
Ionomic Approaches for Discovery of Novel Stress-Resilient Genes in Plants

Abstract
:1. Introduction
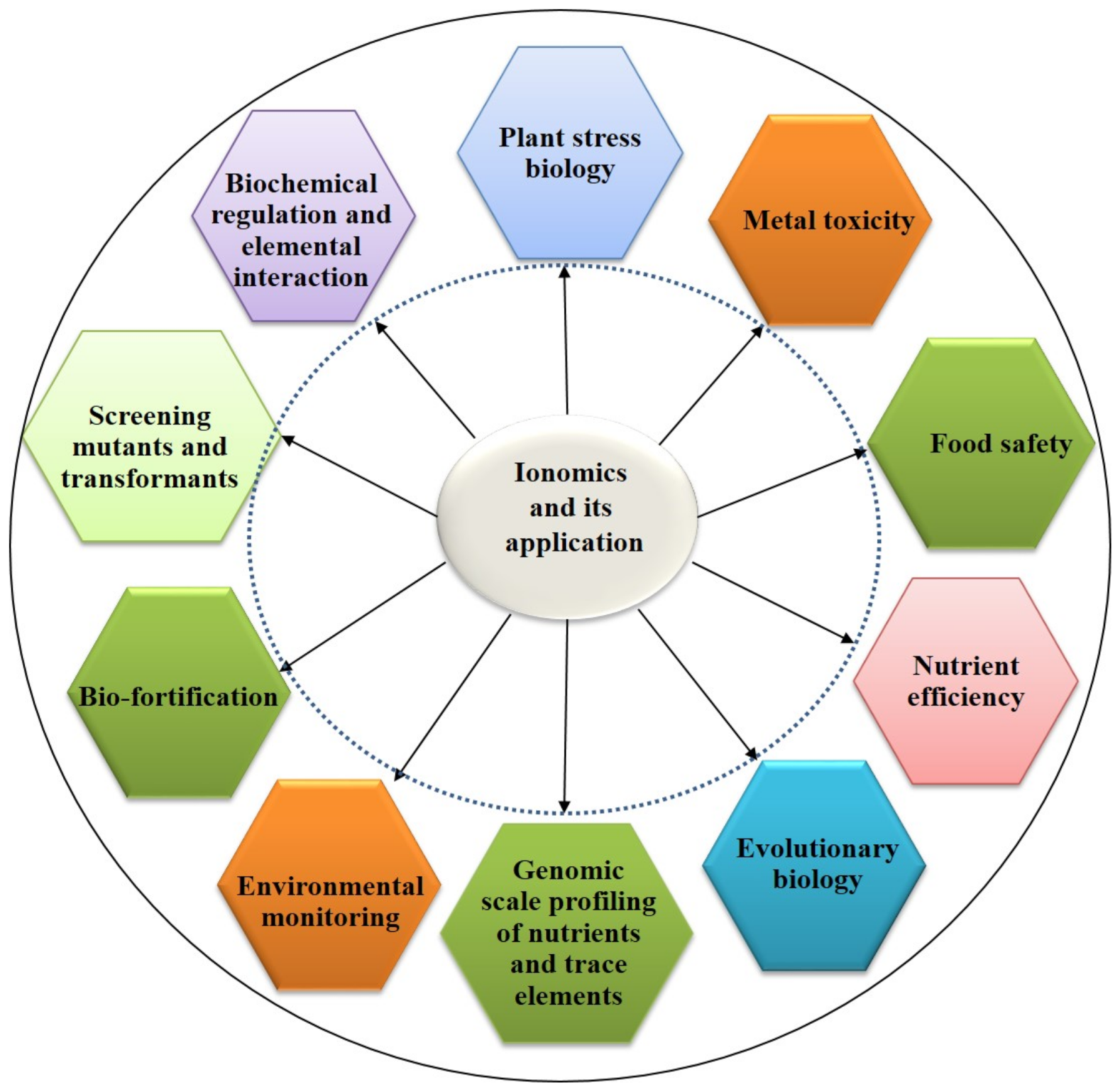
2. Importance of Ionomics in Human Health, Food, and Agriculture
3. Ionomics and Plant Biotic Stress
4. Ionomics for Enhancement of Abiotic Stress Resilience in Crops
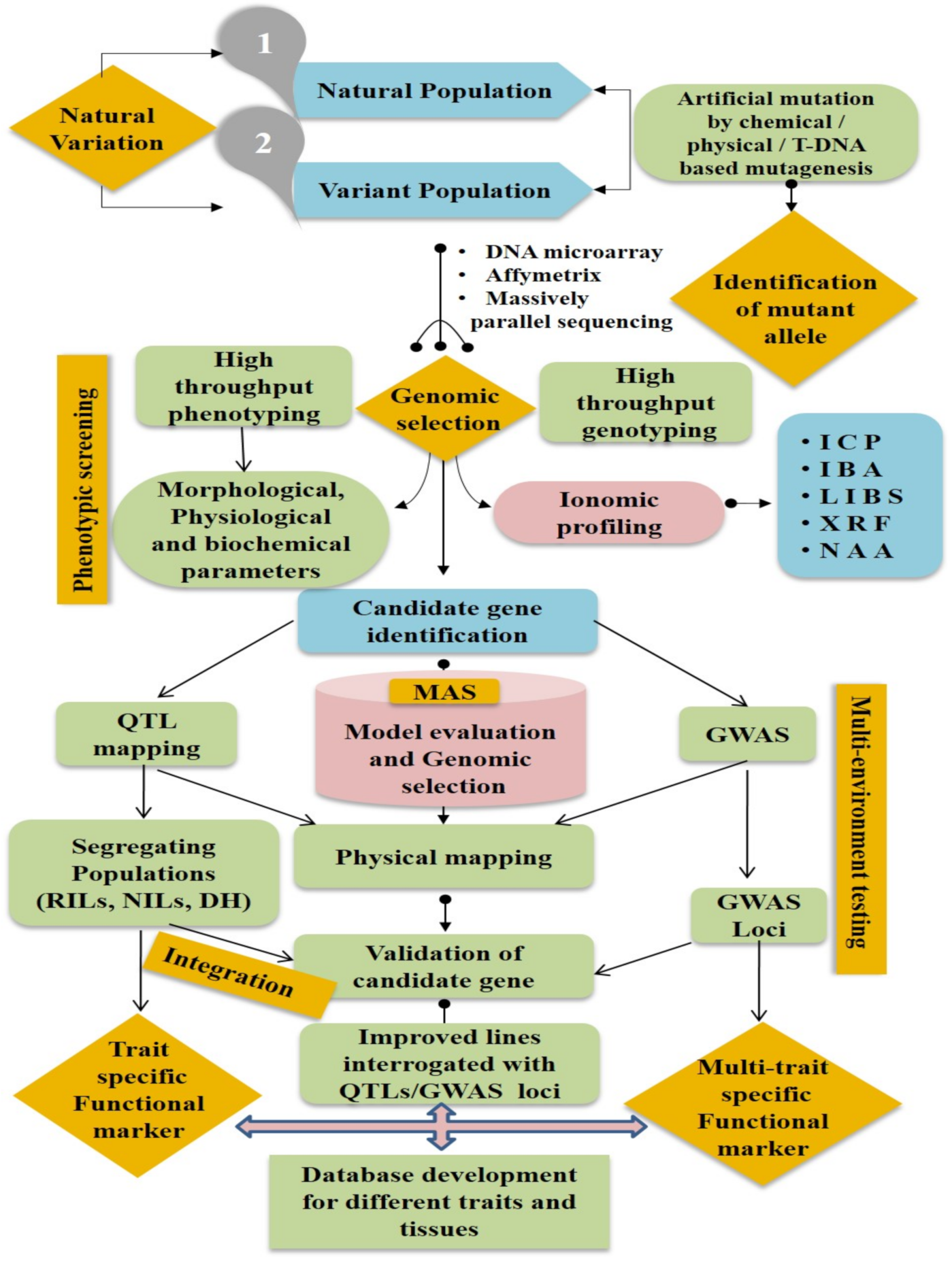
5. Quantitative Trait Loci (QTLs) and Genes Regulating Ionomic Traits
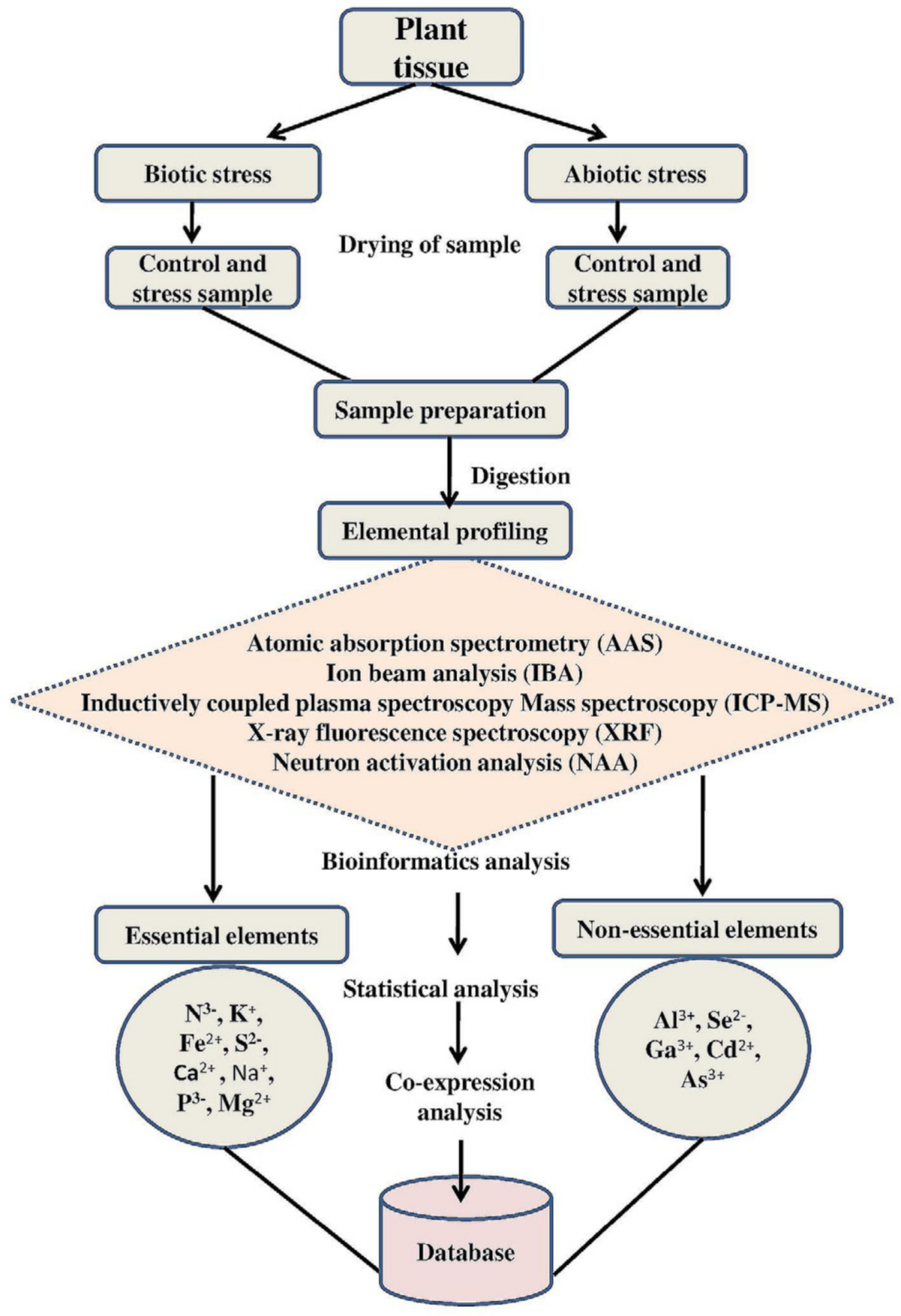
6. Tools and Technological Advancements for Ionomics Studies
7. Transporters and Channels Involved in Elemental Uptake and Transport
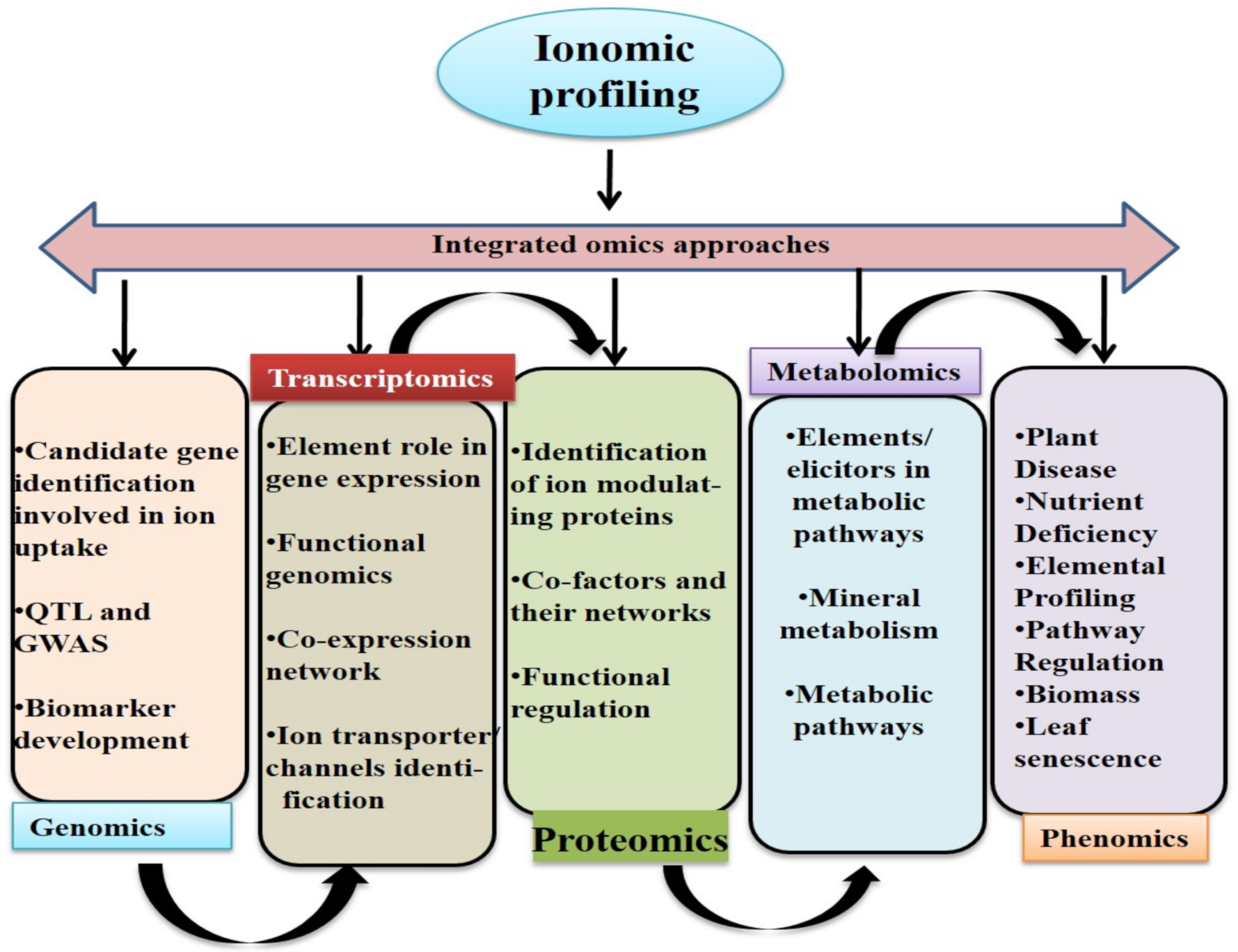
8. Ionomics and Integrated Omics Approaches
9. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Atkinson, N.J.; Urwin, P.E. The interaction of plant biotic and abiotic stresses: From genes to the field. J. Exp. Bot. 2012, 63, 3523–3543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, N.; Rivero, R.M.; Shulaev, V.; Blumwald, E.; Mittler, R. Abiotic and Biotic Stress Combinations. New Phytol. 2014, 203, 32–43. [Google Scholar] [CrossRef] [PubMed]
- He, M.; He, C.Q.; Ding, N.Z. Abiotic Stresses: General Defenses of Land Plants and Chances for Engineering Multistress Tolerance. Front. Plant Sci. 2018, 1771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, S.; Mir, Z.A.; Tyagi, A.; Bhat, J.A.; Chandrashekar, N.; Papolu, P.K.; Rawat, S.; Grover, A. Identification and comparative analysis of Brassica juncea pathogenesis-related genes in response to hormonal, biotic and abiotic stresses. Acta Physiol. Plant. 2017, 39, 268. [Google Scholar] [CrossRef]
- Pandey, A.; Ramegowda, V.; Senthil-Kumar, M. Shared and Unique Responses of Plants to Multiple Individual Stresses and Stress Combinations: Physiological and Molecular Mechanisms. Front. Plant Sci. 2015, 723. [Google Scholar] [CrossRef] [Green Version]
- Raza, A.; Razzaq, A.; Mehmood, S.S.; Zou, X.; Zhang, X.; Lv, Y.; Xu, J. Impact of Climate Change on Crops Adaptation and Strategies to Tackle Its Outcome: A Review. Plants 2019, 8, 34. [Google Scholar] [CrossRef] [Green Version]
- Muszyńska, E.; Labudda, M. Dual Role of Metallic Trace Elements in Stress Biology—From Negative to Beneficial Impact on Plants. Int. J. Mol. Sci. 2019, 20, 3117. [Google Scholar] [CrossRef] [Green Version]
- Campos, A.C.A.L.; Kruijer, W.; Alexander, R.; Akkers, R.C.; Danku, J.; Salt, D.E.; Aarts, M.G.M. Natural variation in Arabidopsis thaliana reveals shoot ionome, biomass, and gene expression changes as biomarkers for zinc deficiency tolerance. J. Exp. Bot. 2017, 68, 3643–3656. [Google Scholar] [CrossRef] [Green Version]
- Maathuis, F.J. Physiological Functions of Mineral Macronutrients. Curr. Opin. Plant Biol. 2009, 12, 250–258. [Google Scholar] [CrossRef]
- Williams, L.; Salt, D.E. The Plant Ionome Coming into Focus. Curr. Opin. Plant Biol. 2009, 12, 247–249. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, T.; Urayama, M.; Shinano, T.; Okada, R.; Osaki, M. Application of Ionomics to Plant and Soil in Fields under Long-Term Fertilizer Trials. SpringerPlus 2015, 4, 781. [Google Scholar] [CrossRef]
- Fattorini, L.; Ronzan, M.; Piacentini, D.; Della Rovere, F.; de Virgilio, C.; Sofo, A.; Altamura, M.M.; Falasca, G. Cadmium and Arsenic Affect Quiescent Centre Formation and Maintenance in Arabidopsis thaliana Post-Embryonic Roots Disrupting Auxin Biosynthesis and Transport. Environ. Exp. Bot. 2017, 144, 37–48. [Google Scholar] [CrossRef]
- Shahid, M.; Khalid, S.; Abbas, G.; Shahid, N.; Nadeem, M.; Sabir, M.; Aslam, M.; Dumat, C. Heavy Metal Stress and Crop Productivity. In Crop Production and Global Environmental Issues; Springer: Cham, Switzerland, 2015; pp. 1–25. [Google Scholar]
- Salt, D.E.; Baxter, I.; Lahner, B. Ionomics and the Study of the Plant Ionome. Annu. Rev. Plant. Biol. 2008, 59, 709–733. [Google Scholar] [CrossRef] [Green Version]
- Pita-Barbosa, A.; Ricachenevsky, F.K.; Wilson, M.; Dottorini, T.; Salt, D.E. Transcriptional Plasticity Buffers Genetic Variation in Zinc Homeostasis. Sci. Rep. 2019, 9, 19482. [Google Scholar] [CrossRef] [Green Version]
- Lahner, B.; Gong, J.; Mahmoudian, M.; Smith, E.L.; Abid, K.B.; Rogers, E.E.; Guerinot, M.L.; Harper, J.F.; Ward, J.M.; McIntyre, L.; et al. Genomic Scale Profiling of Nutrient and Trace Elements in Arabidopsis thaliana. Nat. Biotechnol. 2003, 21, 1215–1221. [Google Scholar] [CrossRef]
- Danku, J.M.C.; Lahner, B.; Yakubova, E.; Salt, D.E. Large-Scale Plant Ionomics. Methods Mol. Biol. 2013, 953, 255–276. [Google Scholar]
- Ziegler, G.; Terauchi, A.; Becker, A.; Armstrong, P.; Hudson, K.; Baxter, I. Ionomic Screening of Field-Grown Soybean Identifies Mutants with Altered Seed Elemental Composition. Plant Genome 2013, 6. [Google Scholar] [CrossRef] [Green Version]
- Baxter, I.R.; Ziegler, G.; Lahner, B.; Mickelbart, M.V.; Foley, R.; Danku, J.; Armstrong, P.; Salt, D.E.; Hoekenga, O.A. Single-Kernel Ionomic Profiles Are Highly Heritable Indicators of Genetic and Environmental Influences on Elemental Accumulation in Maize Grain (Zea mays). PLoS ONE 2014, 9, e87628. [Google Scholar] [CrossRef] [Green Version]
- Pinson, S.R.M.; Tarpley, L.; Yan, W.; Yeater, K.; Lahner, B.; Yakubova, E.; Huang, X.-Y.; Zhang, M.; Guerinot, M.L.; Salt, D.E.; et al. Worldwide Genetic Diversity for Mineral Element Concentrations in Rice Grain. Crop Sci. 2015, 55, 294–311. [Google Scholar] [CrossRef]
- Thomas, C.L.; Alcock, T.D.; Graham, N.S.; Hayden, R.; Matterson, S.; Wilson, L.; Young, S.D.; Dupuy, L.X.; White, P.J.; Hammond, J.P.; et al. Root Morphology and Seed and Leaf Ionomic Traits in a Brassica Napus L: Diversity Panel Show Wide Phenotypic Variation and Are Characteristic of Crop Habit. BMC Plant Biol. 2016, 16, 214. [Google Scholar] [CrossRef] [Green Version]
- D’Attoma, G.; Morelli, M.; Saldarelli, P.; Saponari, M.; Giampetruzzi, A.; Boscia, D.; Savino, V.N.; la Fuente, L.D.; Cobine, P.A. Ionomic Differences between Susceptible and Resistant Olive Cultivars Infected by Xylella Fastidiosa in the Outbreak Area of Salento, Italy. Pathogens 2019, 8, 272. [Google Scholar] [CrossRef] [Green Version]
- Cesco, S.; Tolotti, A.; Nadalini, S.; Rizzi, S.; Valentinuzzi, F.; Mimmo, T.; Porfido, C.; Allegretta, I.; Giovannini, O.; Perazzolli, M.; et al. Plasmopara Viticola Infection Affects Mineral Elements Allocation and Distribution in Vitis Vinifera Leaves. Sci. Rep. 2020, 10, 18759. [Google Scholar] [CrossRef] [PubMed]
- Baxter, I.R.; Vitek, O.; Lahner, B.; Muthukumar, B.; Borghi, M.; Morrissey, J.; Guerinot, M.L.; Salt, D.E. The Leaf Ionome as a Multivariable System to Detect a Plant’s Physiological Status. Proc. Natl. Acad. Sci. USA 2008, 105, 12081–12086. [Google Scholar] [CrossRef] [Green Version]
- Chao, D.Y.; Baraniecka, P.; Danku, J.; Koprivova, A.; Lahner, B.; Luo, H.; Yakubova, E.; Dilkes, B.; Kopriva, S.; Salt, D.E. Variation in Sulfur and Selenium Accumulation Is Controlled by Naturally Occurring Isoforms of the Key Sulfur Assimilation Enzyme Adenosine 5’-Phosphosulfate Reductase2 across the Arabidopsis Species Range. Plant Physiol. 2014, 166, 1593–1608. [Google Scholar] [CrossRef] [Green Version]
- White, P.J.; Broadley, M.R.; Thompson, J.A.; McNicol, J.W.; Crawley, M.J.; Poulton, P.R.; Johnston, A.E. Testing the Distinctness of Shoot Ionomes of Angiosperm Families Using the Rothamsted Park Grass Continuous Hay Experiment. New Phytol. 2012, 196, 101–109. [Google Scholar] [CrossRef] [Green Version]
- Shakoor, N.; Ziegler, G.; Dilkes, B.P.; Brenton, Z.; Boyles, R.; Connolly, E.L.; Kresovich, S.; Baxter, I. Integration of Experiments across Diverse Environments Identifies the Genetic Determinants of Variation in Sorghum Bicolor Seed Element Composition. Plant Physiol. 2016, 170, 1989–1998. [Google Scholar] [CrossRef] [Green Version]
- Kamiya, T.; Borghi, M.; Wang, P.; Danku, J.M.C.; Kalmbach, L.; Hosmani, P.S.; Naseer, S.; Fujiwara, T.; Geldner, N.; Salt, D.E. The MYB36 Transcription Factor Orchestrates Casparian Strip Formation. Proc. Natl. Acad. Sci. USA 2015, 112, 10533–10538. [Google Scholar] [CrossRef] [Green Version]
- Hindt, M.N.; Akmakjian, G.Z.; Pivarski, K.L.; Punshon, T.; Baxter, I.; Salt, D.E.; Guerinot, M. Lou. BRUTUS and Its Paralogs, BTS LIKE1 and BTS LIKE2, Encode Important Negative Regulators of the Iron Deficiency Response in Arabidopsis thaliana. Metallomics 2017, 9, 876–890. [Google Scholar] [CrossRef]
- Whitt, L.; Ricachenevsky, F.K.; Ziegler, G.Z.; Clemens, S.; Walker, E.; Maathuis, F.J.; Kear, P.; Baxter, I. A curated list of genes that affect the plant ionome. Plant Direct 2020, 4, e00272. [Google Scholar] [CrossRef]
- Kisko, M.; Bouain, N.; Safi, A.; Medici, A.; Akkers, R.C.; Secco, D.; Fouret, G.; Krouk, G.; Aarts, M.G.M.; Busch, W.; et al. LPCAT1 controls phosphate homeostasis in a zinc-dependent manner. eLife 2018, 7, e32077. [Google Scholar] [CrossRef]
- Barberon, M.; Dubeaux, G.; Kolb, C.; Isono, E.; Zelazny, E.; Vert, G. Polarization of IRON-REGULATED TRANSPORTER 1 (IRT1) to the plant-soil interface plays crucial role in metal homeostasis. Proc. Natl. Acad. Sci. USA 2014, 111, 8293–8298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Shi, S.; Wang, L.; Tang, Z.; Lv, T.; Zhu, X.; Ding, X.; Wang, Y.; Zhao, F.J.; Zhongchang, W.U. OsHAC4 Is critical for arsenate tolerance and regulates arsenic accumulation in rice. New Phytol. 2017, 215, 1090–1101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Miao, Y.U.; Xin-Qiao, D.U.; Wang, Z.F.; Wei-Hua, W.U.; Quintero, F.J.; Jin, X.H.; Li, H.D.; Wang, Y.I. NRT1.5/NPF7.3 functions as a proton-coupled H+/K+ antiporter for K+ loading into the xylem in arabidopsis. Plant Cell 2017, 29, 2016–2026. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.-Y.; Chao, D.-Y.; Koprivova, A.; Danku, J.; Wirtz, M.; Müller, S.; Sandoval, F.J.; Bauwe, H.; Roje, S.; Dilkes, B.; et al. Nuclear localised More Sulphur Accumulation1 epigenetically regulates sulphur homeostasis in Arabidopsis thaliana. PLoS Genet. 2016, 12, e1006298. [Google Scholar] [CrossRef]
- Yan, J.; Chia, J.-C.; Sheng, H.; Jung, H.-I.; Tetiana-Olena Zavodna, L.U.; Zhang, R.H.; Zavodna, T.O.; Zhang, L.; Huang, R.; Jiao, C.; et al. Arabidopsis pollen fertility requires the transcription factors CITF1 and SPL7 that regulate copper delivery to anthers and jasmonic acid synthesis. Plant Cell 2017, 29, 3012–3029. [Google Scholar] [CrossRef] [Green Version]
- Olsen, L.I.; Hansen, T.H.; Larue, C.; Osterberg, J.T.; Hoffmann, R.D.; Liesche, J.; Kramer, U.; Surblé, S.; Cadarsi, S.; Samson, V.A.; et al. Mother-plant-mediated pumping of zinc into the developing seed. Nat. Plants 2016, 2, 16036. [Google Scholar] [CrossRef]
- Zhu, J.; Lau, K.; Puschmann, R.; Harmel, R.K.; Zhang, Y.; Pries, V.; Gaugler, P.; Broger, L.; Dutta, A.K.; Jessen, H.J.; et al. Two bifunctional inositol pyrophosphate kinases/phosphatases control plant phosphate homeostasis. eLife 2019, 8, e43582. [Google Scholar] [CrossRef]
