
Reelin Affects Signaling Pathways of a Group of Inhibitory Neurons and the Development of Inhibitory Synapses in Primary Neurons

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents and Antibodies
2.2. Primary Neuronal Culture
2.3. Production of Recombinant Reelin and Semi-Purification
2.4. RNA Isolation and RT-PCR Analysis
2.5. Western Blot Analysis
2.6. Immunofluorescence
2.7. Dendrite Analysis and Quantification of Gephyrin–VGAT-Positive Clusters
2.8. Statistical Analysis
3. Results
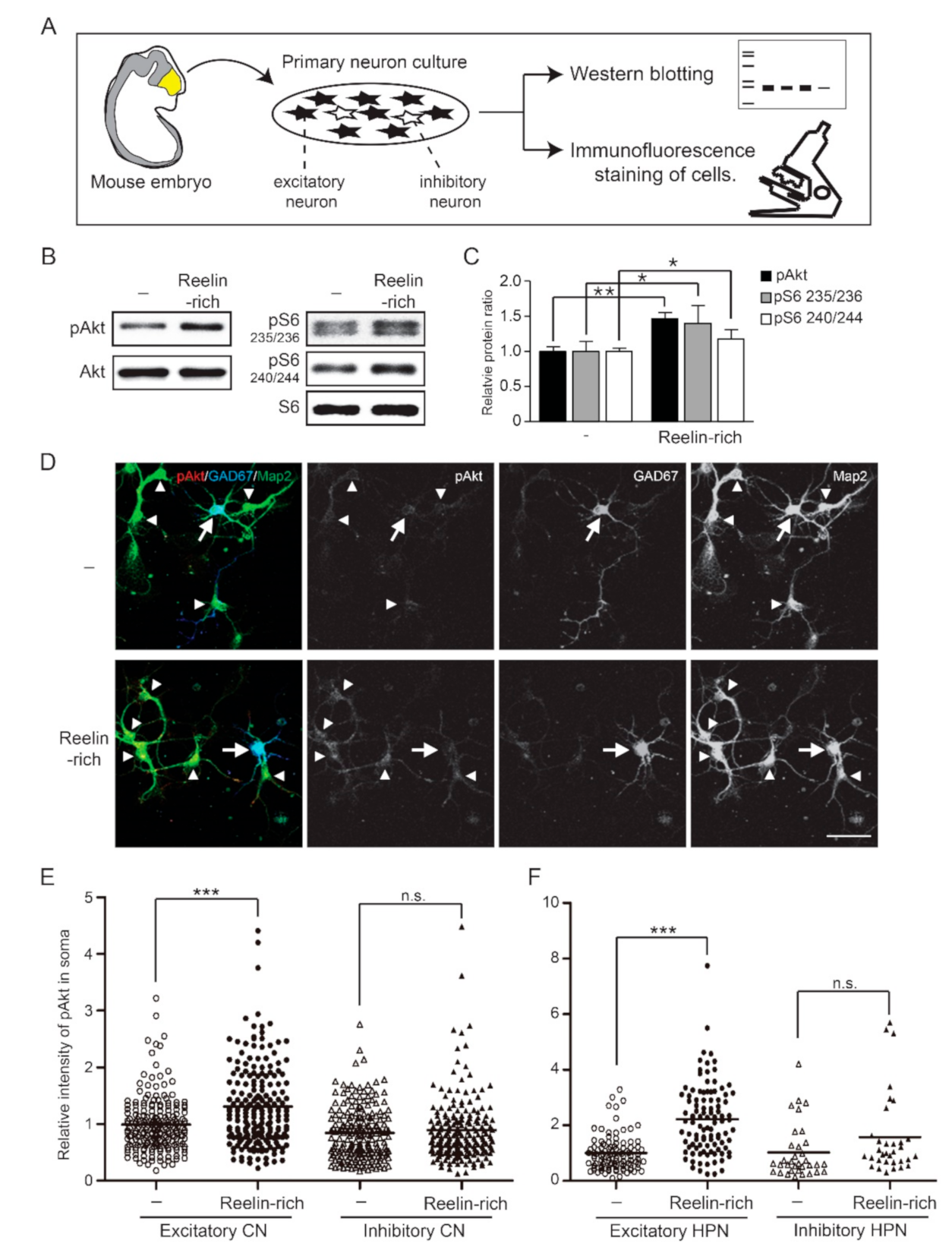
3.1. Reelin Does Not Phosphorylate Akt or S6 Protein in Numerous Inhibitory Neurons
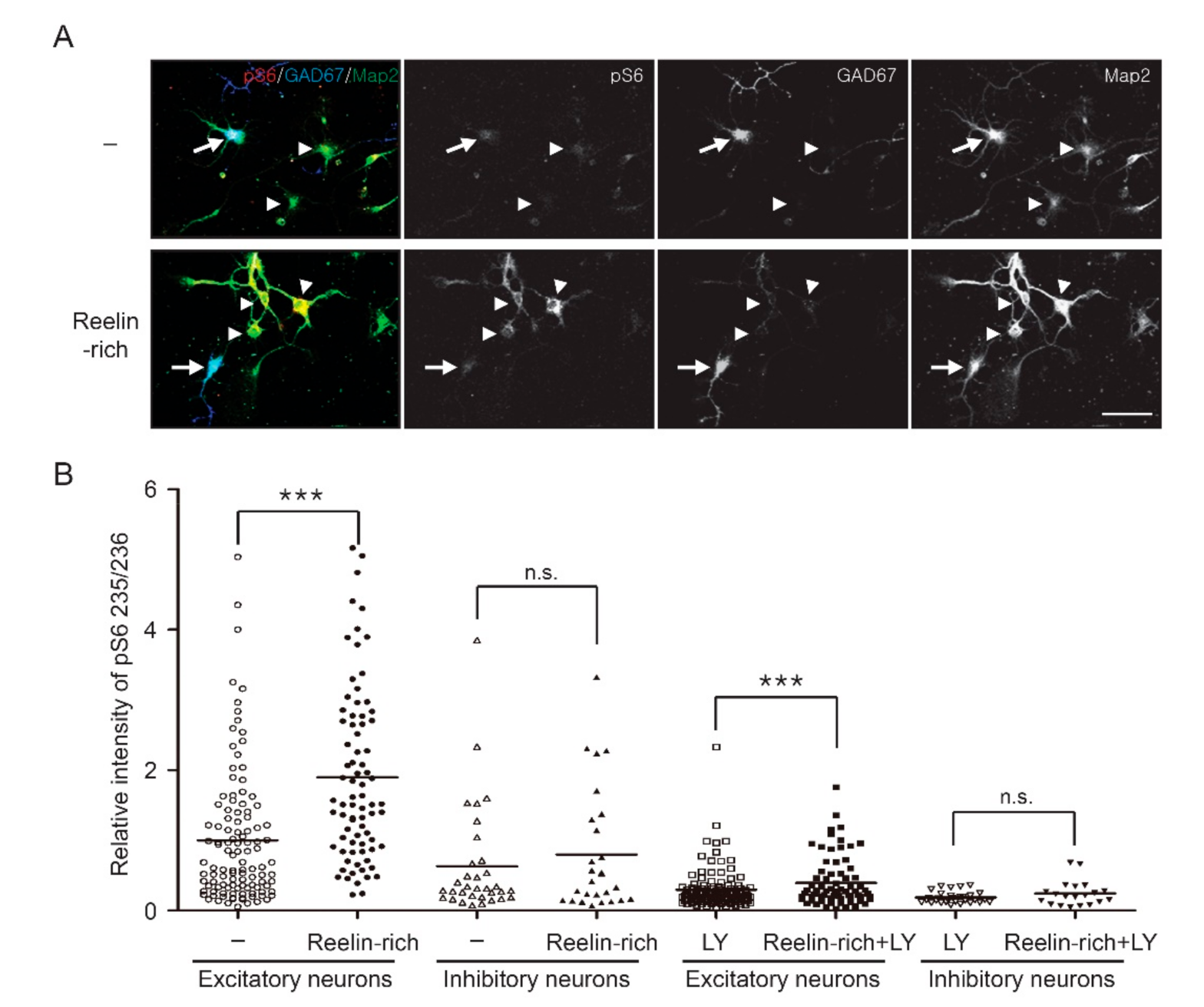
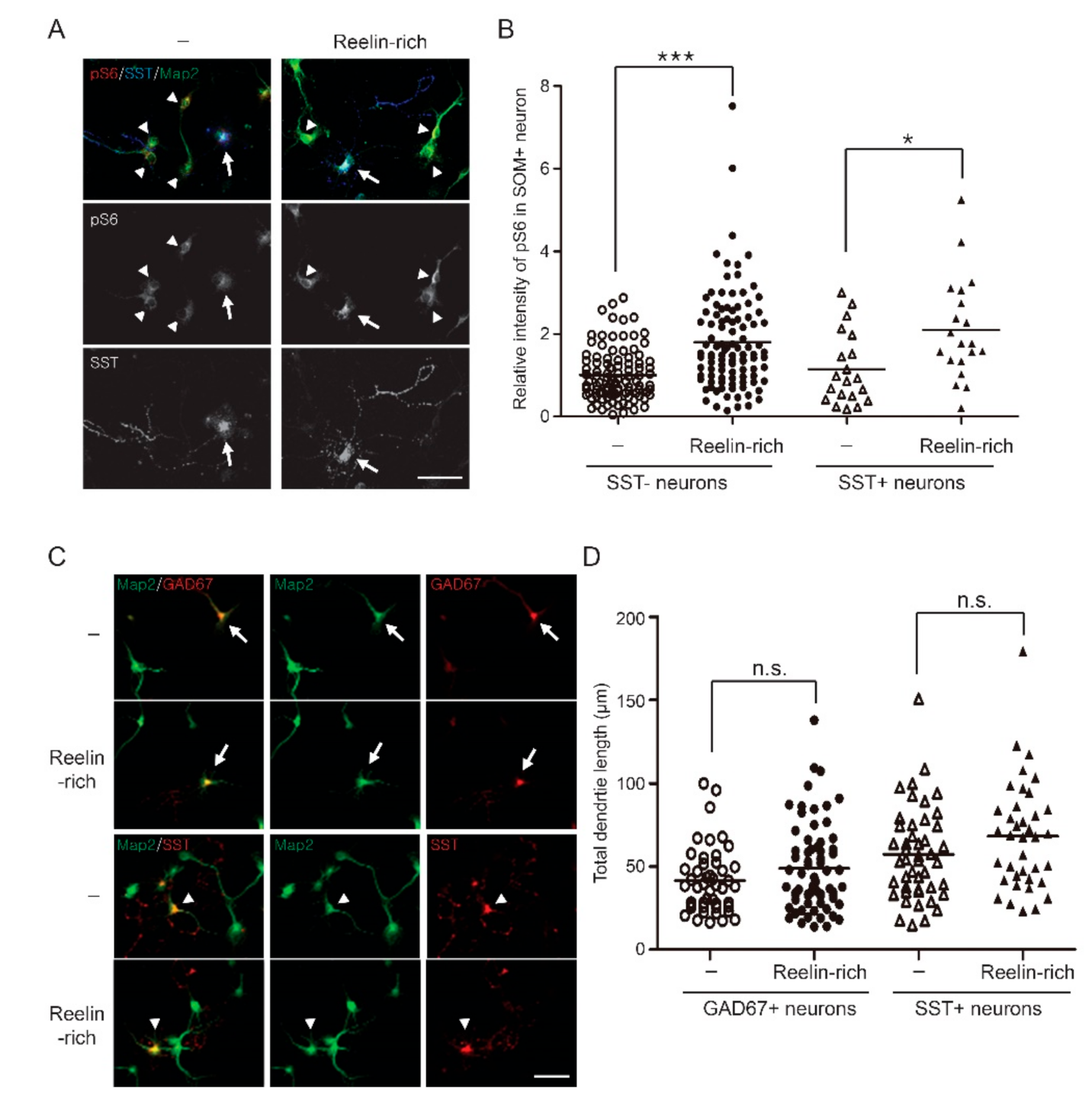
3.2. Reelin-Rich Protein Supplementation Induces Phosphorylation of S6 in Somatostatin-Positive Inhibitory Neurons
3.3. Reelin-Rich Protein Supplementation Does Not Affect Dendrite Outgrowth of Inhibitory Neurons in Primary Cultures
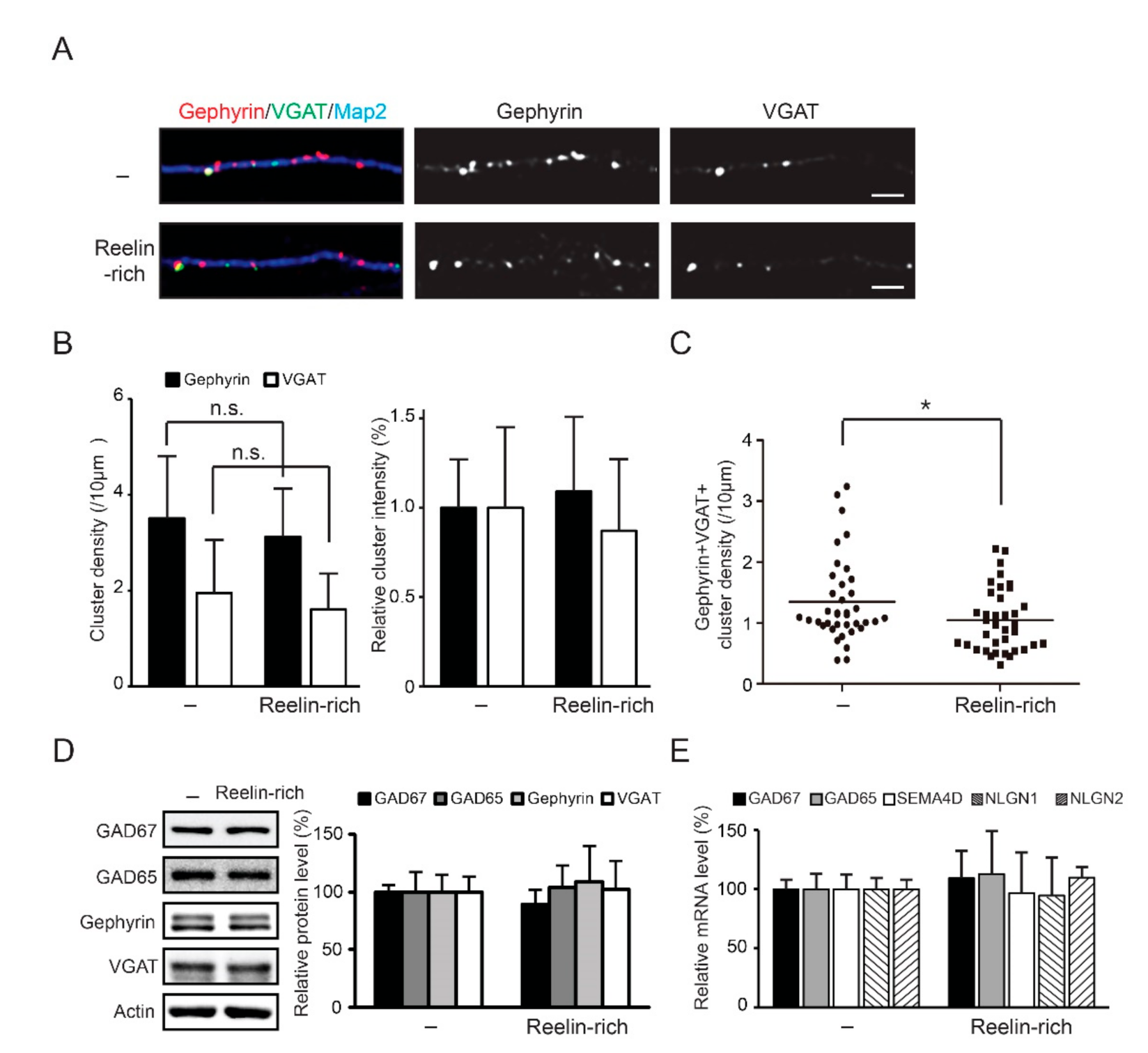
3.4. Reelin-Rich Protein Supplementation Reduces the Density of Inhibitory Synapses
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- D’Arcangelo, G.; Miao, G.G.; Chen, S.-C.; Scares, H.D.; Morgan, J.I.; Curran, T. A protein related to extracellular matrix proteins deleted in the mouse mutant reeler. Nat. Cell Biol. 1995, 374, 719–723. [Google Scholar] [CrossRef]
- Alcantara, S.; Ruiz, M.; D’Arcangelo, G.; Ezan, F.; de Lecea, L.; Curran, T.; Sotelo, C.; Soriano, E. Regional and cellular patterns of reelin mRNA expression in the forebrain of the developing and adult mouse. J. Neurosci. Off. J. Soc. Neurosci. 1998, 18, 7779–7799. [Google Scholar] [CrossRef] [Green Version]
