
Influence of Cannabinoid Receptor 1 Genetic Variants on the Subjective Effects of Smoked Cannabis


 ,
,  ,
, 
Abstract
:1. Introduction
2. Results
2.1. Subject Pool and Genotyping Results
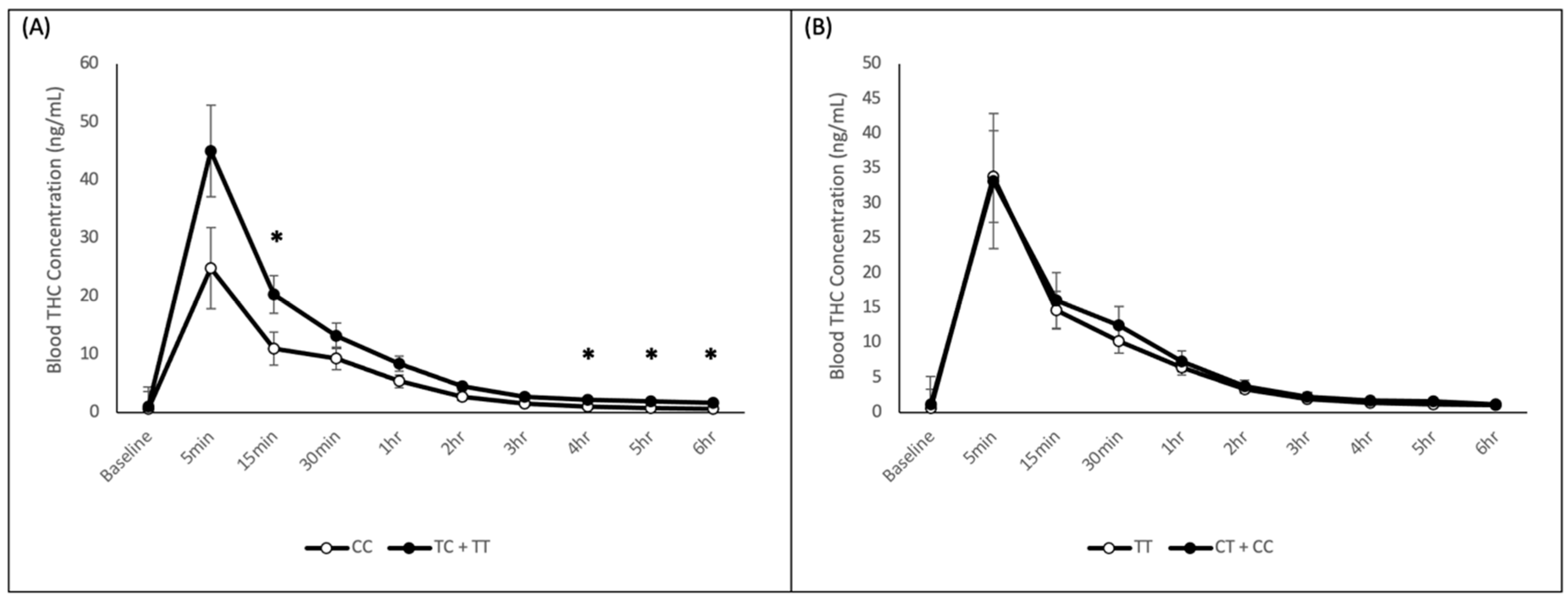
2.2. Blood THC Analysis
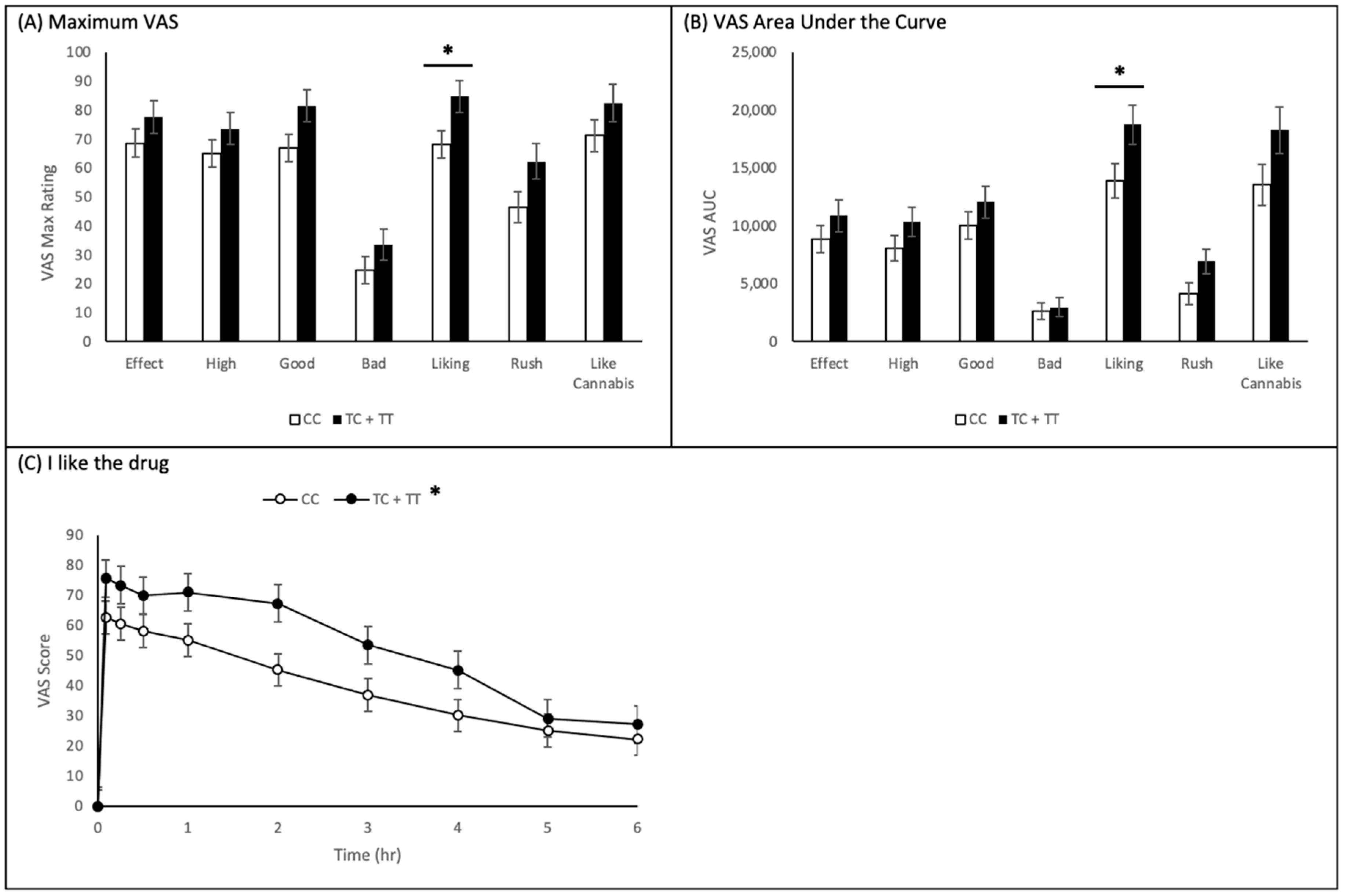
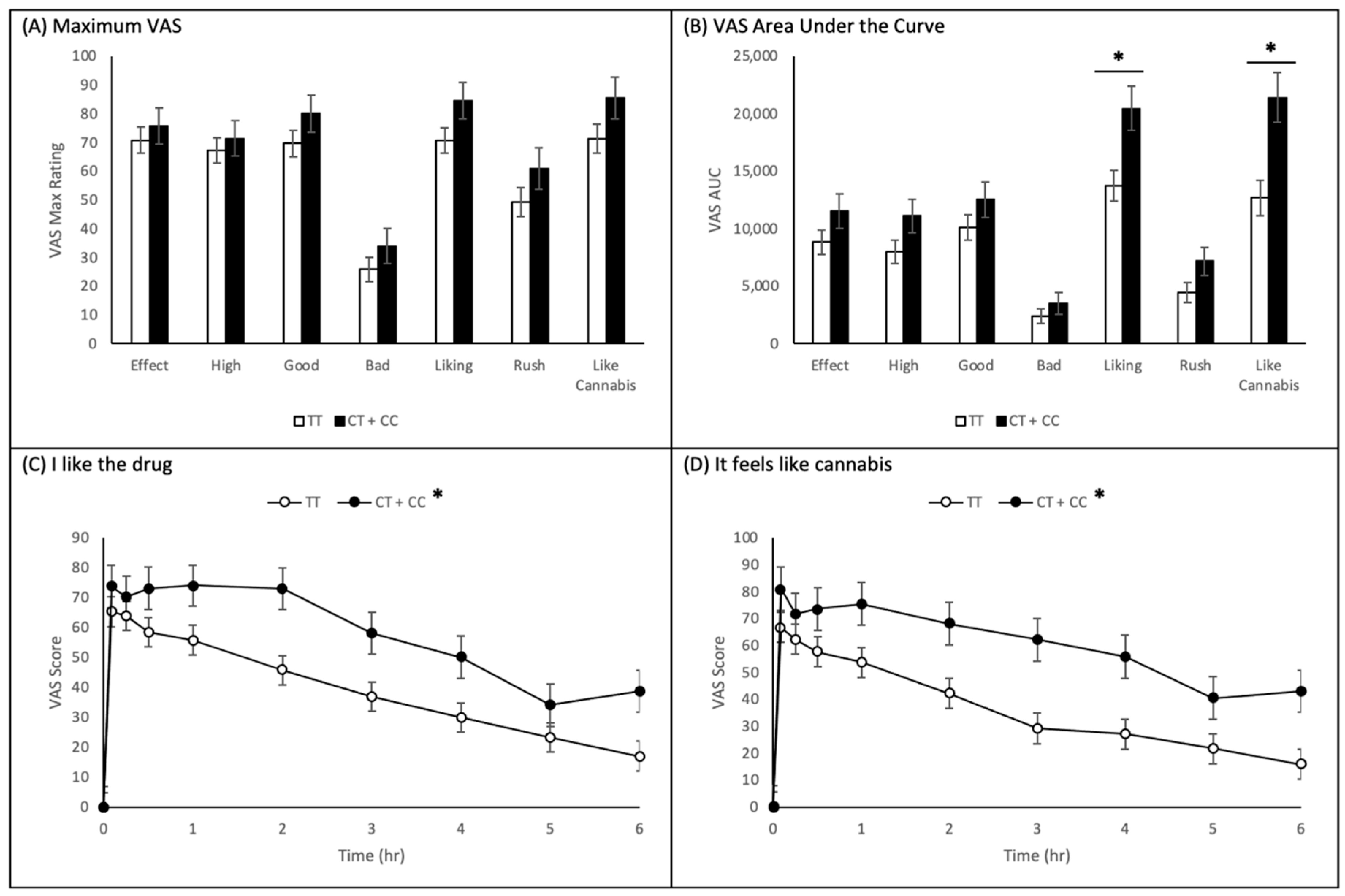
2.3. Visual Analogue Scales
2.3.1. CNR1 rs1049353 Polymorphism
2.3.2. CNR1 rs2023239 Polymorphism
2.3.3. Adjusted Results
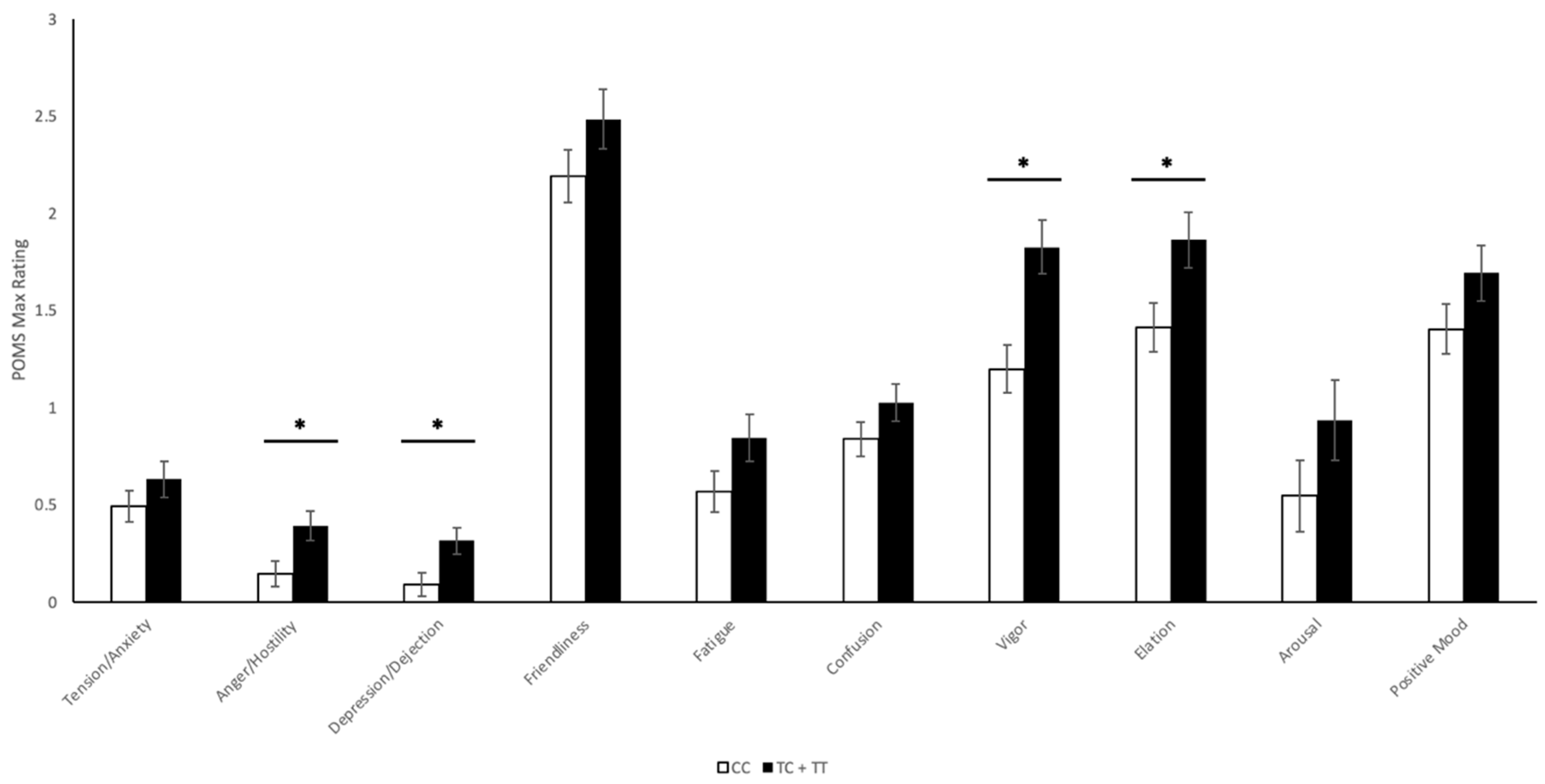
2.4. Profile of Mood States
2.4.1. CNR1 rs1049353 Polymorphism
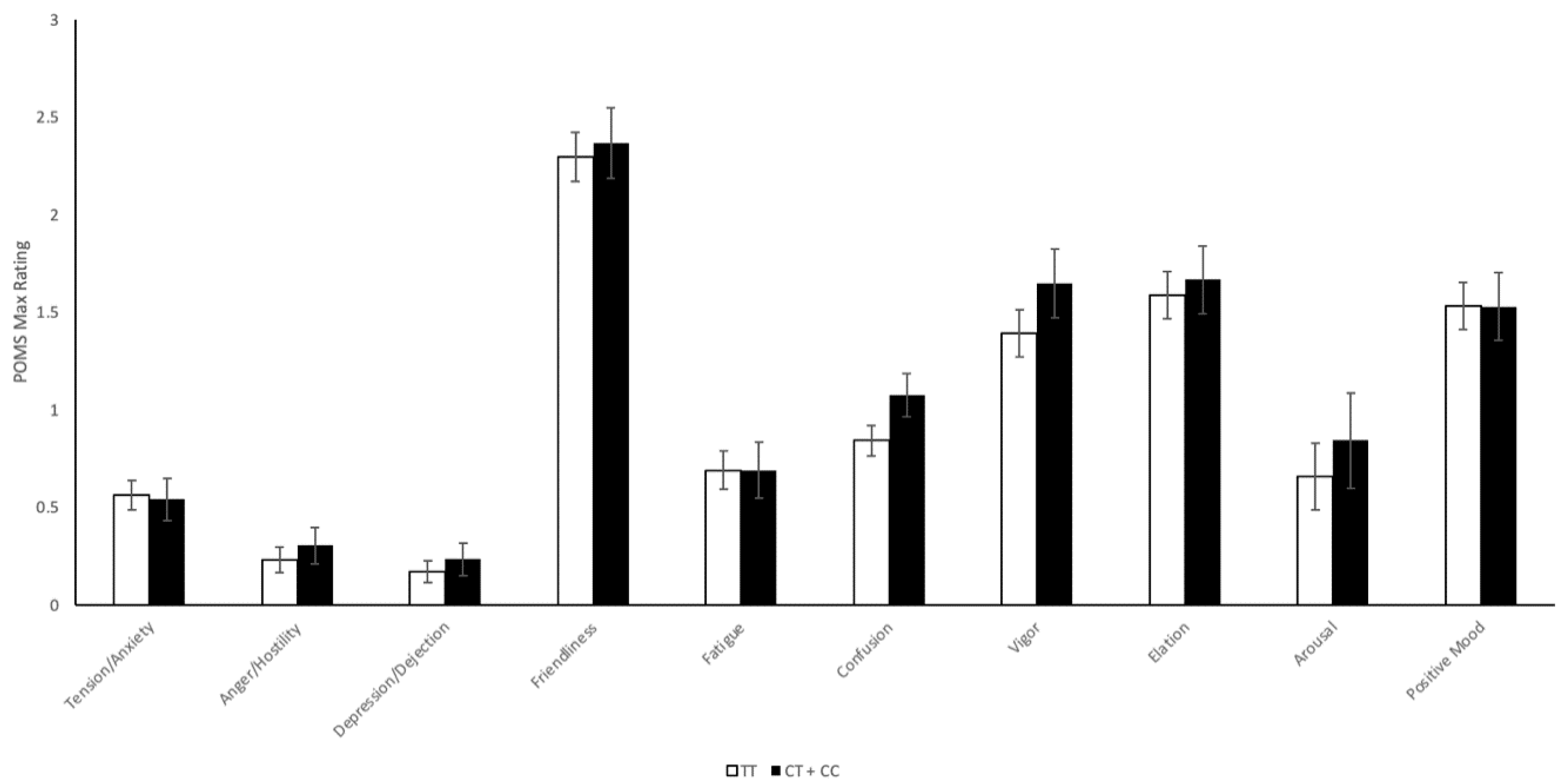
2.4.2. CNR1 rs2023239 Polymorphism
2.4.3. Adjusted Results