- Lindsay, E.R.; Maathuis, F.J.M. Arabidopsis thaliana NIP7;1 is involved in tissue arsenic distribution and tolerance in response to arsenate. FEBS Lett. 2016, 590, 779–786. [Google Scholar] [CrossRef] [Green Version]
- Ueno, D.; Sasaki, A.; Yamaji, N.; Miyaji, T.; Fujii, Y.; Takemoto, Y.; Moriyama, S.; Che, J.; Moriyama, Y.; Iwasaki, K.; et al. A polarly localized transporter for efficient manganese uptake in rice. Nat. Plants 2015, 1, 15170. [Google Scholar] [CrossRef]
- Kobayashi, T.; Nagasaka, S.; Senoura, T.; Itai, R.N.; Nakanishi, H.; Nishizawa, N.K. Iron-binding haemerythrin RING ubiquitin ligases regulate plant iron responses and accumulation. Nat. Comm. 2013, 4, 2792. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.I.; Sun, S.-K.; Tang, Z.; Liu, G.; Moore, K.L.; Maathuis, F.J.M.; Miller, A.J.; McGrath, S.P.; Zhao, F.-J. The nodulin 26-like intrinsic membrane protein OsNIP3;2 is involved in arsenite uptake by lateral roots in rice. J. Exp. Bot. 2017, 68, 3007–3016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, X.-Y.; Deng, F.; Yamaji, N.; Pinson, S.R.M.; Fujii-Kashino, M.; Danku, J.; Douglas, A.; Guerinot, M.L.; Salt, D.E.; Ma, J.F. A heavy metal P-Type ATPase OsHMA4 prevents copper accumulation in rice grain. Nat. Commun. 2016, 7, 12138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, X.-Y.; Liu, H.; Zhu, Y.-F.; Pinson, S.R.M.; Lin, H.-X.; Guerinot, M.L.; Zhao, F.-J.; Salt, D.E. Natural variation in a molybdate transporter controls grain molybdenum concentration in rice. New Phytol. 2019, 221, 1983–1997. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.-K.; Chen, Y.I.; Che, J.; Konishi, N.; Tang, Z.; Miller, A.J.; Ma, J.F.; Zhao, F.-J. Decreasing arsenic accumulation in rice by overexpressing OsNIP1;1 and OsNIP3;3 through disrupting arsenite radial transport in roots. New Phytol. 2018, 219, 641–653. [Google Scholar] [CrossRef] [Green Version]
- Takemoto, Y.; Tsunemitsu, Y.; Fujii-Kashino, M.; Mitani-Ueno, N.; Yamaji, N.; Ma, J.F.; Kato, S.-I.; Iwasaki, K.; Ueno, D. The tonoplast-localized transporter MTP8.2 contributes to manganese detoxification in the shoots and roots of Oryza Sativa L. Plant Cell Physiol. 2017, 58, 1573–1582. [Google Scholar] [CrossRef]
- Rai, H.; Yokoyama, S.; Satoh-Nagasawa, N.; Furukawa, J.; Nomi, T.; Ito, Y.; Fujimura, S.; Takahashi, H.; Suzuki, R.; Yousra, E.; et al. Cesium uptake by rice roots largely depends upon a single gene, HAK1, which encodes a potassium transporter. Plant Cell Physiol. 2017, 58, 1486–1493. [Google Scholar] [CrossRef] [Green Version]
- Senoura, T.; Sakashita, E.; Kobayashi, T.; Takahashi, M.; Aung, M.S.; Masuda, H.; Nakanishi, H.; Nishizawa, N.K. The iron-chelate transporter OsYSL9 plays a role in iron distribution in developing rice grains. Plant Mol. Biol. 2017, 95, 375–387. [Google Scholar] [CrossRef]
- Hayashi, S.; Kuramata, M.; Abe, T.; Takagi, H.; Ozawa, K.; Ishikawa, S. Phytochelatin synthase OsPCS1 plays a crucial role in reducing arsenic levels in rice grains. Plant J. 2017, 91, 840–848. [Google Scholar] [CrossRef] [Green Version]
- Uraguchi, S.; Tanaka, N.; Hofmann, C.; Abiko, K.; Ohkama-Ohtsu, N.; Weber, M.; Kamiya, T.; Sone, Y.; Nakamura, R.; Takanezawa, Y.; et al. Phytochelatin synthase has contrasting effects on cadmium and arsenic accumulation in rice grains. Plant Cell Physiol. 2017, 58, 1730–1742. [Google Scholar] [CrossRef]
- Yamaji, N.; Takemoto, Y.; Miyaji, T.; Mitani-Ueno, N.; Yoshida, K.T.; Ma, J.F. Reducing phosphorus accumulation in rice grains with an impaired transporter in the node. Nature 2017, 541, 92–95. [Google Scholar] [CrossRef]
- Che, J.; Yokosho, K.; Yamaji, N.; Ma, J.F. A Vacuolar phytosiderophore transporter alters iron and zinc accumulation in polished rice grains. Plant Physiol. 2019, 181, 276–288. [Google Scholar] [CrossRef] [Green Version]
- Aggarwal, S.; Kumar, A.; Bhati, K.K.; Kaur, G.; Shukla, V.; Tiwari, S.; Pandey, A.K. RNAi-mediated downregulation of inositol pentakisphosphate kinase (IPK1) in wheat grains decreases phytic acid levels and increases Fe and Zn Accumulation. Front. Plant Sci. 2018, 9, 259. [Google Scholar] [CrossRef]
- Wang, J.; Sun, J.; Miao, J.; Guo, J.; Shi, Z.; He, M.; Chen, Y.; Zhao, X.; Li, B.; Han, F.; et al. A phosphate starvation response regulator Ta-PHR1 is involved in phosphate signalling and increases grain yield in wheat. Ann. Bot. 2013, 111, 1139–1153. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.U.; Ying, Y.; Narsai, R.; Ye, L.; Zheng, L.; Tian, J.; Whelan, J.; Shou, H. Identification of OsbHLH133 as a regulator of iron distribution between roots and shoots in Oryza Sativa. Plant Cell Environ. 2013, 36, 224–236. [Google Scholar] [CrossRef]
- Bhati, K.K.; Alok, A.; Kumar, A.; Kaur, J.; Tiwari, S.; Pandey, A.K. Silencing of ABCC13 transporter in wheat reveals its involvement in grain development, phytic acid accumulation and lateral root formation. J. Exp. Bot. 2016, 67, 4379–4389. [Google Scholar] [CrossRef] [Green Version]
- Laurie, S.; Feeney, K.A.; Maathuis, F.J.M.; Heard, P.J.; Brown, S.J.; Leigh, R.A. A role for HKT1 in sodium uptake by wheat roots. Plant J. 2002, 32, 139–149. [Google Scholar] [CrossRef]
- Zhang, M.; Cao, Y.; Wang, Z.; Wang, Z.Q.; Shi, J.; Liang, X.; Song, W.; Chen, Q.; Lai, J.; Jiang, C. A retrotransposon in an HKT1 family sodium transporter causes variation of leaf Na+ exclusion and salt tolerance in maize. New Phytol. 2018, 217, 1161–1176. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Chen, K.; Zhao, F.J.; Sun, C.; Jin, C.; Shi, Y.; Sun, Y.; Li, Y.; Yang, M.; Jing, X.; et al. OsATX1 interacts with heavy metal P1B-Type ATPases and affects copper transport and distribution. Plant Physiol. 2018, 178, 329–344. [Google Scholar] [CrossRef] [Green Version]
- Von Wiren, N.; Mori, S.; Marschner, H.; Romheld, V. Iron inefficiency in maize mutant ys1 (Zea mays L. Cv Yellow-Stripe) is caused by a defect in uptake of iron phytosidero-phores. Plant Physiol. 1994, 106, 71–77. [Google Scholar] [CrossRef] [Green Version]
- Chan-Rodriguez, D.; Walker, E.L. Analysis of yellow striped mutants of Zea mays reveals novel loci contributing to iron deficiency chlorosis. Front. Plant Sci. 2018, 9, 157. [Google Scholar] [CrossRef] [Green Version]
- Qin, Y.J.; Wei-Hua, W.U.; Wang, Y.I. ZmHAK5 and ZmHAK1 function in K+ uptake and distribution in maize under low K+ conditions. J. Int. Plant Biol. 2019, 61, 691–705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durbak, A.R.; Phillips, K.A.; Pike, S.; O’Neill, M.A.; Mares, J.; Gallavotti, A.; Malcomber, S.T.; Gassmann, W.; McSteen, P. Transport of boron by the tassel-less1 aquaporin is critical for vegetative and reproductive development in maize. Plant Cell 2014, 26, 2978–2995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gil-Díez, P.; Tejada-Jiménez, M.; León-Mediavilla, J.; Wen, J.; Mysore, K.S.; Imperial, J.; González-Guerrero, M. MtMOT1.2 is responsible for molybdate supply to Medicago truncatula nodules. Plant Cell Environ. 2018, 42, 310–320. [Google Scholar] [CrossRef] [Green Version]
- Tejada-Jiménez, M.; Gil-Díez, P.; León-Mediavilla, J.; Wen, J.; Mysore, K.S.; Imperial, J.; González-Guerrero, M. Medicago truncatula Molybdate Transporter Type 1 (MtMOT1.3) is a plasma membrane molybdenum transporter required for nitrogenase activity in root nodules under molybdenum deficiency. New Phytol. 2017, 216, 1223–1235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tejada-Jimenez, M.; Castro-Rodriguez, R.; Igor Kryvoruchko, M.; Lucas, M.; Udvardi, M.; Imperial, J.; Gonzalez-Guerrero, M. Medicago truncatula natural resistance-associated macrophage protein1 is required for iron uptake by rhizobia-infected nodule cells. Plant Physiol. 2015, 168, 258–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Senovilla, M.; Castro-Rodríguez, R.; Abreu, I.; Escudero, V.; Kryvoruchko, I.; Udvardi, M.K.; Imperial, J.; González-Guerrero, M. Medicago truncatula copper transporter 1 (MtCOPT1) delivers copper for symbiotic nitrogen fixation. New Phytol. 2018, 218, 696–709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- León-Mediavilla, J.; Senovilla, M.; Montiel, J.; Gil-Díez, P.; Saez, A.; Kryvoruchko, I.S.; Reguera, M.; Udvardi, M.K.; Imperial, J.; González-Guerrero, M. MtMTP2-facilitated zinc transport into intracellular compartments is essential for nodule development in Medicago truncatula. Front. Plant Sci. 2018, 9, 990. [Google Scholar] [CrossRef] [PubMed]
- Abreu, I.; Saez, A.; Castro-Rodríguez, R.; Escudero, V.; Rodríguez-Haas, B.; Senovilla, M.; Larue, C.; Grolimund, D.; Tejada-Jiménez, M.; Imperial, J.; et al. Medicago truncatula zinc-iron permease 6 provides zinc to rhizobia-infected nodule cells. Plant Cell Environ. 2017, 40, 2706–2719. [Google Scholar] [CrossRef]
- White, P.J.; Brown, P.H. Plant Nutrition for Sustainable Development and Global Health. Ann. Bot. 2010, 105, 1073–1080. [Google Scholar] [CrossRef] [Green Version]
- Younis, U.; Qayyum, M.F.; Shah, M.H.R.; Danish, S.; Shahzad, A.N.; Malik, S.A.; Mahmood, S. Growth, Survival, and Heavy Metal (Cd and Ni) Uptake of Spinach (Spinacia Oleracea) and Fenugreek (Trigonella Corniculata) in a Biochar-Amended Sewage-Irrigated Contaminated Soil. J. Plant Nutr. Soil Sci. 2015, 178, 209–217. [Google Scholar] [CrossRef]
- White, P.J.; Broadley, M.R. Biofortification of Crops with Seven Mineral Elements Often Lacking in Human Diets—Iron, Zinc, Copper, Calcium, Magnesium, Selenium and Iodine. New Phytol. 2009, 182, 49–84. [Google Scholar] [CrossRef]
- Bouis, H.E.; Saltzman, A. Improving nutrition through biofortification: A review of evidence from HarvestPlus, 2003 through 2016. Glob. Food Sec. 2017, 12, 49–58. [Google Scholar] [CrossRef]
- Buturi, C.V.; Mauro, R.P.; Fogliano, V.; Leonardi, C.; Giuffrida, F. Mineral biofortification of vegetables as a tool to improve human diet. Foods 2021, 10, 223. [Google Scholar] [CrossRef]
- Kanter, U.; Hauser, A.; Michalke, B.; Draxl, S.; Schäffner, A.R. Caesium and strontium accumulation in shoots of Arabidopsis thaliana: Genetic and physiological aspects. J. Exp. Bot. 2010, 61, 3995–4009. [Google Scholar] [CrossRef] [Green Version]
- Pawlowski, M.L.; Helfenstein, J.; Frossard, E.; Hartman, G.L. Boron and Zinc Deficiencies and Toxicities and Their Interactions with Other Nutrients in Soybean Roots, Leaves, and Seeds. J. Plant Nutr. 2019, 42, 634–649. [Google Scholar] [CrossRef]
- Afshan, S.; Ali, S.; Bharwana, S.A.; Rizwan, M.; Farid, M.; Abbas, F.; Ibrahim, M.; Mehmood, M.A.; Abbasi, G.H. Citric Acid Enhances the Phytoextraction of Chromium, Plant Growth, and Photosynthesis by Alleviating the Oxidative Damages in Brassica napus L. Environ. Sci. Pollut. Res. 2015, 22, 11679–11689. [Google Scholar] [CrossRef]