- Lee, G.H.; D’Arcangelo, G. New Insights into Reelin-Mediated Signaling Pathways. Front. Cell. Neurosci. 2016, 10, 122. [Google Scholar] [CrossRef]
- Lee, G.H.; Chhangawala, Z.; von Daake, S.; Savas, J.N.; Yates, J.R.; Comoletti, D.; D’Arcangelo, G. Reelin Induces Erk1/2 Signaling in Cortical Neurons Through a Non-canonical Pathway*. J. Biol. Chem. 2014, 289, 20307–20317. [Google Scholar] [CrossRef] [Green Version]
- Trotter, J.; Lee, G.H.; Kazdoba, T.M.; Crowell, B.; Domogauer, J.; Mahoney, H.M.; Franco, S.J.; Müller, U.; Weeber, E.J.; D’Arcangelo, G. Dab1 is required for synaptic plasticity and associative learning. J. Neurosci. 2013, 33, 15652–15668. [Google Scholar] [CrossRef] [Green Version]
- Niu, S.; Yabut, O.; D’Arcangelo, G. The Reelin Signaling Pathway Promotes Dendritic Spine Development in Hippocampal Neurons. J. Neurosci. 2008, 28, 10339–10348. [Google Scholar] [CrossRef]
- Niu, S.; Renfro, A.; Quattrocchi, C.C.; Sheldon, M.; D’Arcangelo, G. Reelin promotes hippocampal dendrite development through the VLDLR/ApoER2-Dab1 pathway. Neuron 2004, 41, 71–84. [Google Scholar] [CrossRef] [Green Version]
- Da Costa, E.M. REELIN and Schizophrenia: A Disease at the Interface of the Genome and the Epigenome. Mol. Interv. 2002, 2, 47–57. [Google Scholar] [CrossRef]
- De Rubeis, S.; He, X.; Goldberg, A.P.; Poultney, C.S.; Samocha, K.; Cicek, A.E.; Kou, Y.; Liu, L.; Fromer, M.; Walker, S.; et al. Synaptic, transcriptional and chromatin genes disrupted in autism. Nat. Cell Biol. 2014, 515, 209–215. [Google Scholar] [CrossRef]
