
Bacterial Flagellar Filament: A Supramolecular Multifunctional Nanostructure



Abstract
:1. Introduction
2. Historical Overview
3. Flagellins
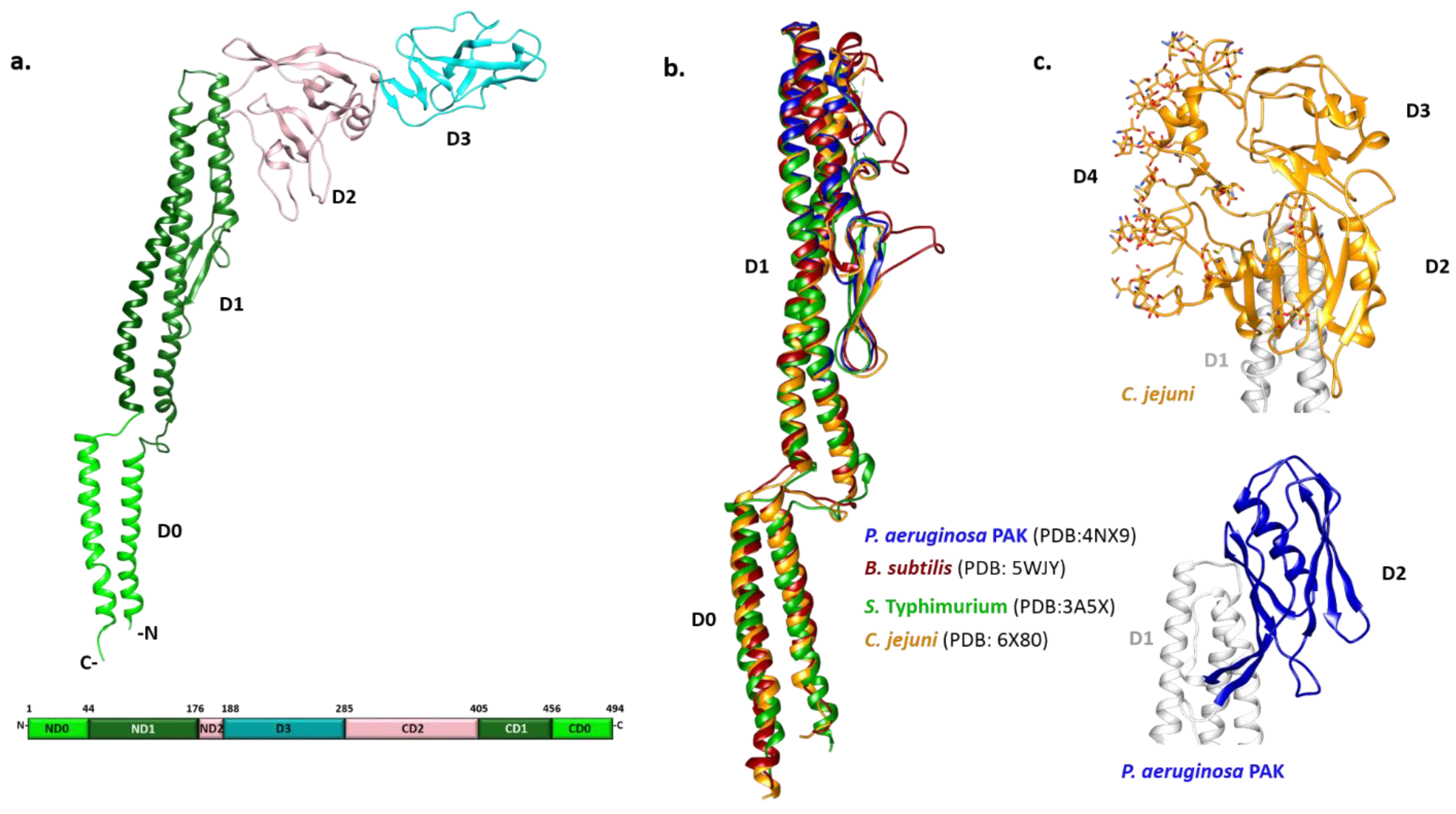
3.1. Flagellin Structure
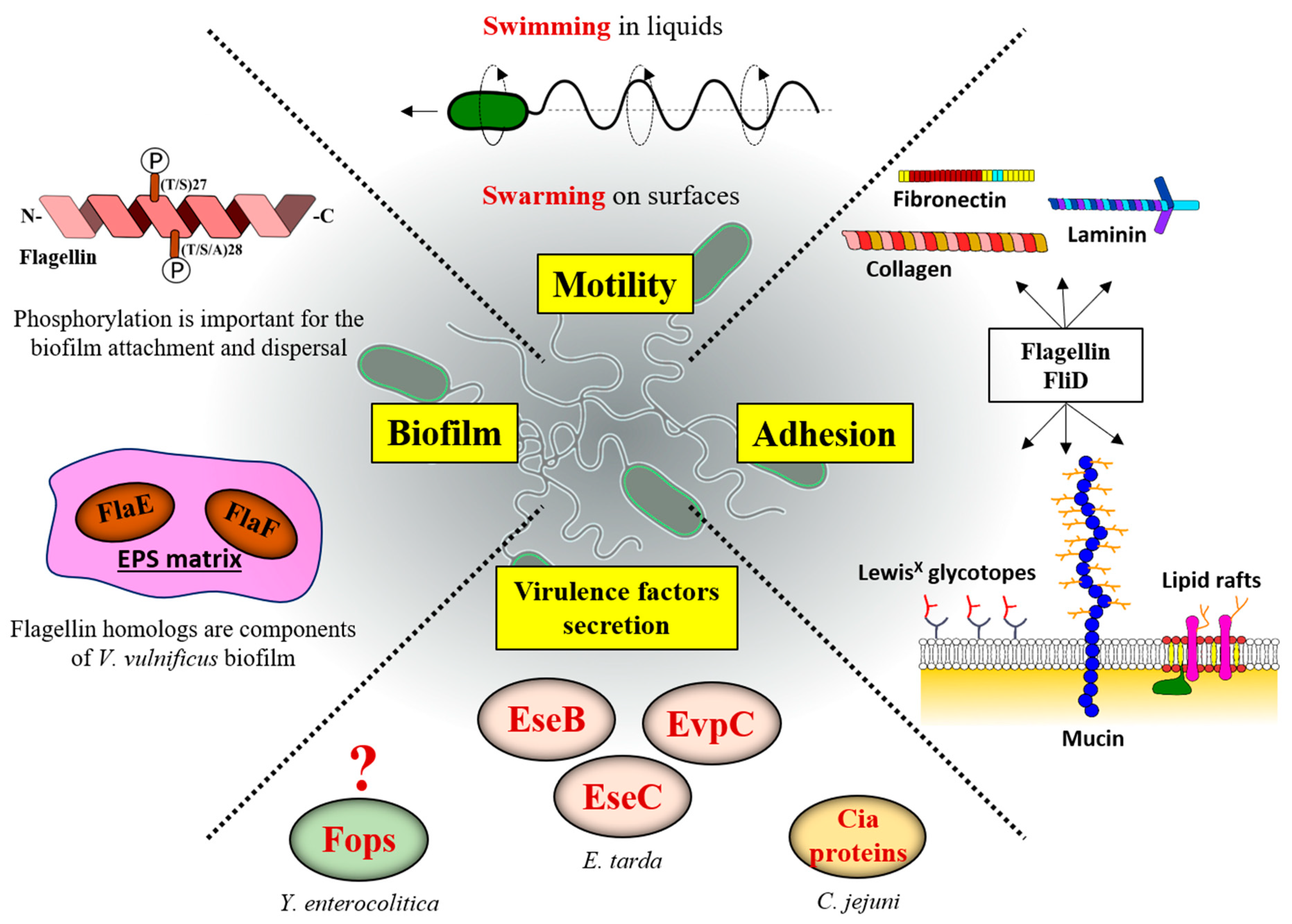
3.2. Posttranslational Modifications of Flagellins
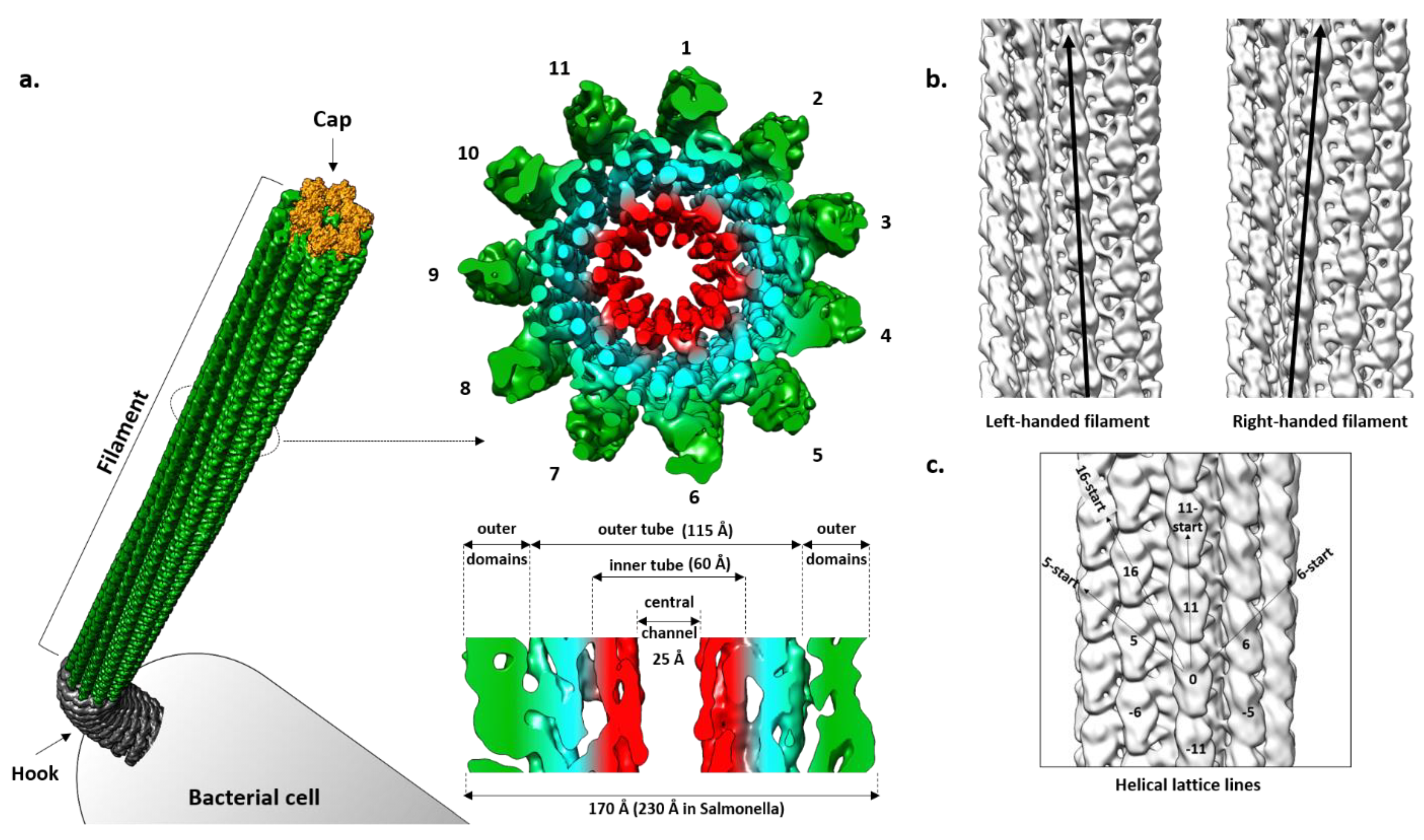
4. Filament
4.1. Comparison of L- and R-Filaments
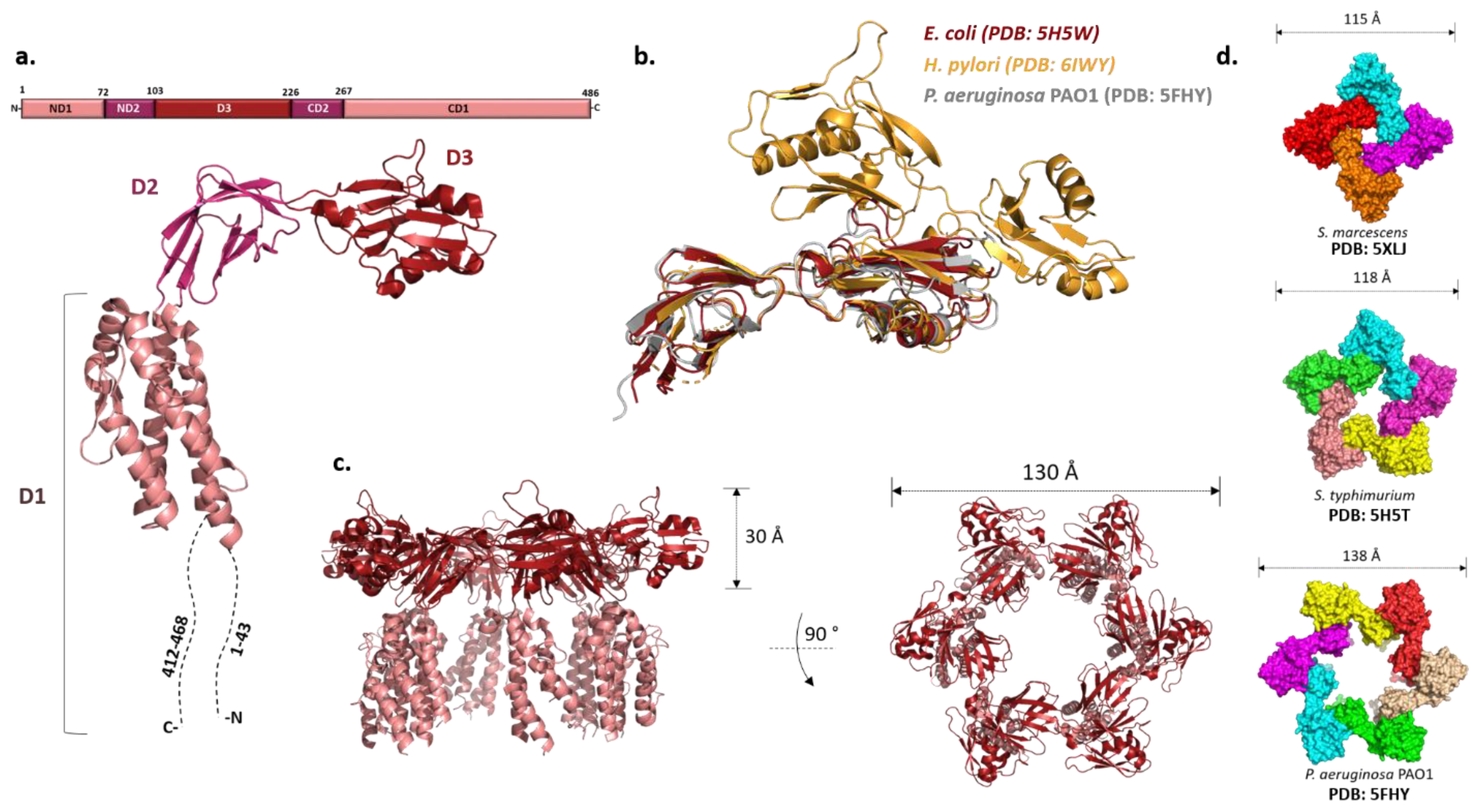
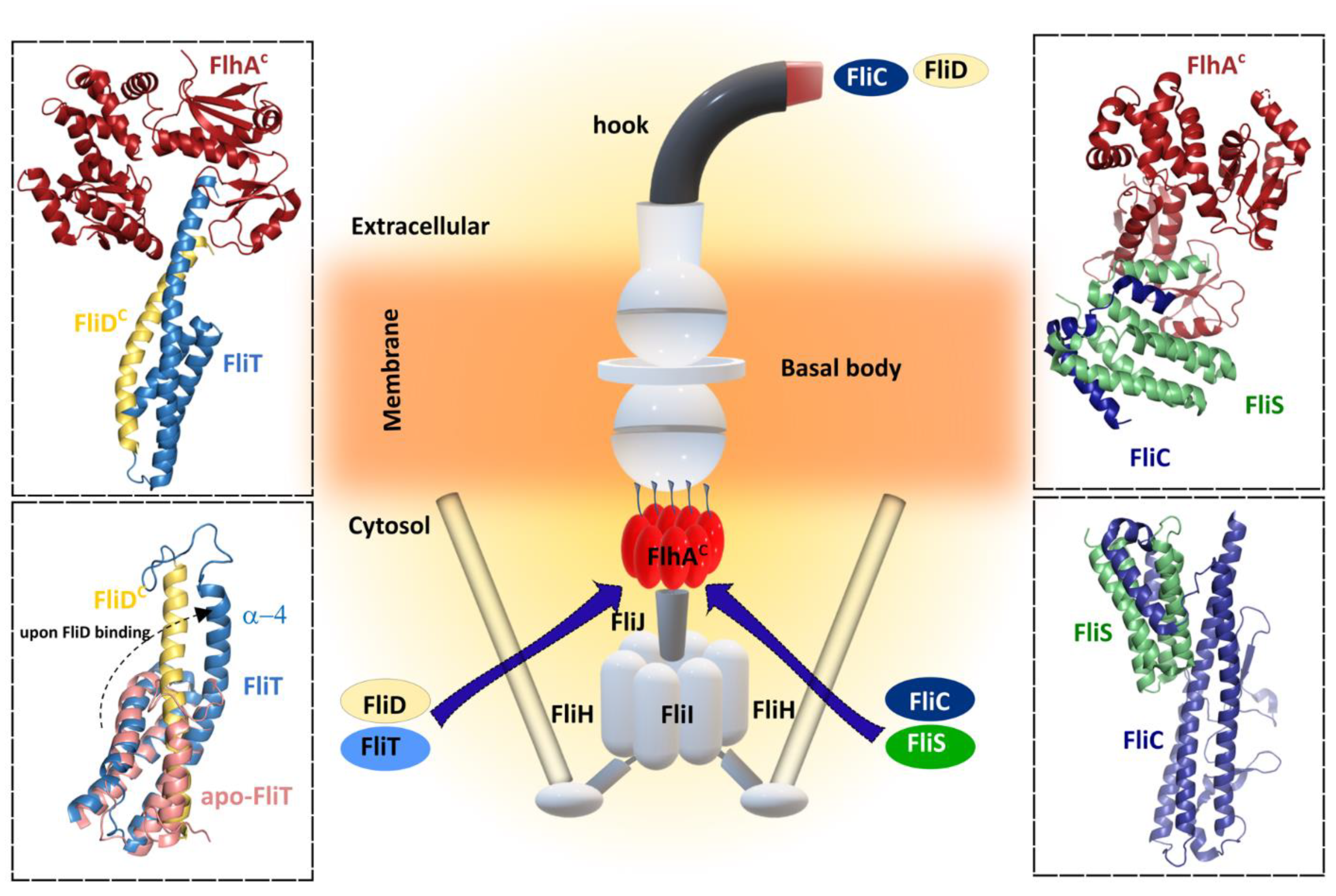
4.2. Filament Cap
5. Protein Synthesis, Export and Filament Elongation
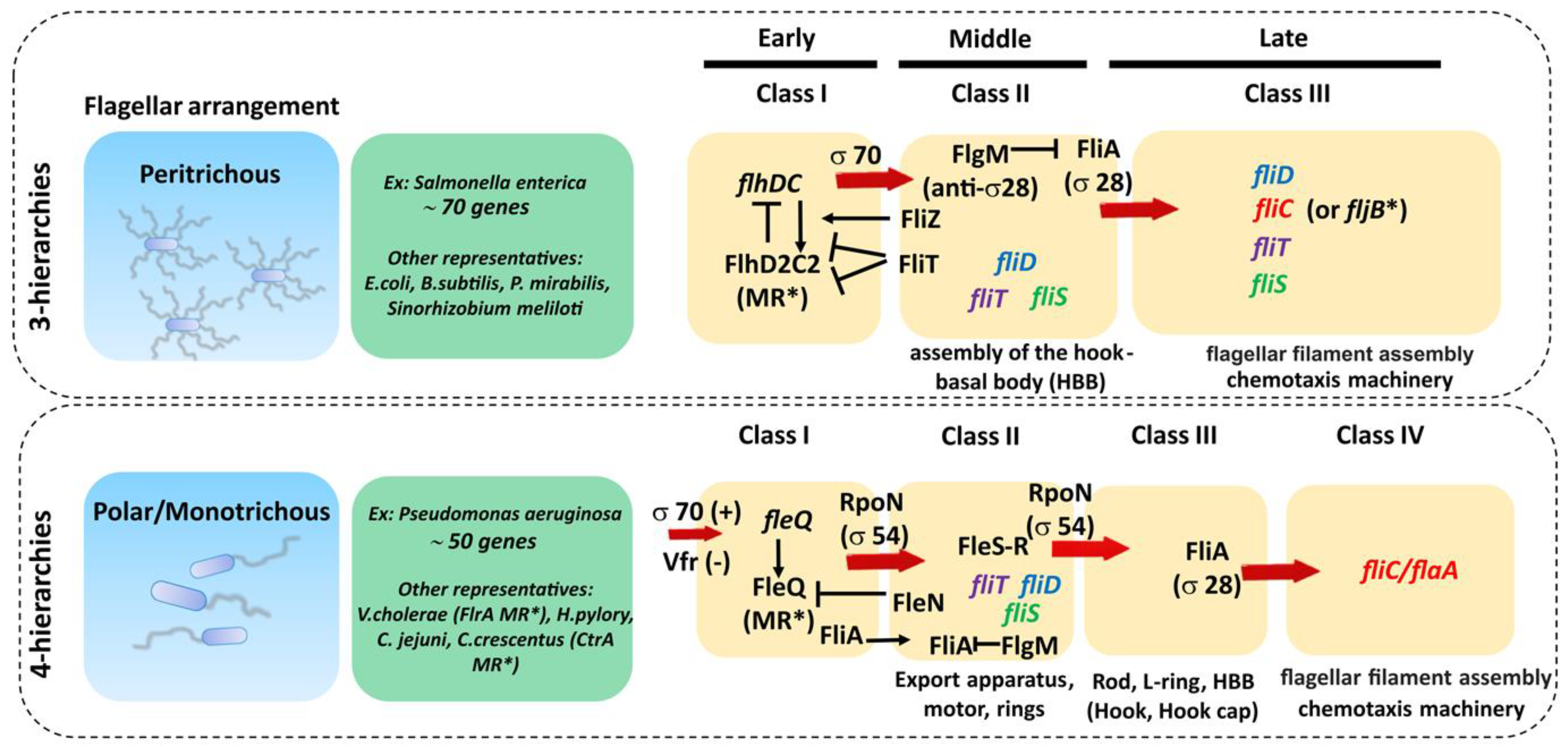
5.1. Genetic Regulation of FliC and FliD
5.2. Chaperones
5.2.1. FliS—Flagellin Chaperone
5.2.2. FliT—Chaperone of FliD
5.3. Flagellin Export and Assembly into the Growing Filament
6. Filament–Host Interaction
6.1. Filaments Are Directly Involved in Surface Adhesion
6.2. Flagellar Filament in Biofilm Formation
6.3. Secretion of Non-Flagellar Proteins during Host Invasion
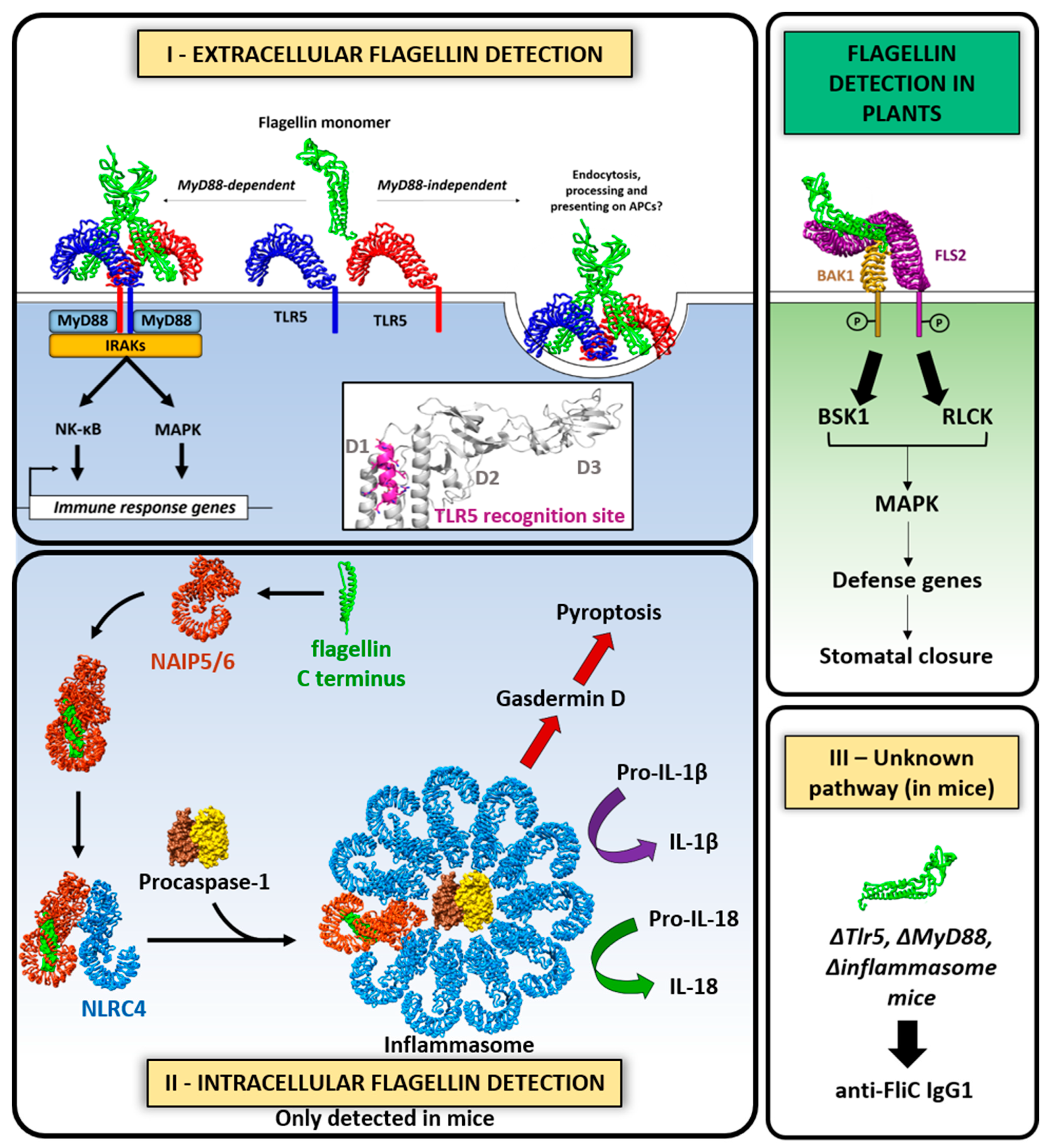
6.4. Host Immune Response to Flagella
6.5. Flagellin in Vaccines
6.6. Additional Applications of Flagellin
7. Future Perspectives
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Schuhmacher, J.S.; Thormann, K.M.; Bange, G. How bacteria maintain location and number of flagella? FEMS Microbiol. Rev. 2015, 39, 812–822. [Google Scholar] [CrossRef] [Green Version]
- Macnab, R.M. How bacteria assemble flagella. Annu. Rev. Microbiol. 2003, 57, 77–100. [Google Scholar] [CrossRef]
- Chaban, B.; Hughes, H.V.; Beeby, M. The flagellum in bacterial pathogens: For motility and a whole lot more. Semin. Cell Dev. Biol. 2015, 46, 91–103. [Google Scholar] [CrossRef] [Green Version]
- Larsen, S.H.; Reader, R.W.; Kort, E.N.; Tso, W.W.; Adler, J. Change in direction of flagellar rotation is the basis of the chemotactic response in Escherichia coli. Nature 1974, 249, 74–77. [Google Scholar] [CrossRef] [PubMed]
- Turner, L.; Ryu, W.S.; Berg, H.C. Real-time imaging of fluorescent flagellar filaments. J. Bacteriol. 2000, 182, 2793–2801. [Google Scholar] [CrossRef] [Green Version]
- Shimada, K.; Kamiya, R.; Asakura, S. Left-handed to right-handed helix conversion in Salmonella flagella. Nature 1975, 254, 332–334. [Google Scholar] [CrossRef] [PubMed]
- Chu, J.; Liu, J.; Hoover, T.R. Phylogenetic Distribution, Ultrastructure, and Function of Bacterial Flagellar Sheaths. Biomolecules 2020, 10, 363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, Y.; Liu, J. Architecture and Assembly of Periplasmic Flagellum. Microbiol. Spectr. 2019, 7. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, S.; Minamino, T. Flagella-Driven Motility of Bacteria. Biomolecules 2019, 9, 279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morimoto, Y.V.; Minamino, T. Architecture and Assembly of the Bacterial Flagellar Motor Complex. Subcell. Biochem. 2021, 96, 297–321. [Google Scholar] [CrossRef]
- Nord, A.L.; Pedaci, F. Mechanisms and Dynamics of the Bacterial Flagellar Motor. Adv. Exp. Med. Biol. 2020, 1267, 81–100. [Google Scholar] [CrossRef]
- Khan, S. The Architectural Dynamics of the Bacterial Flagellar Motor Switch. Biomolecules 2020, 10, 833. [Google Scholar] [CrossRef]
- Münch, R. Robert Koch. Microbes Infect. 2003, 5, 69–74. [Google Scholar] [CrossRef]
- Strick, J. Swimming against the tide: Adrianus Pijper and the debate over bacterial flagella, 1946–1956. Isis 1996, 87, 274–305. [Google Scholar] [CrossRef]
- Boivin, A.; Mesrobeanu, L. Sur la résistance à l’acide trichloracétique de l’antigène flagellaire (antigène H) du bacille typhique et sur la nature chimique possible de cet antigène. CR Soc. Biol. (Paris) 1938, 129, 136–138. [Google Scholar]
- Weibull, C. Some chemical and physico-chemical properties of the flagella of Proteus vulgaris. Biochim. Biophys. Acta 1948, 2, 351–361. [Google Scholar] [CrossRef]
- Weibull, C. Chemical and physicochemical properties of the flagella of Proteus vulgaris and Bacillus subtilis a comparison. Biochim. Biophys. Acta 1949, 3, 378–382. [Google Scholar] [CrossRef]
- Weibull, C. Electrophoretic and Titrimetric Measurements on Bacterial Flagella. Acta Chem. Scand. 1950, 4, 260–267. [Google Scholar] [CrossRef]
- Weibull, C. The Free Amino Groups of the Proteus Flagella Protein-Quantitative Determination of Dinitrophenyl Amino Acids Using Paper Chromatography. Acta Chem. Scand. 1953, 7, 335–339. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, T.; Rinker, J.N.; Koffler, H. Purification and chemical properties of flagellin. Arch. Biochem. Biophys. 1959, 84, 342–362. [Google Scholar] [CrossRef]
- Koffler, H.; Kobayashi, T. Purification of flagella and flagellin with ammonium sulfate. Arch. Biochem. Biophys. 1957, 67, 246–248. [Google Scholar] [CrossRef]
- Koffler, H.; Kobayashi, T.; Mallett, G.E. Cysteine-cystine content and the free amino groups of flagellin. Arch. Biochem. Biophys. 1956, 64, 509–511. [Google Scholar] [CrossRef]
- Astbury, W.T.; Weibull, C. X-ray diffraction study of the structure of bacterial flagella. Nature 1949, 163, 280–282. [Google Scholar] [CrossRef]
- Astbury, W.T.; Beighton, C.; Weibull, C. The structure of bacterial flagella. Symp. Soc. Exp. Biol. 1955, 9, 282–305. [Google Scholar]
- Ada, G.L.; Nossal, G.J.; Pye, J.; Abbot, A. Behaviour of Active Bacterial Antigens during the Induction of the Immune Response. I. Properties of Flagellar Antigens from Salmonella. Nature 1963, 199, 1257–1259. [Google Scholar] [CrossRef]
- Asakura, S.; Eguchi, G.; Iino, T. Reconstitution of bacterial flagella in vitro. J. Mol. Biol. 1964, 10, 42–56. [Google Scholar] [CrossRef]
- Lowy, J.; McDonough, M.W. Structure of Filaments Produced by Re-Aggregation of Salmonella Flagellin. Nature 1964, 204, 125–127. [Google Scholar] [CrossRef] [PubMed]
- Asakura, S.; Eguchi, G.; Iino, T. Unidirectional growth of Salmonella flagella in vitro. J. Mol. Biol. 1968, 35, 227–236. [Google Scholar] [CrossRef]
- Iino, T. Polarity of flagellar growth in salmonella. J. Gen. Microbiol. 1969, 56, 227–239. [Google Scholar] [CrossRef] [Green Version]
- Kerridge, D.; Horne, R.W.; Glauert, A.M. Structural components of flagella from Salmonella typhimurium. J. Mol. Biol. 1962, 4, 227–238. [Google Scholar] [CrossRef]
- Lowy, J.; Hanson, J. Structure of Bacterial Flagella. Nature 1964, 202, 538–540. [Google Scholar] [CrossRef] [PubMed]
- Champness, J.N. X-ray and optical diffraction studies of bacterial flagella. J. Mol. Biol. 1971, 56, 295–310. [Google Scholar] [CrossRef]
- O’Brien, E.J.; Bennett, P.M. Structure of Straight flagella from a mutant Salmonella. J. Mol. Biol. 1972, 70, 133–152. [Google Scholar] [CrossRef]
- Ikeda, T.; Asakura, S.; Kamiya, R. “Cap” on the tip of Salmonella flagella. J. Mol. Biol. 1985, 184, 735–737. [Google Scholar] [CrossRef]
- Ikeda, T.; Homma, M.; Iino, T.; Asakura, S.; Kamiya, R. Localization and stoichiometry of hook-associated proteins within Salmonella typhimurium flagella. J. Bacteriol. 1987, 169, 1168–1173. [Google Scholar] [CrossRef] [Green Version]
- Homma, M.; Kutsukake, K.; Iino, T.; Yamaguchi, S. Hook-associated proteins essential for flagellar filament formation in Salmonella typhimurium. J. Bacteriol. 1984, 157, 100–108. [Google Scholar] [CrossRef] [Green Version]
- Il Kim, M.; Lee, C.; Park, J.; Jeon, B.Y.; Hong, M. Crystal structure of Bacillus cereus flagellin and structure-guided fusion-protein designs. Sci. Rep. 2018, 8, 5814. [Google Scholar] [CrossRef]