- Adrees, M.; Ali, S.; Rizwan, M.; Zia-ur-Rehman, M.; Ibrahim, M.; Abbas, F.; Farid, M.; Qayyum, M.F.; Irshad, M.K. Mechanisms of Silicon-Mediated Alleviation of Heavy Metal Toxicity in Plants: A Review. Ecotoxicol. Environ. Saf. 2015, 119, 186–197. [Google Scholar] [CrossRef]
- Liang, Y.; Sun, W.; Zhu, Y.G.; Christie, P. Mechanisms of silicon-mediated alleviation of abiotic stresses in higher plants: A review. Environ. Pollut. 2007, 147, 422–428. [Google Scholar] [CrossRef] [Green Version]
- Maksimović, D.J.; Mojović, M.; Maksimović, V.; Römheld, V.; Nikolic, M. Silicon ameliorates manganese toxicity in cucumber by decreasing hydroxyl radical accumulation in the leaf apoplast. J. Exp. Bot. 2012, 63, 2411–2420. [Google Scholar] [CrossRef] [Green Version]
- Punshon, T.; Jackson, B.P. Essential Micronutrient and Toxic Trace Element Concentrations in Gluten Containing and Gluten-Free Foods. Food Chem. 2018, 252, 258–264. [Google Scholar] [CrossRef]
- Dordas, C. Role of Nutrients in Controlling Plant Diseases in Sustainable Agriculture: A Review. Agron. Sustain. Dev. 2009, 28, 33–46. [Google Scholar] [CrossRef] [Green Version]
- Navarrete, F.; De La Fuente, L. Zinc Detoxification Is Required for Full Virulence and Modification of the Host Leaf Ionome by Xylella fastidiosa. Mol. Plant Microbe Interact. 2015, 28, 497–507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walters, D.R.; Bingham, I.J. Influence of Nutrition on Disease Development Caused by Fungal Pathogens: Implications for Plant Disease Control. Ann. Appl. Biol. 2007, 151, 307–324. [Google Scholar] [CrossRef]
- Fones, H.; Davis, C.A.R.; Rico, A.; Fang, F.; Smith, J.A.C.; Preston, G.M. Metal Hyperaccumulation Armors Plants against Disease. PLoS Pathog. 2010, 6, e1001093. [Google Scholar] [CrossRef] [PubMed]
- Hoffland, E.; Jeger, M.J.; van Beusichem, M.L. Effect of nitrogen supply rate on disease resistance in tomato depends on the pathogen. Plant Soil 2000, 218, 239–247. [Google Scholar] [CrossRef]
- Sun, X.; Pan, B.; Wang, Y.; Xu, W.; Zhang, S. Exogenous Calcium Improved Resistance to Botryosphaeria dothidea by Increasing Autophagy Activity and Salicylic Acid Level in Pear. Mol. Plant Microbe Interact. 2020, 33, 1150–1160. [Google Scholar] [CrossRef] [PubMed]
- Clarkson, D.T.; Marschner, H. Mineral Nutrition of Higher Plants. Second Edition. 889pp; London: Academic Press, £29.95 (Paperback). Ann. Bot. 1996, 78, 527–528. [Google Scholar] [CrossRef] [Green Version]
- Cakmak, I. Tansley Review No. 111. Possible roles of zinc in protecting plant cells from damage by reactive oxygen species. New Phytol. 2000, 146, 185–205. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Parihar, P.; Singh, R.; Singh, V.P.; Prasad, S.M. Heavy Metal Tolerance in Plants: Role of Transcriptomics, Proteomics, Metabolomics, and Ionomics. Front. Plant Sci. 2016, 6, 1143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kieu, N.P.; Aznar, A.; Segond, D.; Rigault, M.; Simond-Côte, E.; Kunz, C.; Soulie, M.C.; Expert, D.; Dellagi, A. Iron Deficiency Affects Plant Defence Responses and Confers Resistance to Dickeya Dadantii and Botrytis Cinerea. Mol. Plant Pathol. 2012, 13, 816–827. [Google Scholar] [CrossRef] [PubMed]
- Del Coco, L.; Migoni, D.; Girelli, C.R.; Angilè, F.; Scortichini, M.; Fanizzi, F.P. Soil and Leaf Ionome Heterogeneity in Xylella fastidiosa Subsp. Pauca—Infected, Non-Infected and Treated Olive Groves in Apulia, Italy. Plants 2020, 9, 760. [Google Scholar] [CrossRef]
- Datnoff, L.E.; Rodrigues, F.A.; Seebold, K.W. Silicon and Plant Nutrition. In Mineral Nutrition and Plant Disease; Datnoff, L.E., Elmer, W.H., Huber, D.M., Eds.; American Phytopathological Society (APS Press): St. Paul, MN, USA, 2007; pp. 233–246. [Google Scholar]
- Cobine, P.A.; Cruz, L.F.; Navarrete, F.; Duncan, D.; Tygart, M.; de la Fuente, L. Xylella Fastidiosa Differentially Accumulates Mineral Elements in Biofilm and Planktonic Cells. PLoS ONE 2013, 8, e54936. [Google Scholar] [CrossRef] [PubMed]
- De La Fuente, L.; Parker, J.K.; Oliver, J.E.; Granger, S.; Brannen, P.M.; van Santen, E.; Cobine, P.A. The Bacterial Pathogen Xylella Fastidiosa Affects the Leaf Ionome of Plant Hosts during Infection. PLoS ONE 2013, 8, e62945. [Google Scholar] [CrossRef] [Green Version]
- Nicolas, O.; Charles, M.T.; Jenni, S.; Toussaint, V.; Parent, S.-É.; Beaulieu, C. The Ionomics of Lettuce Infected by Xanthomonas Campestris Pv. Vitians. Front. Plant Sci. 2019, 10, 351. [Google Scholar] [CrossRef]
- Yuan, M.; Wang, S.; Chu, Z.; Li, X.; Xu, C. The Bacterial Pathogen Xanthomonas oryzae Overcomes Rice Defenses by Regulating Host Copper Redistribution. Plant Cell 2010, 22, 3164–3176. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Sun, R.; Albrecht, U.; Padmanabhan, C.; Wang, A.; Coffey, M.D.; Girke, T.; Wang, Z.; Close, T.J.; Roose, M.; et al. Small RNA Profiling Reveals Phosphorus Deficiency as a Contributing Factor in Symptom Expression for Citrus Huanglongbing Disease. Mol. Plant 2013, 6, 301–310. [Google Scholar] [CrossRef] [Green Version]
- Yasin, N.A.; Zaheer, M.M.; Khan, W.U.; Ahmad, S.R.; Ahmad, A.; Ali, A.; Akram, W. The beneficial role of potassium in Cd-induced stress alleviation and growth improvement in Gladiolus grandiflora L. Int. J. Phytoremediat. 2018, 20, 274–283. [Google Scholar] [CrossRef]
- Han, Y.; Chaouch, S.; Mhamdi, A.; Queval, G.; Zechmann, B.; Noctor, G. Functional analysis of Arabidopsis mutants points to novel roles for glutathione in coupling H2O2 to activation of salicylic acid accumulation and signaling. Antioxid. Redox Signal. 2013, 18, 2106–2121. [Google Scholar] [CrossRef] [Green Version]
- Ghanta, S.; Bhattacharyya, D.; Sinha, R.; Banerjee, A.; Chattopadhyay, S. Nicotiana tabacum overexpressing γ-ECS exhibits biotic stress tolerance likely through NPR1-dependent salicylic acid-mediated pathway. Planta 2011, 233, 895–910. [Google Scholar] [CrossRef]
- Yadeta, K.A.; Elmore, J.M.; Creer, A.Y.; Feng, B.; Franco, J.Y.; Rufian, J.S.; He, P.; Phinney, B.; Coaker, G. A cysteine-rich protein kinase associates with a membrane immune complex and the cysteine residues are required for cell death. Plant Physiol. 2017, 173, 771–787. [Google Scholar] [CrossRef] [Green Version]
- Bollig, K.; Specht, A.; Myint, S.S.; Zahn, M.; Horst, W.J. Sulphur supply impairs spread of Verticillium dahliae in tomato. Eur. J. Plant Pathol. 2013, 135, 81–96. [Google Scholar] [CrossRef]
- Kruse, C.; Haas, F.H.; Jost, R.; Reiser, B.; Reichelt, M.; Wirtz, M.; Gershenzon, J.; Schnug, E.; Hell, R. Improved sulfur nutrition provides the basis for enhanced production of sulfur-containing defense compounds in Arabidopsis thaliana upon inoculation with Alternaria brassicicola. J. Plant Physiol. 2012, 169, 740–743. [Google Scholar] [CrossRef] [PubMed]
- Amtmann, A.; Troufflard, S.; Armengaud, P. The effect of potassium nutrition on pest and disease resistance in plants. Physiol. Plant. 2008, 133, 682–691. [Google Scholar] [CrossRef] [PubMed]
- Holzmueller, E.J.; Jose, S.; Jenkins, M.A. Influence of calcium, potassium, and magnesium on Cornus florida L. density and resistance to dogwood anthracnose. Plant Soil 2007, 290, 189–199. [Google Scholar] [CrossRef]
- Delhaize, E. A Metal-Accumulator Mutant of Arabidopsis thaliana. Plant. Physiol. 1996, 111, 849–855. [Google Scholar] [CrossRef] [Green Version]
- Rogers, E.E.; Guerinot, M. Lou. FRD3, a Member of the Multidrug and Toxin Efflux Family, Controls Iron Deficiency Responses in Arabidopsis. Plant Cell 2002, 14, 1787–1799. [Google Scholar] [CrossRef] [Green Version]
- Durrett, T.P.; Gassmann, W.; Rogers, E.E. The FRD3-Mediated Efflux of Citrate into the Root Vasculature Is Necessary for Efficient Iron Translocation. Plant Physiol. 2007, 144, 197–205. [Google Scholar] [CrossRef] [Green Version]
- Rus, A.; Baxter, I.; Muthukumar, B.; Gustin, J.; Lahner, B.; Yakubova, E.; Salt, D.E. Natural Variants of AtHKT1 Enhance Na+ Accumulation in Two Wild Populations of Arabidopsis. PLoS Genet. 2006, 2, e210. [Google Scholar] [CrossRef] [Green Version]
- Baxter, I.; Brazelton, J.N.; Yu, D.; Huang, Y.S.; Lahner, B.; Yakubova, E.; Li, Y.; Bergelson, J.; Borevitz, J.O.; Nordborg, M.; et al. A Coastal Cline in Sodium Accumulation in Arabidopsis thaliana is Driven by Natural Variation of the Sodium Transporter AtHKT1;1. PLoS Genet. 2010, 6, e1001193. [Google Scholar] [CrossRef] [Green Version]
- Loudet, O.; Saliba-Colombani, V.; Camilleri, C.; Calenge, F.; Gaudon, V.; Koprivova, A.; North, K.A.; Kopriva, S.; Daniel-Vedele, F. Natural Variation for Sulfate Content in Arabidopsis thaliana is Highly Controlled by APR2. Nat. Genet. 2007, 39, 896–900. [Google Scholar] [CrossRef]
- Baxter, I.; Muthukumar, B.; Park, H.C.; Buchner, P.; Lahner, B.; Danku, J.; Zhao, K.; Lee, J.; Hawkesford, M.J.; Guerinot, M.; et al. Variation in Molybdenum Content Across Broadly Distributed Populations of Arabidopsis thaliana Is Controlled by a Mitochondrial Molybdenum Transporter (MOT1). PLoS Genet. 2008, 4, e1000004. [Google Scholar] [CrossRef] [Green Version]
- Poormohammad Kiani, S.; Trontin, C.; Andreatta, M.; Simon, M.; Robert, T.; Salt, D.E.; Loudet, O. Allelic Heterogeneity and Trade-Off Shape Natural Variation for Response to Soil Micronutrient. PLoS Genet. 2012, 8, e1002814. [Google Scholar] [CrossRef] [Green Version]
- Baxter, I.; Hosmani, P.S.; Rus, A.; Lahner, B.; Borevitz, J.O.; Muthukumar, B.; Mickelbart, M.V.; Schreiber, L.; Franke, R.B.; Salt, D.E. Root Suberin Forms an Extracellular Barrier That Affects Water Relations and Mineral Nutrition in Arabidopsis. PLoS Genet. 2009, 5, e1000492. [Google Scholar] [CrossRef] [Green Version]
- Hosmani, P.S.; Kamiya, T.; Danku, J.; Naseer, S.; Geldner, N.; Guerinot, M.L.; Salt, D.E. Dirigent Domain-Containing Protein Is Part of the Machinery Required for Formation of the Lignin-Based Casparian Strip in the Root. Proc. Natl. Acad. Sci. USA 2013, 110, 14498–14503. [Google Scholar] [CrossRef] [Green Version]
- Tian, H.; Baxter, I.R.; Lahner, B.; Reinders, A.; Salt, D.E.; Ward, J.M. Arabidopsis NPCC6/NaKR1 Is a Phloem Mobile Metal Binding Protein Necessary for Phloem Function and Root Meristem Maintenance. Plant Cell 2010, 22, 3963–3979. [Google Scholar] [CrossRef] [Green Version]
- Alassimone, J.; Fujita, S.; Doblas, V.G.; van Dop, M.; Barberon, M.; Kalmbach, L.; Vermeer, J.E.M.; Rojas-Murcia, N.; Santuari, L.; Hardtke, C.S.; et al. Polarly Localized Kinase SGN1 Is Required for Casparian Strip Integrity and Positioning. Nat. Plants 2016, 2, 16113. [Google Scholar] [CrossRef]
- Borghi, M.; Rus, A.; Salt, D.E. Loss-of-Function of Constitutive Expresser of Pathogenesis Related Genes5 Affects Potassium Homeostasis in Arabidopsis thaliana. PLoS ONE 2011, 6, e26360. [Google Scholar] [CrossRef] [Green Version]