- Fatemi, S.H.; A Earle, J.; McMenomy, T. Reduction in Reelin immunoreactivity in hippocampus of subjects with schizophrenia, bipolar disorder and major depression. Mol. Psychiatry 2000, 5, 654–663. [Google Scholar] [CrossRef] [Green Version]
- Telese, F.; Ma, Q.; Perez, P.M.; Notani, D.; Oh, S.; Li, W.; Comoletti, D.; Ohgi, K.A.; Taylor, H.; Rosenfeld, M.G. LRP8-Reelin-Regulated Neuronal Enhancer Signature Underlying Learning and Memory Formation. Neuron 2015, 86, 696–710. [Google Scholar] [CrossRef] [Green Version]
- Giovannetti, E.A.; Fuhrmann, M. Unsupervised excitation: GABAergic dysfunctions in Alzheimer’s disease. Brain Res. 2019, 1707, 216–226. [Google Scholar] [CrossRef]
- Chao, H.-T.; Chen, H.; Samaco, R.; Xue, M.; Chahrour, M.; Yoo, J.; Neul, J.; Gong, S.; Lu, H.-C.; Heintz, N.; et al. Dysfunction in GABA signalling mediates autism-like stereotypies and Rett syndrome phenotypes. Nat. Cell Biol. 2010, 468, 263–269. [Google Scholar] [CrossRef]
- Lisman, J. Excitation, inhibition, local oscillations, or large-scale loops: What causes the symptoms of schizophrenia? Curr. Opin. Neurobiol. 2012, 22, 537–544. [Google Scholar] [CrossRef] [Green Version]
- Cline, H. Synaptogenesis: A Balancing Act between Excitation and Inhibition. Curr. Biol. 2005, 15, R203–R205. [Google Scholar] [CrossRef] [Green Version]
- Rubenstein, J.L. Three hypotheses for developmental defects that may underlie some forms of autism spectrum disorder. Curr. Opin. Neurol. 2010, 23, 118–123. [Google Scholar] [CrossRef] [Green Version]