- Beatson, S.A.; Minamino, T.; Pallen, M.J. Variation in bacterial flagellins: From sequence to structure. Trends Microbiol. 2006, 14, 151–155. [Google Scholar] [CrossRef]
- Steimle, A.; Menz, S.; Bender, A.; Ball, B.; Weber, A.N.R.; Hagemann, T.; Lange, A.; Maerz, J.K.; Parusel, R.; Michaelis, L.; et al. Flagellin hypervariable region determines symbiotic properties of commensal Escherichia coli strains. PLoS Biol. 2019, 17, e3000334. [Google Scholar] [CrossRef]
- Yoshioka, K.; Aizawa, S.; Yamaguchi, S. Flagellar filament structure and cell motility of Salmonella typhimurium mutants lacking part of the outer domain of flagellin. J. Bacteriol. 1995, 177, 1090–1093. [Google Scholar] [CrossRef] [Green Version]
- LaVallie, E.R.; Stahl, M.L. Cloning of the flagellin gene from Bacillus subtilis and complementation studies of an in vitro-derived deletion mutation. J. Bacteriol. 1989, 171, 3085–3094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faulds-Pain, A.; Birchall, C.; Aldridge, C.; Smith, W.D.; Grimaldi, G.; Nakamura, S.; Miyata, T.; Gray, J.; Li, G.; Tang, J.X.; et al. Flagellin redundancy in Caulobacter crescentus and its implications for flagellar filament assembly. J. Bacteriol. 2011, 193, 2695–2707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macnab, R.M. Genetics and biogenesis of bacterial flagella. Annu. Rev. Genet. 1992, 26, 131–158. [Google Scholar] [CrossRef] [PubMed]
- Silverman, M.; Zieg, J.; Simon, M. Flagellar-phase variation: Isolation of the rh1 gene. J. Bacteriol. 1979, 137, 517–523. [Google Scholar] [CrossRef] [Green Version]
- Ikeda, J.S.; Schmitt, C.K.; Darnell, S.C.; Watson, P.R.; Bispham, J.; Wallis, T.S.; Weinstein, D.L.; Metcalf, E.S.; Adams, P.; O’Connor, C.D.; et al. Flagellar phase variation of Salmonella enterica serovar Typhimurium contributes to virulence in the murine typhoid infection model but does not influence Salmonella-induced enteropathogenesis. Infect. Immun. 2001, 69, 3021–3030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horstmann, J.A.; Zschieschang, E.; Truschel, T.; de Diego, J.; Lunelli, M.; Rohde, M.; May, T.; Strowig, T.; Stradal, T.; Kolbe, M.; et al. Flagellin phase-dependent swimming on epithelial cell surfaces contributes to productive Salmonella gut colonisation. Cell Microbiol. 2017, 19, e12739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kostrzynska, M.; Betts, J.D.; Austin, J.W.; Trust, T.J. Identification, characterization, and spatial localization of two flagellin species in Helicobacter pylori flagella. J. Bacteriol. 1991, 173, 937–946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nuijten, P.J.; van Asten, F.J.; Gaastra, W.; van der Zeijst, B.A. Structural and functional analysis of two Campylobacter jejuni flagellin genes. J. Biol. Chem. 1990, 265, 17798–17804. [Google Scholar] [CrossRef]
- Wassenaar, T.M.; Bleumink-Pluym, N.M.; Newell, D.G.; Nuijten, P.J.; van der Zeijst, B.A. Differential flagellin expression in a flaA flaB+ mutant of Campylobacter jejuni. Infect. Immun. 1994, 62, 3901–3906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuhn, M.J.; Schmidt, F.K.; Farthing, N.E.; Rossmann, F.M.; Helm, B.; Wilson, L.G.; Eckhardt, B.; Thormann, K.M. Spatial arrangement of several flagellins within bacterial flagella improves motility in different environments. Nat. Commun. 2018, 9, 5369. [Google Scholar] [CrossRef] [Green Version]
- Guerry, P.; Alm, R.A.; Power, M.E.; Logan, S.M.; Trust, T.J. Role of two flagellin genes in Campylobacter motility. J. Bacteriol. 1991, 173, 4757–4764. [Google Scholar] [CrossRef] [Green Version]
- Lis, L.; Connerton, I.F. The Minor Flagellin of Campylobacter jejuni (FlaB) Confers Defensive Properties against Bacteriophage Infection. Front. Microbiol. 2016, 7, 1908. [Google Scholar] [CrossRef] [Green Version]
- Allison, J.S.; Dawson, M.; Drake, D.; Montie, T.C. Electrophoretic separation and molecular weight characterization of Pseudomonas aeruginosa H-antigen flagellins. Infect. Immun. 1985, 49, 770–774. [Google Scholar] [CrossRef] [Green Version]
- Morgan, J.A.; Bellingham, N.F.; Winstanley, C.; Ousley, M.A.; Hart, C.A.; Saunders, J.R. Comparison of flagellin genes from clinical and environmental Pseudomonas aeruginosa isolates. Appl. Environ. Microbiol. 1999, 65, 1175–1179. [Google Scholar] [CrossRef] [Green Version]
- Brimer, C.D.; Montie, T.C. Cloning and comparison of fliC genes and identification of glycosylation in the flagellin of Pseudomonas aeruginosa a-type strains. J. Bacteriol. 1998, 180, 3209–3217. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.Y.; Thanh, X.T.; Jeong, K.; Kim, S.B.; Pan, S.O.; Jung, C.H.; Hong, S.H.; Lee, S.E.; Rhee, J.H. Contribution of six flagellin genes to the flagellum biogenesis of Vibrio vulnificus and in vivo invasion. Infect. Immun. 2014, 82, 29–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tambalo, D.D.; Bustard, D.E.; Del Bel, K.L.; Koval, S.F.; Khan, M.F.; Hynes, M.F. Characterization and functional analysis of seven flagellin genes in Rhizobium leguminosarum bv. viciae. Characterization of R. leguminosarum flagellins. BMC Microbiol. 2010, 10, 219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vonderviszt, F.; Kanto, S.; Aizawa, S.-I.; Namba, K. Terminal regions of flagellin are disordered in solution. J. Mol. Biol. 1989, 209, 127–133. [Google Scholar] [CrossRef]
- Namba, K.; Yamashita, I.; Vonderviszt, F. Structure of the core and central channel of bacterial flagella. Nature 1989, 342, 648–654. [Google Scholar] [CrossRef] [PubMed]
- Samatey, F.A.; Imada, K.; Vonderviszt, F.; Shirakihara, Y.; Namba, K. Crystallization of the F41 fragment of flagellin and data collection from extremely thin crystals. J. Struct. Biol. 2000, 132, 106–111. [Google Scholar] [CrossRef]
- Samatey, F.A.; Imada, K.; Nagashima, S.; Vonderviszt, F.; Kumasaka, T.; Yamamoto, M.; Namba, K. Structure of the bacterial flagellar protofilament and implications for a switch for supercoiling. Nature 2001, 410, 331–337. [Google Scholar] [CrossRef]
- Yonekura, K.; Maki-Yonekura, S.; Namba, K. Complete atomic model of the bacterial flagellar filament by electron cryomicroscopy. Nature 2003, 424, 643–650. [Google Scholar] [CrossRef] [PubMed]
- Song, W.S.; Yoon, S.I. Crystal structure of FliC flagellin from Pseudomonas aeruginosa and its implication in TLR5 binding and formation of the flagellar filament. Biochem. Biophys. Res. Commun. 2014, 444, 109–115. [Google Scholar] [CrossRef]
- Kreutzberger, M.A.B.; Ewing, C.; Poly, F.; Wang, F.; Egelman, E.H. Atomic structure of the Campylobacter jejuni flagellar filament reveals how epsilon Proteobacteria escaped Toll-like receptor 5 surveillance. Proc. Natl. Acad. Sci. USA 2020, 117, 16985–16991. [Google Scholar] [CrossRef]
- Arora, S.K.; Bangera, M.; Lory, S.; Ramphal, R. A genomic island in Pseudomonas aeruginosa carries the determinants of flagellin glycosylation. Proc. Natl. Acad. Sci. USA 2001, 98, 9342–9347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schirm, M.; Kalmokoff, M.; Aubry, A.; Thibault, P.; Sandoz, M.; Logan, S.M. Flagellin from Listeria monocytogenes is glycosylated with beta-O-linked N-acetylglucosamine. J. Bacteriol. 2004, 186, 6721–6727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verma, A.; Schirm, M.; Arora, S.K.; Thibault, P.; Logan, S.M.; Ramphal, R. Glycosylation of b-Type flagellin of Pseudomonas aeruginosa: Structural and genetic basis. J. Bacteriol. 2006, 188, 4395–4403. [Google Scholar] [CrossRef] [Green Version]
