
Bactericidal Activity of a Self-Biodegradable Lysine-Containing Dendrimer against Clinical Isolates of Acinetobacter Genus

 ,
, 
 and
and
Abstract
:
1. Introduction
2. Results and Discussion
2.1. Uncharged Polyester-Based Dendrimer Inner Scaffold (G5-PD-OH)
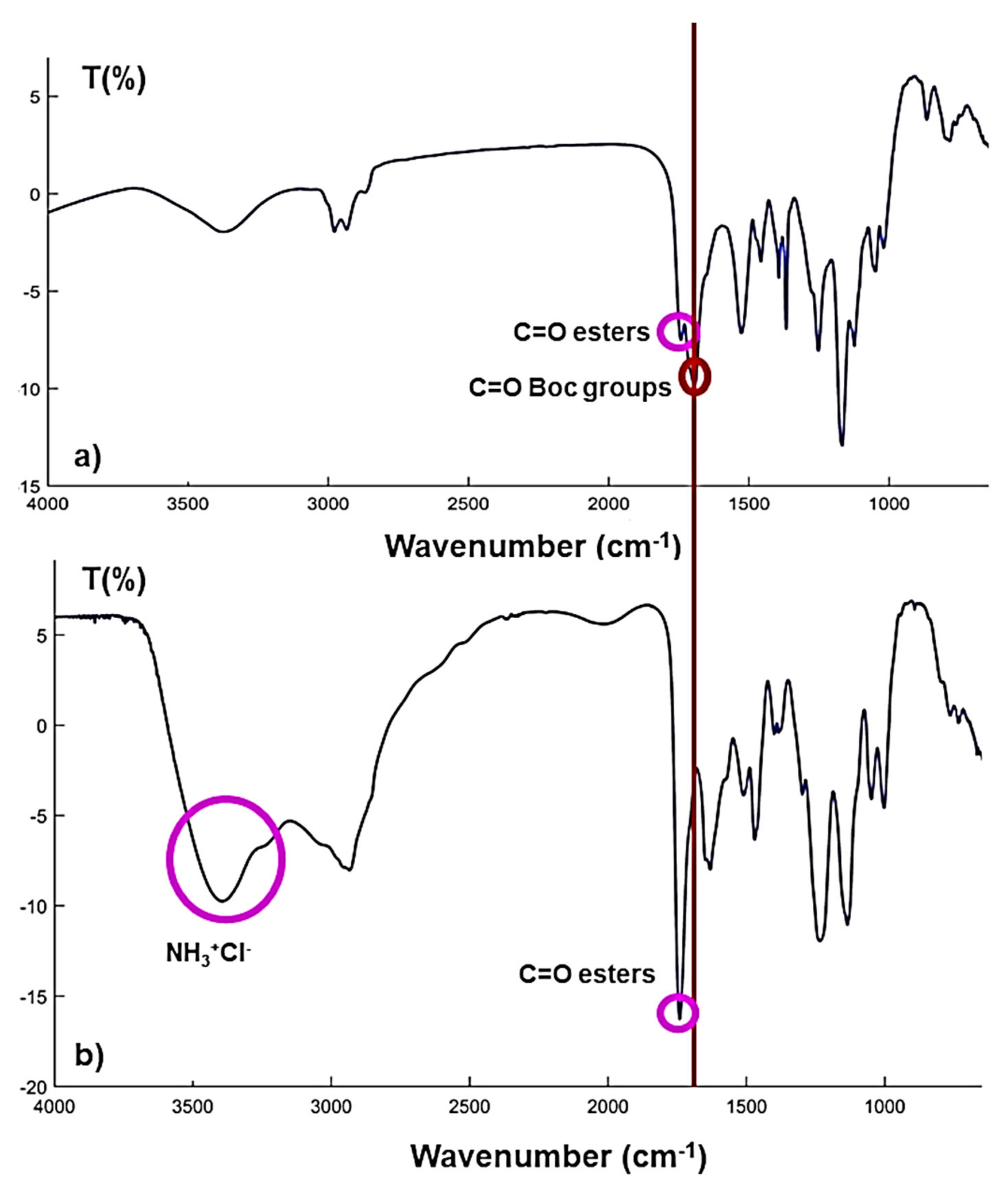
2.2. Synthesis and Spectroscopic Characterization of the Polyester-Based Cationic Dendrimer (G5-PDK * 128 HCl)
2.2.1. Synthesis and Spectroscopic Characterization of G5-PD-BK
2.2.2. Synthesis and Spectroscopic Characterization of G5-PDK * 128 HCl
2.3. MW Determination by Volumetric Titrations
2.4. Potentiometric Titration of G5-PDK
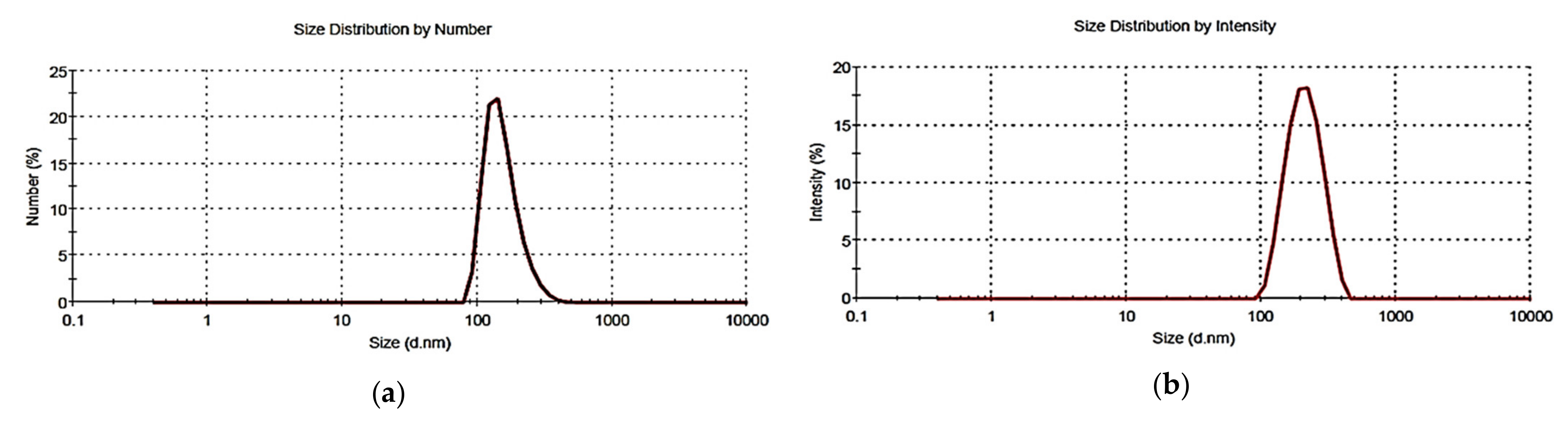
2.5. Dynamic Light Scattering Analysis
2.6. Antibacterial Activity of G5-PDK
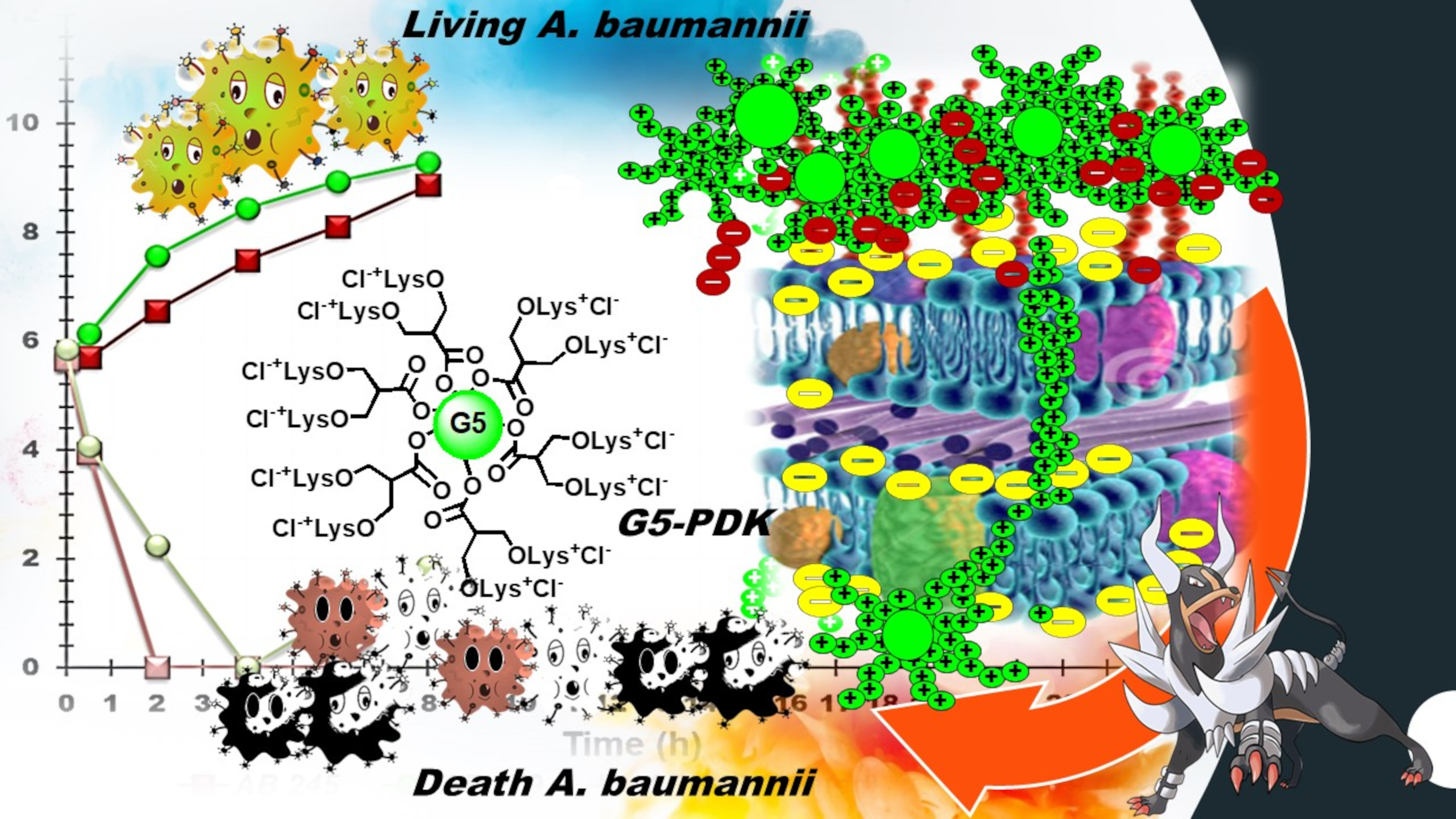
2.6.1. Design of the Structure of G5-PDK
2.6.2. MIC Values Displayed by G5-PDK
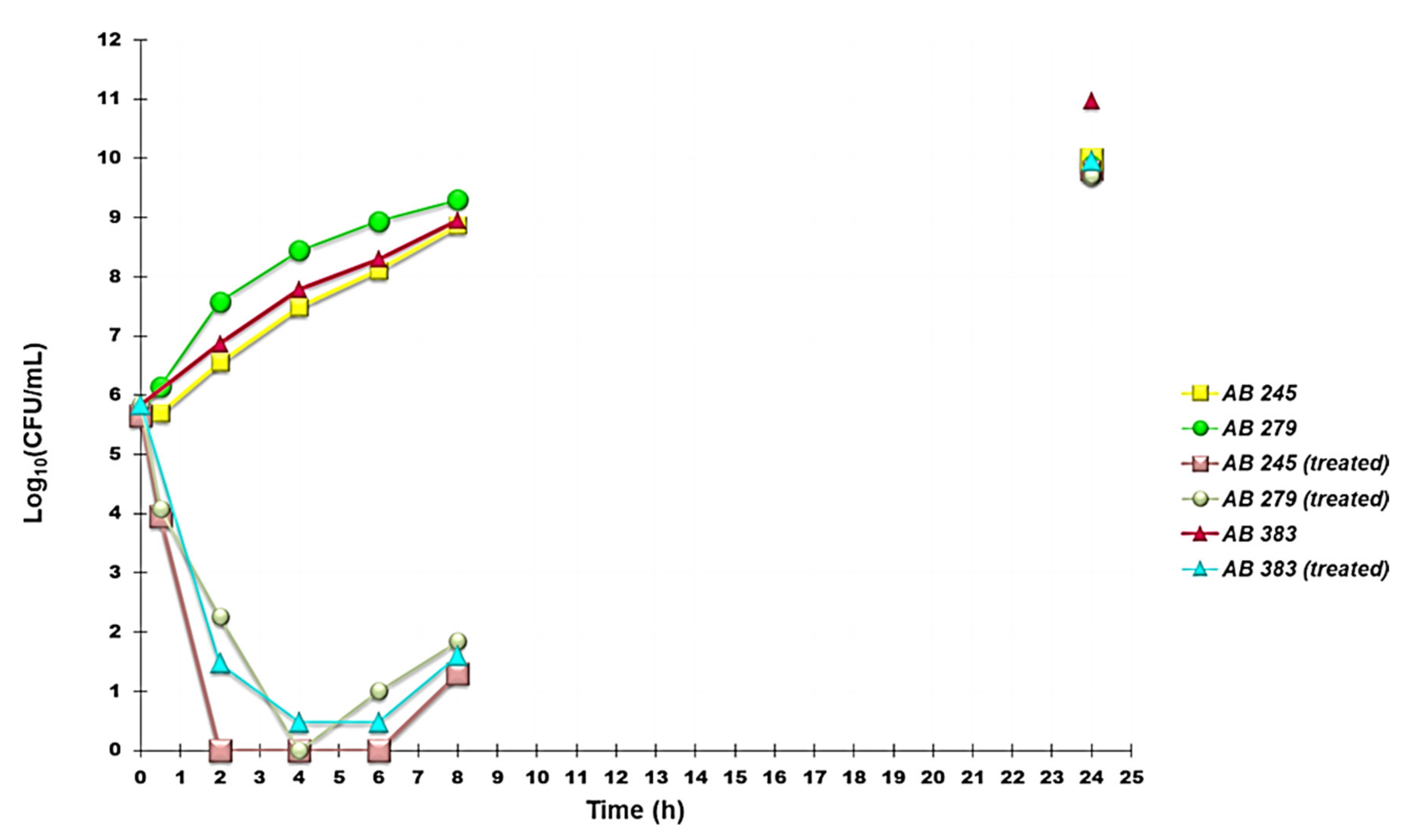
2.6.3. Time-Kill Curves
2.6.4. Authors Consideration about the Antibacterial Activity of G5-PDK
3. Materials and Methods
3.1. Chemicals and Instruments
3.2. FTIR, NMR Spectral Data, and Elemental Analysis Results of G5-PD-OH
3.3. Synthesis of Lysine-Modified Cationic Dendrimer G5-PDK (128 HCl)
3.3.1. Synthesis of Lysine-Modified Boc-Protected Dendrimer G5-PD-BK
3.3.2. Acidic Deprotection of G5-PD-BK to Obtain G5-PDK * 128 HCl
3.4. Determination of Experimental MW of G5-PDK by Volumetric Titration
Preparation of the Standard Solution of HClO4 in AcOH
3.5. Potentiometric Titration of G5-PDK
3.6. Dynamic Light Scattering (DLS) Analysis
3.7. Microbiology
3.7.1. Microorganisms
3.7.2. Determination of the MIC
3.7.3. Killing Curves
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Martín-Aspas, A.; Guerrero-Sánchez, F.M.; García-Colchero, F.; Rodríguez-Roca, S.; Girón-González, J.-A. Differential characteristics of Acinetobacter baumannii colonization and infection: Risk factors, clinical picture, and mortality. Infect. Drug Resist. 2018, 11, 861–872. [Google Scholar] [CrossRef] [Green Version]
- Roca, I.; Espinal, P.; Vila-Farrés, X.; Vila, J. The Acinetobacter baumannii oxymoron: Commensal hospital dweller turned pan-drug-resistant menace. Front. Microbiol. 2012, 3, 148. [Google Scholar] [CrossRef] [Green Version]