3. Materials and Methods
3.1. CADRI Study
- Self-reports of drug effects using Visual Analog Scales (VAS)
- ○
- VAS questions: (1) I feel a drug effect, (2) I feel this high, (3) I feel the drug’s good effects, (4) I feel the drug’s bad effects, (5) I like the drug, (6) I feel a rush, and (7) It feels like cannabis.
- ○
- Assessed on a 10 cm continuum and scored from 0 to 100.
- ○
- Baseline, 5, 15, 30 min and 1, 2, 3, 4, 5, 6, 24, and 48 h post-dose.
- ○
- Psychometric rating scales used to assess a variety of transient mood states.
- ○
- POMS scales: (1) Tension/Anxiety, (2) Anger/Hostility, (3) Depression/Dejection, (4) Friendliness, (5) Fatigue, (6) Confusion, (7) Vigor, (8) Elation, (9) Arousal, and (10) Positive Mood.
- ○
- Baseline, 1, 24, and 48 h post-dose.
3.2. Genotyping Experiment
3.3. Statistical Analysis
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Russo, E.B.; Jiang, H.E.; Li, X.; Sutton, A.; Carboni, A.; del Bianco, F.; Mandolino, G.; Potter, D.J.; Zhao, Y.X.; Bera, S.; et al. Phytochemical and genetic analyses of ancient cannabis from Central Asia. J. Exp. Bot. 2008, 59, 4171–4182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Connor, J.P.; Stjepanovic, D.; Le Foll, B.; Hoch, E.; Budney, A.J.; Hall, W.D. Cannabis use and cannabis use disorder. Nat. Rev. Dis. Primers 2021, 7, 16. [Google Scholar] [CrossRef] [PubMed]
- Adams, I.B.; Martin, B.R. Cannabis: Pharmacology and toxicology in animals and humans. Addiction 1996, 91, 1585–1614. [Google Scholar] [CrossRef] [PubMed]
- Johnston, L.D.; O’Malley, P.M.; Bachman, J.G. National Survey Results on Drug Use from the Monitoring the Future Study; National Institute on Drug Abuse, US Department of Health and Human Services, Public Health Service, National Institutes of Health: Washington, DC, USA, 1995; pp. 1975–1997.
- Adamson, P. Child Well-being in Rich Countries: A comparative overview. UNICEF Off. Res. 2013. Available online: https://time.com/wp-content/uploads/2015/04/rc11_eng.pdf (accessed on 5 July 2021).