- Arora, S.K.; Neely, A.N.; Blair, B.; Lory, S.; Ramphal, R. Role of motility and flagellin glycosylation in the pathogenesis of Pseudomonas aeruginosa burn wound infections. Infect. Immun. 2005, 73, 4395–4398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khemiri, A.; Naudin, B.; Franck, X.; Song, P.C.; Jouenne, T.; Cosette, P. N-glycosidase treatment with 18O labeling and de novo sequencing argues for flagellin FliC glycopolymorphism in Pseudomonas aeruginosa. Anal. Bioanal. Chem. 2013, 405, 9835–9842. [Google Scholar] [CrossRef]
- Doig, P.; Kinsella, N.; Guerry, P.; Trust, T.J. Characterization of a post-translational modification of Campylobacter flagellin: Identification of a sero-specific glycosyl moiety. Mol. Microbiol. 1996, 19, 379–387. [Google Scholar] [CrossRef]
- Thibault, P.; Logan, S.M.; Kelly, J.F.; Brisson, J.R.; Ewing, C.P.; Trust, T.J.; Guerry, P. Identification of the carbohydrate moieties and glycosylation motifs in Campylobacter jejuni flagellin. J. Biol. Chem. 2001, 276, 34862–34870. [Google Scholar] [CrossRef] [Green Version]
- Logan, S.M.; Kelly, J.F.; Thibault, P.; Ewing, C.P.; Guerry, P. Structural heterogeneity of carbohydrate modifications affects serospecificity of Campylobacter flagellins. Mol. Microbiol. 2002, 46, 587–597. [Google Scholar] [CrossRef]
- Szymanski, C.M.; Logan, S.M.; Linton, D.; Wren, B.W. Campylobacter—A tale of two protein glycosylation systems. Trends Microbiol. 2003, 11, 233–238. [Google Scholar] [CrossRef]
- Goon, S.; Kelly, J.F.; Logan, S.M.; Ewing, C.P.; Guerry, P. Pseudaminic acid, the major modification on Campylobacter flagellin, is synthesized via the Cj1293 gene. Mol. Microbiol. 2003, 50, 659–671. [Google Scholar] [CrossRef] [PubMed]
- Schirm, M.; Soo, E.C.; Aubry, A.J.; Austin, J.; Thibault, P.; Logan, S.M. Structural, genetic and functional characterization of the flagellin glycosylation process in Helicobacter pylori. Mol. Microbiol. 2003, 48, 1579–1592. [Google Scholar] [CrossRef]
- Taguchi, F.; Shibata, S.; Suzuki, T.; Ogawa, Y.; Aizawa, S.; Takeuchi, K.; Ichinose, Y. Effects of glycosylation on swimming ability and flagellar polymorphic transformation in Pseudomonas syringae pv. tabaci 6605. J. Bacteriol. 2008, 190, 764–768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiku, K.; Yamamoto, M.; Ohnishi-Kameyama, M.; Ishii, T.; Yoshida, M.; Taguchi, F.; Ichinose, Y.; Ono, H. Comparative analysis of flagellin glycans among pathovars of phytopathogenic Pseudomonas syringae. Carbohydr. Res. 2013, 375, 100–104. [Google Scholar] [CrossRef] [PubMed]
- Ambler, R.P.; Rees, M.W. Epsilon-N-Methyl-lysine in bacterial flagellar protein. Nature 1959, 184, 56–57. [Google Scholar] [CrossRef]
- Frye, J.; Karlinsey, J.E.; Felise, H.R.; Marzolf, B.; Dowidar, N.; McClelland, M.; Hughes, K.T. Identification of new flagellar genes of Salmonella enterica serovar Typhimurium. J. Bacteriol. 2006, 188, 2233–2243. [Google Scholar] [CrossRef] [Green Version]
- Horstmann, J.A.; Lunelli, M.; Cazzola, H.; Heidemann, J.; Kuhne, C.; Steffen, P.; Szefs, S.; Rossi, C.; Lokareddy, R.K.; Wang, C.; et al. Methylation of Salmonella Typhimurium flagella promotes bacterial adhesion and host cell invasion. Nat. Commun. 2020, 11, 2013. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Jin, M.; Ding, W.; Yuan, J.; Kelly, J.; Gao, H. Posttranslational modification of flagellin FlaB in Shewanella oneidensis. J. Bacteriol. 2013, 195, 2550–2561. [Google Scholar] [CrossRef] [Green Version]
- Yonekura, K. Growth mechanism of the bacterial flagellar filament. Res. Microbiol. 2002, 153, 191–197. [Google Scholar] [CrossRef]
- Kamiya, R.; Asakura, S.; Yamaguchi, S. Formation of helical filaments by copolymerization of two types of ‘straight’ flagellins. Nature 1980, 286, 628–630. [Google Scholar] [CrossRef] [PubMed]
- Calladine, C.R.; Luisi, B.F.; Pratap, J.V. A “mechanistic” explanation of the multiple helical forms adopted by bacterial flagellar filaments. J. Mol. Biol. 2013, 425, 914–928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trachtenberg, S.; DeRosier, D.J. Three-dimensional structure of the frozen-hydrated flagellar filament. J. Mol. Biol. 1987, 195, 581–601. [Google Scholar] [CrossRef]
- Wang, F.; Burrage, A.M.; Postel, S.; Clark, R.E.; Orlova, A.; Sundberg, E.J.; Kearns, D.B.; Egelman, E.H. A structural model of flagellar filament switching across multiple bacterial species. Nat. Commun. 2017, 8, 960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mimori, Y.; Yamashita, I.; Murata, K.; Fujiyoshi, Y.; Yonekura, K.; Toyoshima, C.; Namba, K. The structure of the R-type straight flagellar filament of Salmonella at 9 A resolution by electron cryomicroscopy. J. Mol. Biol. 1995, 249, 69–87. [Google Scholar] [CrossRef] [PubMed]
- Morgan, D.G.; Owen, C.; Melanson, L.A.; DeRosier, D.J. Structure of bacterial flagellar filaments at 11 A resolution: Packing of the alpha-helices. J. Mol. Biol. 1995, 249, 88–110. [Google Scholar] [CrossRef]
- Mimori-Kiyosue, Y.; Yamashita, I.; Fujiyoshi, Y.; Yamaguchi, S.; Namba, K. Role of the outermost subdomain of Salmonella flagellin in the filament structure revealed by electron cryomicroscopy. J. Mol. Biol. 1998, 284, 521–530. [Google Scholar] [CrossRef] [PubMed]
- Maki-Yonekura, S.; Yonekura, K.; Namba, K. Conformational change of flagellin for polymorphic supercoiling of the flagellar filament. Nat. Struct. Mol. Biol. 2010, 17, 417–422. [Google Scholar] [CrossRef]
- Yamashita, I.; Hasegawa, K.; Suzuki, H.; Vonderviszt, F.; Mimori-Kiyosue, Y.; Namba, K. Structure and switching of bacterial flagellar filaments studied by X-ray fiber diffraction. Nat. Struct. Biol. 1998, 5, 125–132. [Google Scholar] [CrossRef]
- Kitao, A.; Yonekura, K.; Maki-Yonekura, S.; Samatey, F.A.; Imada, K.; Namba, K.; Go, N. Switch interactions control energy frustration and multiple flagellar filament structures. Proc. Natl. Acad. Sci. USA 2006, 103, 4894–4899. [Google Scholar] [CrossRef] [Green Version]
- Vonderviszt, F.; Imada, K.; Furukawa, Y.; Uedaira, H.; Taniguchi, H.; Namba, K. Mechanism of self-association and filament capping by flagellar HAP2. J. Mol. Biol. 1998, 284, 1399–1416. [Google Scholar] [CrossRef]
- Postel, S.; Deredge, D.; Bonsor, D.A.; Yu, X.; Diederichs, K.; Helmsing, S.; Vromen, A.; Friedler, A.; Hust, M.; Egelman, E.H.; et al. Bacterial flagellar capping proteins adopt diverse oligomeric states. Elife 2016, 5, e18857. [Google Scholar] [CrossRef]
- Song, W.S.; Cho, S.Y.; Hong, H.J.; Park, S.C.; Yoon, S.I. Self-Oligomerizing Structure of the Flagellar Cap Protein FliD and Its Implication in Filament Assembly. J. Mol. Biol. 2017, 429, 847–857. [Google Scholar] [CrossRef]
- Cho, S.Y.; Song, W.S.; Hong, H.J.; Lee, G.S.; Kang, S.G.; Ko, H.J.; Kim, P.H.; Yoon, S.I. Tetrameric structure of the flagellar cap protein FliD from Serratia marcescens. Biochem. Biophys. Res. Commun. 2017, 489, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.Y.; Song, W.S.; Oh, H.B.; Kim, H.U.; Jung, H.S.; Yoon, S.I. Structural analysis of the flagellar capping protein FliD from Helicobacter pylori. Biochem. Biophys. Res. Commun. 2019, 514, 98–104. [Google Scholar] [CrossRef] [PubMed]
- Al-Otaibi, N.S.; Taylor, A.J.; Farrell, D.P.; Tzokov, S.B.; DiMaio, F.; Kelly, D.J.; Bergeron, J.R.C. The cryo-EM structure of the bacterial flagellum cap complex suggests a molecular mechanism for filament elongation. Nat. Commun. 2020, 11, 3210. [Google Scholar] [CrossRef]