- Kaila, K.; Ruusuvuori, E.; Seja, P.; Voipio, J.; Puskarjov, M. GABA actions and ionic plasticity in epilepsy. Curr. Opin. Neurobiol. 2014, 26, 34–41. [Google Scholar] [CrossRef] [Green Version]
- Curley, A.A.; Arion, M.; Volk, D.W.; Asafu-Adjei, J.K.; Sampson, A.R.; Fish, K.N.; Lewis, D.A. Cortical Deficits of Glutamic Acid Decarboxylase 67 Expression in Schizophrenia: Clinical, Protein, and Cell Type-Specific Features. Am. J. Psychiatry 2011, 168, 921–929. [Google Scholar] [CrossRef] [Green Version]
- Pla, R.; Borrell, V.; Flames, N.; Marín, O. Layer Acquisition by Cortical GABAergic Interneurons Is Independent of Reelin Signaling. J. Neurosci. 2006, 26, 6924–6934. [Google Scholar] [CrossRef] [Green Version]
- Pahle, J.; Muhia, M.; Wagener, R.J.; Tippmann, A.; Bock, H.H.; Graw, J.; Herz, J.; Staiger, J.F.; Drakew, A.; Kneussel, M.; et al. Selective Inactivation of Reelin in Inhibitory Interneurons Leads to Subtle Changes in the Dentate Gyrus but Leaves Cortical Layering and Behavior Unaffected. Cereb. Cortex 2019, 30, 1688–1707. [Google Scholar] [CrossRef]
- Kuzirian, M.S.; Paradis, S. Emerging themes in GABAergic synapse development. Prog. Neurobiol. 2011, 95, 68–87. [Google Scholar] [CrossRef] [Green Version]
- Lu, W.; Bromley-Coolidge, S.; Li, J. Regulation of GABAergic synapse development by postsynaptic membrane proteins. Brain Res. Bull. 2017, 129, 30–42. [Google Scholar] [CrossRef] [Green Version]