- Health Canada. Summary of results: Canadian Student Tobacco, Alcohol and Drugs Survey 2014–2015. 2016. Available online: https://www.canada.ca/en/health-canada/services/canadian-student-tobacco-alcohol-drugs-survey/2014-2015-summary.html (accessed on 5 July 2021).
- Statistics Canada. Analysis of trends in the prevalence of cannabis use in Canada, 1985 to 2015. Health Rep. 2018, 29, 10–20. [Google Scholar]
- Azofeifa, A.; Mattson, M.E.; Schauer, G.; McAfee, T.; Grant, A.; Lyerla, R. National Estimates of Marijuana Use and Related Indicators—National Survey on Drug Use and Health, United States, 2002–2014. Morb. Mortal. Wkly. Rep. 2016, 65, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Rotermann, M.; Langlois, K. Prevalence and Correlates of Marijuana Use in Canada, 2012. Health Rep. 2015, 26, 10–15. [Google Scholar] [PubMed]
- United Nations Office on Drugs and Crime. World Drug Report 2019. 2019. Available online: https://onlinelibrary.wiley.com/doi/abs/10.1002/9781118929803.ewac0543 (accessed on 5 July 2021).
- Statistics Canada. National Cannabis Survey, first quarter 2019. 2019. Available online: https://www150.statcan.gc.ca/n1/pub/11-627-m/11-627-m2019032-eng.htm (accessed on 5 July 2021).
- Rotermann, M. Analysis of trends in the prevalence of cannabis use and related metrics in Canada. Health Rep. 2019, 30, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Russell, C.; Rueda, S.; Room, R.; Tyndall, M.; Fischer, B. Routes of administration for cannabis use-basic prevalence and related health outcomes: A scoping review and synthesis. Int. J. Drug Policy 2018, 52, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Health Canada. Canadian Tobacco, Alcohol and Drugs Survey (CTADS): Summary of Results for 2017. 2019. Available online: https://www.canada.ca/en/health-canada/services/canadian-tobacco-alcohol-drugs-survey/2017-summary.html (accessed on 5 July 2021).
- Atakan, Z. Cannabis, a complex plant: Different compounds and different effects on individuals. Adv. Psychopharmacol 2012, 2, 241–254. [Google Scholar] [CrossRef] [Green Version]
- Di Marzo, V.; De Petrocellis, L.; Bisogno, T. Endocannabinoids Part I: Molecular basis of endocannabinoid formation, action and inactivation and development of selective inhibitors. Expert. Opin. Targets 2001, 5, 241–265. [Google Scholar] [CrossRef]
- Sugiura, T.; Waku, K. Cannabinoid receptors and their endogenous ligands. J. Biochem. 2002, 132, 7–12. [Google Scholar] [CrossRef]
- Gonzalez-Mariscal, I.; Krzysik-Walker, S.M.; Doyle, M.E.; Liu, Q.R.; Cimbro, R.; Santa-Cruz Calvo, S.; Ghosh, S.; Ciesla, L.; Moaddel, R.; Carlson, O.D.; et al. Human CB1 Receptor Isoforms, present in Hepatocytes and beta-cells, are Involved in Regulating Metabolism. Sci. Rep. 2016, 6, 33302. [Google Scholar] [CrossRef] [Green Version]
- Pertwee, R.G. Cannabinoid pharmacology: The first 66 years. Br. J. Pharm. 2006, 147 (Suppl. 1), S163–S171. [Google Scholar] [CrossRef] [Green Version]
- Pertwee, R.G. Pharmacology of cannabinoid CB1 and CB2 receptors. Pharmacology 1997, 74, 129–180. [Google Scholar] [CrossRef]
- Ashton, C.H. Pharmacology and effects of cannabis: A brief review. Br. J. Psychiatry 2001, 178, 101–106. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. The health and Social Effects of Nonmedical Cannabis Use. 2016. Available online: https://www.who.int/substance_abuse/publications/msbcannabis.pdf (accessed on 5 July 2021).