- Chevance, F.F.; Hughes, K.T. Coordinating assembly of a bacterial macromolecular machine. Nat. Rev. Microbiol. 2008, 6, 455–465. [Google Scholar] [CrossRef] [Green Version]
- Wilhelms, M.; Molero, R.; Shaw, J.G.; Tomas, J.M.; Merino, S. Transcriptional hierarchy of Aeromonas hydrophila polar-flagellum genes. J. Bacteriol. 2011, 193, 5179–5190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dasgupta, N.; Wolfgang, M.C.; Goodman, A.L.; Arora, S.K.; Jyot, J.; Lory, S.; Ramphal, R. A four-tiered transcriptional regulatory circuit controls flagellar biogenesis in Pseudomonas aeruginosa. Mol. Microbiol. 2003, 50, 809–824. [Google Scholar] [CrossRef] [Green Version]
- McCarter, L.L. Regulation of flagella. Curr. Opin. Microbiol. 2006, 9, 180–186. [Google Scholar] [CrossRef]
- Jang, M.S.; Mouri, Y.; Uchida, K.; Aizawa, S.; Hayakawa, M.; Fujita, N.; Tezuka, T.; Ohnishi, Y. Genetic and Transcriptional Analyses of the Flagellar Gene Cluster in Actinoplanes missouriensis. J. Bacteriol. 2016, 198, 2219–2227. [Google Scholar] [CrossRef] [Green Version]
- Ikeda, T.; Oosawa, K.; Hotani, H. Self-assembly of the filament capping protein, FliD, of bacterial flagella into an annular structure. J. Mol. Biol. 1996, 259, 679–686. [Google Scholar] [CrossRef] [PubMed]
- Aldridge, P.; Gnerer, J.; Karlinsey, J.E.; Hughes, K.T. Transcriptional and translational control of the Salmonella fliC gene. J. Bacteriol. 2006, 188, 4487–4496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koirala, S.; Mears, P.; Sim, M.; Golding, I.; Chemla, Y.R.; Aldridge, P.D.; Rao, C.V. A nutrient-tunable bistable switch controls motility in Salmonella enterica serovar Typhimurium. MBio 2014, 5, e01611–e01614. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Koirala, S.; Aldridge, P.D.; Rao, C.V. Two Tandem Mechanisms Control Bimodal Expression of the Flagellar Genes in Salmonella enterica. J. Bacteriol. 2020, 202, e00787-19. [Google Scholar] [CrossRef] [PubMed]
- Niehus, E.; Gressmann, H.; Ye, F.; Schlapbach, R.; Dehio, M.; Dehio, C.; Stack, A.; Meyer, T.F.; Suerbaum, S.; Josenhans, C. Genome-wide analysis of transcriptional hierarchy and feedback regulation in the flagellar system of Helicobacter pylori. Mol. Microbiol. 2004, 52, 947–961. [Google Scholar] [CrossRef]
- Alm, R.A.; Guerry, P.; Trust, T.J. Significance of duplicated flagellin genes in Campylobacter. J. Mol. Biol. 1993, 230, 359–363. [Google Scholar] [CrossRef] [Green Version]
- Prouty, M.G.; Correa, N.E.; Klose, K.E. The novel sigma54- and sigma28-dependent flagellar gene transcription hierarchy of Vibrio cholerae. Mol. Microbiol. 2001, 39, 1595–1609. [Google Scholar] [CrossRef]
- Neville, B.A.; Sheridan, P.O.; Harris, H.M.; Coughlan, S.; Flint, H.J.; Duncan, S.H.; Jeffery, I.B.; Claesson, M.J.; Ross, R.P.; Scott, K.P.; et al. Pro-inflammatory flagellin proteins of prevalent motile commensal bacteria are variably abundant in the intestinal microbiome of elderly humans. PLoS ONE 2013, 8, e68919. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Chai, Y.; Guo, J.H.; Losick, R. Evidence for cyclic Di-GMP-mediated signaling in Bacillus subtilis. J. Bacteriol. 2012, 194, 5080–5090. [Google Scholar] [CrossRef] [PubMed]
- Purcell, E.B.; McKee, R.W.; McBride, S.M.; Waters, C.M.; Tamayo, R. Cyclic diguanylate inversely regulates motility and aggregation in Clostridium difficile. J. Bacteriol. 2012, 194, 3307–3316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simm, R.; Morr, M.; Kader, A.; Nimtz, M.; Romling, U. GGDEF and EAL domains inversely regulate cyclic di-GMP levels and transition from sessility to motility. Mol. Microbiol. 2004, 53, 1123–1134. [Google Scholar] [CrossRef] [PubMed]
- Kuchma, S.L.; Brothers, K.M.; Merritt, J.H.; Liberati, N.T.; Ausubel, F.M.; O’Toole, G.A. BifA, a cyclic-Di-GMP phosphodiesterase, inversely regulates biofilm formation and swarming motility by Pseudomonas aeruginosa PA14. J. Bacteriol. 2007, 189, 8165–8178. [Google Scholar] [CrossRef] [Green Version]
- Paul, K.; Nieto, V.; Carlquist, W.C.; Blair, D.F.; Harshey, R.M. The c-di-GMP binding protein YcgR controls flagellar motor direction and speed to affect chemotaxis by a “backstop brake” mechanism. Mol. Cell 2010, 38, 128–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, Y.; Yu, Z.; Liu, S.; Chen, B.; Zhu, L.; Li, Z.; Chou, S.H.; He, J. c-di-GMP Regulates Various Phenotypes and Insecticidal Activity of Gram-Positive Bacillus thuringiensis. Front. Microbiol. 2018, 9, 45. [Google Scholar] [CrossRef] [PubMed]
- Verberkmoes, N.C.; Russell, A.L.; Shah, M.; Godzik, A.; Rosenquist, M.; Halfvarson, J.; Lefsrud, M.G.; Apajalahti, J.; Tysk, C.; Hettich, R.L.; et al. Shotgun metaproteomics of the human distal gut microbiota. ISME J. 2009, 3, 179–189. [Google Scholar] [CrossRef] [Green Version]
- Cullender, T.C.; Chassaing, B.; Janzon, A.; Kumar, K.; Muller, C.E.; Werner, J.J.; Angenent, L.T.; Bell, M.E.; Hay, A.G.; Peterson, D.A.; et al. Innate and adaptive immunity interact to quench microbiome flagellar motility in the gut. Cell Host Microbe 2013, 14, 571–581. [Google Scholar] [CrossRef] [Green Version]
- Gauger, E.J.; Leatham, M.P.; Mercado-Lubo, R.; Laux, D.C.; Conway, T.; Cohen, P.S. Role of motility and the flhDC Operon in Escherichia coli MG1655 colonization of the mouse intestine. Infect. Immun. 2007, 75, 3315–3324. [Google Scholar] [CrossRef] [Green Version]
- Eriksson, S.; Lucchini, S.; Thompson, A.; Rhen, M.; Hinton, J.C. Unravelling the biology of macrophage infection by gene expression profiling of intracellular Salmonella enterica. Mol. Microbiol. 2003, 47, 103–118. [Google Scholar] [CrossRef] [PubMed]
- Ott, M.; Messner, P.; Heesemann, J.; Marre, R.; Hacker, J. Temperature-dependent expression of flagella in Legionella. J. Gen. Microbiol. 1991, 137, 1955–1961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamp, H.D.; Higgins, D.E. A protein thermometer controls temperature-dependent transcription of flagellar motility genes in Listeria monocytogenes. PLoS Pathog. 2011, 7, e1002153. [Google Scholar] [CrossRef] [PubMed]
- Anderson, P.E.; Gober, J.W. FlbT, the post-transcriptional regulator of flagellin synthesis in Caulobacter crescentus, interacts with the 5′ untranslated region of flagellin mRNA. Mol. Microbiol. 2000, 38, 41–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, S.; Kutsukake, K. FljA-mediated posttranscriptional control of phase 1 flagellin expression in flagellar phase variation of Salmonella enterica serovar Typhimurium. J. Bacteriol. 2006, 188, 958–967. [Google Scholar] [CrossRef] [Green Version]
- Yakhnin, H.; Pandit, P.; Petty, T.J.; Baker, C.S.; Romeo, T.; Babitzke, P. CsrA of Bacillus subtilis regulates translation initiation of the gene encoding the flagellin protein (hag) by blocking ribosome binding. Mol. Microbiol. 2007, 64, 1605–1620. [Google Scholar] [CrossRef]
- Anderson, D.K.; Newton, A. Posttranscriptional regulation of Caulobacter flagellin genes by a late flagellum assembly checkpoint. J. Bacteriol. 1997, 179, 2281–2288. [Google Scholar] [CrossRef] [Green Version]
- Sal, M.S.; Li, C.; Motalab, M.A.; Shibata, S.; Aizawa, S.; Charon, N.W. Borrelia burgdorferi uniquely regulates its motility genes and has an intricate flagellar hook-basal body structure. J. Bacteriol. 2008, 190, 1912–1921. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, S.; Yakhnin, H.; Kysela, D.; Sokoloski, J.; Babitzke, P.; Kearns, D.B. CsrA-FliW interaction governs flagellin homeostasis and a checkpoint on flagellar morphogenesis in Bacillus subtilis. Mol. Microbiol. 2011, 82, 447–461. [Google Scholar] [CrossRef] [Green Version]