- Chao, D.Y.; Gable, K.; Chen, M.; Baxter, I.; Dietrich, C.R.; Cahoon, E.B.; Guerinot, M.L.; Lahner, B.; Lü, S.; Markham, J.E.; et al. Sphingolipids in the Root Play an Important Role in Regulating the Leaf Ionome in Arab. Thaliana. Plant Cell 2011, 23, 1061–1081. [Google Scholar] [CrossRef] [Green Version]
- Chao, D.-Y.; Silva, A.; Baxter, I.; Huang, Y.S.; Nordborg, M.; Danku, J.; Lahner, B.; Yakubova, E.; Salt, D.E. Genome-Wide Association Studies Identify Heavy Metal ATPase3 as the Primary Determinant of Natural Variation in Leaf Cadmium in Arabidopsis thaliana. PLoS Genet. 2012, 8, e1002923. [Google Scholar] [CrossRef] [Green Version]
- Koprivova, A.; Giovannetti, M.; Baraniecka, P.; Lee, B.R.; Grondin, C.; Loudet, O.; Kopriva, S. Natural Variation in the ATPS1 Isoform of ATP Sulfurylase Contributes to the Control of Sulfate Levels in Arabidopsis. Plant Physiol. 2013, 163, 1133–1141. [Google Scholar] [CrossRef] [Green Version]
- Pfister, A.; Barberon, M.; Alassimone, J.; Kalmbach, L.; Lee, Y.; Vermeer, J.E.M.; Yamazaki, M.; Li, G.; Maurel, C.; Takano, J.; et al. A Receptor-like Kinase Mutant with Absent Endodermal Diffusion Barrier Displays Selective Nutrient Homeostasis Defects. eLife 2014, 3, e03115. [Google Scholar] [CrossRef]
- Chao, D.Y.; Chen, Y.; Chen, J.; Shi, S.; Chen, Z.; Wang, C.; Danku, J.M.; Zhao, F.J.; Salt, D.E. Genome-wide Association Mapping identifies a New Arsenate Reductase Enzyme Critical for Limiting Arsenic Accumulation in Plants. PLoS Biol. 2014, 12, e1002009. [Google Scholar] [CrossRef] [Green Version]
- Stolpe, C.; Giehren, F.; Krämer, U.; Müller, C. Both Heavy Metal-Amendment of Soil and Aphid-Infestation Increase Cd and Zn Concentrations in Phloem Exudates of a Metal-Hyperaccumulating Plant. Phytochemistry 2017, 139, 109–117. [Google Scholar] [CrossRef]
- Kumar, S.; Parekh, M.J.; Patel, C.B.; Zala, H.N.; Sharma, R.; Kulkarni, K.S.; Fougat, R.S.; Bhatt, R.K.; Sakure, A.A. Development and Validation of EST-Derived SSR Markers and Diversity Analysis in Cluster Bean (Cyamopsis tetragonoloba). J. Plant Biochem. Biotechnol. 2016, 25, 263–269. [Google Scholar] [CrossRef]
- Wang, Y.Q.; Feechan, A.; Yun, B.W.; Shafiei, R.; Hofmann, A.; Taylor, P.; Xue, P.; Yang, F.Q.; Xie, Z.S.; Pallas, J.A.; et al. S-Nitrosylation of AtSABP3 Antagonizes the Expression of Plant Immunity. J. Biol. Chem. 2009, 284, 2131–2137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calic, I.; Koch, J.; Carey, D.; Addo-Quaye, C.; Carlson, J.E.; Neale, D.B. Genome-Wide Association Study Identifies a Major Gene for Beech Bark Disease Resistance in American Beech (Fagus grandifolia Ehrh.). BMC Genom. 2017, 18, 547. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, Y.; Liu, P.; Ding, Y.; Mu, X.; Liu, X.; Wang, X.; Zhao, M.; Huai, B.; Huang, L.; et al. TaRar1 Is Involved in Wheat Defense against Stripe Rust Pathogen Mediated by YrSu. Front. Plant Sci. 2017, 8, 156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, X.; Zhu, Y.; Zhang, R.; Zhu, Z.; Zhao, T.; Cheng, L.; Gao, L.; Liu, B.; Zhang, X.; Wang, Y. Ionomic and Metabolomic Analyses Reveal the Resistance Response Mechanism to Saline-Alkali Stress in Malus halliana Seedlings. Plant Physiol. Biochem. 2020, 147, 77–90. [Google Scholar] [CrossRef] [PubMed]
- Pilon-Smits, E.A.; Quinn, C.F.; Tapken, W.; Malagoli, M.; Schiavon, M. Physiological Functions of Beneficial Elements. Curr. Opin Plant Biol. 2009, 12, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Deshmukh, R.; Sonah, H.; Patil, G.; Chen, W.; Prince, S.; Mutava, R.; Vuong, T.; Valliyodan, B.; Nguyen, H.T. Integrating Omic Approaches for Abiotic Stress Tolerance in Soybean. Front. Plant Sci. 2014, 244. [Google Scholar] [CrossRef]
- Ahmad, P.; Abd Allah, E.F.; Alyemeni, M.N.; Wijaya, L.; Alam, P.; Bhardwaj, R.; Siddique, K.H.M. Exogenous Application of Calcium to 24-Epibrassinosteroid Pre-Treated Tomato Seedlings Mitigates NaCl Toxicity by Modifying Ascorbate-Glutathione Cycle and Secondary Metabolites. Sci. Rep. 2018, 8, 13515. [Google Scholar] [CrossRef]
- Pankovic, D. Effects of Nitrogen Nutrition on Photosynthesis in Cd-Treated Sunflower Plants. Ann. Bot. 2000, 86, 841–847. [Google Scholar] [CrossRef] [Green Version]
- Sarwar, N.; Malhi, S.S.; Zia, M.H.; Naeem, A.; Bibi, S.; Farid, G. Role of mineral nutrition in minimizing cadmium accumulation by plants. J. Sci. Food Agric. 2010, 90, 925–937. [Google Scholar] [CrossRef]
- Riyazuddin, R.; Verma, R.; Singh, K.; Nisha, N.; Keisham, M.; Bhati, K.K.; Kim, S.T.; Gupta, R. Ethylene: A master regulator of salinity stress tolerance in plants. Biomolecules 2020, 10, 959. [Google Scholar] [CrossRef]
- Astolfi, S.; Zuchi, S. Adequate S supply protects barley plants from adverse effects of salinity stress by increasing thiol contents. Acta Physiol. Plant. 2013, 35, 175–181. [Google Scholar] [CrossRef]
- Usmani, M.M.; Nawaz, F.; Majeed, S. Sulfate-mediated Drought Tolerance in Maize Involves Regulation at Physiological and Biochemical Levels. Sci Rep. 2020, 10, 1147. [Google Scholar] [CrossRef] [Green Version]
- Misra, B.B.; Reichman, S.M.; Chen, S. The guard cell ionome: Understanding the role of ions in guard cell functions. Prog. Biophys. Mol. Biol. 2019, 146, 50–62. [Google Scholar] [CrossRef]
- Abid, M.; Schneider, U.A.; Scheffran, J. Adaptation to Climate Change and Its Impacts on Food Productivity and Crop Income: Perspectives of Farmers in Rural Pakistan. J. Rural Stud. 2016, 47, 254–266. [Google Scholar] [CrossRef]
- Luo, Z.; Kong, X.; Dai, J.; Dong, H. Soil plus foliar nitrogen application increases cotton growth and salinity tolerance. J. Plant Nutr. 2015, 38, 443–455. [Google Scholar] [CrossRef]
- Khan, A.; Ahmad, I.; Shah, A.; Ahmad, F.; Ghani, A.; Nawaz, M.; Shaheen, F.; Fatima, H.U.; Pervaiz, F.; Javed, S.; et al. Amelioration of Salinity Stress in Wheat (Triticum Aestivum) by Foliar Application of Phosphorus. Phyton 2013, 82, 281–287. [Google Scholar]
- Lopez-Delacalle, M.; Camejo, D.M.; García-Martí, M.; Nortes, P.A.; Nieves-Cordones, M.; Martínez, V.; Rubio, F.; Mittler, R.; Rivero, R.M. Using Tomato Recombinant Lines to Improve Plant Tolerance to Stress Combination Through a More Efficient Nitrogen Metabolism. Front. Plant Sci. 2020, 10, 1702. [Google Scholar] [CrossRef] [PubMed]
- Brito, C.; Dinis, L.T.; Ferreira, H.; Coutinho, J.; Moutinho-Pereira, J.; Correia, C.M. Salicylic acid increases drought adaptability of young olive trees by changes on redox status and ionome. Plant Physiol. Biochem. 2019, 141, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Abbas, S.; Javed, M.T.; Ali, Q.; Akram, M.S.; Tanwir, K.; Ali, S.; Chaudhary, H.J.; Iqbal, N. Elucidating Cd-mediated distinct rhizospheric and in planta ionomic and physio-biochemical responses of two contrasting Zea mays L. cultivars. Physiol. Mol. Biol. Plants 2021, 27, 297–312. [Google Scholar] [CrossRef]
- Lee, B.-R.; Zaman, R.; Avice, J.-C.; Ourry, A.; Kim, T.H. Sulfur Use Efficiency is a Significant Determinant of Drought Stress Tolerance in Relation to Photosynthetic Activity in Brassica napus Cultivars. Front. Plant Sci. 2016, 7, 459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hossain, A.; Skalicky, M.; Brestic, M.; Maitra, S.; Ashraful Alam, M.; Syed, M.A.; Hossain, J.; Sarkar, S.; Saha, S.; Bhadra, P.; et al. Consequences and Mitigation Strategies of Abiotic Stresses in Wheat (Triticum aestivum L.) under the Changing Climate. Agronomy 2021, 11, 241. [Google Scholar] [CrossRef]
- Peng, C.Y.; Xu, X.F.; Zhu, H.Y.; Ren, Y.F.; Niu, H.L.; Hou, R.Y.; Wan, X.C.; Cai, H.M. Metabolics and ionomics responses of tea leaves (Camellia sinensis (L.) O. Kuntze) to fluoride stress. Plant Physiol. Biochem. 2021, 158, 65–75. [Google Scholar] [CrossRef]
- Banerjee, A.; Singh, A.; Sudarshan, M.; Roychoudhury, A. Silicon Nanoparticle-Pulsing Mitigates Fluoride Stress in Rice by Fine-Tuning the Ionomic and Metabolomic Balance and Refining Agronomic Traits. Chemosphere 2021, 262, 127826. [Google Scholar] [CrossRef]
- Ahmad, P.; Ahanger, M.A.; Alam, P. Silicon (Si) Supplementation Alleviates NaCl Toxicity in Mung Bean [Vigna radiata (L. ) Wilczek] Through the Modifications of Physio-biochemical Attributes and Key Antioxidant Enzymes. J. Plant Growth Regul. 2019, 38, 70–82. [Google Scholar] [CrossRef]
- Pandit, A.; Rai, V.; Bal, S.; Sinha, S.; Kumar, V.; Chauhan, M.; Gautam, R.K.; Singh, R.; Sharma, P.C.; Singh, A.K.; et al. Combining QTL Mapping and Transcriptome Profiling of Bulked RILs for Identification of Functional Polymorphism for Salt Tolerance Genes in Rice (Oryza sativa L.). Mol. Genet. Genom. 2010, 284, 121–136. [Google Scholar] [CrossRef]
- Sonah, H.; O’Donoughue, L.; Cober, E.; Rajcan, I.; Belzile, F. Identification of Loci Governing Eight Agronomic Traits Using a GBS-GWAS Approach and Validation by QTL Mapping in Soya Bean. Plant Biotechnol. J. 2015, 13, 211–221. [Google Scholar] [CrossRef]
- Deshmukh, R.; Singh, A.; Jain, N.; Anand, S.; Gacche, R.; Singh, A.; Gaikwad, K.; Sharma, T.; Mohapatra, T.; Singh, N. Identification of Candidate Genes for Grain Number in Rice (Oryza sativa L.). Funct. Integr. Genom. 2010, 10, 339–347. [Google Scholar] [CrossRef]
- Sonah, H.; Deshmukh, R.; Chand, S.; Srinivasprasad, M.; Rao, G.; Upreti, H.; Singh, A.; Singh, N.; Sharma, T. Molecular Mapping of Quantitative Trait Loci for Flag Leaf Length and Other Agronomic Traits in Rice (Oryza sativa). Cereal Res. Commun. 2012, 40, 362–372. [Google Scholar] [CrossRef]
- Bentsink, L.; Yuan, K.; Koornneef, M.; Vreugdenhil, D. The Genetics of Phytate and Phosphate Accumulation in Seeds and Leaves of Arabidopsis thaliana, Using Natural Variation. Theor. Appl. Genet. 2003, 106, 1234–1243. [Google Scholar] [CrossRef]
- Payne, K.A.; Bowen, H.C.; Hammond, J.P.; Hampton, C.R.; Lynn, J.R.; Mead, A.; Swarup, K.; Bennett, M.J.; White, P.J.; Broadley, M.R. Natural Genetic Variation in Caesium (Cs) Accumulation by Arabidopsis thaliana. New Phytol. 2004, 162, 535–548. [Google Scholar] [CrossRef]
- Rauh, B.L.; Basten, C.; Buckler IV, E.S. Quantitative Trait Loci Analysis of Growth Response to Varying Nitrogen Sources in Arab. Thaliana. Theor. Appl. Genet. 2002, 104, 743–750. [Google Scholar] [CrossRef]
- Hoekenga, O.A.; Maron, L.G.; Piñeros, M.A.; Cançado, G.M.A.; Shaff, J.; Kobayashi, Y.; Ryan, P.R.; Dong, B.; Delhaize, E.; Sasaki, T.; et al. AtALMT1, Which Encodes a Malate Transporter, Is Identified as One of Several Genes Critical for Aluminum Tolerance in Arabidopsis. Proc. Natl. Acad. Sci. USA 2006, 103, 9738–9743. [Google Scholar] [CrossRef] [Green Version]
- Buescher, E.; Achberger, T.; Amusan, I.; Giannini, A.; Ochsenfeld, C.; Rus, A.; Lahner, B.; Hoekenga, O.; Yakubova, E.; Harper, J.F.; et al. Natural Genetic Variation in Selected Populations of Arabidopsis thaliana Is Associated with Ionomic Differences. PLoS ONE 2010, 5, e11081. [Google Scholar] [CrossRef] [Green Version]