- Shlaes, D.M.; Bradford, P.A. Antibiotics—From There to Where? Pathog. Immun. 2018, 3, 19–43. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.-L.; Lee, Y.-T.; Kuo, S.-C.; Yang, S.-P.; Fung, C.-P.; Lee, S.-D. Rapid identification of Acinetobacter baumannii, Acinetobacter nosocomialis and Acinetobacter pittii with a multiplex PCR assay. J. Med. Microbiol. 2014, 63, 1154–1159. [Google Scholar] [CrossRef] [PubMed]
- Linde, H.-J.; Hahn, J.; Holler, E.; Reischl, U.; Lehn, N. Septicemia Due to Acinetobacter junii. J. Clin. Microbiol. 2002, 40, 2696–2697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cayô, R.; Segundo, L.Y.; del Molino Bernal, I.C.P.; de la Fuente, C.G.; Rodríguez, M.A.B.; Calvo, J.; Martínez, L.M. Bloodstream infection caused by Acinetobacter junii in a patient with acute lymphoblastic leukaemia after allogenic haematopoietic cell transplantation. J. Med. Microbiol. 2011, 60, 375–377. [Google Scholar] [CrossRef]
- Henao-Martínez, A.; González-Fontal, G.R.; Johnson, S. Celulitis por Acinetobacter junii-johnsonii adquirida en la comunidad: Una presentación de caso. Biomédica 2012, 32, 179–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonomo, R.A.; Szabo, D. Mechanisms of Multidrug Resistance in Acinetobacter Species and Pseudomonas aeruginosa. Clin. Infect. Dis. 2006, 43, S49–S56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Powers, M.J.; Trent, M.S. Expanding the paradigm for the outer membrane: Acinetobacter baumannii in the absence of endotoxin. Mol. Microbiol. 2018, 107, 47–56. [Google Scholar] [CrossRef] [Green Version]
- Harding, C.M.; Hennon, S.W.; Feldman, M.F. Uncovering the mechanisms of Acinetobacter baumannii virulence. Nat. Rev. Genet. 2018, 16, 91–102. [Google Scholar] [CrossRef]
- Dexter, C.; Murray, G.; Paulsen, I.; Peleg, A. Community-acquiredAcinetobacter baumannii: Clinical characteristics, epidemiology and pathogenesis. Expert Rev. Anti-Infect. Ther. 2015, 13, 567–573. [Google Scholar] [CrossRef]
- Garmendia, J.L.G.; Ortiz‐Leyba, C.; Garnacho-Montero, J.; Jiménez‐Jiménez, F.; Pérez‐Paredes, C.; Barrero‐Almodóvar, A.E.; Gili‐Miner, M. Risk Factors for Acinetobacter baumannii Nosocomial Bacteremia in Critically Ill Patients: A Cohort Study. Clin. Infect. Dis. 2001, 33, 939–946. [Google Scholar] [CrossRef] [Green Version]