- Khanra, N.; Rossi, P.; Economou, A.; Kalodimos, C.G. Recognition and targeting mechanisms by chaperones in flagellum assembly and operation. Proc. Natl. Acad. Sci. USA 2016, 113, 9798–9803. [Google Scholar] [CrossRef] [Green Version]
- Auvray, F.; Thomas, J.; Fraser, G.M.; Hughes, C. Flagellin polymerisation control by a cytosolic export chaperone. J. Mol. Biol. 2001, 308, 221–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozin, A.J.; Claret, L.; Auvray, F.; Hughes, C. The FliS chaperone selectively binds the disordered flagellin C-terminal D0 domain central to polymerisation. FEMS Microbiol. Lett. 2003, 219, 219–224. [Google Scholar] [CrossRef]
- Muskotal, A.; Kiraly, R.; Sebestyen, A.; Gugolya, Z.; Vegh, B.M.; Vonderviszt, F. Interaction of FliS flagellar chaperone with flagellin. FEBS Lett. 2006, 580, 3916–3920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evdokimov, A.G.; Phan, J.; Tropea, J.E.; Routzahn, K.M.; Peters, H.K.; Pokross, M.; Waugh, D.S. Similar modes of polypeptide recognition by export chaperones in flagellar biosynthesis and type III secretion. Nat. Struct. Biol. 2003, 10, 789–793. [Google Scholar] [CrossRef] [PubMed]
- Galeva, A.; Moroz, N.; Yoon, Y.H.; Hughes, K.T.; Samatey, F.A.; Kostyukova, A.S. Bacterial flagellin-specific chaperone FliS interacts with anti-sigma factor FlgM. J. Bacteriol. 2014, 196, 1215–1221. [Google Scholar] [CrossRef] [Green Version]
- Furukawa, Y.; Inoue, Y.; Sakaguchi, A.; Mori, Y.; Fukumura, T.; Miyata, T.; Namba, K.; Minamino, T. Structural stability of flagellin subunit affects the rate of flagellin export in the absence of FliS chaperone. Mol. Microbiol. 2016, 102, 405–416. [Google Scholar] [CrossRef]
- Bange, G.; Kummerer, N.; Engel, C.; Bozkurt, G.; Wild, K.; Sinning, I. FlhA provides the adaptor for coordinated delivery of late flagella building blocks to the type III secretion system. Proc. Natl. Acad. Sci. USA 2010, 107, 11295–11300. [Google Scholar] [CrossRef] [Green Version]
- Kinoshita, M.; Hara, N.; Imada, K.; Namba, K.; Minamino, T. Interactions of bacterial flagellar chaperone-substrate complexes with FlhA contribute to co-ordinating assembly of the flagellar filament. Mol. Microbiol. 2013, 90, 1249–1261. [Google Scholar] [CrossRef] [Green Version]
- Terahara, N.; Inoue, Y.; Kodera, N.; Morimoto, Y.V.; Uchihashi, T.; Imada, K.; Ando, T.; Namba, K.; Minamino, T. Insight into structural remodeling of the FlhA ring responsible for bacterial flagellar type III protein export. Sci. Adv. 2018, 4, eaao7054. [Google Scholar] [CrossRef] [Green Version]
- Xing, Q.; Shi, K.; Portaliou, A.; Rossi, P.; Economou, A.; Kalodimos, C.G. Structures of chaperone-substrate complexes docked onto the export gate in a type III secretion system. Nat. Commun. 2018, 9, 1773. [Google Scholar] [CrossRef]
- Altegoer, F.; Mukherjee, S.; Steinchen, W.; Bedrunka, P.; Linne, U.; Kearns, D.B.; Bange, G. FliS/flagellin/FliW heterotrimer couples type III secretion and flagellin homeostasis. Sci. Rep. 2018, 8, 11552. [Google Scholar] [CrossRef]
- Yokoseki, T.; Iino, T.; Kutsukake, K. Negative regulation by fliD, fliS, and fliT of the export of the flagellum-specific anti-sigma factor, FlgM, in Salmonella typhimurium. J. Bacteriol. 1996, 178, 899–901. [Google Scholar] [CrossRef] [Green Version]
- Yokoseki, T.; Kutsukake, K.; Ohnishi, K.; Iino, T. Functional analysis of the flagellar genes in the fliD operon of Salmonella typhimurium. Microbiology (Reading) 1995, 141, 1715–1722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, S.; Peng, Z.; Cui, B.; Wang, T.; Song, Y.; Zhang, L.; Wei, G.; Wang, Y.; Shen, X. FliS modulates FlgM activity by acting as a non-canonical chaperone to control late flagellar gene expression, motility and biofilm formation in Yersinia pseudotuberculosis. Environ. Microbiol. 2014, 16, 1090–1104. [Google Scholar] [CrossRef] [PubMed]
- Imada, K.; Minamino, T.; Kinoshita, M.; Furukawa, Y.; Namba, K. Structural insight into the regulatory mechanisms of interactions of the flagellar type III chaperone FliT with its binding partners. Proc. Natl. Acad. Sci. USA 2010, 107, 8812–8817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibuki, T.; Imada, K.; Minamino, T.; Kato, T.; Miyata, T.; Namba, K. Common architecture of the flagellar type III protein export apparatus and F- and V-type ATPases. Nat. Struct. Mol. Biol. 2011, 18, 277–282. [Google Scholar] [CrossRef]
- Erhardt, M.; Mertens, M.E.; Fabiani, F.D.; Hughes, K.T. ATPase-independent type-III protein secretion in Salmonella enterica. PLoS Genet. 2014, 10, e1004800. [Google Scholar] [CrossRef]
- Minamino, T.; Kawamoto, A.; Kinoshita, M.; Namba, K. Molecular Organization and Assembly of the Export Apparatus of Flagellar Type III Secretion Systems. Curr. Top. Microbiol. Immunol. 2020, 427, 91–107. [Google Scholar] [CrossRef]
- Hughes, K.T.; Gillen, K.L.; Semon, M.J.; Karlinsey, J.E. Sensing structural intermediates in bacterial flagellar assembly by export of a negative regulator. Science 1993, 262, 1277–1280. [Google Scholar] [CrossRef]
- Calvo, R.A.; Kearns, D.B. FlgM is secreted by the flagellar export apparatus in Bacillus subtilis. J. Bacteriol. 2015, 197, 81–91. [Google Scholar] [CrossRef] [Green Version]
- Evans, L.D.; Hughes, C.; Fraser, G.M. Building a flagellum in biological outer space. Microb. Cell 2014, 1, 64–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, L.D.; Poulter, S.; Terentjev, E.M.; Hughes, C.; Fraser, G.M. A chain mechanism for flagellum growth. Nature 2013, 504, 287–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, L.; Stern, A.S.; Berg, H.C. Growth of flagellar filaments of Escherichia coli is independent of filament length. J. Bacteriol. 2012, 194, 2437–2442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stern, A.S.; Berg, H.C. Single-file diffusion of flagellin in flagellar filaments. Biophys. J. 2013, 105, 182–184. [Google Scholar] [CrossRef] [Green Version]
- Tanner, D.E.; Ma, W.; Chen, Z.; Schulten, K. Theoretical and computational investigation of flagellin translocation and bacterial flagellum growth. Biophys. J. 2011, 100, 2548–2556. [Google Scholar] [CrossRef] [Green Version]
- Renault, T.T.; Abraham, A.O.; Bergmiller, T.; Paradis, G.; Rainville, S.; Charpentier, E.; Guet, C.C.; Tu, Y.; Namba, K.; Keener, J.P.; et al. Bacterial flagella grow through an injection-diffusion mechanism. Elife 2017, 6, e23136. [Google Scholar] [CrossRef]
- Shibata, S.; Takahashi, N.; Chevance, F.F.; Karlinsey, J.E.; Hughes, K.T.; Aizawa, S. FliK regulates flagellar hook length as an internal ruler. Mol. Microbiol. 2007, 64, 1404–1415. [Google Scholar] [CrossRef]
- Burtchett, T.; Love, C.; Sarkar, R.; Tripp, B.C. A structure-function study of C-terminal residues predicted to line the export channel in Salmonella Flagellin. Biochim. Biophys. Acta Gen. Subj. 2021, 1865, 129748. [Google Scholar] [CrossRef]
- Haiko, J.; Westerlund-Wikstrom, B. The role of the bacterial flagellum in adhesion and virulence. Biology (Basel) 2013, 2, 1242–1267. [Google Scholar] [CrossRef] [Green Version]
- Duan, Q.; Zhou, M.; Zhu, L.; Zhu, G. Flagella and bacterial pathogenicity. J. Basic. Microbiol. 2013, 53, 1–8. [Google Scholar] [CrossRef]