- Tiwari, V.K.; Rawat, N.; Chhuneja, P.; Neelam, K.; Aggarwal, R.; Randhawa, G.S.; Dhaliwal, H.S.; Keller, B.; Singh, K. Mapping of Quantitative Trait Loci for Grain Iron and Zinc Concentration in Diploid a Genome Wheat. J. Hered. 2009, 100, 771–776. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Yang, J.; Li, R.; Shi, L.; Zhang, C.; Long, Y.; Xu, F.; Meng, J. Analysis of Genetic Factors That Control Shoot Mineral Concentrations in Rapeseed (Brassica napus) in Different Boron Environments. Plant. Soil 2009, 320, 255–266. [Google Scholar] [CrossRef]
- Satismruti, K.; Senthil, N.; Vellaikumar, S.; Ranjani, R.V.; Raveendran, M. Plant Ionomics: A Platform for Identifying Novel Gene Regulating Plant Mineral Nutrition. Am. J. Plant Sci. 2013, 4, 1309–1315. [Google Scholar] [CrossRef] [Green Version]
- Chaudhary, J.; Patil, G.B.; Sonah, H.; Deshmukh, R.K.; Vuong, T.D.; Valliyodan, B.; Nguyen, H.T. Expanding Omics Resources for Improvement of Soybean Seed Composition Traits. Front. Plant Sci. 2015, 6, 1021. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Zeng, L.; Tao, Y.; Vuong, T.; Wan, J.; Boerma, R.; Noe, J.; Li, Z.; Finnerty, S.; Pathan, S.M.; et al. Pinpointing Genes Underlying the Quantitative Trait Loci for Root-Knot Nematode Resistance in Palaeopolyploid Soybean by Whole Genome Resequencing. Proc. Natl. Acad. Sci. USA 2013, 110, 13469–13474. [Google Scholar] [CrossRef] [Green Version]
- Deshmukh, R.; Bélanger, R.R. Molecular Evolution of Aquaporins and Silicon Influx in Plants. Funct. Ecol. 2016, 30, 1277–1285. [Google Scholar] [CrossRef]
- Deshmukh, R.K.; Ma, J.F.; Bélanger, R.R. Editorial: Role of Silicon in Plants. Front. Plant Sci. 2017, 8, 1858. [Google Scholar] [CrossRef] [Green Version]
- Kadam, S.; Vuong, T.D.; Qiu, D.; Meinhardt, C.G.; Song, L.; Deshmukh, R.; Patil, G.; Wan, J.; Valliyodan, B.; Scaboo, A.M.; et al. Genomic-Assisted Phylogenetic Analysis and Marker Development for next Generation Soybean Cyst Nematode Resistance Breeding. Plant Sci. 2016, 242, 342–350. [Google Scholar] [CrossRef] [Green Version]
- Induri, B.R.; Ellis, D.R.; Slavov, G.T.; Yin, T.; Zhang, X.; Muchero, W.; Tuskan, G.A.; DiFazio, S.P. Identification of Quantitative Trait Loci and Candidate Genes for Cadmium Tolerance in Populus. Tree Physiol. 2012, 32, 626–638. [Google Scholar] [CrossRef] [Green Version]
- Ziegler, G.; Nelson, R.; Granada, S.; Krishnan, H.B.; Gillman, J.D.; Baxter, I. Genomewide Association Study of Ionomic Traits on Diverse Soybean Populations from Germplasm Collections. Plant Direct 2017, 2, e00033. [Google Scholar] [CrossRef]
- Yang, M.; Lu, K.; Zhao, F.J.; Xie, W.; Ramakrishna, P.; Wang, G.; Du, Q.; Liang, L.; Sun, C.; Zhao, H.; et al. Genome-Wide Association Studies Reveal the Genetic Basis of Ionomic Variation in Rice. Plant Cell 2018, 30, 2720–2740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Norton, G.J.; Deacon, C.M.; Xiong, L.; Huang, S.; Meharg, A.A.; Price, A.H. Genetic mapping of the rice ionome in leaves and grain: Identification of QTLs for 17 elements including arsenic, cadmium, iron and selenium. Plant Soil 2010, 329, 139–153. [Google Scholar] [CrossRef]
- Blair, M.W.; Wu, X.; Bhandari, D.; Astudillo, C. Genetic Dissection of ICP-Detected Nutrient Accumulation in the Whole Seed of Common Bean (Phaseolus vulgaris L.). Front. Plant Sci. 2016, 7, 219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lowry, D.B.; Sheng, C.C.; Zhu, Z.; Juenger, T.E.; Lahner, B.; Salt, D.E.; Willis, J.H. Mapping of ionomic traits in Mimulus guttatus reveals Mo and Cd QTLs that colocalize with MOT1 homologues. PLoS ONE 2012, 7, e30730. [Google Scholar] [CrossRef] [Green Version]
- Gyawali, S.; Otte, M.L.; Chao, S.; Jilal, A.; Jacob, D.L.; Amezrou, R.; Verma, R.P.S. Genome wide association studies (GWAS) of element contents in grain with a special focus on zinc and iron in a world collection of barley (Hordeum vulgare L.). J. Cereal Sci. 2017, 77, 266–274. [Google Scholar] [CrossRef]
- Bastien, M.; Sonah, H.; Belzile, F. Genome Wide Association Mapping of Sclerotinia Sclerotiorum Resistance in Soybean with a Genotyping-by-Sequencing Approach. Plant Genome 2014, 7. [Google Scholar] [CrossRef] [Green Version]
- Tardivel, A.; Sonah, H.; Belzile, F.; O’Donoughue, L.S. Rapid Identification of Alleles at the Soybean Maturity Gene E3 Using Genotyping by Sequencing and a Haplotype-Based Approach. Plant Genome 2014, 7. [Google Scholar] [CrossRef]
- Iquira, E.; Humira, S.; François, B. Association Mapping of QTLs for Sclerotinia Stem Rot Resistance in a Collection of Soybean Plant Introductions Using a Genotyping by Sequencing (GBS) Approach. BMC Plant Biol. 2015, 15, 5. [Google Scholar] [CrossRef] [Green Version]
- Weigel, D.; Nordborg, M. Population Genomics for Understanding Adaptation in Wild Plant Species. Annu. Rev. Genet. 2015, 49, 315–338. [Google Scholar] [CrossRef] [Green Version]
- Atwell, S.; Huang, Y.; Vilhjálmsson, B.; Willmems, G.; Horton, M.; Li, Y.; Meng, D.; Platt, A.; Tarone, A.M.; Hu, T.T.; et al. Genome-wide association study of 107 phenotypes in Arabidopsis thaliana inbred lines. Nature 2010, 465, 627–631. [Google Scholar] [CrossRef]
- Mills, R.F.; Valdes, B.; Duke, M.; Peaston, K.A.; Lahner, B.; Salt, D.E.; Williams, L.E. Functional significance of AtHMA4 C-Terminal Domain in Planta. PLoS ONE 2010, 5, e13388. [Google Scholar] [CrossRef] [Green Version]
- Hanikenne, M.; Talke, I.N.; Haydon, M.J.; Lanz, C.; Nolte, A.; Motte, P.; Kroymann, J.; Weigel, D.; Krämer, U. Evolution of metal hyperaccumulation required cis-regulatory changes and triplication of HMA4. Nature 2008, 453, 391–395. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, M.L.; Schaefer, R.; Dang, P.; Jiang, T.; Chen, C. GWAS and Coexpression Network Reveal Ionomic Variation in Cultivated Peanut. J. Agric. Food Chem. 2019, 67, 12026–12036. [Google Scholar] [CrossRef]
- Descalsota-Empleo, G.I.; Amparado, A.; Inabangan-Asilo, M.A.; Tesoro, F.; Stangoulis, J.; Reinke, R.; Swamy, B.P.M. Genetic Mapping of QTL for Agronomic Traits and Grain Mineral Elements in Rice. Crop J. 2019, 7, 560–572. [Google Scholar] [CrossRef]
- Fikas, A.A.; Dilkes, B.P.; Baxter, I. Multivariate Analysis Reveals Environmental and Genetic Determinants of Element Covariation in the Maize Grain Ionome. Plant Direct 2019, 3, e00139. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Zhong, H.; Meng, X.; Sun, T.; Li, Y.; Pinson, S.R.; Chang, S.K.; Peng, Z. Genome-wide association studies of ionomic and agronomic traits in USDA mini core collection of rice and comparative analyses of different mapping methods. BMC Plant Biol. 2020, 20, 441. [Google Scholar] [CrossRef]
- Li, G.; Nunes, L.; Wang, Y.; Williams, P.N.; Zheng, M.; Zhang, Q.; Zhu, Y. Profiling the ionome of rice and its use in discriminating geographical origins at the regional scale. China J. Environ. Sci. 2013, 25, 144–154. [Google Scholar] [CrossRef]
- Wickramasinghe, W.A.; Anusha, D.L.; Mubiana, V.K.; Blust, R. The Effects of Heavy Metal Concentration on Bio-Accumulation, Productivity and Pigment Content of Two Species of Marine Macro Algae. Sri Lanka J. Aquat. Sci. 2017, 22. [Google Scholar] [CrossRef] [Green Version]
- Ardini, F.; Soggia, F.; Abelmoschi, M.L.; Magi, E.; Grotti, M. Ionomic profiling of Nicotiana langsdorffii wild-type and mutant genotypes exposed to abiotic stresses. Anal. Bioanal. Chem. 2013, 405, 665–677. [Google Scholar] [CrossRef]
- Chen, A.; Hansen, T.H.; Olsen, L.I.; Palmgren, M.; Husted, S.; Schjoerring, J.K.; Persson, D.P. Towards single-cell ionomics: A novel micro-scaled method for multi-element analysis of nanogram-sized biological samples. Plant Methods 2020, 16, 31. [Google Scholar] [CrossRef]
- Ko, J.A.; Furuta, N.; Lim, H. Bin. Quantitative Mapping of Elements in Basil Leaves (Ocimum basilicum) Based on Cesium Concentration and Growth Period Using Laser Ablation ICP-MS. Chemosphere 2018, 190, 368–374. [Google Scholar] [CrossRef]
- Leśniewicz, A.; Zyrnicki, W. Extraction of Trace Elements from Spruce Needles. Commun. Soil Sci. Plant. Anal. 2003, 34, 629–643. [Google Scholar] [CrossRef]
- Galinha, C.; Freitas, M.C.; Pacheco, A.M.G. Enrichment factors and transfer coefficients from soil to rye plants by INAA. J. Radioanal. Nucl. Chem. 2010, 286, 583–589. [Google Scholar] [CrossRef]
- Hamidatou, L.; Slamene, H.; Akhal, T.; Zouranen, B. Concepts, Instrumentation and Techniques of Neutron Activation Analysis. In Imaging and Radioanalytical Techniques in Interdisciplinary Research—Fundamentals and Cutting Edge Applications; InTechOpen: Rijeka, Croatia: London, UK, 2013; pp. 141–178. [Google Scholar] [CrossRef] [Green Version]
- Adomako, D.; Nyarko, B.J.B.; Dampare, S.B.; Serfor-Armah, Y.; Osae, S.; Fianko, J.R.; Akaho, E.H.K. Determination of toxic elements in waters and sediments from River Subin in the Ashanti Region of Ghana. Environ. Monit. Assess. 2008, 141, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Messaoudi, M.; Begaa, S.; Hamidatou, L.; Salhi, M. Determination of selenium in roasted beans coffee samples consumed in Algeria by radiochemical neutron activation analysis method. Radiochim. Acta 2018, 106, 141–146. [Google Scholar] [CrossRef]
- Djingova, R.; Mihaylova, V.; Lyubomirova, V.; Tsalev, D.L. Multielement Analytical Spectroscopy in Plant Ionomics Research. Appl. Spectrosc. Rev. 2013, 48, 384–424. [Google Scholar] [CrossRef]
- Bamford, S.; Wegrzynek, D.; Chinea-Cano, E.; Markowicz, A. Application of X-ray fluorescence techniques for the determination of hazardous and essential trace elements in environmental and biological materials. Nukleonika 2004, 49, 87–95. [Google Scholar]
- Young, L.W.; Westcott, N.D.; Attenkofer, K.; Reaney, M.J.T. A High-Throughput Determination of Metal Concentrations in Whole Intact Arabidopsis thaliana Seeds Using Synchrotron-Based X-Ray Fluorescence Spectroscopy. J. Synchrotron. Radiat. 2006, 13, 304–313. [Google Scholar] [CrossRef]
- Ratcliffe, S.; Jugdaohsingh, R.; Vivancos, J.; Marron, A.; Deshmukh, E.R.; Ma, J.F.; Mitani-Ueno, N.; Robertson, J.; Wills, J.; Boekschoten, M.V.; et al. Identification of a mammalian silicon transporter. Am. J. Physiol. Cell Physiol. 2017, 312, C550–C561. [Google Scholar] [CrossRef] [Green Version]
- Ouellette, S.; Marie-Hélène, G.; Labbé, C.; Laur, J.; Gaudreau, L.; Gosselin, A.; Dorais, M.; Deshmukh, R.K.; Bélanger, R.R. Silicon transporters and effects of silicon amendments in strawberry under high tunnel and field conditions. Front. Plant Sci. 2017, 8, 949. [Google Scholar] [CrossRef]