- Arimura, N.; Kaibuchi, K. Neuronal polarity: From extracellular signals to intracellular mechanisms. Nat. Rev. Neurosci. 2007, 8, 194–205. [Google Scholar] [CrossRef]
- Nikolaeva, I.; Kazdoba, T.M.; Crowell, B.; D’Arcangelo, G. Differential roles for Akt and mTORC1 in the hypertrophy of Pten mutant neurons, a cellular model of brain overgrowth disorders. Neuroscience 2017, 354, 196–207. [Google Scholar] [CrossRef]
- Takano, T.; Xu, C.; Funahashi, Y.; Namba, T.; Kaibuchi, K. Neuronal polarization. Development 2015, 142, 2088–2093. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Markram, H.; Goodman, P.H.; Berger, T.K.; Goldmann-Rakic, P.; Goldman-Rakic, P.S. Heterogeneity in the pyramidal network of the medial prefrontal cortex. Nat. Neurosci. 2006, 9, 534–542. [Google Scholar] [CrossRef]
- Previtera, M.L.; Firestein, B.L. Glutamate affects dendritic morphology of neurons grown on compliant substrates. Biotechnol. Prog. 2015, 31, 1128–1132. [Google Scholar] [CrossRef] [Green Version]
- Nelson, S.B.; Sugino, K.; Hempel, C.M. The problem of neuronal cell types: A physiological genomics approach. Trends Neurosci. 2006, 29, 339–345. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Rudy, B.; Fishell, G.; Lee, S.; Leffler, J.H. Three groups of interneurons account for nearly 100% of neocortical GABAergic neurons. Dev. Neurobiol. 2010, 71, 45–61. [Google Scholar] [CrossRef] [Green Version]
- Harris, K.D.; Hochgerner, H.; Skene, N.; Magno, L.; Katona, L.; Gonzales, C.B.; Somogyi, P.; Kessaris, N.; Linnarsson, S.; Leffler, J.H. Classes and continua of hippocampal CA1 inhibitory neurons revealed by single-cell transcriptomics. PLoS Biol. 2018, 16, e2006387. [Google Scholar] [CrossRef] [Green Version]
