesis, characterisation and biomedical applications of curcumin conjugated chitosan microspheres. Int. J. Biol. Macromol. 2018, 110, 227–233. [Google Scholar] [CrossRef]
- Maghsoudi, A.; Yazdian, F.; Shahmoradi, S.; Ghaderi, L.; Hemati, M.; Amoabediny, G. Curcumin-loaded polysaccharide nanoparticles: Optimization and anticariogenic activity against Streptococcus mutans. Mater. Sci. Eng. C 2017, 75, 1259–1267. [Google Scholar] [CrossRef] [Green Version]
- Jahanizadeh, S.; Yazdian, F.; Marjani, A.; Omidi, M.; Rashedi, H. Curcumin-loaded chitosan/carboxymethyl starch/montmorillonite bio-nanocomposite for reduction of dental bacterial biofilm formation. Int. J. Biol. Macromol. 2017, 105, 757–763. [Google Scholar] [CrossRef]
- Zhao, Y.; Liu, J.; Chen, W.; Yu, A. Efficacy of thermosensitive chitosan/β-glycerophosphate hydrogel loaded with β-cyclodextrin-curcumin for the treatment of cutaneous wound infection in rats. Exp. Med. 2017, 15, 1304–1313. [Google Scholar] [CrossRef] [Green Version]
- Niranjan, R.; Kaushik, M.; Prakash, J.; Venkataprasanna, K.S.; Arpana, C.; Balashanmugam, P.; Venkatasubbu, G.D. Enhanced wound healing by PVA/chitosan/curcumin patches: In vitro and in vivo study. Colloids Surf. B Biointerfaces 2019, 182, 110339. [Google Scholar] [CrossRef]
- Abbas, M.; Hussain, T.; Arshad, M.; Ansari, A.R.; Irshad, A.; Nisar, J.; Hussain, F.; Masood, N.; Nazir, A.; Iqbal, M. Wound healing potential of curcumin cross-linked chitosan/polyvinyl alcohol. Int. J. Biol. Macromol. 2019, 140, 871–876. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.; Moser, D.; Han, F.; Leonhard, M.; Schneider-Stickler, B.; Tan, Y. Preparation and antibiofilm studies of curcumin loaded chitosan nanoparticles against polymicrobial biofilms of Candida albicans and Staphylococcus aureus. Carbohydr. Polym. 2020, 241, 116254. [Google Scholar] [CrossRef] [PubMed]
- Loutfy, S.A.; Elberry, M.H.; Farroh, K.Y.; Mohamed, H.T.; Mohamed, A.A.; Mohamed, E.B.; Faraag, A.H.I.; Mousa, S.A. Antiviral activity of chitosan nanoparticles encapsulating curcumin against Hepatitis C virus genotype 4a in human hepatoma cell lines. Int. J. Nanomed. 2020, 15, 2699–2715. [Google Scholar] [CrossRef] [Green Version]
- Taha, S.H.; El-Sherbiny, I.M.; Salem, A.S.; Abdel-Hamid, M.; Hamed, A.H.; Ahmed, G.A. Antiviral activity of curcumin loaded milk proteins nanoparticles on Potato virus Y. Pak. J. Biol. Sci. 2019, 22, 614–622. [Google Scholar] [CrossRef]
- Hegge, A.B.; Andersen, T.; Melvik, J.; Bruzell, E.; Kristensen, S.; Tønnesen, H. Formulation and bacterial phototoxicity of curcumin loaded alginate foams for wound treatment applications: Studies on curcumin and curcuminoides XLII. J. Pharm. Sci. 2011, 100, 174–185. [Google Scholar] [CrossRef]
- Bhawana Basniwal, R.K.; Buttar, H.S.; Jain, V.K.; Jain, N. Curcumin nanoparticles: Preparation, characterization, and antimicrobial study. J. Agric. Food Chem. 2011, 59, 2056–2061. [Google Scholar] [CrossRef]
- Adahoun, M.A.; Al-Akhras, M.-A.; Jaafar, M.S.; Bououdina, M. Enhanced anti-cancer and antimicrobial activities of curcumin nanoparticles. Artif. Cells Nanomed. Biotechnol. 2016, 45, 98–107. [Google Scholar] [CrossRef]
- Shlar, I.; Poverenov, E.; Vinokur, Y.; Horev, B.; Droby, S.; Rodov, V. High-throughput screening of nanoparticle-stabilizing ligands: Application to preparing antimicrobial curcumin nanoparticles by antisolvent precipitation. Nano-Micro Lett. 2015, 7, 68–79. [Google Scholar] [CrossRef] [Green Version]
- No, D.S.; AlGburi, A.; Huynh, P.; Moret, A.; Ringard, M.; Comito, N.; Drider, D.; Takhistov, P.; Chikindas, M.L. Antimicrobial efficacy of curcumin nanoparticles against Listeria monocytogenes is mediated by surface charge. J. Food Saf. 2017, 37, e12353. [Google Scholar] [CrossRef]
- Gutierrez, J.K.T.; Zanatta, G.C.; Ortega, A.L.M.; Balastegui, M.I.C.; Sanitá, P.V.; Pavarina, A.C.; Barbugli, P.; Mima, E.G.D.O. Encapsulation of curcumin in polymeric nanoparticles for antimicrobial photodynamic therapy. PLoS ONE 2017, 12, e0187418. [Google Scholar] [CrossRef] [Green Version]
- Sakima, V.T.; Barbugli, P.A.; Cerri, P.S.; Chorilli, M.; Carmello, J.C.; Pavarina, A.C.; Mima, E.G.D.O. Antimicrobial photodynamic therapy mediated by curcumin-loaded polymeric nanoparticles in a murine model of oral candidiasis. Molecules 2018, 23, 2075. [Google Scholar] [CrossRef] [Green Version]
- Shariati, A.; Asadian, E.; Fallah, F.; Azimi, T.; Hashemi, A.; Sharahi, J.Y.; Moghadam, M.T. Evaluation of nano-curcumin effects on expression levels of virulence genes and biofilm production of multidrug-resistant Pseudomonas aeruginosa isolated from burn wound infection in Tehran, Iran. Infect. Drug Resist. 2019, 12, 2223–2235. [Google Scholar] [CrossRef] [Green Version]
- Preis, E.; Baghdan, E.; Agel, M.R.; Anders, T.; Pourasghar, M.; Schneider, M.; Bakowsky, U. Spray dried curcumin loaded nanoparticles for antimicrobial photodynamic therapy. Eur. J. Pharm. Biopharm. 2019, 142, 531–539. [Google Scholar] [CrossRef]
- Jaguezeski, A.M.; Souza, C.F.; Perin, G.; Reis, J.H.; Gomes, T.M.; Baldissera, M.D.; Vaucher, R.A.; de Andrade, C.M.; Stefani, L.M.; Gundel, S.S.; et al. Effect of free and nano-encapsulated curcumin on treatment and energetic metabolism of gerbils infected by Listeria monocytogenes. Microb. Pathog. 2019, 134, 103564. [Google Scholar] [CrossRef]
- Pourhajibagher, M.; Omrani, L.R.; Noroozian, M.; Ghorbanzadeh, Z.; Bahador, A. In vitro antibacterial activity and durability of a nano-curcumin-containing pulp capping agent combined with antimicrobial photodynamic therapy. Photodiagn. Photodyn. 2021, 33, 102150. [Google Scholar] [CrossRef]