- Shivaraj, S.M.; Deshmukh, R.; Bhat, J.A.; Sonah, H.; Bélanger, R.R. Understanding aquaporin transport system in eelgrass (Zostera marina L.), an aquatic plant species. Front. Plant Sci. 2017, 8, 1334. [Google Scholar] [CrossRef] [Green Version]
- Fahrni, C.J. Biological applications of X-ray fluorescence microscopy: Exploring the subcellular topography and speciation of transition metals. Curr. Opin. Chem. Biol. 2007, 11, 121–127. [Google Scholar] [CrossRef]
- Qin, Z.; Caruso, J.A.; Lai, B.; Matusch, A.; Becker, J.S. Trace metal imaging with high spatial resolution: Applications in biomedicine. Metallomics 2011, 3, 28–37. [Google Scholar] [CrossRef] [Green Version]
- Sarret, G.; Schroeder, W.H.; Marcus, M.A.; Geoffroy, N.; Manceau, A. Localization and speciation of Zn in mycorrhized roots by µSXRF and µEXAFS. J. Phys. 2003, 107, 1193–1196. [Google Scholar]
- Bulska, E.; Wysocka, I.A.; Wierzbicka, M.L.; Proost, K.; Janssens, K.; Falkenberg, G. In vivo investigation of the distribution and the local speciation of selenium in Allium cepa L. by means of microscopic X-ray absorption near-edge structure spectroscopy and confocal microscopic X-ray fluorescence analysis. Anal Chem. 2006, 78, 7616–7624. [Google Scholar] [CrossRef]
- Meharg, A.A.; Williams, P.N.; Adomako, E.; Lawgali, Y.Y.; Deacon, C.; Villada, A.; Cambell, R.C.; Sun, G.; Zhu, Y.G.; Feldmann, J.; et al. Geographical variation in total and inorganic arsenic content of polished (white) rice. Environ. Sci. Technol. 2009, 43, 1612–1617. [Google Scholar] [CrossRef]
- Takahashi, M.; Nozoye, T.; Kitajima, N.; Fukuda, N.; Hokura, A.; Terada, Y.; Nakai, I.; Ishimaru, Y.; Kobayashi, T.; Nakanishi, H.; et al. In vivo analysis of metal distribution and expression of metal transporters in rice seed during germination process by microarray and X-ray fluorescence imaging of Fe, Zn, Mn, and Cu. Plant Soil 2009, 325, 39–51. [Google Scholar] [CrossRef] [Green Version]
- Ricachenevsky, F.K.; Punshon, T.; Lee, S.; Oliveira, B.H.N.; Trenz, T.S.; Maraschin, F.S.; Hindt, M.N.; Danku, J.; Salt, D.E.; Fett, J.P.; et al. Elemental Profiling of Rice FOX Lines Leads to Characterization of a New Zn Plasma Membrane Transporter, OsZIP7. Front. Plant Sci. 2018, 9, 865. [Google Scholar] [CrossRef] [Green Version]
- Hacisalihoglu, G.; Settles, A.M. Quantification of seed ionome variation in 90 diverse soybean (Glycine max) lines. J. Plant Nutr. 2017, 40, 2808–2817. [Google Scholar] [CrossRef]
- Li, H.; Zhu, Y.; Hu, Y.; Han, W.; Gong, H. Beneficial effects of silicon in alleviating salinity stress of tomato seedlings grown under sand culture. Acta Physiol. Plant. 2015, 37, 71. [Google Scholar] [CrossRef]
- Chen, Z.; Watanabe, T.; Shinano, T.; Ezawa, T.; Wasaki, J.; Kimura, K.; Osaki, M.; Zhu, Y.G. Element interconnections in Lotus japonicus: A systematic study of the effects of element additions on different natural variants. Soil Sci. Plant Nutr. 2008, 55, 91–101. [Google Scholar] [CrossRef] [Green Version]
- Wu, D.; Shen, Q.; Cai, S.; Chen, Z.H.; Dai, F.; Zhang, G. Ionomic Responses and Correlations Between Elements and Metabolites Under Salt Stress in Wild and Cultivated Barley. Plant Cell Physiol. 2013, 54, 1976–1988. [Google Scholar] [CrossRef] [Green Version]
- Begaa, S.; Messaoudi, M.; Benarfa, A. Statistical Approach and Neutron Activation Analysis for Determining Essential and Toxic Elements in Two Kinds of Algerian Artemisia Plant. Biol. Trace Elem. Res. 2021, 199, 2399–2405. [Google Scholar] [CrossRef]
- Barbosa, J.T.P.; Santos, C.M.M.; Dos Santos Bispo, L.; Lyra, F.H.; David, J.M.; Korn, M.D.G.A.; Flores, E.M.M. Bromine, Chlorine, and Iodine Determination in Soybean and its Products by ICP-MS After Digestion Using Microwave-Induced Combustion. Food Anal. Methods 2013, 6, 1065–1070. [Google Scholar] [CrossRef]
- Xu, Z.; Ren, T.; Marowa, P.; You, X.; Lu, X.; Li, Y.; Zhang, C. Establishment of a Cultivation Mode of Glycine soja, the Bridge of Phytoremediation and Industrial Utilization. Agronomy 2020, 10, 595. [Google Scholar] [CrossRef] [Green Version]
- Sanchez, D.H.; Pieckenstain, F.L.; Escaray, F.; Erban, A.; Kraemer, U.; Udvardi, M.K.; Kopka, J. Comparative ionomics and metabolomics in extremophile and glycophytic Lotus species under salt stress challenge the metabolic pre-adaptation hypothesis. Plant Cell Environ. 2011, 34, 605–617. [Google Scholar] [CrossRef]
- Da Costa, L.F.; Tormena, C.F.; Arruda, M.A.Z. Ionomics and lipidomics for evaluating the transgenic (cp4-EPSPS gene) and non-transgenic soybean seed generations. Microchem. J. 2021, 165, 106130. [Google Scholar] [CrossRef]
- Jyothsna, S.; Manjula, G.; Suthari, S.; Rao, A.S.N. Qualitative elemental analysis of selected potential anti-asthmatic medicinal plant taxa using EDXRF technique. Heliyon 2020, 6, e03260. [Google Scholar] [CrossRef]
- Dragut, E.C.; Ouzzani, M.; Madkour, A.; Mohamed, N.; Baker, P.; Salt, D.E. Lonomics Atlas: A tool to explore interconnected ionomic, genomic and environmental data. In Proceedings of the 21st ACM International Conference on Information and Knowledge Management, Maui, HI, USA, 29 October–2 November 2012; pp. 2680–2682. [Google Scholar]
- Zhang, M.; Pinson, S.R.; Tarpley, L.; Huang, X.Y.; Lahner, B.; Yakubova, E.; Baxter, I.; Guerinot, M.L.; Salt, D.E. Mapping and validation of quantitative trait loci associated with concentrations of 16 elements in unmilled rice grain. Theor. Appl. Genet. 2014, 127, 37–165. [Google Scholar] [CrossRef] [Green Version]
- Yu, D.; Danku, J.M.; Baxter, I.; Kim, S.; Vatamaniuk, O.K.; Vitek, O.; Ouzzani, M.; Salt, D.E. High-resolution genome-wide scan of genes, gene-networks and cellular systems impacting the yeast ionome. BMC Genom. 2012, 13, 623. [Google Scholar] [CrossRef] [Green Version]
- Maillard, A.; Sorin, E.; Etienne, P.; Diquélou, S.; Koprivova, A.; Kopriva, S.; Arkoun, M.; Gallardo, K.; Turner, M.; Cruz, F.; et al. Non-specific root transport of nutrient gives access to an early nutritional indicator: The case of sulfate and molybdate. PLoS ONE 2016, 11, e0166910. [Google Scholar] [CrossRef]
- Sasaki, A.; Yamaji, N.; Ma, J.F. Transporters involved in mineral nutrient uptake in rice. J. Exp. Bot. 2016, 67, 3645–3653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, J.A.; Williams, L.E. Transition metal transporters in plants. J. Exp. Bot. 2003, 54, 2601–2613. [Google Scholar] [CrossRef]
- Williams, L.E.; Pittman, J.K.; Hall, J.L. Emerging mechanisms for heavy metal transport in plants. Biochim. Biophys. Acta Biomembr. 2000, 1465, 104–126. [Google Scholar] [CrossRef]
- Axelsen, K.B.; Palmgren, M.G. Evolution of substrate specificities in the P-type ATPase superfamily. J. Mol. Evol. 1998, 46, 84–101. [Google Scholar] [CrossRef] [PubMed]
- Papoyan, A.; Leon, V.K. Identification of Thlaspi caerulescens genes that may be involved in heavy metal hyperaccumulation and tolerance. Characterization of a novel heavy metal transporting ATPase. Plant Physiol. 2004, 136, 3814–3823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, R.; Bashir, K.; Ishimaru, Y.; Nishizawa, N.K.; Nakanishi, H. The role of heavy-metal ATPases, HMAs, in zinc and cadmium transport in rice. Plant Signal. Behav. 2012, 7, 1605–1607. [Google Scholar] [CrossRef] [PubMed]
- Gravot, A.; Lieutaud, A.; Verret, F.; Auroy, P.; Vavasseur, A.; Richaud, P. AtHMA3, a plant P1B-ATPase, functions as a Cd/Pb transporter in yeast. FEBS Lett. 2004, 561, 22–28. [Google Scholar] [CrossRef] [Green Version]
- Morel, M.; Crouzet, J.; Gravot, A.; Auroy, P.; Leonhardt, N.; Vavasseur, A.; Richaud, P. AtHMA3, a P1B-ATPase allowing Cd/Zn/co/Pb vacuolar storage in Arabidopsis. Plant Physiol. 2009, 149, 894–904. [Google Scholar] [CrossRef] [Green Version]
- Eren, E.; Argüello, J.M. Arabidopsis HMA2, a divalent heavy metal-transporting PIB-type ATPase, is involved in cytoplasmic Zn2+ homeostasis. Plant Physiol. 2004, 136, 3712–3723. [Google Scholar] [CrossRef] [Green Version]
- Mills, R.F.; Krijger, G.C.; Baccarini, P.J.; Hall, J.L.; Williams, L.E. Functional expression of AtHMA4, a P1B-type ATPase of the Zn/Co/Cd/Pb subclass. Plant J. 2003, 35, 164–176. [Google Scholar] [CrossRef]
- Mills, R.F.; Krijger, G.C.; Baccarini, P.J.; Hall, J.L.; Williams, L.E. The plant P1B-type ATPase AtHMA4 transports Zn and Cd and plays a role in detoxification of transition metals supplied at elevated levels. FEBS Lett. 2005, 579, 783–791. [Google Scholar] [CrossRef] [Green Version]
- Verret, F.; Gravot, A.; Auroy, P.; Preveral, S.; Forestier, C.; Vavasseur, A.; Richaud, P. Heavy metal transport by AtHMA4 involves the N-terminal degenerated metal binding domain and the C-terminal His11 stretch. FEBS Lett. 2005, 579, 1515–1522. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, M.; Bashir, K.; Inoue, H.; Takahashi, M.; Nakanishi, H.; Nishizawa, N.K. Accumulation of starch in Zn-deficient rice. Rice 2012, 5, 9. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, R.; Ishimaru, Y.; Shimo, H.; Ogo, Y.; Senoura, T.; Nishizawa, N.K.; Nakanishi, H. The OsHMA2 transporter is involved in root-to-shoot translocation of Zn and Cd in rice. Plant Cell Environ. 2012, 35, 1948–1957. [Google Scholar] [CrossRef]
- Miyadate, H.; Adachi, S.; Hiraizumi, A.; Tezuka, K.; Nakazawa, N.; Kawamoto, T.; Katou, K.; Kodama, I.; Sakurai, K.; Takahashi, H.; et al. OsHMA3, a P1B-type of ATPase affects root-to-shoot cadmium translocation in rice by mediating efflux into vacuoles. New Phytol. 2011, 189, 190–199. [Google Scholar] [CrossRef]
- Deng, F.; Yamaji, N.; Xia, J.; Ma, J.F. A member of the heavy metal P-type ATPase OsHMA5 is involved in xylem loading of copper in rice. Plant. Physiol. 2013, 163, 1353–1362. [Google Scholar] [CrossRef] [Green Version]
- Zhiguo, E.; Li, T.T.; Chen, C.; Wang, L. Genome-wide survey and expression analysis of P-1B-ATPases in rice, maize and sorghum. Rice Sci. 2018, 25, 208–217. [Google Scholar] [CrossRef]
- Mäser, P.; Thomine, S.; Schroeder, J.I.; Ward, J.M.; Hirschi, K.; Sze, H.; Talke, I.N.; Amtmann, A.; Maathuis, F.J.; Sanders, D.; et al. Phylogenetic relationships within cation transporter families of Arabidopsis. Plant Physiol. 2001, 126, 1646–1667. [Google Scholar] [CrossRef] [Green Version]
- Thomine, S.; Wang, R.; Ward, J.M.; Crawford, N.M.; Schroeder, J.I. Cadmium and iron transport by members of a plant metal transporter family in Arabidopsis with homology to Nramp genes. Proc. Natl. Acad. Sci. USA 2000, 97, 4991–4996. [Google Scholar] [CrossRef] [Green Version]
- Kolaj-Robin, O.; Russell, D.; Hayes, K.A.; Pembroke, J.T.; Soulimane, T. Cation Diffusion Facilitator family: Structure and function. FEBS Lett. 2015, 589, 1283–1295. [Google Scholar] [CrossRef]
- Montanini, B.; Blaudez, D.; Jeandroz, S.; Sanders, D.; Chalot, M. Phylogenetic and functional analysis of the cation diffusion facilitator (CDF) family: Improved signature and prediction of substrate specificity. BMC Genom. 2007, 8, 107. [Google Scholar] [CrossRef] [Green Version]