- Nitsch, R.; Bergmann, I.; Küppers, K.; Mueller, G.; Frotscher, M. Late appearance of parvalbumin-immunoreactivity in the development of GABAergic neurons in the rat hippocampus. Neurosci. Lett. 1990, 118, 147–150. [Google Scholar] [CrossRef]
- Jossin, Y.; Goffinet, A.M. Reelin Signals through Phosphatidylinositol 3-Kinase and Akt To Control Cortical Development and through mTor To Regulate Dendritic Growth. Mol. Cell. Biol. 2007, 27, 7113–7124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boivin, J.R.; Nedivi, E. Functional implications of inhibitory synapse placement on signal processing in pyramidal neuron dendrites. Curr. Opin. Neurobiol. 2018, 51, 16–22. [Google Scholar] [CrossRef]
- Artinian, J.; Jordan, A.; Khlaifia, A.; Honoré, E.; La Fontaine, A.; Racine, A.-S.; Laplante, I.; Lacaille, J.-C. Regulation of Hippocampal Memory by mTORC1 in Somatostatin Interneurons. J. Neurosci. 2019, 39, 8439–8456. [Google Scholar] [CrossRef] [PubMed]
- Ventruti, A.; Kazdoba, T.; Niu, S.; D’Arcangelo, G. Reelin deficiency causes specific defects in the molecular composition of the synapses in the adult brain. Neuroscience 2011, 189, 32–42. [Google Scholar] [CrossRef]
- Rice, D.; Sheldon, M.; D’Arcangelo, G.; Nakajima, K.; Goldowitz, D.; Curran, T. Disabled-1 acts downstream of Reelin in a signaling pathway that controls laminar organization in the mammalian brain. Development 1998, 125, 3719–3729. [Google Scholar] [CrossRef]