- Pourhajibagher, M.; Plotino, G.; Chiniforush, N.; Bahador, A. Dual wavelength irradiation antimicrobial photodynamic therapy using indocyanine green and metformin doped with nano-curcumin as an efficient adjunctive endodontic treatment modality. Photodiagn. Photodyn. 2020, 29, 101628. [Google Scholar] [CrossRef]
- Chen, L.; Song, Z.; Zhi, X.; Du, B. Photoinduced antimicrobial activity of curcumin-containing coatings: Molecular interaction, stability and potential application in food decontamination. ACS Omega 2020, 5, 31044–31054. [Google Scholar] [CrossRef]
- Ahmadi, H.; Haddadi-Asl, V.; Ghafari, H.-A.; Ghorbanzadeh, R.; Mazlum, Y.; Bahador, A. Shear bond strength, adhesive remnant index, and anti-biofilm effects of a photoexcited modified orthodontic adhesive containing curcumin doped poly lactic-co-glycolic acid nanoparticles: An ex-vivo biofilm model of S. mutans on the enamel slab bonded brackets. Photodiagn. Photodyn. 2020, 30, 101674. [Google Scholar] [CrossRef]
- Kiti, K.; Suwantong, O. The potential use of curcumin-β-cyclodextrin inclusion complex/chitosan-loaded cellulose sponges for the treatment of chronic wound. Int. J. Biol. Macromol. 2020, 164, 3250–3258. [Google Scholar] [CrossRef]
- Shakeri, A.; Panahi, Y.; Johnston, T.P.; Sahebkar, A. Biological properties of metal complexes of curcumin. BioFactors 2019, 45, 304–317. [Google Scholar] [CrossRef]
- Bajpai, S.; Ahuja, S.; Chand, N.; Bajpai, M. Nano cellulose dispersed chitosan film with Ag NPs/Curcumin: An in vivo study on albino rats for wound dressing. Int. J. Biol. Macromol. 2017, 104, 1012–1019. [Google Scholar] [CrossRef]
- De Freitas, C.F.; Kimura, E.; Rubira, A.F.; Muniz, E.C. Curcumin and silver nanoparticles carried out from polysaccharide-based hydrogels improved the photodynamic properties of curcumin through metal-enhanced singlet oxygen effect. Mater. Sci. Eng. C 2020, 112, 110853. [Google Scholar] [CrossRef]
- Durán, N.; Durán, M.; de Jesus, M.B.; Seabra, A.B.; Fávaro, W.J.; Nakazato, G. Silver nanoparticles: A new view on mechanistic aspects on antimicrobial activity. Nanomed. Nanotechnol. Biol. Med. 2016, 12, 789–799. [Google Scholar] [CrossRef]
- Jaiswal, S.; Mishra, P. Antimicrobial and antibiofilm activity of curcumin-silver nanoparticles with improved stability and selective toxicity to bacteria over mammalian cells. Med. Microbiol. Immunol. 2017, 207, 39–53. [Google Scholar] [CrossRef]
- El Khoury, E.; Abiad, M.; Kassaify, Z.G.; Patra, D. Green synthesis of curcumin conjugated nanosilver for the applications in nucleic acid sensing and anti-bacterial activity. Colloids Surf. B Biointerfaces 2015, 127, 274–280. [Google Scholar] [CrossRef]
- Loo, J.C.Y.; Rohanizadeh, R.; Young, P.M.; Traini, D.; Cavaliere, R.; Whitchurch, C.B.; Lee, W.H. Combination of silver nanoparticles and curcumin nanoparticles for enhanced anti-biofilm activities. J. Agric. Food Chem. 2016, 64, 2513–2522. [Google Scholar] [CrossRef] [PubMed]
- Abdellah, A.M.; Sliem, M.A.; Bakr, M.; Amin, R.M. Green synthesis and biological activity of silver-curcumin nanoconjugates. Futur. Med. Chem. 2018, 10, 2577–2588. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.; Wu, Y.; Wang, H.; Han, H. Synergistic antibacterial effects of curcumin modified silver nanoparticles through ROS-mediated pathways. Mater. Sci. Eng. C 2019, 99, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, P.; Shukla, M.; Kaul, G.; Chopra, S.; Patra, A.K. Rationally designed curcumin based ruthenium(ii) antimicrobials effective against drug-resistant Staphylococcus aureus. Dalton Trans. 2019, 48, 11822–11828. [Google Scholar] [CrossRef] [PubMed]
- Varaprasad, K.; Vimala, K.; Ravindra, S.; Reddy, N.N.; Reddy, G.V.S.; Raju, K.M. Fabrication of silver nanocomposite films impregnated with curcumin for superior antibacterial applications. J. Mater. Sci. Mater. Med. 2011, 22, 1863–1872. [Google Scholar] [CrossRef]
- Srivatsan, K.V.; Duraipandy, N.; Begum, S.; Lakra, R.; Ramamurthy, U.; Korrapati, P.S.; Kiran, M.S. Effect of curcumin caged silver nanoparticle on collagen stabilization for biomedical applications. Int. J. Biol. Macromol. 2015, 75, 306–315. [Google Scholar] [CrossRef]
- Anjum, S.; Gupta, A.; Sharma, D.; Gautam, D.; Bhan, S.; Sharma, A.; Kapil, A.; Gupta, B. Development of novel wound care systems based on nanosilver nanohydrogels of polymethacrylic acid with Aloe vera and curcumin. Mater. Sci. Eng. C 2016, 64, 157–166. [Google Scholar] [CrossRef]
- Soumya, K.R.; Snigdha, S.; Sugathan, S.; Mathew, J.; Radhakrishnan, E.K. Zinc oxide–curcumin nanocomposite loaded collagen membrane as an effective material against methicillin-resistant coagulase-negative Staphylococci. 3 Biotech 2017, 7, 238. [Google Scholar] [CrossRef]
- Alves, T.; Chaud, M.; Grotto, D.; Jozala, A.F.; Pandit, R.; Rai, M.; Dos Santos, C.A. Association of silver nanoparticles and curcumin solid dispersion: Antimicrobial and antioxidant properties. Aaps Pharmscitech 2017, 19, 225–231. [Google Scholar] [CrossRef]
- Paul, S.; Mohanram, K.; Kannan, I. Antifungal activity of curcumin-silver nanoparticles against fluconazole-resistant clinical isolates of Candida species. AYU 2018, 39, 182–186. [Google Scholar] [CrossRef]
- Khanh, L.L.; Truc, N.T.; Dat, N.T.; Nghi, N.T.P.; van Toi, V.; Hoai, N.T.T.; Quyen, T.N.; Loan, T.T.T.; Hiep, N.T. Gelatin-stabilized composites of silver nanoparticles and curcumin: Characterization, antibacterial and antioxidant study. Sci. Technol. Adv. Mater. 2019, 20, 276–290. [Google Scholar] [CrossRef] [Green Version]