- Ajeesh Krishna, T.P.; Maharajan, T.; Victor Roch, G.; Ignacimuthu, S.; Antony Ceasar, S. Structure, Function, Regulation and Phylogenetic Relationship of ZIP Family Transporters of Plants. Front. Plant Sci. 2020, 11, 662. [Google Scholar] [CrossRef]
- Tiong, J.; McDonald, G.; Genc, Y.; Shirley, N.; Langridge, P.; Huang, C.Y. Increased expression of six ZIP family genes by zinc (Zn) deficiency is associated with enhanced uptake and root−to−shoot translocation of Zn in barley (Hordeum vulgare). New Phytol. 2015, 207, 1097–1109. [Google Scholar] [CrossRef] [PubMed]
- Assuncao, A.G.; Herrero, E.; Lin, Y.F.; Huettel, B.; Talukdar, S.; Smaczniak, C.; Immink, R.G.; van Eldik, M.; Fiers, M.; Schat, H.; et al. Arabidopsis thalianatranscription factors bZIP19 and bZIP23 regulate the adaptation to zinc deficiency. Proc. Natl. Acad. Sci. USA 2010, 107, 10296–10301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, L.; Sun, L.; Tan, L. Progress in ZIP transporter gene family in rice. Yi Chuan 2018, 40, 33–43. [Google Scholar] [CrossRef] [PubMed]
- Santos, A.L.D.; Chaves-Silva, S.; Yang, L. Global analysis of the MATE gene family of metabolite transporters in tomato. BMC Plant Biol. 2017, 17, 185. [Google Scholar] [CrossRef]
- Wang, L.; Bei, X.; Gao, J.; Li, Y.; Yan, Y.; Hu, Y. The similar and different evolutionary trends of MATE family occurred between rice and Arabidopsis thaliana. BMC Plant Biol. 2016, 16, 207. [Google Scholar] [CrossRef] [Green Version]
- Zhu, H.; Wu, J.; Jiang, Y.; Jin, J.; Zhou, W.E.I.; Wang, Y.U.; Han, G.; Zhao, Y.; Cheng, B. Genome wide analysis of MATE-type gene family in maize reveals microsynteny and their expression patterns under aluminum treatment. J. Genet. 2016, 95, 691–704. [Google Scholar] [CrossRef]
- Zhao, J.; Dixon, R.A. MATE transporters facilitate vacuolar uptake of epicatechin 3′-O-glucoside for proanthocyanidin biosynthesis in Medicago truncatula and Arabidopsis. Plant Cell 2009, 21, 2323–2340. [Google Scholar] [CrossRef] [Green Version]
- Ogawa, I.; Nakanishi, H.; Mori, S.; Nishizawa, N.K. Time course analysis of gene regulation under cadmium stress in rice. Plant Soil 2009, 325, 97–108. [Google Scholar] [CrossRef] [Green Version]
- Guidotti, G. ATP transport and ABC proteins. Chem. Biol. 1996, 3, 703–706. [Google Scholar] [CrossRef] [Green Version]
- Garcia, O.; Philippe, B.; Cyrille, F.; Elie, D. Inventory and comparative analysis of rice and Arabidopsis ATP-binding cassette (ABC) systems. J. Mol. Biol. 2004, 343, 249–265. [Google Scholar] [CrossRef]
- Kuromori, T.; Eriko, S.; Kazuo, S. Arabidopsis mutants of AtABCG22, an ABC transporter gene, increase water transpiration and drought susceptibility. Plant J. 2011, 67, 885–894. [Google Scholar] [CrossRef]
- Rice, A.J.; Aekyung, P.; Heather, W.P. Diversity in ABC transporters: Type I, II and III importers. Crit. Rev. Biochem. Mol. Biol. 2014, 49, 426–437. [Google Scholar] [CrossRef] [Green Version]
- Boominathan, R.; Doran, P.M. Cadmium tolerance and antioxidative defenses in hairy roots of the cadmium hyperaccumulator, Thlaspi caerulescens. Biotechnol. Bioengin. 2003, 83, 158–167. [Google Scholar] [CrossRef]
- Park, J.; Song, W.Y.; Ko, D.; Eom, Y.; Hansen, T.H.; Schiller, M.; Lee, T.G.; Martinoia, E.; Lee, Y. The phytochelatin transporters AtABCC1 and AtABCC2 mediate tolerance to cadmium and mercury. Plant J. 2012, 69, 278–288. [Google Scholar] [CrossRef]
- Larsen, P.B.; Geisler, M.J.; Jones, C.A.; Williams, K.M.; Cancel, J.D. ALS3 encodes a phloem-localized ABC transporter-like protein that is required for aluminum tolerance in Arabidopsis. Plant J. 2005, 41, 353–363. [Google Scholar] [CrossRef]
- Sonah, H.; Deshmukh, R.K.; Labbé, C.; Bélanger, R.R. Analysis of aquaporins in Brassicaceae species reveals high-level of conservation and dynamic role against biotic and abiotic stress in canola. Sci. Rep. 2017, 7, 2771. [Google Scholar] [CrossRef]
- Ma, J.F.; Yamaji, N.; Mitani, N.; Tamai, K.; Konishi, S.; Fujiwara, T.; Katsuhara, M.; Yano, M. An efflux transporter of silicon in rice. Nature 2007, 448, 209–212. [Google Scholar] [CrossRef]
- Vivancos, J.; Deshmukh, R.; Grégoire, C.; Rémus-Borel, W.; Belzile, F.; Bélanger, R.R. Identification and characterization of silicon efflux transporters in horsetail (Equisetum arvense). J. Plant Physiol. 2016, 200, 82–89. [Google Scholar] [CrossRef]
- Chiba, Y.; Mitani, N.; Yamaji, N.; Ma, J.F. HvLsi1 is a silicon influx transporter in barley. Plant J. 2009, 57, 810–818. [Google Scholar] [CrossRef]
- Deshmukh, R.K.; Vivancos, J.; Guérin, V.; Sonah, H.; Labbé, C.; Belzile, F.; Bélanger, R.R. Identification and functional characterization of silicon transporters in soybean using comparative genomics of major intrinsic proteins in Arabidopsis and rice. Plant Mol. Biol. 2013, 83, 303–315. [Google Scholar] [CrossRef]
- Mitani, N.; Chiba, Y.; Yamaji, N.; Ma, J.F. Identification and characterization of maize and barley Lsi2-like silicon efflux transporters reveals a distinct silicon uptake system from that in rice. Plant Cell 2009, 21, 2133–2142. [Google Scholar] [CrossRef] [Green Version]
- Montpetit, J.; Vivancos, J.; Mitani-Ueno, N.; Yamaji, N.; Rémus-Borel, W.; Belzile, F.; Ma, J.F.; Bélanger, R.R. Cloning, functional characterization and heterologous expression of TaLsi1, a wheat silicon transporter gene. Plant Mol. Biol. 2012, 79, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Salt, D.E. Update on ionomics. Plant Physiol. 2004, 136, 2451–2456. [Google Scholar] [CrossRef] [Green Version]
- Arbona, V.; Manzi, M.; Ollas, C.D.; Gómez-Cadenas, A. Metabolomics as a tool to investigate abiotic stress tolerance in plants. Int. J. Mol. Sci. 2013, 14, 4885–4911. [Google Scholar] [CrossRef] [PubMed]
- Bajwa, A.A.; Farooq, M.; Nawaz, A. Seed priming with sorghum extracts and benzyl aminopurine improves the tolerance against salt stress in wheat (Triticum aestivum L.). Physiol. Mol. Biol. Plants 2018, 24, 239–249. [Google Scholar] [CrossRef]
- Bowne, J.B.; Erwin, T.A.; Juttner, J.; Schnurbusch, T.; Langridge, P.; Bacic, A.; Roessner, U. Drought responses of leaf tissues from wheat cultivars of differing drought tolerance at the metabolite level. Mol. Plant 2012, 5, 418–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.M.; Shi, W.M. Quantitative characterization of nitrogen regulation of OsAMT1; 1, OsAMT1; 2, and OsAMT2; 2 expression in rice seedlings. Russ. J. Plant Physiol. 2006, 53, 837–843. [Google Scholar] [CrossRef]
- Narayanan, S.; Tamura, P.J.; Roth, M.R.; Prasad, P.V.; Welti, R. Wheat leaf lipids during heat stress: I. High day and night temperatures result in major lipid alterations. Plant Cell Environ. 2016, 39, 787–803. [Google Scholar] [CrossRef] [Green Version]
- Pant, B.D.; Burgos, A.; Pant, P.; Cuadros-Inostroza, A.; Willmitzer, L.; Scheible, W.R. The transcription factor PHR1 regulates lipid remodeling and triacylglycerol accumulation in Arabidopsis thaliana during phosphorus starvation. J. Exp. Bot. 2015, 66, 1907–1918. [Google Scholar] [CrossRef] [Green Version]
- Skirycz, A.; De Bodt, S.; Obata, T.; De Clercq, I.; Claeys, H.; De Rycke, R.; Andriankaja, M.; Van Aken, O.; van Breusegem, F.; Fernie, A.R.; et al. Developmental stage specificity and the role of mitochondrial metabolism in the response of Arabidopsis leaves to prolonged mild osmotic stress. Plant Physiol. 2010, 152, 226–244. [Google Scholar] [CrossRef] [Green Version]
- Zhu, G.; Wang, S.; Huang, Z.; Zhang, S.; Liao, Q.; Zhang, C.; Lin, T.; Qin, M.; Peng, M.; Yang, C.; et al. Rewiring of the fruit metabolome in tomato breeding. Cell 2018, 172, 249–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ichihashi, Y.; Date, Y.; Shino, A.; Shimizu, T.; Shibata, A.; Kumaishi, K.; Funahashi, F.; Wakayama, K.; Yamazaki, K.; Umezawa, A.; et al. Multi-omics analysis on an agroecosystem reveals the significant role of organic nitrogen to increase agricultural crop yield. Proc. Natl. Acad. Sci. USA 2020, 117, 14552–14560. [Google Scholar] [CrossRef] [PubMed]
- Moreno, J.C.; Martinez-Jaime, S.; Kosmacz, M.; Sokolowska, E.M.; Schulz, P.; Fischer, A.; Luzarowska, U.; Havaux, M.; Skirycz, A. A multi-OMICs approach sheds light on the higher yield phenotype and enhanced abiotic stress tolerance in tobacco lines expressing the carrot lycopene β-cyclase1 gene. Front. Plant Sci. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Xiangchi, Z.; Chao, L.; Wen, W.; Cheng, L. Ionomic and metabolic responses to drought stress in elite wheat seedlings under two phosphorus levels. Res. Sq. 2021, 1–20. [Google Scholar] [CrossRef]
- Huang, X.Y.; Salt, D.E. Plant ionomics: From elemental profiling to environmental adaptation. Mol. Plant 2016, 9, 787–797. [Google Scholar] [CrossRef] [Green Version]
- Guo, H.; Li, S.; Min, W.; Ye, J.; Hou, Z. Ionomic and transcriptomic analyses of two cotton cultivars (Gossypium hirsutum L.) provide insights into the ion balance mechanism of cotton under salt stress. PLoS ONE 2019, 14, e0226776. [Google Scholar] [CrossRef] [Green Version]
- Guo, R.; Shi, L.; Yan, C.; Zhong, X.; Gu, F.; Liu, Q.; Xia, X.; Li, H. Ionomic and metabolic responses to neutral salt or alkaline salt stresses in maize (Zea mays L.) seedlings. BMC Plant Biol. 2017, 17, 41. [Google Scholar] [CrossRef] [Green Version]
- Ho, W.W.H.; Hill, C.B.; Doblin, M.S.; Shelden, M.C.; van de Meene, A.; Rupasinghe, T.; Bacic, A.; Roessner, U. Integrative multi-omics analyses of barley rootzones under salinity stress reveal two distinctive salt tolerance mechanisms. Plant. Comm. 2020, 1, 100031. [Google Scholar] [CrossRef]
- Groskinsky, D.K.; Syaifullah, S.J.; Roitsch, T. Integration of multi-omics techniques and physiological phenotyping within a holistic phenomics approach to study senescence in model and crop plants. J. Exp. Bot. 2018, 69, 825–844. [Google Scholar] [CrossRef] [Green Version]
- Raza, A.; Razzaq, A.; Mehmood, S.S.; Hussain, M.A.; Wei, S.; He, H.; Zaman, Q.U.; Xuekun, Z.; Hasanuzzaman, M. Omics: The way forward to enhance abiotic stress tolerance in Brassica napus L. GM Crop. Food 2021, 12, 251–281. [Google Scholar] [CrossRef]
- Fischer, N.; Efferth, T. The impact of “omics” technologies for grapevine (Vitis vinifera) research. J. Berr. Res. 2020, 1–15. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Total Ionome Regulatory Genes | Primary Gene Names | Target Elements | Tissues | References |
|---|---|---|---|---|---|