- Tasic, B.; Menon, V.; Nguyen, T.N.; Kim, T.K.; Jarsky, T.; Yao, Z.; Levi, B.P.; Graybuck, L.; Sorensen, S.A.; A Dolbeare, T.; et al. Adult mouse cortical cell taxonomy revealed by single cell transcriptomics. Nat. Neurosci. 2016, 19, 335–346. [Google Scholar] [CrossRef] [Green Version]
- Loo, L.; Simon, J.M.; Xing, L.; McCoy, E.S.; Niehaus, J.K.; Guo, J.; Anton, E.S.; Zylka, M.J. Single-cell transcriptomic analysis of mouse neocortical development. Nat. Commun. 2019, 10, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Rosato-Siri, M.; Grandolfo, M.; Ballerini, L. Activity-dependent modulation of GABAergic synapses in developing rat spinal networks in vitro. Eur. J. Neurosci. 2002, 16, 2123–2135. [Google Scholar] [CrossRef]
- Marty, S.; Wehrlé, R.; Sotelo, C. Neuronal Activity and Brain-Derived Neurotrophic Factor Regulate the Density of Inhibitory Synapses in Organotypic Slice Cultures of Postnatal Hippocampus. J. Neurosci. 2000, 20, 8087–8095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartman, K.N.; Pal, S.K.; Burrone, J.; Murthy, V. Activity-dependent regulation of inhibitory synaptic transmission in hippocampal neurons. Nat. Neurosci. 2006, 9, 642–649. [Google Scholar] [CrossRef]
- Aamodt, S.M.; Shi, J.; Colonnese, M.; Veras, W.; Constantine-Paton, M. Chronic NMDA Exposure Accelerates Development of GABAergic Inhibition in the Superior Colliculus. J. Neurophysiol. 2000, 83, 1580–1591. [Google Scholar] [CrossRef] [PubMed]