- McSweegan, E.; Walker, R.I. Identification and characterization of two Campylobacter jejuni adhesins for cellular and mucous substrates. Infect. Immun. 1986, 53, 141–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirov, S.M.; Castrisios, M.; Shaw, J.G. Aeromonas flagella (polar and lateral) are enterocyte adhesins that contribute to biofilm formation on surfaces. Infect. Immun. 2004, 72, 1939–1945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arora, S.K.; Ritchings, B.W.; Almira, E.C.; Lory, S.; Ramphal, R. The Pseudomonas aeruginosa flagellar cap protein, FliD, is responsible for mucin adhesion. Infect. Immun. 1998, 66, 1000–1007. [Google Scholar] [CrossRef] [Green Version]
- Simpson, D.A.; Ramphal, R.; Lory, S. Genetic analysis of Pseudomonas aeruginosa adherence: Distinct genetic loci control attachment to epithelial cells and mucins. Infect. Immun. 1992, 60, 3771–3779. [Google Scholar] [CrossRef] [Green Version]
- Scharfman, A.; Arora, S.K.; Delmotte, P.; Van Brussel, E.; Mazurier, J.; Ramphal, R.; Roussel, P. Recognition of Lewis x derivatives present on mucins by flagellar components of Pseudomonas aeruginosa. Infect. Immun. 2001, 69, 5243–5248. [Google Scholar] [CrossRef] [Green Version]
- Lillehoj, E.P.; Kim, B.T.; Kim, K.C. Identification of Pseudomonas aeruginosa flagellin as an adhesin for Muc1 mucin. Am. J. Physiol. Lung Cell. Mol. Physiol. 2002, 282, L751–L756. [Google Scholar] [CrossRef] [Green Version]
- Giron, J.A.; Torres, A.G.; Freer, E.; Kaper, J.B. The flagella of enteropathogenic Escherichia coli mediate adherence to epithelial cells. Mol. Microbiol. 2002, 44, 361–379. [Google Scholar] [CrossRef]
- Roy, K.; Hilliard, G.M.; Hamilton, D.J.; Luo, J.; Ostmann, M.M.; Fleckenstein, J.M. Enterotoxigenic Escherichia coli EtpA mediates adhesion between flagella and host cells. Nature 2009, 457, 594–598. [Google Scholar] [CrossRef] [PubMed]
- Erdem, A.L.; Avelino, F.; Xicohtencatl-Cortes, J.; Giron, J.A. Host protein binding and adhesive properties of H6 and H7 flagella of attaching and effacing Escherichia coli. J. Bacteriol. 2007, 189, 7426–7435. [Google Scholar] [CrossRef] [Green Version]
- Mahajan, A.; Currie, C.G.; Mackie, S.; Tree, J.; McAteer, S.; McKendrick, I.; McNeilly, T.N.; Roe, A.; La Ragione, R.M.; Woodward, M.J.; et al. An investigation of the expression and adhesin function of H7 flagella in the interaction of Escherichia coli O157: H7 with bovine intestinal epithelium. Cell Microbiol. 2009, 11, 121–137. [Google Scholar] [CrossRef] [Green Version]
- Troge, A.; Scheppach, W.; Schroeder, B.O.; Rund, S.A.; Heuner, K.; Wehkamp, J.; Stange, E.F.; Oelschlaeger, T.A. More than a marine propeller—The flagellum of the probiotic Escherichia coli strain Nissle 1917 is the major adhesin mediating binding to human mucus. Int. J. Med. Microbiol. 2012, 302, 304–314. [Google Scholar] [CrossRef] [PubMed]
- Tasteyre, A.; Barc, M.C.; Collignon, A.; Boureau, H.; Karjalainen, T. Role of FliC and FliD flagellar proteins of Clostridium difficile in adherence and gut colonization. Infect. Immun. 2001, 69, 7937–7940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dingle, T.C.; Mulvey, G.L.; Armstrong, G.D. Mutagenic analysis of the Clostridium difficile flagellar proteins, FliC and FliD, and their contribution to virulence in hamsters. Infect. Immun. 2011, 79, 4061–4067. [Google Scholar] [CrossRef] [Green Version]
- Sheu, B.S.; Yang, H.B.; Yeh, Y.C.; Wu, J.J. Helicobacter pylori colonization of the human gastric epithelium: A bug’s first step is a novel target for us. J. Gastroenterol. Hepatol. 2010, 25, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Cazzola, H.; Lemaire, L.; Acket, S.; Prost, E.; Duma, L.; Erhardt, M.; Cechova, P.; Trouillas, P.; Mohareb, F.; Rossi, C.; et al. The Impact of Plasma Membrane Lipid Composition on Flagellum-Mediated Adhesion of Enterohemorrhagic Escherichia coli. MSphere 2020, 5, e00702-20. [Google Scholar] [CrossRef] [PubMed]
- Crawford, R.W.; Reeve, K.E.; Gunn, J.S. Flagellated but not hyperfimbriated Salmonella enterica serovar Typhimurium attaches to and forms biofilms on cholesterol-coated surfaces. J. Bacteriol. 2010, 192, 2981–2990. [Google Scholar] [CrossRef] [Green Version]
- Guttenplan, S.B.; Kearns, D.B. Regulation of flagellar motility during biofilm formation. FEMS Microbiol. Rev. 2013, 37, 849–871. [Google Scholar] [CrossRef] [Green Version]
- Kim, T.J.; Young, B.M.; Young, G.M. Effect of flagellar mutations on Yersinia enterocolitica biofilm formation. Appl. Environ. Microbiol. 2008, 74, 5466–5474. [Google Scholar] [CrossRef] [Green Version]
- Zhou, M.; Guo, Z.; Yang, Y.; Duan, Q.; Zhang, Q.; Yao, F.; Zhu, J.; Zhang, X.; Hardwidge, P.R.; Zhu, G. Flagellin and F4 fimbriae have opposite effects on biofilm formation and quorum sensing in F4ac+ enterotoxigenic Escherichia coli. Vet. Microbiol. 2014, 168, 148–153. [Google Scholar] [CrossRef]
- Hossain, M.M.; Tsuyumu, S. Flagella-mediated motility is required for biofilm formation by Erwinia carotovora subsp. carotovora. J. Gen. Plant Pathol. 2006, 72, 34–39. [Google Scholar] [CrossRef]
- He, Y.; Xu, T.; Fossheim, L.E.; Zhang, X.H. FliC, a flagellin protein, is essential for the growth and virulence of fish pathogen Edwardsiella tarda. PLoS ONE 2012, 7, e45070. [Google Scholar] [CrossRef] [Green Version]
- Hartmann, I.; Carranza, P.; Lehner, A.; Stephan, R.; Eberl, L.; Riedel, K. Genes involved in Cronobacter sakazakii biofilm formation. Appl. Environ. Microbiol. 2010, 76, 2251–2261. [Google Scholar] [CrossRef] [Green Version]
- Suriyanarayanan, T.; Periasamy, S.; Lin, M.H.; Ishihama, Y.; Swarup, S. Flagellin FliC Phosphorylation Affects Type 2 Protease Secretion and Biofilm Dispersal in Pseudomonas aeruginosa PAO1. PLoS ONE 2016, 11, e0164155. [Google Scholar] [CrossRef]
- Jung, Y.C.; Lee, M.A.; Lee, K.H. Role of Flagellin-Homologous Proteins in Biofilm Formation by Pathogenic Vibrio Species. MBio 2019, 10, e01793-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, G.M.; Schmiel, D.H.; Miller, V.L. A new pathway for the secretion of virulence factors by bacteria: The flagellar export apparatus functions as a protein-secretion system. Proc. Natl. Acad. Sci. USA 1999, 96, 6456–6461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmiel, D.H.; Wagar, E.; Karamanou, L.; Weeks, D.; Miller, V.L. Phospholipase A of Yersinia enterocolitica contributes to pathogenesis in a mouse model. Infect. Immun. 1998, 66, 3941–3951. [Google Scholar] [CrossRef] [Green Version]
- Warren, S.M.; Young, G.M. An amino-terminal secretion signal is required for YplA export by the Ysa, Ysc, and flagellar type III secretion systems of Yersinia enterocolitica biovar 1B. J. Bacteriol. 2005, 187, 6075–6083. [Google Scholar] [CrossRef] [Green Version]
- Young, B.M.; Young, G.M. YplA is exported by the Ysc, Ysa, and flagellar type III secretion systems of Yersinia enterocolitica. J. Bacteriol. 2002, 184, 1324–1334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buelow, D.R.; Christensen, J.E.; Neal-McKinney, J.M.; Konkel, M.E. Campylobacter jejuni survival within human epithelial cells is enhanced by the secreted protein CiaI. Mol. Microbiol. 2011, 80, 1296–1312. [Google Scholar] [CrossRef] [Green Version]
- Christensen, J.E.; Pacheco, S.A.; Konkel, M.E. Identification of a Campylobacter jejuni-secreted protein required for maximal invasion of host cells. Mol. Microbiol. 2009, 73, 650–662. [Google Scholar] [CrossRef] [Green Version]
- Konkel, M.E.; Kim, B.J.; Rivera-Amill, V.; Garvis, S.G. Bacterial secreted proteins are required for the internaliztion of Campylobacter jejuni into cultured mammalian cells. Mol. Microbiol. 1999, 32, 691–701. [Google Scholar] [CrossRef]