- Anagha, B.; George, D.; Maheswari, P.U.; Begum, K.M.M.S. Biomass derived antimicrobial hybrid cellulose hydrogel with green ZnO nanoparticles for curcumin delivery and its kinetic modelling. J. Polym. Environ. 2019, 27, 2054–2067. [Google Scholar] [CrossRef]
- George, D.; Maheswari, P.U.; Begum, K.M.M.S.; Arthanareeswaran, G.; Arthanaresswarn, G. Biomass-derived dialdehyde cellulose cross-linked chitosan-based nanocomposite hydrogel with phytosynthesized zinc oxide nanoparticles for enhanced curcumin delivery and bioactivity. J. Agric. Food Chem. 2019, 67, 10880–10890. [Google Scholar] [CrossRef]
- Masoule, S.F.; Pourhajibagher, M.; Safari, J.; Khoobi, M. Base-free green synthesis of copper(II) oxide nanoparticles using highly cross-linked poly(curcumin) nanospheres: Synergistically improved antimicrobial activity. Res. Chem. Intermed. 2019, 45, 4449–4462. [Google Scholar] [CrossRef]
- Lyu, Y.; Yu, M.; Liu, Q.; Zhang, Q.; Liu, Z.; Tian, Y.; Li, D.; Mu, C. Synthesis of silver nanoparticles using oxidized amylose and combination with curcumin for enhanced antibacterial activity. Carbohydr. Polym. 2020, 230, 115573. [Google Scholar] [CrossRef]
- Gupta, A.; Briffa, S.M.; Swingler, S.; Gibson, H.; Kannappan, V.; Adamus, G.; Kowalczuk, M.M.; Martin, C.; Radecka, I. Synthesis of silver nanoparticles using curcumin-cyclodextrins loaded into bacterial cellulose-based hydrogels for wound dressing applications. Biomacromolecules 2020, 21, 1802–1811. [Google Scholar] [CrossRef]
- El-Nahhal, I.M.; Salem, J.; Anbar, R.; Kodeh, F.S.; Elmanama, A. Preparation and antimicrobial activity of ZnO-NPs coated cotton/starch and their functionalized ZnO-Ag/cotton and Zn(II) curcumin/cotton materials. Sci. Rep. 2020, 10, 5410. [Google Scholar] [CrossRef]
- Ghaffari, S.-B.; Sarrafzadeh, M.-H.; Salami, M.; Khorramizadeh, M. A pH-sensitive delivery system based on N-succinyl chitosan-ZnO nanoparticles for improving antibacterial and anticancer activities of curcumin. Int. J. Biol. Macromol. 2020, 151, 428–440. [Google Scholar] [CrossRef]
- Marulasiddeshwara, R.; Jyothi, M.; Soontarapa, K.; Keri, R.S.; Velmurugan, R. Nonwoven fabric supported, chitosan membrane anchored with curcumin/TiO2 complex: Scaffolds for MRSA infected wound skin reconstruction. Int. J. Biol. Macromol. 2020, 144, 85–93. [Google Scholar] [CrossRef]
- Muniyappan, N.; Pandeeswaran, M.; Amalraj, A. Green synthesis of gold nanoparticles using Curcuma pseudomontana isolated curcumin: Its characterization, antimicrobial, antioxidant and anti- inflammatory activities. Environ. Chem. Ecotoxicol. 2021, 3, 117–124. [Google Scholar] [CrossRef]
- Davis, M.E. Ordered porous materials for emerging applications. Nat. Cell Biol. 2002, 417, 813–821. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.-Y.; Chen, L.-H.; Li, Y.; Rooke, J.C.; Sanchez, C.; Su, B.-L. Hierarchically porous materials: Synthesis strategies and structure design. Chem. Soc. Rev. 2017, 46, 481–558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agostini, A.; Sancenón, F.; Martínez-Máñez, R.; Marcos, M.D.; Soto, J.; Amorós, P. A photoactivated molecular gate. Chem. A Eur. J. 2012, 18, 12218–12221. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhao, Q.; Han, N.; Bai, L.; Li, J.; Che, E.; Hu, L.; Zhang, Q.; Jiang, T.; Wang, S. Mesoporous silica nanoparticles in drug delivery and biomedical applications. Nanomed. Nanotechnol. Biol. Med. 2015, 11, 313–327. [Google Scholar] [CrossRef] [PubMed]
- Vallet-Regí, M.; Balas, F.; Arcos, D. Mesoporous materials for drug delivery. Angew. Chem. Int. Ed. 2007, 46, 7548–7558. [Google Scholar] [CrossRef]
- Kuthati, Y.; Kankala, R.K.; Busa, P.; Lin, S.-X.; Deng, J.-P.; Mou, C.-Y.; Lee, C.-H. Phototherapeutic spectrum expansion through synergistic effect of mesoporous silica trio-nanohybrids against antibiotic-resistant gram-negative bacterium. J. Photochem. Photobiol. B Biol. 2017, 169, 124–133. [Google Scholar] [CrossRef]
- Wu, C.; Zhu, Y.; Wu, T.; Wang, L.; Yuan, Y.; Chen, J.; Hu, Y.; Pang, J. Enhanced functional properties of biopolymer film incorporated with curcurmin-loaded mesoporous silica nanoparticles for food packaging. Food Chem. 2019, 288, 139–145. [Google Scholar] [CrossRef]
- Sharma, A.; Yadav, A.; Gupta, N.; Sharma, S.; Kakkar, R.; Cwiklinski, K.; Quaye, E.; Mahajan, S.D.; Schwartz, S.A.; Sharma, R.K. Multifunctional mesoporous curcumin encapsulated iron-phenanthroline nanocluster: A new anti-HIV agent. Colloids Surf. B Biointerfaces 2019, 180, 289–297. [Google Scholar] [CrossRef]
- Cheng, Y.; Zhang, Y.; Deng, W.; Hu, J. Antibacterial and anticancer activities of asymmetric lollipop-like mesoporous silica nanoparticles loaded with curcumin and gentamicin sulfate. Colloids Surf. B Biointerfaces 2020, 186, 110744. [Google Scholar] [CrossRef]
- Song, Y.; Cai, L.; Tian, Z.; Wu, Y.; Chen, J. Phytochemical curcumin-coformulated, silver-decorated melanin-like polydopamine/mesoporous silica composites with improved antibacterial and chemotherapeutic effects against drug-resistant cancer cells. ACS Omega 2020, 5, 15083–15094. [Google Scholar] [CrossRef]
- Lawal, A.T. Graphene-based nano composites and their applications. A review. Biosens. Bioelectron. 2019, 141, 111384. [Google Scholar] [CrossRef]