- Hill, S.Y.; Sharma, V.; Jones, B.L. Lifetime use of cannabis from longitudinal assessments, cannabinoid receptor (CNR1) variation, and reduced volume of the right anterior cingulate. Psychiatry Res. Neuroimaging 2016, 255, 24–34. [Google Scholar] [CrossRef] [Green Version]
- Hindocha, C.; Freeman, T.P.; Schafer, G.; Gardner, C.; Bloomfield, M.A.P.; Bramon, E.; Morgan, C.J.A.; Curran, H.V. Acute effects of cannabinoids on addiction endophenotypes are moderated by genes encoding the CB1 receptor and FAAH enzyme. Addict. Biol. 2020, 25, e12762. [Google Scholar] [CrossRef]
- Gerra, M.C.; Jayanthi, S.; Manfredini, M.; Walther, D.; Schroeder, J.; Phillips, K.A.; Cadet, J.L.; Donnini, C. Gene variants and educational attainment in cannabis use: Mediating role of DNA methylation. Transl. Psychiatry 2018, 8, 23. [Google Scholar] [CrossRef]
- Palmer, R.H.C.; McGeary, J.E.; Knopik, V.S.; Bidwell, L.C.; Metrik, J.M. CNR1 and FAAH variation and affective states induced by marijuana smoking. Am. J. Drug Alcohol Abus. 2019, 45, 514–526. [Google Scholar] [CrossRef]
- Hartman, C.A.; Hopfer, C.J.; Haberstick, B.; Rhee, S.H.; Crowley, T.J.; Corley, R.P.; Hewitt, J.K.; Ehringer, M.A. The association between cannabinoid receptor 1 gene (CNR1) and cannabis dependence symptoms in adolescents and young adults. Drug Alcohol Depend. 2009, 104, 11–16. [Google Scholar] [CrossRef] [Green Version]
- Isir, A.B.; Baransel, C.; Nacak, M. An Information Theoretical Study of the Epistasis Between the CNR1 1359 G/A Polymorphism and the Taq1A and Taq1B DRD2 Polymorphisms: Assessing the Susceptibility to Cannabis Addiction in a Turkish Population. J. Mol. Neurosci. 2016, 58, 456–460. [Google Scholar] [CrossRef]
- Hirvonen, J.; Zanotti-Fregonara, P.; Umhau, J.C.; George, D.T.; Rallis-Frutos, D.; Lyoo, C.H.; Li, C.T.; Hines, C.S.; Sun, H.; Terry, G.E.; et al. Reduced cannabinoid CB1 receptor binding in alcohol dependence measured with positron emission tomography. Mol. Psychiatry 2013, 18, 916–921. [Google Scholar] [CrossRef] [Green Version]
- Hutchison, K.E.; Haughey, H.; Niculescu, M.; Schacht, J.; Kaiser, A.; Stitzel, J.; Horton, W.J.; Filbey, F. The incentive salience of alcohol: Translating the effects of genetic variant in CNR1. Arch. Gen. Psychiatry 2008, 65, 841–850. [Google Scholar] [CrossRef] [Green Version]
- Ketcherside, A.; Noble, L.J.; McIntyre, C.K.; Filbey, F.M. Cannabinoid Receptor 1 Gene by Cannabis Use Interaction on CB1 Receptor Density. Cannabis Cannabinoid Res. 2017, 2, 202–209. [Google Scholar] [CrossRef] [Green Version]
- Filbey, F.M.; Schacht, J.P.; Myers, U.S.; Chavez, R.S.; Hutchison, K.E. Individual and additive effects of the CNR1 and FAAH genes on brain response to marijuana cues. Neuropsychopharmacology 2010, 35, 967–975. [Google Scholar] [CrossRef] [Green Version]
- Haughey, H.M.; Marshall, E.; Schacht, J.P.; Louis, A.; Hutchison, K.E. Marijuana withdrawal and craving: Influence of the cannabinoid receptor 1 (CNR1) and fatty acid amide hydrolase (FAAH) genes. Addiction 2008, 103, 1678–1686. [Google Scholar] [CrossRef]
- Brands, B.; Mann, R.E.; Wickens, C.M.; Sproule, B.; Stoduto, G.; Sayer, G.S.; Burston, J.; Pan, J.F.; Matheson, J.; Stefan, C.; et al. Acute and residual effects of smoked cannabis: Impact on driving speed and lateral control, heart rate, and self-reported drug effects. Drug Alcohol Depend. 2019, 205, 107641. [Google Scholar] [CrossRef]
- Matheson, J.; Mann, R.E.; Sproule, B.; Huestis, M.A.; Wickens, C.M.; Stoduto, G.; George, T.P.; Rehm, J.; Le Foll, B.; Brands, B. Acute and residual mood and cognitive performance of young adults following smoked cannabis. Pharm. Biochem. Behav. 2020, 194, 172937. [Google Scholar] [CrossRef] [PubMed]
- McNair, D.; Loor, M.; Droppleman, L. Profile of Mood States; Educational and Industrial Testing Service: San Diego, CA, USA, 1971. [Google Scholar]
- Phan, L.; Jin, Y.; Zhang, H.; Qiang, W.; Shekhtman, E.; Shao, D.; Revoe, D.; Villamarin, R.; Ivanchenko, E.; Kimura, M.; et al. ALFA: Allele Frequency Aggregator. Available online: https://www.ncbi.nlm.nih.gov/snp/docs/gsr/alfa/ (accessed on 5 July 2021).