- Robenshtok, E.; Paul, M.; Leibovici, L.; Fraser, A.; Pitlik, S.; Ostfeld, I.; Samra, Z.; Perez, S.; Lev, B.; Weinberger, M. The significance of Acinetobacter baumannii bacteraemia compared with Klebsiella pneumoniae bacteraemia: Risk factors and outcomes. J. Hosp. Infect. 2006, 64, 282–287. [Google Scholar] [CrossRef]
- Karageorgopoulos, D.E.; Falagas, M.E. Current control and treatment of multidrug-resistant Acinetobacter baumannii infections. Lancet Infect. Dis. 2008, 8, 751–762. [Google Scholar] [CrossRef]
- Wong, D.; Nielsen, T.B.; Bonomo, R.A.; Pantapalangkoor, P.; Luna, B.; Spellberg, B. Clinical and Pathophysiological Overview of Acinetobacter Infections: A Century of Challenges. Clin. Microbiol. Rev. 2017, 30, 409–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maragakis, L.L.; Perl, T.M. Antimicrobial Resistance:Acinetobacter baumannii:Epidemiology, Antimicrobial Resistance, and Treatment Options. Clin. Infect. Dis. 2008, 46, 1254–1263. [Google Scholar] [CrossRef] [Green Version]
- Falagas, M.E.; Karveli, E.A.; Kelesidis, T. Community-acquired Acinetobacter infections. Eur. J. Clin. Microbiol. Infect. Dis. 2007, 26, 857–868. [Google Scholar] [CrossRef] [PubMed]
- Anstey, N.M.; Currie, B.J.; Withnall, K.M. Community-Acquired Acinetobacter Pneumonia in the Northern Territory of Australia. Clin. Infect. Dis. 1992, 14, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Patamatamkul, S.; Klungboonkrong, V.; Praisarnti, P.; Jirakiat, K. A case-control study of community-acquired Acinetobacter baumannii pneumonia and melioidosis pneumonia in northeast Thailand: An emerging fatal disease with unique clinical features. Diagn. Microbiol. Infect. Dis. 2017, 87, 79–86. [Google Scholar] [CrossRef]
- Yang, Y.-S.; Lee, Y.-T.; Huang, T.-W.; Sun, J.-R.; Kuo, S.-C.; Yang, C.-H.; Chen, T.-L.; Lin, J.-C.; Fung, C.-P.; Chang, F.-Y. Acinetobacter baumannii nosocomial pneumonia: Is the outcome more favorable in non-ventilated than ventilated patients? BMC Infect. Dis. 2013, 13, 142. [Google Scholar] [CrossRef] [Green Version]
- Joly-Guillou, M.-L. Clinical impact and pathogenicity of Acinetobacter. Clin. Microbiol. Infect. 2005, 11, 868–873. [Google Scholar] [CrossRef] [Green Version]