- Farani, M.R.; Parsi, P.K.; Riazi, G.; Ardestani, M.S.; Rad, H.S. Extending the application of a magnetic PEG three-part drug release device on a graphene substrate for the removal of Gram-positive and Gram-negative bacteria and cancerous and pathologic cells. Drug Des. Dev. 2019, 13, 1581–1591. [Google Scholar] [CrossRef] [Green Version]
- Ghorbanzadeh, R.; Assadian, H.; Chiniforush, N.; Parker, S.; Pourakbari, B.; Ehsani, B.; Alikhani, M.Y.; Bahador, A. Modulation of virulence in Enterococcus faecalis cells surviving antimicrobial photodynamic inactivation with reduced graphene oxide-curcumin: An ex vivo biofilm model. Photodiagn. Photodyn. 2020, 29, 101643. [Google Scholar] [CrossRef]
- Oves, M.; Rauf, M.A.; Ansari, M.O.; Khan, A.A.P.; Qari, H.; Alajmi, M.F.; Sau, S.; Iyer, A.K. Graphene decorated zinc oxide and curcumin to disinfect the methicillin-resistant Staphylococcus aureus. Nanomaterials 2020, 10, 1004. [Google Scholar] [CrossRef]
- Singh, A.; Prakash, P.; Singh, R.; Nandy, N.; Firdaus, Z.; Bansal, M.; Singh, R.K.; Srivastava, A.; Roy, J.K.; Mishra, B.; et al. Curcumin quantum dots mediated degradation of bacterial biofilms. Front. Microbiol. 2017, 8, 1517. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.; Chang, L.; Chu, H.; Lin, H.; Chang, P.; Wang, R.Y.L.; Unnikrishnan, B.; Mao, J.; Chen, S.; Huang, C. High Amplification of the antiviral activity of curcumin through transformation into carbon quantum dots. Small 2019, 15, e1902641. [Google Scholar] [CrossRef]
- Singh, A.K.; Mishra, H.; Firdaus, Z.; Yadav, S.; Aditi, P.; Nandy, N.; Sharma, K.; Bose, P.; Pandey, A.K.; Chauhan, B.S.; et al. MoS2-modified curcumin nanostructures: The novel theranostic hybrid having potent antibacterial and antibiofilm activities against multidrug-resistant hypervirulent Klebsiella pneumoniae. Chem. Res. Toxicol. 2019, 32, 1599–1618. [Google Scholar] [CrossRef] [Green Version]
- Pourhajibagher, M.; Parker, S.; Chiniforush, N.; Bahador, A. Photoexcitation triggering via semiconductor graphene quantum dots by photochemical doping with curcumin versus perio-pathogens mixed biofilms. Photodiagn. Photodyn. 2019, 28, 125–131. [Google Scholar] [CrossRef]
- Iravani, S.; Varma, R.S. Green synthesis, biomedical and biotechnological applications of carbon and graphene quantum dots. A review. Environ. Chem. Lett. 2020, 18, 703–727. [Google Scholar] [CrossRef] [Green Version]
- Mirzahosseinipour, M.; Khorsandi, K.; Hosseinzadeh, R.; Ghazaeian, M.; Shahidi, F.K. Antimicrobial photodynamic and wound healing activity of curcumin encapsulated in silica nanoparticles. Photodiagn. Photodyn. 2020, 29, 101639. [Google Scholar] [CrossRef]
- Fakhrullina, G.; Khakimova, E.; Akhatova, F.; Lazzara, G.; Parisi, F.; Fakhrullin, R.F. Selective antimicrobial effects of curcumin@halloysite nanoformulation: A Caenorhabditis elegans Study. ACS Appl. Mater. Interfaces 2019, 11, 23050–23064. [Google Scholar] [CrossRef]
- Zou, X.; Yuan, M.; Zhang, T.; Wei, H.; Xu, S.; Jiang, N.; Zheng, N.; Wu, Z. Extracellular vesicles expressing a single-chain variable fragment of an HIV-1 specific antibody selectively target Env+ tissues. Theranostics 2019, 9, 5657–5671. [Google Scholar] [CrossRef]
- Sotomil, J.M.; Munchow, E.; Pankajakshan, D.; Spolnik, K.J.; Ferreira, J.A.; Gregory, R.L.; Bottino, M.C. Curcumin-a natural medicament for root canal disinfection: Effects of irrigation, drug release, and photoactivation. J. Endod. 2019, 45, 1371–1377. [Google Scholar] [CrossRef]
- Sattaria, S.; Tehrani, A.D.; Adelia, M.; Soleimania, K.; Rashidipourb, M. Fabrication of new generation of co-delivery systems based on graphene-g-cyclodextrin/chitosan nanofiber. Int. J. Biol. Macromol. 2020, 156, 1126–1134. [Google Scholar] [CrossRef]
- Yang, Y.-N.; Lu, K.-Y.; Wang, P.; Ho, Y.-C.; Tsai, M.-L.; Mi, F.-L. Development of bacterial cellulose/chitin multi-nanofibers based smart films containing natural active microspheres and nanoparticles formed in situ. Carbohydr. Polym. 2020, 228, 115370. [Google Scholar] [CrossRef] [PubMed]
- Esmaeili, E.; Eslami-Arshaghi, T.; Hosseinzadeh, S.; Elahirad, E.; Jamalpoor, Z.; Hatamie, S.; Soleimani, M. The biomedical potential of cellulose acetate/polyurethane nanofibrous mats containing reduced graphene oxide/silver nanocomposites and curcumin: Antimicrobial performance and cutaneous wound healing. Int. J. Biol. Macromol. 2020, 152, 418–427. [Google Scholar] [CrossRef] [PubMed]
- Xi, Y.; Ge, J.; Wang, M.; Chen, M.; Niu, W.; Cheng, W.; Xue, Y.; Lin, C.; Lei, B. Bioactive anti-inflammatory, antibacterial, antioxidative silicon-based nanofibrous dressing enables cutaneous tumor photothermo-chemo therapy and infection-induced wound healing. ACS Nano 2020, 14, 2904–2916. [Google Scholar] [CrossRef]
- Shababdoust, A.; Zandi, M.; Ehsani, M.; Shokrollahi, P.; Foudazi, R. Controlled curcumin release from nanofibers based on amphiphilic-block segmented polyurethanes. Int. J. Pharm. 2020, 575, 118947. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zhuang, B.; Du, G.; Han, G.; Jin, Y. Curcumin solid dispersion-loaded in situ hydrogels for local treatment of injured vaginal bacterial infection and improvement of vaginal wound healing. J. Pharm. Pharm. 2019, 71, 1044–1054. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Rhim, J.-W. Preparation of antimicrobial and antioxidant gelatin/curcumin composite films for active food packaging application. Colloids Surf. B Biointerfaces 2020, 188, 110761. [Google Scholar] [CrossRef]
- Roy, S.; Rhim, J.-W. Carboxymethyl cellulose-based antioxidant and antimicrobial active packaging film incorporated with curcumin and zinc oxide. Int. J. Biol. Macromol. 2020, 148, 666–676. [Google Scholar] [CrossRef] [PubMed]