- Le Strat, Y.; Ramoz, N.; Horwood, J.; Falissard, B.; Hassler, C.; Romo, L.; Choquet, M.; Fergusson, D.; Gorwood, P. First positive reactions to cannabis constitute a priority risk factor for cannabis dependence. Addiction 2009, 104, 1710–1717. [Google Scholar] [CrossRef] [PubMed]
- Marcos, M.; Pastor, I.; de la Calle, C.; Barrio-Real, L.; Laso, F.J.; Gonzalez-Sarmiento, R. Cannabinoid receptor 1 gene is associated with alcohol dependence. Alcohol Clin. Exp. Res. 2012, 36, 267–271. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, L.G.; Samochowiec, J.; Finckh, U.; Fiszer-Piosik, E.; Horodnicki, J.; Wendel, B.; Rommelspacher, H.; Hoehe, M.R. Association of a CB1 cannabinoid receptor gene (CNR1) polymorphism with severe alcohol dependence. Drug Alcohol Depend. 2002, 65, 221–224. [Google Scholar] [CrossRef]
- Yang, Y.; Chen, Y.; Wu, J.; Xing, Y.; Zeng, F.; Huang, Y.; Cheng, B.W. Association study of CNR1, GAD1 and BDNF polymorphisms with male heroin dependence in the Dai population in Yunnan. Yi Chuan 2014, 36, 888–896. [Google Scholar] [CrossRef]
- Courtney, K.E.; Infante, M.A.; Brown, G.G.; Tapert, S.F.; Simmons, A.N.; Smith, T.L.; Schuckit, M.A. The Relationship between Regional Cerebral Blood Flow Estimates and Alcohol Problems at 5-Year Follow-Up: The Role of Level of Response. Alcohol Clin. Exp. Res. 2019, 43, 812–821. [Google Scholar] [CrossRef]
- Schuckit, M.A.; Smith, T.L.; Trim, R.S.; Heron, J.; Horwood, J.; Davis, J.; Hibbeln, J.; Team, A.S. The self-rating of the effects of alcohol questionnaire as a predictor of alcohol-related outcomes in 12-year-old subjects. Alcohol Alcohol. 2008, 43, 641–646. [Google Scholar] [CrossRef] [Green Version]
- D‘Silva, J.; Cohn, A.M.; Johnson, A.L.; Villanti, A.C. Differences in Subjective Experiences to First Use of Menthol and Nonmenthol Cigarettes in a National Sample of Young Adult Cigarette Smokers. Nicotine Tob Res. 2018, 20, 1062–1068. [Google Scholar] [CrossRef]
- DiFranza, J.R.; Savageau, J.A.; Fletcher, K.; Pbert, L.; O‘Loughlin, J.; McNeill, A.D.; Ockene, J.K.; Friedman, K.; Hazelton, J.; Wood, C.; et al. Susceptibility to nicotine dependence: The Development and Assessment of Nicotine Dependence in Youth 2 study. Pediatrics 2007, 120, e974–e983. [Google Scholar] [CrossRef] [Green Version]
- Buchmann, A.F.; Hohm, E.; Witt, S.H.; Blomeyer, D.; Jennen-Steinmetz, C.; Schmidt, M.H.; Esser, G.; Banaschewski, T.; Brandeis, D.; Laucht, M. Role of CNR1 polymorphisms in moderating the effects of psychosocial adversity on impulsivity in adolescents. J. Neural. Transm. 2015, 122, 455–463. [Google Scholar] [CrossRef]
- Ehlers, C.L.; Slutske, W.S.; Lind, P.A.; Wilhelmsen, K.C. Association between single nucleotide polymorphisms in the cannabinoid receptor gene (CNR1) and impulsivity in southwest California Indians. Twin Res. Hum. Genet. 2007, 10, 805–811. [Google Scholar] [CrossRef] [Green Version]
- de Wit, H. Impulsivity as a determinant and consequence of drug use: A review of underlying processes. Addict. Biol. 2009, 14, 22–31. [Google Scholar] [CrossRef]
- Reynolds, B. A review of delay-discounting research with humans: Relations to drug use and gambling. Behav. Pharm. 2006, 17, 651–667. [Google Scholar] [CrossRef]
- Qin, H.; Zeng, J.; Chen, H.; Deng, L.; Su, L. Can Your DNA Influence Your Bet-Placing? The Impact of Cannabinoid Receptor 1 Gene on Gambling Tasks. Front. Hum. Neurosci. 2018, 12, 458. [Google Scholar] [CrossRef]
- Bidwell, L.C.; Metrik, J.; McGeary, J.; Palmer, R.H.; Francazio, S.; Knopik, V.S. Impulsivity, variation in the cannabinoid receptor (CNR1) and fatty acid amide hydrolase (FAAH) genes, and marijuana-related problems. J. Stud. Alcohol Drugs 2013, 74, 867–878. [Google Scholar] [CrossRef] [Green Version]
- van den Wildenberg, E.; Janssen, R.G.; Hutchison, K.E.; van Breukelen, G.J.; Wiers, R.W. Polymorphisms of the dopamine D4 receptor gene (DRD4 VNTR) and cannabinoid CB1 receptor gene (CNR1) are not strongly related to cue-reactivity after alcohol exposure. Addict. Biol. 2007, 12, 210–220. [Google Scholar] [CrossRef]
- Chen, X.; Williamson, V.S.; An, S.S.; Hettema, J.M.; Aggen, S.H.; Neale, M.C.; Kendler, K.S. Cannabinoid receptor 1 gene association with nicotine dependence. Arch. Gen. Psychiatry 2008, 65, 816–824. [Google Scholar] [CrossRef]
- Evans, D.E.; Sutton, S.K.; Jentink, K.G.; Lin, H.Y.; Park, J.Y.; Drobes, D.J. Cannabinoid receptor 1 (CNR1) gene variant moderates neural index of cognitive disruption during nicotine withdrawal. Genes Brain Behav. 2016, 15, 621–626. [Google Scholar] [CrossRef] [Green Version]