- Fan, B.; Guan, J.; Wang, X.; Cong, Y. Activity of Colistin in Combination with Meropenem, Tigecycline, Fosfomycin, Fusidic Acid, Rifampin or Sulbactam against Extensively Drug-Resistant Acinetobacter baumannii in a Murine Thigh-Infection Model. PLoS ONE 2016, 11, e0157757. [Google Scholar] [CrossRef]
- Kim, W.-Y.; Moon, J.-Y.; Huh, J.W.; Choi, S.-H.; Lim, C.-M.; Koh, Y.; Chong, Y.P.; Hong, S.-B. Comparable Efficacy of Tigecycline versus Colistin Therapy for Multidrug-Resistant and Extensively Drug-Resistant Acinetobacter baumannii Pneumonia in Critically Ill Patients. PLoS ONE 2016, 11, e0150642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, G.G.; Ly, N.S.; Diep, J.; Forrest, A.; Bulitta, J.B.; Holden, P.N.; Nation, R.; Li, J.; Tsuji, B.T. Combinatorial pharmacodynamics of polymyxin B and tigecycline against heteroresistant Acinetobacter baumannii. Int. J. Antimicrob. Agents 2016, 48, 331–336. [Google Scholar] [CrossRef] [Green Version]
- Alfei, S.; Schito, A.M. Positively Charged Polymers as Promising Devices against Multidrug Resistant Gram-Negative Bacteria: A Review. Polymers 2020, 12, 1195. [Google Scholar] [CrossRef] [PubMed]
- Alfei, S.; Schito, A.M. From Nanobiotechnology, Positively Charged Biomimetic Dendrimers as Novel An-tibacterial Agents: A Review. Nanomaterials 2020, 10, 2022. [Google Scholar] [CrossRef] [PubMed]
- Schito, A.; Schito, G.; Alfei, S. Synthesis and Antibacterial Activity of Cationic Amino Acid-Conjugated Dendrimers Loaded with a Mixture of Two Triterpenoid Acids. Polymers 2021, 13, 521. [Google Scholar] [CrossRef] [PubMed]
- Schito, A.M.; Alfei, S. Antibacterial Activity of Non-Cytotoxic, Amino Acid-Modified Polycationic Dendrimers against Pseudomonas aeruginosa and Other Non-Fermenting Gram-Negative Bacteria. Polymers 2020, 12, 1818. [Google Scholar] [CrossRef]
- Alfei, S.; Piatti, G.; Caviglia, D.; Schito, A. Synthesis, Characterization, and Bactericidal Activity of a 4-Ammoniumbuthylstyrene-Based Random Copolymer. Polymers 2021, 13, 1140. [Google Scholar] [CrossRef]
- Alfei, S.; Marengo, B.; Valenti, G.; Domenicotti, C. Synthesis of Polystyrene-Based Cationic Nanomaterials with Pro-Oxidant Cytotoxic Activity on Etoposide-Resistant Neuroblastoma Cells. Nanomaterials 2021, 11, 977. [Google Scholar] [CrossRef]