- Hamad, M.I.K.; Jbara, A.; Rabaya, O.; Petrova, P.; Daoud, S.; Melliti, N.; Meseke, M.; Lutz, D.; Petrasch-Parwez, E.; Schwitalla, J.C.; et al. Reelin signaling modulates GABA B receptor function in the neocortex. J. Neurochem. 2021, 156, 589–603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouamrane, L.; Scheyer, A.; Lassalle, O.; Iafrati, J.; Thomazeau, A.; Chavis, P. Reelin-Haploinsufficiency Disrupts the Developmental Trajectory of the E/I Balance in the Prefrontal Cortex. Front. Cell. Neurosci. 2017, 10, 308. [Google Scholar] [CrossRef] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.-E.; Lee, G.H. Reelin Affects Signaling Pathways of a Group of Inhibitory Neurons and the Development of Inhibitory Synapses in Primary Neurons. Int. J. Mol. Sci. 2021, 22, 7510. https://doi.org/10.3390/ijms22147510
Lee S-E, Lee GH. Reelin Affects Signaling Pathways of a Group of Inhibitory Neurons and the Development of Inhibitory Synapses in Primary Neurons. International Journal of Molecular Sciences. 2021; 22(14):7510. https://doi.org/10.3390/ijms22147510
Chicago/Turabian StyleLee, Seong-Eun, and Gum Hwa Lee. 2021. "Reelin Affects Signaling Pathways of a Group of Inhibitory Neurons and the Development of Inhibitory Synapses in Primary Neurons" International Journal of Molecular Sciences 22, no. 14: 7510. https://doi.org/10.3390/ijms22147510
APA StyleLee, S. -E., & Lee, G. H. (2021). Reelin Affects Signaling Pathways of a Group of Inhibitory Neurons and the Development of Inhibitory Synapses in Primary Neurons. International Journal of Molecular Sciences, 22(14), 7510. https://doi.org/10.3390/ijms22147510