- Konkel, M.E.; Klena, J.D.; Rivera-Amill, V.; Monteville, M.R.; Biswas, D.; Raphael, B.; Mickelson, J. Secretion of virulence proteins from Campylobacter jejuni is dependent on a functional flagellar export apparatus. J. Bacteriol. 2004, 186, 3296–3303. [Google Scholar] [CrossRef] [Green Version]
- Grant, C.C.; Konkel, M.E.; Cieplak, W., Jr.; Tompkins, L.S. Role of flagella in adherence, internalization, and translocation of Campylobacter jejuni in nonpolarized and polarized epithelial cell cultures. Infect. Immun. 1993, 61, 1764–1771. [Google Scholar] [CrossRef] [Green Version]
- Neal-McKinney, J.M.; Konkel, M.E. The Campylobacter jejuni CiaC virulence protein is secreted from the flagellum and delivered to the cytosol of host cells. Front. Cell Infect. Microbiol. 2012, 2, 31. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.C.; Jin, S.; Louie, H.; Ng, D.; Lau, R.; Zhang, Y.; Weerasekera, R.; Al Rashid, S.; Ward, L.A.; Der, S.D.; et al. FlaC, a protein of Campylobacter jejuni TGH9011 (ATCC43431) secreted through the flagellar apparatus, binds epithelial cells and influences cell invasion. Mol. Microbiol. 2004, 53, 541–553. [Google Scholar] [CrossRef]
- Samuelson, D.R.; Eucker, T.P.; Bell, J.A.; Dybas, L.; Mansfield, L.S.; Konkel, M.E. The Campylobacter jejuni CiaD effector protein activates MAP kinase signaling pathways and is required for the development of disease. Cell Commun. Signal. 2013, 11, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samuelson, D.R.; Konkel, M.E. Serine phosphorylation of cortactin is required for maximal host cell invasion by Campylobacter jejuni. Cell Commun. Signal. 2013, 11, 82. [Google Scholar] [CrossRef] [Green Version]
- Barrero-Tobon, A.M.; Hendrixson, D.R. Identification and analysis of flagellar coexpressed determinants (Feds) of Campylobacter jejuni involved in colonization. Mol. Microbiol. 2012, 84, 352–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuwajima, G.; Kawagishi, I.; Homma, M.; Asaka, J.; Kondo, E.; Macnab, R.M. Export of an N-terminal fragment of Escherichia coli flagellin by a flagellum-specific pathway. Proc. Natl. Acad. Sci. USA 1989, 86, 4953–4957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vegh, B.M.; Gal, P.; Dobo, J.; Zavodszky, P.; Vonderviszt, F. Localization of the flagellum-specific secretion signal in Salmonella flagellin. Biochem. Biophys. Res. Commun. 2006, 345, 93–98. [Google Scholar] [CrossRef]
- Smith, K.D.; Andersen-Nissen, E.; Hayashi, F.; Strobe, K.; Bergman, M.A.; Barrett, S.L.; Cookson, B.T.; Aderem, A. Toll-like receptor 5 recognizes a conserved site on flagellin required for protofilament formation and bacterial motility. Nat. Immunol. 2003, 4, 1247–1253. [Google Scholar] [CrossRef]
- Zhao, Y.; Yang, J.; Shi, J.; Gong, Y.N.; Lu, Q.; Xu, H.; Liu, L.; Shao, F. The NLRC4 inflammasome receptors for bacterial flagellin and type III secretion apparatus. Nature 2011, 477, 596–600. [Google Scholar] [CrossRef]
- Lopez-Yglesias, A.H.; Lu, C.C.; Zhao, X.; Chou, T.; VandenBos, T.; Strong, R.K.; Smith, K.D. FliC’s Hypervariable D3 Domain Is Required for Robust Anti-Flagellin Primary Antibody Responses. Immunohorizons 2019, 3, 422–432. [Google Scholar] [CrossRef]
- Andersen-Nissen, E.; Smith, K.D.; Bonneau, R.; Strong, R.K.; Aderem, A. A conserved surface on Toll-like receptor 5 recognizes bacterial flagellin. J. Exp. Med. 2007, 204, 393–403. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, F.; Smith, K.D.; Ozinsky, A.; Hawn, T.R.; Yi, E.C.; Goodlett, D.R.; Eng, J.K.; Akira, S.; Underhill, D.M.; Aderem, A. The innate immune response to bacterial flagellin is mediated by Toll-like receptor 5. Nature 2001, 410, 1099–1103. [Google Scholar] [CrossRef] [PubMed]
- Gay, N.J.; Gangloff, M.; Weber, A.N. Toll-like receptors as molecular switches. Nat. Rev. Immunol. 2006, 6, 693–698. [Google Scholar] [CrossRef] [PubMed]
- Hajam, I.A.; Dar, P.A.; Shahnawaz, I.; Jaume, J.C.; Lee, J.H. Bacterial flagellin-a potent immunomodulatory agent. Exp. Mol. Med. 2017, 49, e373. [Google Scholar] [CrossRef] [PubMed]
- Gewirtz, A.T.; Navas, T.A.; Lyons, S.; Godowski, P.J.; Madara, J.L. Cutting edge: Bacterial flagellin activates basolaterally expressed TLR5 to induce epithelial proinflammatory gene expression. J. Immunol. 2001, 167, 1882–1885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Araki, A.; Kanai, T.; Ishikura, T.; Makita, S.; Uraushihara, K.; Iiyama, R.; Totsuka, T.; Takeda, K.; Akira, S.; Watanabe, M. MyD88-deficient mice develop severe intestinal inflammation in dextran sodium sulfate colitis. J. Gastroenterol. 2005, 40, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Xie, Y.; Tan, M.; Zheng, K.; Xiao, Y.; Jiang, C.; Zhao, F.; Zeng, T.; Wu, Y. The N-terminal D1 domain of Treponema pallidum flagellin binding to TLR5 is required but not sufficient in activation of TLR5. J. Cell Mol. Med. 2019, 23, 7490–7504. [Google Scholar] [CrossRef]
- Andersen-Nissen, E.; Smith, K.D.; Strobe, K.L.; Barrett, S.L.; Cookson, B.T.; Logan, S.M.; Aderem, A. Evasion of Toll-like receptor 5 by flagellated bacteria. Proc. Natl. Acad. Sci. USA 2005, 102, 9247–9252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voogdt, C.G.P.; Wagenaar, J.A.; van Putten, J.P.M. Duplicated TLR5 of zebrafish functions as a heterodimeric receptor. Proc. Natl. Acad. Sci. USA 2018, 115, E3221–E3229. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Gómez, L.; Boller, T. FLS2: An LRR Receptor–like Kinase Involved in the Perception of the Bacterial Elicitor Flagellin in Arabidopsis. Mol. Cell 2000, 5, 1003–1011. [Google Scholar] [CrossRef]
- Robatzek, S.; Wirthmueller, L. Mapping FLS2 function to structure: LRRs, kinase and its working bits. Protoplasma 2013, 250, 671–681. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Li, L.; Macho, A.P.; Han, Z.; Hu, Z.; Zipfel, C.; Zhou, J.M.; Chai, J. Structural basis for flg22-induced activation of the Arabidopsis FLS2-BAK1 immune complex. Science 2013, 342, 624–628. [Google Scholar] [CrossRef]
- He, K.; Wu, Y. Receptor-Like Kinases and Regulation of Plant Innate Immunity. Enzymes 2016, 40, 105–142. [Google Scholar] [CrossRef] [PubMed]
- Tenthorey, J.L.; Haloupek, N.; Lopez-Blanco, J.R.; Grob, P.; Adamson, E.; Hartenian, E.; Lind, N.A.; Bourgeois, N.M.; Chacon, P.; Nogales, E.; et al. The structural basis of flagellin detection by NAIP5: A strategy to limit pathogen immune evasion. Science 2017, 358, 888–893. [Google Scholar] [CrossRef] [Green Version]
- Nempont, C.; Cayet, D.; Rumbo, M.; Bompard, C.; Villeret, V.; Sirard, J.C. Deletion of flagellin’s hypervariable region abrogates antibody-mediated neutralization and systemic activation of TLR5-dependent immunity. J. Immunol. 2008, 181, 2036–2043. [Google Scholar] [CrossRef] [Green Version]
- Cui, B.; Liu, X.; Fang, Y.; Zhou, P.; Zhang, Y.; Wang, Y. Flagellin as a vaccine adjuvant. Expert Rev. Vaccines 2018, 17, 335–349. [Google Scholar] [CrossRef]
- Honko, A.N.; Sriranganathan, N.; Lees, C.J.; Mizel, S.B. Flagellin is an effective adjuvant for immunization against lethal respiratory challenge with Yersinia pestis. Infect. Immun. 2006, 74, 1113–1120. [Google Scholar] [CrossRef] [Green Version]
- Gupta, S.K.; Bajwa, P.; Deb, R.; Chellappa, M.M.; Dey, S. Flagellin a toll-like receptor 5 agonist as an adjuvant in chicken vaccines. Clin. Vaccine Immunol. 2014, 21, 261–270. [Google Scholar] [CrossRef] [Green Version]