- Ezati, P.; Rhim, J.-W. pH-responsive pectin-based multifunctional films incorporated with curcumin and sulfur nanoparticles. Carbohydr. Polym. 2020, 230, 115638. [Google Scholar] [CrossRef]
- Aydogdu, A.; Radke, C.J.; Bezci, S.; Kirtil, E. Characterization of curcumin incorporated guar gum/orange oil antimicrobial emulsion films. Int. J. Biol. Macromol. 2020, 148, 110–120. [Google Scholar] [CrossRef]
- Dai, C.; Wang, Y.; Sharma, G.; Shen, J.; Velkov, T.; Xiao, X. Polymyxins–curcumin combination antimicrobial therapy: Safety implications and efficacy for infection treatment. Antioxidants 2020, 9, 506. [Google Scholar] [CrossRef]
- Hakeem, M.J.; Asseri, K.A.; Ma, L.; Chou, K.C.; Konkel, M.E.; Lu, X. A novel mathematical model for studying antimicrobial interactions against Campylobacter jejuni. Front. Microbiol. 2019, 10, 1038. [Google Scholar] [CrossRef]
- Paschoal, M.A.; Moura, C.M.Z.; Jeremias, F.; Souza, J.F.; Bagnato, V.S.; Giusti, J.S.M.; Santos-Pinto, L. Longitudinal effect of curcumin-photodynamic antimicrobial chemotherapy in adolescents during fixed orthodontic treatment: A single-blind randomized clinical trial study. Lasers Med. Sci. 2014, 30, 2059–2065. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Solvent | Microorganism | Culture | Antimicrobial Method | CUR Concentration | Light/Ultrasonic Parameters | Reference |
|---|---|---|---|---|---|---|
| DMSO (0.4%) | ZIKV | Cell infection | IC50 | 5.62–16.57 µM | - | [19] |
| >DGEV | >IC90 | |||||
| N/R | HPVA | Cell infection | Viral survival | 0.015 mg/mL | - | [20] |
| Tulane V | ||||||
| N/R | KSPV | Infected cells | EC50 | Up to 6.68 µM | - | [21] |
| Aqueous Piper nigrum seed extract | SARS-CoV-2 | Cell infection | IC50 Plaque reduction | 0.4 µg/mL | - | [22] |
| DMSO (<0.4%) | SARS-CoV-1 | Cell infection | Inhibiton of viral replication | 20 µM | - | [23] |
| N/R | SARS-CoV | In vitro | Viral inhibition | 23.5 µM | - | [24] |
| N/R | SARS-CoV | In vitro | papain-like inhibition | 5.7 µM | - | [25] |
| DMSO (1 w/v) | S. aureus | Planktonic | Inhibition zone MIC | 600 and | - | [26] |
| E. coli | 400 µg/mL | |||||
| DMSO | MRSA | Planktonic | MIC FICI | 15.5 µg/mL | - | [27] |
| N/R | S. aureus | Planktonic | Colony count | 100 µg/mL | 8 or 20 J/cm2 | [28] |
| MSSA | ||||||
| MRSA | ||||||
| DMSO (10%) | S. aureus | Biofilm | aPDT | 20, 40, and 80 µM | 5.28 J/cm2 | [29] |
| DMSO | VRSA | Biofilm/animal infection model | MIC MBC | 156.25 µg/mL | 20 J/cm2 | [30] |
| N/R | S. aureus | Animal infection model | aPDT | 78 µg/mL | 60 J/cm2 | [31] |
| DMSO | S. aureus | Infected fruit | Survival fraction | 100 nM | 1.5 and 9 J/cm2 | [32] |
| N/R | S. aureus | Planktonic | PDI | 40 and 80 µM | 15 J/cm2 | [33] |
| E. coli | ||||||
| Tween 80 (0.5%) | S. aureus | Planktonic | CFU/mL | 300 and 500 µM | 0.03–0.05 W/cm2 | [34] |
| N/R | S. aureus | Biofilm | Confocal microscope | N/R | 170 µmol m2 s1 | [35] |
| DMSO (0.5%) | S. aureus | Biofilm | SDT aPDT SPDT | 80 µM | 100 Hz 15 and 70 J/cm2 100 Hz, 15 and 70 J/cm2 | [36] |
| DMSO | E. coli | Planktonic | MIC Inhibition zone | 110, 220 and 330 µg/mL | - | [37] |
| DMSO | E. coli | Planktonic | OD600nm | 8,16, 32, and 64 µg/mL | - | [38] |
| N/R | S. dysenteriae | Planktonic | MIC/MBC | 256 and | - | [39] |
| C. jejuni | 512 µg/mL | |||||
| Edible alcohol | E. coli | Planktonic | aPDT | 5, 10, and 20 µM | 3.6 J/cm2 | [40] |
| DMSO | H. pylori | Planktonic biofilm | MIC MBC aPDT | 50 µg/mL | 10 mW/cm2 | [41] |
| DMSO | P. aeruginosa | Biofilm | aPDT CFU/mL | N/R | 5 and 10 J/cm2 | [42] |
| DMSO | Imipenem-resistant A. baumannii | Planktonic | aPDT | 25, 50, 100, and 200 µM | 5.4 J/cm2 | [43] |
| DMSO (2%) | P. aeruginosa, A. baumannii, K. pneumoniae, E. coli, E. faecalis | Planktonic | MIC/FICI | 128-256 µg/mL | - | [44] |
| N/R | C. difficile, C. sticklandii, B. fragilis, P. bryantii | Planktonic | Viable cell number | 10 µg/mL | - | [45] |
| N/R | B. subtillis, E. coli, S. carnosus, M. smegmatis | Planktonic | MIC/MBC | Up to 25 µM | - | [46] |
| N/R | MRSA | Planktonic/animal infection model | MIC | 4–16 μg/mL | - | [47] |
| MSSA | 2–8 μg/mL | |||||
| E. coli | 8–32 μg/mL | |||||
| N/R | E. faecalis, S. aureus, B. subtillis, P. aeruginosa, E. coli | Planktonic | MIC | 156 μg/mL | - | [48] |
| DMSO (0.5%) | A. hydrophila, E. coli E. faecalis, K. pneumoniae, P. aeruginosa, S. aureus, C. albicans | Planktonic | MIC/MBC/ FICI/aPDT | 37.5–150 µg/mL | N/C | [49] |
| N/R | E. faecalis | Infection model | CFU/mL | 1 µg/mL | - | [50] |
| Commercial solution | E. faecalis | Biofilm | aPDT | 1.5 g/mL | 20.1 J/cm2 | [51] |
| Ethanol 99% | A. hydrophila, V. parahaemolyticus | Planktonic | aPDT/SDT | Up to 15 mg/L | N/C | [52] |
| DMSO (10%) | E. faecalis | Biofilm | MIC/MBC | 120 mg/mL | - | [53] |
| N/R | S. mutans | Planktonic | aPDT | 10 g/100cc | N/C | [54] |
| DMSO: ethyl alcohol | S. mutans, S. pyogenes | Planktonic | aPDT | 3 mg/mL | 28.8 J/cm2 | [55] |
| DMSO (0.8%) | Caries isolated | Biofilm | aPDT | 600 µg/mL | 75 J/cm2 | [56] |
| DMSO | S. mutans, C. albicans | Biofilm single/dual | MBEC | 0.5 mM | - | [57] |
| DMSO (0.05 M) | A. actinomycetemcomitans | Planktonic | aPDT | 40 µg/mL | 300–420 J/cm2 | [58] |
| DMSO (<1%) | P. gingivalis, A. actinomycetemcomitans | Planktonic | aPDT | 20 µg/mL | 6, 12 or 18 J/cm2 | [59] |
| DMSO (0.5%) | P. gingivalis, A. actinomycetemcomitans, C. rectus, E. corrodens, F. nucleatum, P. intermedia, P. micra, T. denticola, T. forsythis | Biofilm | aPDT | 100 mg/L | - | [60] |