- Schito, A.A.; Piatti, G.; Caviglia, D.; Zuccari, G.; Alfei, S. Broad-Spectrum non-lytic Bactericidal Activity of a Synthetic Random Copolymer based on 2-Methoxy-6-(4-vinylbenzyloxy)-benzyl-ammonium chloride. Int. J. Mol. Sci. 2021, 22, 5021. [Google Scholar] [CrossRef] [PubMed]
- Alfei, S.; Catena, S.; Turrini, F. Biodegradable and biocompatible spherical dendrimer nanoparticles with a gallic acid shell and a double-acting strong antioxidant activity as potential device to fight diseases from “oxidative stress”. Drug Deliv. Transl. Res. 2019, 10, 259–270. [Google Scholar] [CrossRef]
- Ihre, H.; Hult, A.; Fréchet, J.M.J.; Gitsov, I. Double-stage convergent approach for the synthesis of function-alized dendritic aliphatic polyesters based on 2,2-bis(hydroxymethyl)propionic acid. Macromolecules 1998, 31, 4061–4068. [Google Scholar] [CrossRef]
- Alfei, S.; Marengo, B.; Domenicotti, C. Polyester-Based Dendrimer Nanoparticles Combined with Etoposide Have an Improved Cytotoxic and Pro-Oxidant Effect on Human Neuroblastoma Cells. Antioxidants 2020, 9, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alfei, S.; Castellaro, S.; Taptue, G.B. Synthesis and NMR characterization of dendrimers based on 2, 2-bis-(hydroxymethyl)-propanoic acid (bis-HMPA) containing peripheral amino acid residues for gene transfection. Org. Commun. 2017, 10, 144–177. [Google Scholar] [CrossRef]
- Alfei, S.; Castellaro, S. Synthesis and characterization of polyester-based dendrimers containing peripheral arginine or mixed amino acids as potential vectors for gene and drug delivery. Macromol. Res. 2017, 25, 1172–1186. [Google Scholar] [CrossRef]
- Vogel, A.I. Elementary Practical Organic Chemistry, Part III; Quantitative Organic Analysis; Longman: London, UK, 1958; Chapter 20; pp. 702–705. [Google Scholar]
- Benns, J.M.; Choi, J.S.; Mahato, R.I.; Park, J.S.; Kim, S.W. pH-sensitive cationic polymer gene delivery ve-hicle: N-Ac-poly(L-histidine)-graft-poly(L-lysine) comb shaped polymer. Bioconj. Chem. 2000, 11, 637–645. [Google Scholar] [CrossRef] [PubMed]
- Microbe Note. Available online: https://microbenotes.com/oxidation-fermentation-of-test (accessed on 3 July 2021).