- Xiao, X.; Zhang, Y.; Wei, Q.; Yin, X. Flagellin FljB as an adjuvant to the recombinant adenovirus rabies glycoprotein vaccine increases immune responses against rabies in mice. Arch. Virol. 2017, 162, 2655–2665. [Google Scholar] [CrossRef]
- Cunningham, A.F.; Khan, M.; Ball, J.; Toellner, K.M.; Serre, K.; Mohr, E.; MacLennan, I.C. Responses to the soluble flagellar protein FliC are Th2, while those to FliC on Salmonella are Th1. Eur. J. Immunol. 2004, 34, 2986–2995. [Google Scholar] [CrossRef] [PubMed]
- Georgel, A.F.; Cayet, D.; Pizzorno, A.; Rosa-Calatrava, M.; Paget, C.; Sencio, V.; Dubuisson, J.; Trottein, F.; Sirard, J.C.; Carnoy, C. Toll-like receptor 5 agonist flagellin reduces influenza A virus replication independently of type I interferon and interleukin 22 and improves antiviral efficacy of oseltamivir. Antivir. Res. 2019, 168, 28–35. [Google Scholar] [CrossRef] [Green Version]
- Song, W.S.; Jeon, Y.J.; Namgung, B.; Hong, M.; Yoon, S.I. A conserved TLR5 binding and activation hot spot on flagellin. Sci. Rep. 2017, 7, 40878. [Google Scholar] [CrossRef] [Green Version]
- Biedma, M.E.; Cayet, D.; Tabareau, J.; Rossi, A.H.; Ivicak-Kocjan, K.; Moreno, G.; Errea, A.; Soulard, D.; Parisi, G.; Jerala, R.; et al. Recombinant flagellins with deletions in domains D1, D2, and D3: Characterization as novel immunoadjuvants. Vaccine 2019, 37, 652–663. [Google Scholar] [CrossRef] [PubMed]
- Campodonico, V.L.; Llosa, N.J.; Grout, M.; Doring, G.; Maira-Litran, T.; Pier, G.B. Evaluation of flagella and flagellin of Pseudomonas aeruginosa as vaccines. Infect. Immun. 2010, 78, 746–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turley, C.B.; Rupp, R.E.; Johnson, C.; Taylor, D.N.; Wolfson, J.; Tussey, L.; Kavita, U.; Stanberry, L.; Shaw, A. Safety and immunogenicity of a recombinant M2e-flagellin influenza vaccine (STF2.4xM2e) in healthy adults. Vaccine 2011, 29, 5145–5152. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Li, Z.; Zhu, X.; Cao, Y.; Chen, X. Improving immunogenicity and safety of flagellin as vaccine carrier by high-density display on virus-like particle surface. Biomaterials 2020, 249, 120030. [Google Scholar] [CrossRef]
- Chakraborty, C.; Sharma, A.R.; Bhattacharya, M.; Sharma, G.; Lee, S.S.; Agoramoorthy, G. Consider TLR5 for new therapeutic development against COVID-19. J. Med. Virol. 2020, 92, 2314–2315. [Google Scholar] [CrossRef] [PubMed]
- Westerlund-Wikström, B. Peptide display on bacterial flagella: Principles and applications. Int. J. Med Microbiol. 2000, 290, 223–230. [Google Scholar] [CrossRef]
- Lu, Z.; Murray, K.S.; Van Cleave, V.; LaVallie, E.R.; Stahl, M.L.; McCoy, J.M. Expression of thioredoxin random peptide libraries on the Escherichia coli cell surface as functional fusions to flagellin: A system designed for exploring protein-protein interactions. Biotechnology 1995, 13, 366–372. [Google Scholar] [CrossRef]
- Szabo, V.; Muskotal, A.; Toth, B.; Mihovilovic, M.D.; Vonderviszt, F. Construction of a xylanase A variant capable of polymerization. PLoS ONE 2011, 6, e25388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein, A.; Toth, B.; Jankovics, H.; Muskotal, A.; Vonderviszt, F. A polymerizable GFP variant. Protein Eng. Des. Sel. 2012, 25, 153–157. [Google Scholar] [CrossRef] [Green Version]
- Kovacs, N.; Patko, D.; Orgovan, N.; Kurunczi, S.; Ramsden, J.J.; Vonderviszt, F.; Horvath, R. Optical anisotropy of flagellin layers: In situ and label-free measurement of adsorbed protein orientation using OWLS. Anal. Chem. 2013, 85, 5382–5389. [Google Scholar] [CrossRef]
- Jankovics, H.; Kovacs, B.; Saftics, A.; Gerecsei, T.; Toth, E.; Szekacs, I.; Vonderviszt, F.; Horvath, R. Grating-coupled interferometry reveals binding kinetics and affinities of Ni ions to genetically engineered protein layers. Sci. Rep. 2020, 10, 22253. [Google Scholar] [CrossRef] [PubMed]
- Kovacs, B.; Patko, D.; Szekacs, I.; Orgovan, N.; Kurunczi, S.; Sulyok, A.; Khanh, N.Q.; Toth, B.; Vonderviszt, F.; Horvath, R. Flagellin based biomimetic coatings: From cell-repellent surfaces to highly adhesive coatings. Acta Biomater. 2016, 42, 66–76. [Google Scholar] [CrossRef] [PubMed]
- Kumara, M.T.; Srividya, N.; Muralidharan, S.; Tripp, B.C. Bioengineered flagella protein nanotubes with cysteine loops: Self-assembly and manipulation in an optical trap. Nano Lett. 2006, 6, 2121–2129. [Google Scholar] [CrossRef]
- Deplanche, K.; Woods, R.D.; Mikheenko, I.P.; Sockett, R.E.; Macaskie, L.E. Manufacture of stable palladium and gold nanoparticles on native and genetically engineered flagella scaffolds. Biotechnol. Bioeng. 2008, 101, 873–880. [Google Scholar] [CrossRef]
- Bereczk-Tompa, E.; Posfai, M.; Toth, B.; Vonderviszt, F. Magnetite-Binding Flagellar Filaments Displaying the MamI Loop Motif. Chembiochem 2016, 17, 2075–2082. [Google Scholar] [CrossRef]
- Kumara, M.T.; Tripp, B.C.; Muralidharan, S. Self-Assembly of Metal Nanoparticles and Nanotubes on Bioengineered Flagella Scaffolds. Chem. Mater. 2007, 19, 2056–2064. [Google Scholar] [CrossRef]
- Farkas, E.; Patko, D.; Khanh, N.Q.; Toth, E.; Vonderviszt, F.; Horvath, R. Self-assembly and structure of flagellin–polyelectrolyte composite layers: Polyelectrolyte induced flagellar filament formation during the alternating deposition process. RSC Adv. 2016, 6, 92159–92167. [Google Scholar] [CrossRef]
- Eckhard, U.; Bandukwala, H.; Mansfield, M.J.; Marino, G.; Cheng, J.; Wallace, I.; Holyoak, T.; Charles, T.C.; Austin, J.; Overall, C.M.; et al. Discovery of a proteolytic flagellin family in diverse bacterial phyla that assembles enzymatically active flagella. Nat. Commun. 2017, 8, 521. [Google Scholar] [CrossRef] [PubMed]
- Mi, Z.; Feng, Z.C.; Li, C.; Yang, X.; Ma, M.T.; Rong, P.F. Salmonella-Mediated Cancer Therapy: An Innovative Therapeutic Strategy. J. Cancer 2019, 10, 4765–4776. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | Function | Sigma Factor | Class in Flagellar Regulatory Hierarchy | Transcriptional Expression Regulators | Post-Transcriptional Regulators | Specific Secretion Chaperone | |
|---|---|---|---|---|---|---|---|
| 3-Tiered | 4-Tiered | ||||||
| Flagellin (FliC, FlaA) | Structural component of the filament | Sigma28 Sigma54 Sigma43 | Class III gene | Class IV gene | CodY Environmental factors (nutrients, c-di-GMP, ppGpp, BCAA, temperature…) | Self-regulating CsrA-FliW | FliS |
| FliD | Filament cap | Sigma70 Sigma28 Sigma54 | Class II and III | Class II | Cognate flagellin | ? | FliT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nedeljković, M.; Sastre, D.E.; Sundberg, E.J. Bacterial Flagellar Filament: A Supramolecular Multifunctional Nanostructure. Int. J. Mol. Sci. 2021, 22, 7521. https://doi.org/10.3390/ijms22147521
Nedeljković M, Sastre DE, Sundberg EJ. Bacterial Flagellar Filament: A Supramolecular Multifunctional Nanostructure. International Journal of Molecular Sciences. 2021; 22(14):7521. https://doi.org/10.3390/ijms22147521
Chicago/Turabian StyleNedeljković, Marko, Diego Emiliano Sastre, and Eric John Sundberg. 2021. "Bacterial Flagellar Filament: A Supramolecular Multifunctional Nanostructure" International Journal of Molecular Sciences 22, no. 14: 7521. https://doi.org/10.3390/ijms22147521
APA StyleNedeljković, M., Sastre, D. E., & Sundberg, E. J. (2021). Bacterial Flagellar Filament: A Supramolecular Multifunctional Nanostructure. International Journal of Molecular Sciences, 22(14), 7521. https://doi.org/10.3390/ijms22147521