| N/R | Subgingival plaque | Biofilm | aPDT | 100 µg/mL | 30 J/cm2 | [61] |
| DMSO | P. gingivalis | Planktonic | MIC | 12.5 µg/mL | - | [62] |
| Ethanol: DMSO (99.9%: 0.1%) | Periodontal pocket | - | aPDT | 100 mg/mL | 7.69 J/cm2 | [63] |
| Tween 80 | Streptococcus spp, Staphylococcus spp, Enterobacteriaceae, C. albicans | Clinical trail | aPDT | 0.75 mg/mL | 20.1 J/cm2 | [64] |
| Sodium hydroxide: PBS | C. albicans, C. parapsilosis, C. glabrata, C.dubliniensis | Planktonic/biofilm | MIC | 0.1–0.5 mg/mL | - | [65] |
| N/R | C. albicans, S. aureus | Planktonic Biofilm | MIC/Biofilm percentag | 200 µg/mL | - | [66] |
| N/R | C. albicans | Biofilm | aPDT | 1.5 g/mL | 20.1 J/cm2 | [67] |
| DMSO (10%) | C. albicans | Biofilm | aPDT | 20, 40, 60 and 80 µM | 2.64, 5.28, 7.92, 10.56, and 13.2 J/cm2 | [68] |
| DMSO (1%) | C. albicans | Biofilm | aPDT | 40 and 80 mM | 37.5 and 50 J/cm2 | [69] |
| N/R | C. albicans | Biofilm | aPDT | 100 µM | 10 J/cm2 | [70] |
| DMSO (2.5%) | Fluconazole-resistant C. albicans | Planktonic/biofilm/infection model | MIC/ aPDT | 40 µM | 5.28 J/cm2 | [71] |
| Fluconazole-susceptible C. albicans | 80 µM | 40.3 J/cm2 | ||||
| DMSO | C. albicans, F. oxysporum, A. flavus, A. niger, C. neoformans | Planktonic | MIC | 137.5–200 μg/mL | - | [72] |
| Type of Micelles | [CUR] Formulation | Microorganism | Type of Culture | Antimicrobial Method | Antimicrobial [CUR] | Light/Ultrasonic Parameters | Reference |
|---|---|---|---|---|---|---|---|
| Mixed polymer micelles | 1000 ppm | E. coli, S. aureus, A. niger | Planktonic | MIC | 350 and 275 µg/mL | - | [92] |
| PCL-b-PAsp and Ag | 2 mg/mL | P. aeruginosa, S. aureus | Planktonic | OD600nm | 8–500 µg/mL | - | [93] |
| mPEG-OA | 1:10 | P. aeruginosa | Planktonic | MIC | 400 µg/mL | - | [94] |
| PEG-PCL | 10 mg | C. albicans | Planktonic | MIC | 256 µg/mL | - | [95] |
| PEG-PE | 50 mM | S. mutans | Planktonic | SACT | 50 mM | 1.56 W/cm2 | [96] |
| DAPMA, SPD, SPM | 0.32 mg/mL | P. aeruginosa | Planktonic | OD600nm and aPDT | 250, 500 nM, 1 µM and 50, 100 nM | 18 and 30 J/cm2 | [97] |
| P123 | 0.5% w/V | S. aureus | Planktonic | aPDT | 7.80 μmol/L | 6.5 J/cm2 | [98] |
| PCL-b-PHMG-b-PCL, STES | 10 mg | S. aureus, E. coli | Planktonic | MIC | 16 and 32 μg/mL * | - | [99] |
| CUR-PLGA-DEX | 1 mg/mL | P. fluorescens, P. putida | Planktonic biofilm | OD600nm antibiofilm | 0.625–5 mg/mL | - | [100] |
| Type of Liposomes or SLN | [CUR] Formulation | Microorganism | Type of Culture | Antimicrobial Method | Antimicrobial [CUR] | Reference |
|---|---|---|---|---|---|---|
| Lecithin and cholesterol | 0.5 mg/mL | A. sobria, C. violaceum, A. tumefaciens | Planktonic biofilm | MIC, antibiofilm | 420, 400, and 460 μg/mL | [105] |
| PCNL | 60.65 ± 1.68 µg/µL | B. subtilis, K. pneumoniae, C. violaceum, E. coli, M. smegmatis, A. niger, C. albicans, F. oxysporum | Planktonic | Disk diffusion assay | N/R | [106] |
| Phosphocolines | 100:1 M | S. aureus | Planktonic | MIC | 7 μg/mL | [107] |
| PLGA: triglycerides: F68 | 0.8 mg/mL | E. coli, S. typhimurium, P. aeruginosa, S. aureus, B. sonorensis, B. licheniformis | Planktonic | MIC | 75 and 100 μg/mL | [108] |
| Soya lecithin and menthol | 0.5 mg/mL | MRSA | Planktonic Biofilm | MIC, microscopy, biomass | 10 and 125 µg/mL | [109] |
| CurSLN | 60 mg/500 mg lipid | S. aureus, S. mutans, V. streptococci, L. acidophilus, E. coli, C. albicans | Planktonic | MIC MBC | 0.09375–3 and 1.5–6 mg/mL | [110] |
| Type of Emulsion | [CUR] Formulation | Microorganism | Type of Culture | Antimicrobial Method | Antimicrobial Concentration | Light/Ultrasonic Parameters | Reference |
|---|---|---|---|---|---|---|---|
| THC ME | 5% | HIV-1 | Cell infection | IC50 | 0.9357 μM | - | [116] |
| CUR-NE | N/R | HPV | - | aPDT | 80 µM | 50 J/cm2 | [117] |
| CUR-NE | N/R | DENV-1 to 4 | Cell infection | Cell viability | 1, 5, 10 µg/mL | - | [118] |
| P60-CUR | 4 mg/L | E. coli | Planktonic | OD 595 nm | N/R | - | [119] |
| PE:CUR | 0.566 mg/mL | S. aureus, S. epidermidis, S. faecalis, C. albicans, E. coli | Planktonic | Inhibition zone | 1 mg/mL * | - | [120] |
| cu-SEDDS | 1% | S. aureus, E. coli, P. aeruginosa, K. pneumonia | Planktonic | MIC | 45–62 µg/mL | - | [121] |
| CUR:NE in microbeads | 0.5 mg/mL | E. coli, S. typhmerium, Y. enterocolitica, P. aeruginosa, S. aureus, B. cereus, L. monocytogenes | Planktonic | Inhibition zone | 90 and 180 mg/mL * | - | [122] |
| Lignin sulfomethylated | 0.3 mg/mL | S. aureus | Planktonic | OD 600 nm | 2.4 mg/mL * | - | [123] |
| C14-EDA/GM/W C14-MEDA/GM/W | N/R | C. albicans | Planktonic, biofilm | Microdilution assay, antibiofilm | 100 µg/mL, 20 µg | - | [124] |
| Type of CD | [CUR] Formulation | Microorganism | Type of Culture | Antimicrobial Method | Antimicrobial [CUR] | Light/Ultrasonic Parameters | Reference |
|---|---|---|---|---|---|---|---|
| PEG-based β-CD or γ-CD | 10 µM | E. coli, E. faecalis | Planktonic | aPDT | 10 µM | 4.8, 29 J/cm2 | [133] |
| HPMC-stabilized hydroxypro pyl-β-CD | 7.64 × 10−3 M | E. coli | Planktonic | aPDT | 10, 25 µM | 5, 14, 28 J/cm2 | [134] |
| methyl-β-CD hyaluronic acid HPMC | 7.64 × 10−3 M | E. faecalis, E. coli | Planktonic | aPDT | 0.5–25 µM | 11, 16, 32 J/cm2 | [135] |
| carboxymethyl-β-CD | 20 µM | E. coli | Planktonic | aPDT | 0.7 ± 0.1 to 4.1 ± 1.6 nmole cm−2 | 1050 ± 250 lx | [136] |
| hydrogel with CUR in hydroxypropyl-β-CD | 15.8 mg/mL | S. aureus | Planktonic | Inhibition zone | 2% (w/v) | - | [137] |
| α- and β-CD | 1 mol/L | E. coli, S. aureus | Planktonic | MIC, OD 600 nm | 0.25 and 0.31 mg/mL | - | [138] |