- Twyman, L.J.; Beezer, A.E.; Esfand, R.; Hardy, M.J.; Mitchell, J. The synthesis of water soluble dendrimers, and their application as possible drug delivery systems. Tetrahedron Lett. 1999, 40, 1743–1746. [Google Scholar] [CrossRef]
- Wang, J.; Mao, W.; Lock, L.L.; Tang, J.; Sui, M.; Sun, W.; Cui, H.; Xu, D.; Shen, Y. The Role of Micelle Size in Tumor Accumulation, Penetration, and Treatment. ACS Nano 2015, 9, 7195–7206. [Google Scholar] [CrossRef]
- Yu, H.; Cui, Z.; Yu, P.; Guo, C.; Feng, B.; Jiang, T.; Wang, S.; Yin, Q.; Zhong, D.; Yang, X.; et al. pH- and NIR Light-Responsive Micelles with Hyperthermia-Triggered Tumor Penetration and Cytoplasm Drug Release to Reverse Doxorubicin Resistance in Breast Cancer. Adv. Funct. Mater. 2015, 25, 2489–2500. [Google Scholar] [CrossRef]
- Ankamwar, B. Size and Shape Effect on Biomedical Applications of Nanomaterials. In Biomedical Engineering-Technical Applications in Medicine; Hudak, R., Penhaker, M., Majernik, J., Eds.; IntechOpen: London, UK, 2012; p. 2439. [Google Scholar]
- Stenström, P.; Hjorth, E.; Zhang, Y.; Andrén, O.C.J.; Guette-Marquet, S.; Schultzberg, M.; Malkoch, M. Syn-thesis and in vitro evaluation of monodisperse amino-functional polyester dendrimers with rapid degra-dability and antibacterial properties. Biomacromolecules 2017, 18, 4323–4330. [Google Scholar] [CrossRef]
- Giacometti, A.; Cirioni, O.; Del Prete, M.S.; Barchiesi, F.; Paggi, A.M.; Petrelli, E.; Scalise, G. Comparative activities of polycationic peptides and clinically used antimicrobial agents against multidrug-resistant nosocomial isolates of Acinetobacter baumannii. J. Antimicrob. Chemother. 2000, 46, 807–810. [Google Scholar] [CrossRef]
- Vila-Farres, X.; de la Maria, C.G.; López-Rojas, R.; Pachón, J.; Giralt, E.; Vila, J. In vitro activity of several antimicrobial peptides against colistin-susceptible and colistin-resistant Acinetobacter baumannii. Clin. Microbiol. Infect. 2012, 18, 383–387. [Google Scholar] [CrossRef] [Green Version]
- Jaśkiewicz, M.; Neubauer, D.; Kazor, K.; Bartoszewska, S.; Kamysz, W. Antimicrobial Activity of Selected Antimicrobial Peptides Against Planktonic Culture and Biofilm of Acinetobacter baumannii. Probiotics Antimicrob. Proteins 2019, 11, 317–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGratha, D.M.; Barbua, E.M.; Driessena, W.H.P.; Lascob, T.M.; Tarrand, J.J.; Okhuysend, P.C.; Kontoyiannise, D.P.; Sidmanf, R.L.; Pasqualinia, P.; Arapa, W. Mechanism of action and initial evaluation of a membrane active all-D-enantiomer antimicrobial peptidomimetic. Proc. Natl. Acad. Sci. USA 2013, 110, 3477–3482. [Google Scholar] [CrossRef] [Green Version]
- Pires, J.; Siriwardena, T.N.; Stach, M.; Tinguely, R.; Kasraian, S.; Luzzaro, F.; Leib, S.L.; Darbre, T.; Reymond, J.-L.; Endimiani, A. In VitroActivity of the Novel Antimicrobial Peptide Dendrimer G3KL against Multidrug-Resistant Acinetobacter baumannii and Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2015, 59, 7915–7918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, D. Cationic antimicrobial peptide and its poly-N-substituted glycine congener: Antibacterial and antibiofilm potential against A. baumannii. Biochem. Biophys. Res. Commun. 2019, 518, e478475. [Google Scholar] [CrossRef]
- Winn, W.; Allen, S.; Janda, W.; Koneman, E.; Procop, G.; Schreckenberger, P.; Woods, G. Color Atlas and Textbook of Diagnostic Microbiology, 6th ed.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2006. [Google Scholar]
- American Society for Microbiology. Available online: https://www.asmscience.org/content/education/protocol/protocol.3151 (accessed on 17 June 2021).