- Herman, A.I.; Kranzler, H.R.; Cubells, J.F.; Gelernter, J.; Covault, J. Association study of the CNR1 gene exon 3 alternative promoter region polymorphisms and substance dependence. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2006, 141B, 499–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okahisa, Y.; Kodama, M.; Takaki, M.; Inada, T.; Uchimura, N.; Yamada, M.; Iwata, N.; Iyo, M.; Sora, I.; Ozaki, N.; et al. Association Study of Two Cannabinoid Receptor Genes, CNR1 and CNR2, with Methamphetamine Dependence. Curr. Neuropharmacol. 2011, 9, 183–189. [Google Scholar] [CrossRef] [Green Version]
- Agrawal, A.; Chou, Y.L.; Carey, C.E.; Baranger, D.A.A.; Zhang, B.; Sherva, R.; Wetherill, L.; Kapoor, M.; Wang, J.C.; Bertelsen, S.; et al. Genome-wide association study identifies a novel locus for cannabis dependence. Mol. Psychiatry 2018, 23, 1293–1302. [Google Scholar] [CrossRef] [Green Version]
- Cornelis, M.C.; Flint, A.; Field, A.E.; Kraft, P.; Han, J.; Rimm, E.B.; van Dam, R.M. A genome-wide investigation of food addiction. Obesity 2016, 24, 1336–1341. [Google Scholar] [CrossRef] [Green Version]
- Levran, O.; Correa da Rosa, J.; Randesi, M.; Rotrosen, J.; Adelson, M.; Kreek, M.J. A non-coding CRHR2 SNP rs255105, a cis-eQTL for a downstream lincRNA AC005154.6, is associated with heroin addiction. PLoS ONE 2018, 13, e0199951. [Google Scholar] [CrossRef]
- Levran, O.; Randesi, M.; Rotrosen, J.; Ott, J.; Adelson, M.; Kreek, M.J. A 3′ UTR SNP rs885863, a cis-eQTL for the circadian gene VIPR2 and lincRNA 689, is associated with opioid addiction. PLoS ONE 2019, 14, e0224399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minica, C.C.; Verweij, K.J.H.; van der Most, P.J.; Mbarek, H.; Bernard, M.; van Eijk, K.R.; Lind, P.A.; Liu, M.Z.; Maciejewski, D.F.; Palviainen, T.; et al. Genome-wide association meta-analysis of age at first cannabis use. Addiction 2018, 113, 2073–2086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheung, E.Y.Y.; Phillips, C.; Eduardoff, M.; Lareu, M.V.; McNevin, D. Performance of ancestry-informative SNP and microhaplotype markers. Forensic. Sci. Int. Genet. 2019, 43, 102141. [Google Scholar] [CrossRef] [PubMed]
- Magi, R.; Horikoshi, M.; Sofer, T.; Mahajan, A.; Kitajima, H.; Franceschini, N.; McCarthy, M.I.; Cogent-Kidney Consortium, T.D.G.C.; Morris, A.P. Trans-ethnic meta-regression of genome-wide association studies accounting for ancestry increases power for discovery and improves fine-mapping resolution. Hum. Mol. Genet. 2017, 26, 3639–3650. [Google Scholar] [CrossRef] [Green Version]
- Zanetti, D.; Weale, M.E. Transethnic differences in GWAS signals: A simulation study. Ann. Hum. Genet. 2018, 82, 280–286. [Google Scholar] [CrossRef]
- Grotenhermen, F. Pharmacokinetics and pharmacodynamics of cannabinoids. Clin. Pharm. 2003, 42, 327–360. [Google Scholar] [CrossRef]
- Wong, K.U.; Baum, C.R. Acute Cannabis Toxicity. Pediatr. Emerg. Care 2019, 35, 799–804. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Participant Demographics | Total (n = 52) | Male (n = 36) | Female (n = 16) |
|---|---|---|---|
| Mean (SD) | |||
| Age (years) | 22.4 (1.9) | 22.1 (2.0) | 22.9 (1.5) |
| BMI (kg/m2) | 24.6 (4.6) | 25.2 (4.9) | 23.1 (3.4) |
| Cannabis Smoking Frequency (times/week) | 2.5 (0.9) | 2.6 (0.8) | 2.4 (1.1) |
| CNR1 Polymorphic Areas | Homozygous Individuals for the Major Allele | Carriers of the Minor Allele | ||
|---|---|---|---|---|
| rs1049353 | CC (n = 29) | TC + TT (n = 23) | ||
| Males (n = 19) | Females (n = 10) | Males (n = 17) | Females (n = 6) | |
| rs2023239 | TT (n = 35) | CT + CC (n = 17) | ||
| Males (n = 25) | Females (n = 10) | Males (n = 11) | Females (n = 6) | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Murphy, T.; Matheson, J.; Mann, R.E.; Brands, B.; Wickens, C.M.; Tiwari, A.K.; Zai, C.C.; Kennedy, J.; Le Foll, B. Influence of Cannabinoid Receptor 1 Genetic Variants on the Subjective Effects of Smoked Cannabis. Int. J. Mol. Sci. 2021, 22, 7388. https://doi.org/10.3390/ijms22147388
Murphy T, Matheson J, Mann RE, Brands B, Wickens CM, Tiwari AK, Zai CC, Kennedy J, Le Foll B. Influence of Cannabinoid Receptor 1 Genetic Variants on the Subjective Effects of Smoked Cannabis. International Journal of Molecular Sciences. 2021; 22(14):7388. https://doi.org/10.3390/ijms22147388
Chicago/Turabian StyleMurphy, Thomas, Justin Matheson, Robert E. Mann, Bruna Brands, Christine M. Wickens, Arun K. Tiwari, Clement C. Zai, James Kennedy, and Bernard Le Foll. 2021. "Influence of Cannabinoid Receptor 1 Genetic Variants on the Subjective Effects of Smoked Cannabis" International Journal of Molecular Sciences 22, no. 14: 7388. https://doi.org/10.3390/ijms22147388
APA StyleMurphy, T., Matheson, J., Mann, R. E., Brands, B., Wickens, C. M., Tiwari, A. K., Zai, C. C., Kennedy, J., & Le Foll, B. (2021). Influence of Cannabinoid Receptor 1 Genetic Variants on the Subjective Effects of Smoked Cannabis. International Journal of Molecular Sciences, 22(14), 7388. https://doi.org/10.3390/ijms22147388