| β-CD or γ-CD in CS | 0.06 mM | E. coli, S. aureus | Planktonic | MIC, Zone of inhibition | 64 and 32 µg/mL | - | [139] |
| γ-CD | 25 mg/L | T. rubrum | Planktonic | MIC, aPDT | N/R | 45 J/cm2 | [140] |
| hydroxypropyl-β-CD | 1:1 | B. subtillis, S. aureus, S. pyrogenes, P. aeruginosa, C. difficile, C. butyricum, L. monocytogenes, E. faecalis, E. coli, K. pneumoniae, P. mirabilis, S. typhimurium, E. aerogens, C. kusei, C. albicans | Planktonic | Inhibition zone | 25 mg/mL | - | [141] |
| methyl-β-CD | 20 mM | E. coli | Planktonic | MIC, MBC, aPDT | 500, 90 µM | 9 J/cm2 | [142] |
| Type of CS | [CUR] Formulation | Microorganism | Type of Culture | Antimicrobial Method | Antimicrobial [CUR] | Reference |
|---|---|---|---|---|---|---|
| PEG-CS | 4.4%, 5 mg/mL | MRSA, P. aeruginosa | Planktonic, Animal model | OD600nm, CFU | 5 and 10 mg/mL * | [148] |
| CCS microspheres | 12.27 mg/mL, 1 mol | S. aureus, E. coli | Planktonic | Zone of inhibition, MIC | N/R | [149] |
| CS nanoparticles | 1.06 mg/mL | S. mutans | Planktonic, biofilm | MIC | 0.114 mg/mL | [150] |
| CS-CMS-MMT | 0.0004–0.004 g | S. mutans | Planktonic, Biofilm | MIC | 0.101 mg/mL | [151] |
| CS-GP-CUR | 148.09 ± 5.01 µg | S. aureus | Planktonic | Zone of inhibition, tissue bacteria count | N/C | [152] |
| PVA-CS-CUR | N/C | E. coli, P. aeruginosa, S. aureus, B. subtilis | Planktonic | Zone of inhibition | N/R | [153] |
| PVA-CS-CUR | 10, 20, 30 mg | P. multocida, S. aureus, E. coli, B. subtilis | Planktonic | Zone of inhibition | 10, 20, 30 mg | [154] |
| CS NPs | 2, 4, 8, 16% | C. albicans, S. aureus | Planktonic, Biofilm | MIC, Colony count | 400 mg/mL | [155] |
| CS NPs | 4 mg/mL | HCV-4 | N/R | Antiviral assay | 15 µg/mL | [156] |
| CS/milk protein nanocomposites | 100 mg | PVY | Plant infection | Antiviral activity | 500, 1000, 1500 mg/100 mL | [157] |
| Type of Polymeric DDS | [CUR] Formulation | Microorganism | Type of Culture | Antimicrobial Method | Antimicrobial [CUR] | Light/Ultrasonic Parameters | Reference |
|---|---|---|---|---|---|---|---|
| PEG 400 γ-CD and PEG + β-CD | 0.18% | E. faecalis E. coli | Planktonic | CFU/mL aPDT | N/R | 9.7 J/cm2 29 J/cm2 | [158] |
| CUR-NP without polymer | 100 mg | S. aureus B. subtillis E. coli P. aeruginosa P. notatum A. niger | Planktonic | MIC Inhibition zone | 100 mg 0.27 mmol | - | [159] |
| CUR-NP without polymer | 100 mg | M. lutues S. aureus E. coli P. aeruginosa | Planktonic | MBC | N/R | - | [160] |
| Mixed polymer NP | 5 mM | E. coli | Planktonic | MIC | 400–500 μM | - | [161] |
| CTAB Tween 20 Sodium dodecylsulfate | 100 mg/mL | L. monicytogenes | Planktonic | Inhibition zone | N/R | - | [162] |
| PLA/dextran sulfate | 4 mg/mL | MRSA C. albicans S. mutans | Planktonic/mono- and –mixed biofilm | aPDT | 260 μM | 43.2 J/cm2 | [163] |
| PLA/dextran sulfate | 0.4% | C. albicans | Animal model | aPDT | 260 μM | 37.5 J/cm2 | [164] |
| Nanocurcumin | N/R | P. aeruginosa (isolates) and standard strain | Planktonic | MIC | 128 µg/mL | - | [165] |
| PLGA | 5 mg | S. saprophyticus subsp. Bovis E. coli | Planktonic | aPDT | 50 µg/mL | 13.2 J/cm2 | [166] |
| Eudragit L-100 | N/C | L. monocytogenes | Planktonic | Animal model infection | N/R | - | [167] |
| nCUR | N/R | S. mutans | Planktonic Biofilm | Inhibition zone aPDT | N/R | 300–420 J/cm2 | [168] |
| nCUR combined with indocyanine | 100 mg | E. faecalis | Biofilm | Metabolic activity | N/R | 500 mW/cm2 | [169] |
| PVAc-CUR-PET-PVDC | 0.02 g | S. aureus S. tiphimurium | Planktonic | aPDT | N/R | 24, 48, and 72 J/cm2 | [170] |
| MOA.CUR-PLGA-NP | Up to 10% | S. mutans | Biofilm | aPDT | 7% wt | 45 J/cm2 | [171] |
| CS- β-CD | N/C | S. aureus E. coli | Planktonic | Colony count | Up to 0.03% | - | [172] |
| Type of Metallic Material | [CUR] Formulation | Microorganism | Type of Culture | Antimicrobial Method | Antimicrobial [CUR] | Reference |
|---|---|---|---|---|---|---|
| CUR-AgNPs | 20 mg/mL | P. aeruginosa, E. coli, B. subtilis, S. aureus | Planktonic | MIC | 20 mg/mL | [177] |
| Ag-CUR-nanoconjugates | 0.1 mM | E. coli, Salmonella spp., Fusarium spp., S. aureus | Planktonic | Zone of Inhibition | 0.1 mM | [178] |
| AgCURNPs | 500 mg | P. aeruginosa S. aureus | Biofilm | CLSM SEM | Up to 400 μg/mL | [179] |
| AgNPs | 7 mg | E. coli | Planktonic | Turbidimetric Assay | 0.005 µM | [180] |
| cAgNPs | 7 mg | E. coli B. subtilis | Planktonic | MIC CFU/mL | 7 mg | [181] |
| Ru II complex | 0.092 g | E. coli, S. aureus, K. pneumoniae, A. baumannii,. P. aeruginosa, Enterococcus sp. | Plakntonic | MIC/FICI | >64 µg/mL | [182] |
| SCMC SNCF nanocomposites with CUR | 0.25 mg/mL | E. coli | Planktonic | Disc Method Count Method | 2 mg/mL | [183] |
| CSCL CUR-AgNP | 0.092 g | E. coli, B. subtilis | Planktonic | Zone of Inhibition | 10 and 20 μM | [184] |
| nSnH | 10% | S. aureus E. coli. | Planktonic | CFU/mL | N/R | [185] |
| Nanocomposite of CUR and ZnO NPs | N/C | S. epidermidis S. hemolyticus S. saprophyticus | Planktonic | Zone of Inhibition | 1000, 750, 500, 250 μg/mL | [186] |
| Thermo-responsive hydrogels | N/C | S. aureus P. aeruginosa E. coli | Planktonic | MIC | 400 μg/mL | [187] |
| CUR-AgNPs | 5 mg/mL | C. albicans, C. glabrata, C. tropicalis, C. parapsilosis, C. krusei, C. kefyr | Planktonic | Zone of Inhibition, MIC | 32.2–250 μg/mL | [188] |
| Gel-CUR-Ag | 20 mg | P. aeruginosa S. aureus | Planktonic | MIC MBC | 20 mg | [189] |
| HGZ-CUR | N/C | S. aureus T. rubrum | Planktonic | Zone of Inhibition | N/C | [190] |