| A. thaliana | 136 | CNGC10, FRO2, MT1A, CPNIFS, LPCAT1, PHO1;H3, PCR2, BTSL2, FYVE1, CIPK23, ABCC1, CCC, NIP3;1, AtNRT1.5/ AtNPF7.3, AtMSA1, myb72, NAS4, PEN3, AtIRT3, AtAPR2, AtHMA5, AtAX1, MYB62, AtCITF1, BTSL1, PHT1;9, NIP6;1, AtNRAMP1, VIT1, SOS1/NHX7, ABH1, AtbZIP23, AtHMA4, AtHAC1, AtNRAMP3, AtMT4b, MOT1, FRU, ESB1, PHT1;5, PHO2, ZAT11, FPN1, PHT1;4, AtMTP11, AtMT4a, CNGC3, AtMTP1, BOR1, VIH2, TSC10a, NIP7, FRD3/MAN1, AtZIP1, myb10, CAX2, mir169a, AtCTL1, BTS, AtATPS1, bHLH34, PHO1, ZIFL2, PYE, AHA4, CAX3, SULTR3;1, bHLH38, bHLH39, AtMTP8, AtMTP3, MGT6, BOR2, GA1, AtHKT1;1, NIP5;1, HAK5, bHLH104, OPT3, IRT1, GSH1, YSL1, AtPHR1, AtHMA2, AtHMA3, CBL10, AtbZIP19, HMA1, NaKR1, ACR2, FPN2, MGT7, ZIF1, VIH1, CNGC2/DND1, APG5, AtSPL7, COPT5, SOS2, AtMfl1, PHT1;1, PCS1, CNGC1, YSL3, ILR3, AtTSB1, AtMYB36, COPT1, CPR5, AtNRAMP3 | K, Ca2+, Mg, Fe, Cd, Zn, As, Se, S, Zn, P, Mn, Co, K, Cd, Na, NO3-, Mo, Cu, B, Ni, Rb, Cs, Li, Sr | Root, shoot, leaf, seed | [29,31,32,33,34,35,36,37,38,39] |
| O. sativa | 141 | OsMTP9, OsHKT1;5, AKT1, OsHORZ1, OsMGT1, OsHAC4, OsHMA4, OsNIP1;1, OsYSL2, OsYSL15, LSI1, OsMTP8.2, OsPHO1;2, OsPT2, OsRab6a, OsMTP8.1, OsMIT, OsNAS2, OsPHR2, OsHAK1, OsVIT1, OsYSL9, OsYSL16, OsHMA5, OsZIP3, OsABCC1, OsRMC, OsPCS1, OsZIP5, OsHRZ2, OsPHO2, OsPCS2, SPDT, OsHMA2, OsHKT2;1, TPKb, OsPHF1, OsHMA3, NRAMP5, OsMOT1;1, OsPRI1, OsNIP3;2, OsNIP3;3, OsATX1, OsVIT2, ZIFL12, OsMIR, OsbHLH133, OsBOR1, OsHRZ1 | Mn, Na+, K, Fe, Mg, Ás, Cu, Mn, Se, P, Zn, Cs, Cd, B | Leaf, shoot, root, seed | [40,41,42,43,44,45,46,47,48,49,50,51,52] |
| T. aestivum | 267 | TaIPK1, Ta-PHR1, TaABCC13, HKT2;1 | Fe, Zn, P, Na+, Ca2+ | Seed, root, shoot | [53,54,55,56,57] |
| Z. mays | 152 | ZmHKT1, YS1, YS3, ZmHAK5, TLS1 | Na+, Fe, K, B | Leaf, root, shoot, anthers | [58,59,60,61,62,63] |
| M. truncatula | 176 | MtMOT1.2, MtNramp1, MtMOT1.3, MtCOPT1, MtMTP2, MtZIP6 | Mo, Fe, Cu, Zn | Nodules | [64,65,66,67,68,69] |
| Candidate Gene | Species Name | Role in Stress | Function | Related Elements | Reference |
|---|---|---|---|---|---|
| FRD3 Ferric Reductase Defective3 | A. thaliana | Biotic | Citrate transporter | High Mn2+ and Co2+ | [107,108,109] |
| HKT1;1 (High-affinity K+ transporter 1) | A. thaliana | Abiotic | Sodium transporter | High Na+ | [110,111] |
| APR2 (Adenylylphosphosulfate reductase) | A. thaliana | Sulfur Assimilation | 50-Phosphosulfate reductase | High sulfate, S2−, and Se2− | [25,112] |
| MOT1 (Molybdate transporter 1) | A. thaliana | Biotic and abiotic | Molybdenum transporter | Low Mo2+ | [113,114] |
| FPN2 (Ferroportin) | A. thaliana | Biotic and abiotic | Ferroportin metal efflux protein | High Co2+ | [30] |
| ESB1 (Enhanced suberin1) | A. thaliana | Biotic and abiotic | Dirigent domain-containing protein | Low Ca2+ and Mn2+; high Na+, S2−, K+, As3+, Se2−, and Mo2+ | [115,116] |
| NaKR1 (Sodium potassium root defective 1) | A. thaliana | Biotic and abiotic | Metal binding protein | High Na+, K+, and Rb+ | [117] |
| SGN1 (Schengen3) | A. thaliana | Biotic and abiotic | Kinase | High Mg2+ | [117,118] |
| CPR5 (Constitutive Expresser of Pathogenesis Related Genes 5) | A. thaliana | Biotic and abiotic | Constitutive expression of pathogen resistance | Low K+ | [119] |
| TSC10A (Ketosphinganine reductase) | A. thaliana | Biotic and abiotic | 3-Ketodihydrosphinganine reductase | Low Mg2+, Ca2+, Fe2+, and Mo2+; high Na+, K+, and Rb+ | [120] |
| HMA3 (Heavy Metal ATPase) | A. thaliana | Biotic and abiotic | Heavy metal ATPase | Low Cd2+ | [121] |
| ATPS1 (ATP sulfurylase 1) | A. thaliana | Biotic and abiotic | ATP sulfurylase | High sulfate | [122] |
| SGN3 (Schengen3) | Hordeum vulgare | Biotic and abiotic | Receptor-like kinase | Low K+; high Mg2+ | [123] |
| ATQ1/HAC1 (Arsenate reductase QTL1/High Arsenic Content 1) | A. thaliana | Biotic and abiotic | Arsenate reductase | High As3+ | [124] |
| MYB36 (MYB Domain protein 36) | A. thaliana | Biotic and abiotic | MYB domain transcription factor | Low Ca2+, Mn2+, and Fe2+; high Na+, Mg2+, and Zn2+ | [28] |
| GSL (Glucosinolate) | Arabidopsis halleri and Brassicaceae | Biotic and abiotic | Increase in phloem of young leaves against Myzuspersicae protection in Arabidopsis | Zn2+, Cd2+ | [125] |
| AD (Alcohol dehydrogenase) | Cyamopsis tetragonoloba | Biotic stress | Up-regulate Zn-binding AD and making pathogen resistant cultivar | Zn2+ | [126] |
| CA (Carbonic anhydrase) | A. thaliana | Biotic stress | Act as salicylic acid binding protein | Zn2+ | [127] |
| MT (Metallothionein-like protein) | Fagus grandifolia | Biotic stress | Resistant to fungal infection | Zn2+ | [128] |
| Znf (Zn finger) | T. aestivum | Biotic stress | Key in the R-gene-specific resistance of plants to pathogens | Zn2+ | [129] |
| NHX8/ZTP1 (Na+/H+ transporter/Zinc transporter protein) | Malus halliana | Abiotic stress | Saline-alkali stress resistance | High Na+ and Fe2+ | [130] |
| AKT1, MRS2-4, and ZTP29 | M. halliana | Abiotic stress | Saline-alkali stress resistance | Low K+, Mg2+, and Zn2+ | [130] |
| ANT, ATP2A, CALM and SOS2 | M. halliana | Abiotic stress | Saline-alkali stress resistance | Ca2+ signal transduction | [130] |
| Plant Species | Number of Genotypes | Plant Tissue Analyzed | Ionomic Tool Used for the Elemental Profiling | Elements Analyzed | Number of Most Significant Loci Associated with Ionomic Trait | Reference |
|---|---|---|---|---|---|---|
| Soybean | 1653 | Seeds | ICP-MS | K, P, Zn, Ca2+, Mg, Na+, S, Ni, Fe, Co, Al, Cu, Cd, Mo Se, Rb | 573 unique SNPs | [169] |
| Rice | 529 | Seeds | ICP-MS | Ca2+, P, N, Na+, Mg, K, Zn, Cu, B, Cr, Mo, Cd, Mn, As, Pb, Co | 72 loci | [170] |
| Rice | 79 | Seeds | Flow injection spectrophotometer, and ICPMS | P, Si, Fe, Zn, Cu, Mn, Ni, Pb, Mo, As, Co, Cd, Al, Se | 36 QTLs | [171] |
| Common bean | 84 | Seeds | ICP-AES | Fe, S, Ca2+, Mg, Cu, Zn, Ni, Mo, Mn, B, Cd, Co, | 21 QTLs | [172] |
| Monkey flower | 168 | Leaves | ICP-MS | K, P, Ca 2+ , Na + , S, Zn, Mg, Fe, Mn, Cu, Rb, B, Sr, Se, As, Cd, Ni, Li, Mo | 7 QTL | [173] |
| Barley | 336 | Grains | ICP-MS | P, S, Si, Na+, Fe, Ba, Mn, Mg, Ca2+, Sr, Zn, Cu | 15 SNP loci | [174] |
| Plant species | Ion | Medium | Tissue Used | Platform | Study | Reference |
|---|---|---|---|---|---|---|
| A. thaliana | B, Ca2+, Mg, Mn, Fe, Cu, Zn, P, Co, Mo, As, Cd | Pot assay | Leaf | ICP-MS | Fe and P homeostasis | [24] |
|
Rice (O. sativa) | 6 elements K, Ca2+, Mn, Zn, Cu, Fe | Lab/tissue culture condition | Seed | SXRF | Characterization of a new Zn plasma membrane transporter, OsZIP7 | [208] |
| Apple (M. halliana) | Ca2+, Fe, Zn, Mg, Mn, Na+, K, Cu, Cl | Hydroponics | Seedlings | ICP-OES, Ion exchange chromatography, LC-MS | To study saline-alkali stress in M. halliana seedlings | [130] |
| Soybean (G.max) | B, Na+, Mg, Al, P, S, K, Ca2+, Mn, Fe, Co, Ni, Cu, Zn, As, Se, Rb, Mo, and Cd | Field conditions | Seed | ICP-MS and SoySNP50k chip data | Identified candidate SNPs controlling elemental accumulation as well as lines with extreme elemental accumulation phenotypes. | [169] |
| Soybean (G. max) | Se, Cu, Fe, Mn | Field conditions | Seed | ICP-MS | C/N and other elements | [209] |
| Tomato (Solanum lycopersicum) | Na+ and Cl | Sand culture | Root, stem and leaf | AAS | Role of Si in mitigating abiotic stress | [210] |
| Lotus (Lotus japonicus) | 15 elements B, Cd, Ca2+, Cu, Cs, Fe, Pb, Mg, Mn, Mo, Ni, K, Na+, Sr and Zn | Hydroponics | Seeds | ICP-MS | To investigate the accumulation of 15 elements in shoots of mutants of L. japonicus | [211] |
| Barley (H. vulgare) | Na+, K, Ca2+, Mg, P, S, Cu, Fe, Mn, and Zn | Hydroponics and pot conditions | Germination/seedlings | ICP-OES | Salinity stress in barley | [212] |
|
Breckland wormwood (Artemisia campestris) | 21 elements Higher: K, Ca2+, Fe, Na+ As, Ba, Br, Ca, Ce, Co, Cr, Cs, Eu, Fe, Hf, K, La, Na, Rb, Sb, Sc, Sm, Sr, Yb, Zn | Field conditions | Leaf | NAA | Determining essential and toxic elements | [213] |
| Tobacco (N. angsdorffii) | 29 elements Ba, Bi, Ca2+, Cd, Co, Cr, Cu, Eu, Fe, Ga, K, Li, Mg, Mn, Mo, Na+, P, Pb, Pt, Rb, S, Sb, Sn, Sr, Te, V, W, Y, and Zn | Agar medium | Root, stem and leaves | ICP-AES/MS | Ionomic profiling of N. langsdorffii wild-type and mutant genotypes exposed to abiotic stresses | [188] |
|
Soybean (Glycine soja (L.) Merr. and G. max) | Br, Cl, and I | Soil | Seeds | ICP-MS | Determination of bromine, chlorine, and iodine in soybean | [214] |
| A. halleri | Zn, Cd | Pot conditions | Leaf | HPLC, ICP-AES | To investigate the effects of the heavy metals Zn and/or Cd on aphid in A. halleri | [125] |
| A. thaliana | Zn | Growth chamber | Leaf tissue | LC-MS/MS | Role in plant immunity | [127] |
| Wheat (T. aestivum) | Zn | Pot conditions | Seedlings | HPLC-MS | key in the R-gene-specific resistance of plants to pathogens | [129] |
| Soybean (G. soja) | Na+, K, Ca2+, and Mg | Field | Roots, shoots, leaves, seeds, and capsules | AAS | The K-Na ratio of seed, leaf, shoot, and capsule were all >1 in the wild | [215] |
| Lotus Extremophile (Lotus creticus) and glycophytic (Lotus corniculatus and Lotus tenuis. L.) | Ca2+, B, P, Mn, S, Zn, Mg, Fe, Cl, K, Na+ (elements) Proline, serine, sucrose, Glyceric acid, citric acid, succinic acid, erythronic acid (metabolites) | Pot and filed conditions | Complete shoots (pooling leaves, petioles, and stems) | Gas chromatography coupled to electron impact ionization-time of flight-mass spectrometry (GC/EI-TOF-MS) | Glyphocites adapted well under salinity | [216] |
| Soybean (G. max) | B, Na+, Mg, Al, P, S, K, Ca2+, Mn, Fe, Co, Ni, Cu, Zn, As, Se, Rb, Mo, and Cd | Filed conditions | Seeds | ICP-MS | Ionomic screening for identifying mutant soybean lines with altered elemental composition | [18] |
| Soybean (G. max) | B, Al, Mn, Fe, Co, Ni, Cu, Zn, Sr, Mo, and Ba, | Soil | Seeds | ICP-MS and NMR | Elemental and lipid profiling of transgenic (cp4-EPSPS gene) and wild type soybean seed generations | [217] |
| Acacia catechu , Argemone mexicana, Aegle marmelos, Datura metel, Phyllanthus emblica, Sapindus emarginatus, Senna occidentalis | 13 elements Major elements found: K, Ca2+, Cl, S, P Lower elements: Cu, Zn, Fe, Mn, Se, Br, Rb, Sr | Field conditions | Leaves, bark, fruits | XRF | Phytomedicine | [218] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ali, S.; Tyagi, A.; Bae, H. Ionomic Approaches for Discovery of Novel Stress-Resilient Genes in Plants. Int. J. Mol. Sci. 2021, 22, 7182. https://doi.org/10.3390/ijms22137182
Ali S, Tyagi A, Bae H. Ionomic Approaches for Discovery of Novel Stress-Resilient Genes in Plants. International Journal of Molecular Sciences. 2021; 22(13):7182. https://doi.org/10.3390/ijms22137182
Chicago/Turabian StyleAli, Sajad, Anshika Tyagi, and Hanhong Bae. 2021. "Ionomic Approaches for Discovery of Novel Stress-Resilient Genes in Plants" International Journal of Molecular Sciences 22, no. 13: 7182. https://doi.org/10.3390/ijms22137182
APA StyleAli, S., Tyagi, A., & Bae, H. (2021). Ionomic Approaches for Discovery of Novel Stress-Resilient Genes in Plants. International Journal of Molecular Sciences, 22(13), 7182. https://doi.org/10.3390/ijms22137182