- Mizutani, M.; Palermo, E.F.; Thoma, L.; Satoh, K.; Kamigaito, M.; Kuroda, K. Design and Synthesis of Self-Degradable Antibacterial Polymers by Simultaneous Chain- and Step-Growth Radical Copolymerization. Biomacromolecules 2012, 13, 1554–1563. [Google Scholar] [CrossRef] [PubMed]
- Lim, Y.-B.; Kim, C.-H.; Kim, K.; Kim, A.S.W.; Park, J.-S. Development of a Safe Gene Delivery System Using Biodegradable Polymer, Poly[α-(4-aminobutyl)-l-glycolic acid]. J. Am. Chem. Soc. 2000, 122, 6524–6525. [Google Scholar] [CrossRef]
- Lim, Y.-B.; Han, S.-O.; Kong, H.-U.; Lee, Y.; Park, J.-S.; Jeong, B.; Kim, S.W. Biodegradable Polyester, Poly[α-(4-Aminobutyl)-l-Glycolic Acid], as a Non-Toxic Gene Carrier. Pharm. Res. 2000, 17, 811–816. [Google Scholar] [CrossRef]
- Marr, A.; Gooderham, W.J.; Hancock, R. Antibacterial peptides for therapeutic use: Obstacles and realistic outlook. Curr. Opin. Pharmacol. 2006, 6, 468–472. [Google Scholar] [CrossRef]
- Tao, Y.; Chen, X.; Jia, F.; Wang, S.; Xiao, C.; Cui, F.; Li, Y.; Bian, Z.; Chen, X.; Wang, X. New Chemosynthetic Route to Linear ε-Poly-Lysine. Chem. Sci. 2015, 6, 6385–6391. [Google Scholar] [CrossRef] [Green Version]
- Barman, S.; Konai, M.M.; Samaddar, S.; Haldar, J. Amino Acid Conjugated Polymers: Antibacterial Agents Effective against Drug-Resistant Acinetobacter baumannii with No Detectable Resistance. ACS Appl. Mater. Interfaces 2019, 11, 33559–33572. [Google Scholar] [CrossRef] [PubMed]
- Lou, W.; Venkataraman, S.; Zhong, G.; Ding, B.; Tan, J.P.; Xu, L.; Fan, W.; Yang, Y.Y. Antimicrobial polymers as therapeutics for treatment of multidrug-resistant Klebsiella pneumoniae lung infection. Acta Biomater. 2018, 78, 78–88. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.; Barman, S.; Mukherjee, R.; Haldar, J. Amphiphilic Cationic Macromolecules Highly Effective Against Multi-Drug Resistant Gram-Positive Bacteria and Fungi With No Detectable Resistance. Front. Bioeng. Biotechnol. 2020, 8, 55. [Google Scholar] [CrossRef]
- Landman, D.; Georgescu, C.; Martin, D.A.; Quale, J. Polymyxins revisited. Clin. Microbiol. Rev. 2008, 21, 449–465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boll, J.M.; Tucker, A.T.; Klein, D.R.; Beltran, A.M.; Brodbelt, J.S.; Davies, B.W.; Trent, M.S. Reinforcing lipid A acylation on the cell surface of Acinetobacter baumannii promotes cationic antimicrobial peptide re-sistance and desiccation survival. MBio 2015, 6, e00478-15. [Google Scholar] [CrossRef] [Green Version]
- Preston, A.; Mandrell, R.E.; Gibson, B.W.; Apicella, M.A. The Lipooligosaccharides of Pathogenic Gram-Negative Bacteria. Crit. Rev. Microbiol. 1996, 22, 139–180. [Google Scholar] [CrossRef] [PubMed]
- Vinogradov, E.V.; Duus, J.Ø.; Brade, H.; Holst, O. The structure of the carbohydrate backbone of the lipo-polysaccharide from Acinetobacter baumannii strain ATCC 19606. Eur. J. Biochem. 2002, 269, 422–430. [Google Scholar] [CrossRef]
- Nikaido, H. Molecular Basis of Bacterial Outer Membrane Permeability Revisited. Microbiol. Mol. Biol. Rev. 2003, 67, 593–656. [Google Scholar] [CrossRef] [Green Version]
- Alfei, S.; Oliveri, P.; Malegori, C. Assessment of the Efficiency of a Nanospherical Gallic Acid Dendrimer for Long‐Term Preservation of Essential Oils: An Integrated Chemometric‐Assisted FTIR Study. ChemistrySelect 2019, 4, 8891–8901. [Google Scholar] [CrossRef]
- Alfei, S.; Signorello, M.G.; Schito, A.; Catena, S.; Turrini, F. Reshaped as polyester-based nanoparticles, gallic acid inhibits platelet aggregation, reactive oxygen species production and multi-resistant Gram-positive bacteria with an efficiency never obtained. Nanoscale Adv. 2019, 1, 4148–4157. [Google Scholar] [CrossRef] [Green Version]
- EUCAST. European Committee on Antimicrobial Susceptibility Testing. Available online: https://www.eucast.org/ast_of_bacteria (accessed on 3 July 2021).