| CHG-ZnO-CUR | N/C | S. aureus T. rubrum | Planktonic | Zone of Inhibition | N/C | [191] |
| Copper (II) oxide NPs | 1 g | E. faecalis P. aeruginosa | Planktonic | Zone of Inhibition CFU/mL | 1 mg/mL | [192] |
| OA-Ag-C | 1 g | P. aeruginosa S. aureus | Planktonic | OD600nm | 2.5 mg/mL | [193] |
| Ag-NP-β-CD-BC | 0.79 g | P. aeruginosa, S. aureus, C. auris | Planktonic | Zone of Inhibition | N/R | [194] |
| Cotton fabrics coated ZnO-NP | 2.71 × 10−3 M | S. aureus, E. coli | Planktonic | Bacterial Count | N/R | [195] |
| CS-ZnO-CUR | 0.2 g | S. aureus, E. coli | Planktonic | MIC MBC | Up to 50 μg/mL | [196] |
| CUR-TiO2 -CS | 100–300 mg | S. aureus, E. coli | Planktonic Animal infection | MIC | 10 mg | [197] |
| CUR-Au-NPs | 1 mg/mL | E. coli, B. subtilis, S. aureus, P. aeruginosa | Planktonic | Zone of Inhibition | 100, 200, 300 μg/mL | [198] |
| Porous DDS | [CUR] Formulation | Microorganism | Type of Culture | Antimicrobial Method | Antimicrobial [CUR] | Light/Ultrasonic Parameters | Reference |
|---|---|---|---|---|---|---|---|
| Cu-SNP/Ag | 1.0 mmol | E. coli | Planktonic | aPDT | N/R | 72 J/cm2 | [204] |
| Bionanocomposite silica/chitosan | 100 mg | E. coli S. aureus | Planktonic | Zone of inhibition | N/R | - | [205] |
| NCIP | 1 mg | HIV-1 | Transfected cells | Immunofluorescent staining | 5–8 mg/mL | - | [206] |
| Lollipop-like MSN | 30 mg L−1 | E. coli S. aureus | Planktonic | OD600nm | N/R | - | [207] |
| SBA-15/PDA/Ag | 2 mg | E. coli S. aureus | Planktonic | CFU/mL | 50 mM | - | [208] |
| Type of Material | [CUR] Formulation | Microorganism | Type of Culture | Antimicrobial Method | Antimicrobial [CUR] | Light/Ultrasonic Parameters | Reference |
|---|---|---|---|---|---|---|---|
| G-NH2–IONP–PEG | 0.004 g | S. aureus E. coli | Planktonic | Colony count | 100, 125 μg/mL | - | [210] |
| CUR-rGO | N/R | E. faecalis | Biofilm | MBIC aPDT | 250 μg/mL | 360 J/cm2 | [211] |
| GrZnO | N/R | MRSA | Planktonic | MIC Inhibition zone Metabolic activity | Up to 62.5 μg/mL | - | [212] |
| CUR-cQDs | 0.6 | S. aureus MRSA E. faecalis E. coli K. pneumoniae P. aeruginosa | Planktonic Biofilm | Grown inhibition Biomass evaluation Confocal microscopy | 3.91–7.825 µg/mL | - | [213] |
| CUR-cQDs | 200 mg | EV-71 | Cell infection Animal infection | MIC Plaque assay TCIC50 assay Western blot PCR | 5 μg/mL | - | [214] |
| CUR-MQD | 2:1 wt% | K. pneumoniae P. aeruginosa S. aureus | Planktonic | MIC MBC Confocal microscopy Fluorescence microscopy Flow cytometry | <0.00625–0.125 μg/mL | - | [215] |
| CUR-GQDs | N/C | A. actinomycetemcomitans P. gingivalis P. intermedia | Mixed- biofilm | aPDT | 100 μg/mL | 60–80 J/cm2 | [216] |
| Type of Material | [CUR] Formulation | Microorganism | Type of Culture | Antimicrobial Method | Antimicrobial [CUR] | Light/Ultrasonic Parameters | Reference |
|---|---|---|---|---|---|---|---|
| CuR-SiNPs | 20 mg | S. aureus P. aeruginosa | Planktonic Biofilm | aPDT | 50 μg/mL 1 mg/mL | 20 J/cm2 | [218] |
| CUR-HNT-DX | 10 mg | S. marcescens E. coli | Planktonic Infection model | Grown inhibition, Confocal microscopy | Up to 0.5 mg/mL | - | [219] |
| Exosomes | N/R | HIV-1 infection | - | Flow cytometry | N/R | - | [220] |
| Electrospun nanofibers | 100 mg/mL | Actinomyces naeslundii | Biofilm | aPDT | 2.5 and 5 mg/mL | 1200 mW/cm2 | [221] |
| Ga NF CD-GO NF | 0.1 mol | B. cereus E. coli | Planktonic | Zone of inhibition MIC | Up to 63.25 µg/mL | - | [222] |
| Multinanofibers-film | 1, 2.5, and 5 mg/mL | S. aureus E. coli | Planktonic | UFC/mL Confocal microscopy | 1 mg/mL | - | [223] |
| Nanofibers scaffolds | 4.0 wt% | S. aureus Pseudomonas sp. | Planktonic | Colony count | N/R | - | [224] |
| Nanofibrous scaffold | 5% | S. aureus E. coli | Planktonic | Colony count | 20 mg | - | [225] |
| Nanofibers | 5 and 10%wt | S. aureus E. coli | Planktonic | OD600nm | Up to 212.5 µg/mL | - | [226] |
| CSDG | 1 w/w | S. aureus E. coli | Planktonic Infection model | Colony count Microscopy | N/R | - | [227] |
| Gelatin film | 0, 0.25, 0.5, 1.0, and 1.5 wt% | E. coli L. monocytogenes | Planktonic | UFC/mL | 0.25 and 1.5 wt% | - | [228] |
| ZnO-CMC film | 0.5 and 1.0 wt% | E. coli L. monocytogenes | Planktonic | UFC/mL | 1 wt% | - | [229] |
| Pectin film | 40 mg | E. coli L. monocytogenes | Planktonic | UFC/mL | N/R | - | [230] |
| Edible film | 0.4% (w/v) | E. coli B. subtilis | Planktonic | Zone of inhibition | 1% wt. | - | [231] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trigo-Gutierrez, J.K.; Vega-Chacón, Y.; Soares, A.B.; Mima, E.G.d.O. Antimicrobial Activity of Curcumin in Nanoformulations: A Comprehensive Review. Int. J. Mol. Sci. 2021, 22, 7130. https://doi.org/10.3390/ijms22137130
Trigo-Gutierrez JK, Vega-Chacón Y, Soares AB, Mima EGdO. Antimicrobial Activity of Curcumin in Nanoformulations: A Comprehensive Review. International Journal of Molecular Sciences. 2021; 22(13):7130. https://doi.org/10.3390/ijms22137130
Chicago/Turabian StyleTrigo-Gutierrez, Jeffersson Krishan, Yuliana Vega-Chacón, Amanda Brandão Soares, and Ewerton Garcia de Oliveira Mima. 2021. "Antimicrobial Activity of Curcumin in Nanoformulations: A Comprehensive Review" International Journal of Molecular Sciences 22, no. 13: 7130. https://doi.org/10.3390/ijms22137130
APA StyleTrigo-Gutierrez, J. K., Vega-Chacón, Y., Soares, A. B., & Mima, E. G. d. O. (2021). Antimicrobial Activity of Curcumin in Nanoformulations: A Comprehensive Review. International Journal of Molecular Sciences, 22(13), 7130. https://doi.org/10.3390/ijms22137130