- Schito, A.M.; Piatti, G.; Stauder, M.; Bisio, A.; Giacomelli, E.; Romussi, G.; Pruzzo, C. Effects of deme-thylfruticuline A and fruticuline A from Salvia corrugata Vahl. on biofilm production in vitro by multire-sistant strains of Staphylococcus aureus, Staphylococcus epidermidis and Enterococcus faecalis. Int. J. Antimicrob. Agents 2011, 37, 129–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Physical Characteristics | G5-PDK | |
|---|---|---|
| N 1 | 128 | |
| MW (calc.) | 20,145.3 | |
| MW (obs.) | 19,961.2 ± 480.2 | |
| Error (%) | −0.9% | |
| Z-Ave (nm) | 203.0 ± 2.6 2 | 203.0 ± 2.6 3 |
| PDI | 0.282 ± 0.028 2 | 0.282 ± 0.028 3 |
| ζ-p (mV) | +19.2 ± 7.3 | |
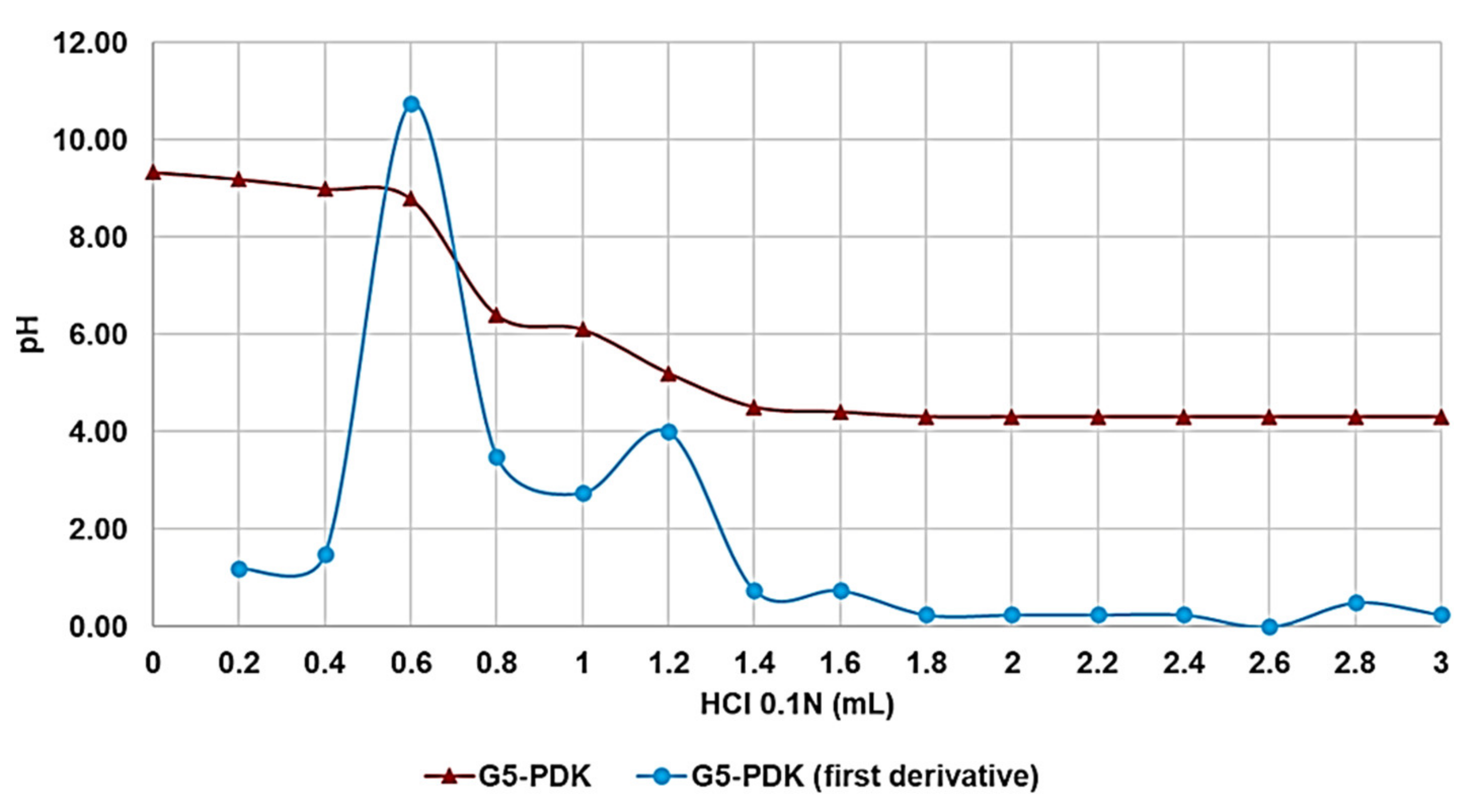
| Max dpH/dV | 10.75 | 4.0 |
| HCl (mL) | 0.6 | 1.2 |
| pH | 6.85 | 4.80 |
| mL HCl 0.1N | pH | dpH/dV |
|---|---|---|
| 0.0 | 9.54 ± 0.02 | ---* |
| 0.2 | 9.30 ± 0.03 | 1.2 |
| 0.4 | 9.00 ± 0.02 | 1.5 |
| 0.6 | 6.85 ± 0.05 | 10.75 |
| 0.8 | 6.15 ± 0.01 | 3.5 |
| 1.0 | 5.60 ± 0.04 | 2.75 |
| 1.2 | 4.80 ± 0.02 | 4 |
| 1.4 | 4.65 ± 0.02 | 0.75 |
| 1.6 | 4.50 ± 0.03 | 0.75 |
| 1.8 | 4.45 ± 0.02 | 0.25 |
| 2.0 | 4.40 ± 0.04 | 0.25 |
| 2.2 | 4.35 ± 0.02 | 0.25 |
| 2.4 | 4.30 ± 0.01 | 0.25 |
| 2.6 | 4.30 ± 0.009 | 0 |
| 2.8 | 4.20 ± 0.009 | 0.5 |
| 3.0 | 4.15 ± 0.01 | 0.25 |
| G5-PDK | ||
| Max dpH/dV | 10.75 | 4.0 |
| HCl (mL) | 0.6 | 1.2 |
| pH | 6.85 | 4.80 |
| G5-PDK (20145) 2 | Ciprofloxacin | |
|---|---|---|
| Strains | MIC µM (µg/mL) | MIC µM (µg/mL) |
| E. faecalis * | >25.4 (>512) | 193.2 (64) |
| E. faecium * | >25.4 (>512) | 772.7 (256) |
| S. aureus ** | >25.4 (>512) | 386.4 (128) |
| S. epidermidis ** | >25.4 (>512) | 193.2 (64) |
| E. coli # | >25.4 (>512) | 96.6 (32) |
| K. pneumoniae # | >25.4 (>512) | 96.6 (32) |
| A. baumannii | 6.3 (128) | 193.2 (64) |
| G5-PDK (20145) 2 | Ciprofloxacin | |
|---|---|---|
| Strains | MIC µM (µg/mL) | MIC µM (µg/mL) |
| A. baumannii 236 | 6.3 (128) | 193.2 (64) |
| A. baumannii 245 | 6.3 (128) | 1.6 (0.5) |
| A. baumannii 257 | 6.3 (128) | 96.6 (32) |
| A. baumannii 279 | 12.7 (256) | 48.3 (16) |
| A. baumannii 383 | 6.3 (128) | 193.2 (64) |
| A. baumannii 406 | 6.3 (128) | 96.6 (32) |
| A. johnsonii 387 | 6.3 (128) | 0.9 (0.3) |
| A. junii 389 | 12.7 (256) | 0.4 (0.125) |
| A. pittii 263 | 6.3 (128) | 3.2 (1) |
| A. pittii 272 | 6.3 (128) | 1.6 (0.5) |
| A. ursingii 388 | 3.2 (64) | 0.4 (0.125) |
| A. ursingii 408 | 6.3 (128) | 0.8 (0.25) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alfei, S.; Caviglia, D.; Piatti, G.; Zuccari, G.; Schito, A.M. Bactericidal Activity of a Self-Biodegradable Lysine-Containing Dendrimer against Clinical Isolates of Acinetobacter Genus. Int. J. Mol. Sci. 2021, 22, 7274. https://doi.org/10.3390/ijms22147274
Alfei S, Caviglia D, Piatti G, Zuccari G, Schito AM. Bactericidal Activity of a Self-Biodegradable Lysine-Containing Dendrimer against Clinical Isolates of Acinetobacter Genus. International Journal of Molecular Sciences. 2021; 22(14):7274. https://doi.org/10.3390/ijms22147274
Chicago/Turabian StyleAlfei, Silvana, Debora Caviglia, Gabriella Piatti, Guendalina Zuccari, and Anna Maria Schito. 2021. "Bactericidal Activity of a Self-Biodegradable Lysine-Containing Dendrimer against Clinical Isolates of Acinetobacter Genus" International Journal of Molecular Sciences 22, no. 14: 7274. https://doi.org/10.3390/ijms22147274
APA StyleAlfei, S., Caviglia, D., Piatti, G., Zuccari, G., & Schito, A. M. (2021). Bactericidal Activity of a Self-Biodegradable Lysine-Containing Dendrimer against Clinical Isolates of Acinetobacter Genus. International Journal of Molecular Sciences, 22(14), 7274. https://doi.org/10.3390/ijms22147274