
Somatotroph Tumors and the Epigenetic Status of the GNAS Locus


 , ,
, ,
Abstract
:1. Introduction
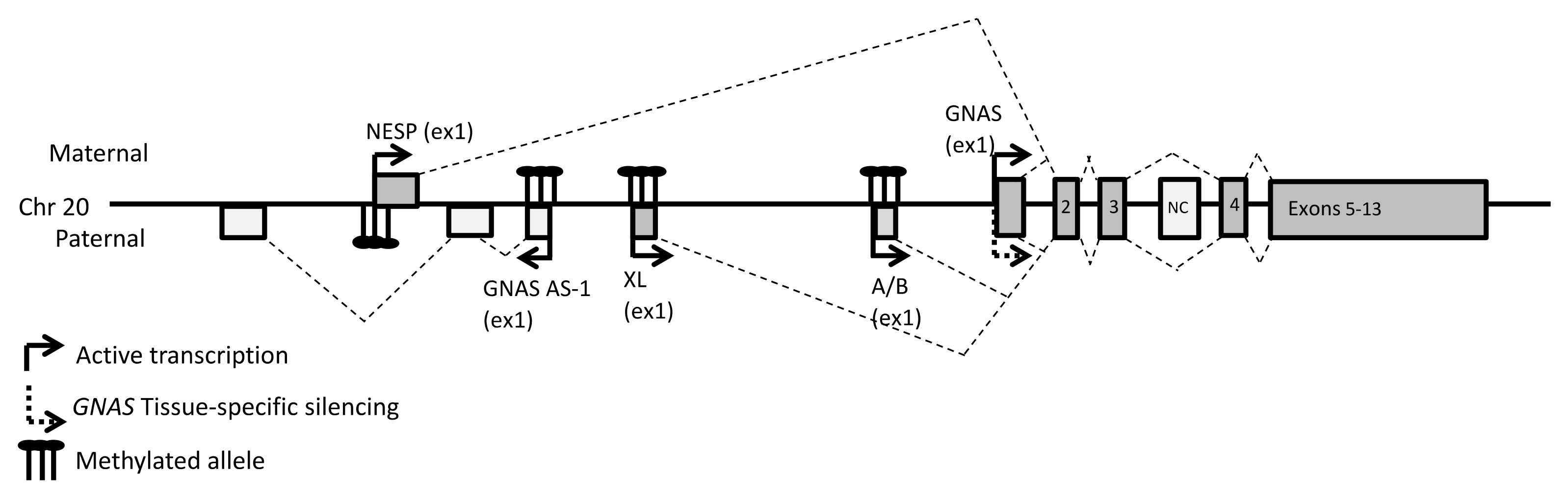
2. Materials and Methods
2.1. Somatotroph Tumors
2.2. Genetic and Epigenetic Analysis
2.3. Expression Analysis of GNAS, SSTR2 and AIP of Cohort II:
2.4. Allelic Quantification (AQ) of GNAS Expression
2.5. Statistical Analysis
3. Results
3.1. Cohort I
3.2. Cohort II
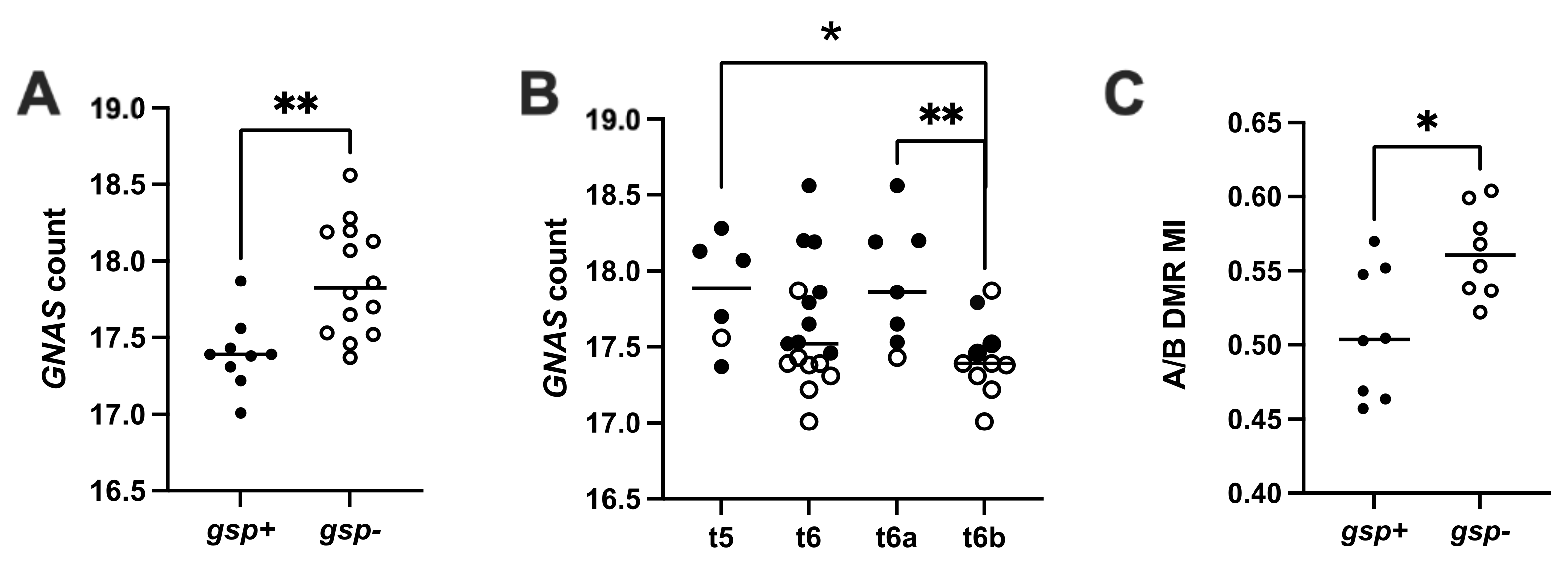
3.2.1. Gsp Oncogene Status and GNAS Expression
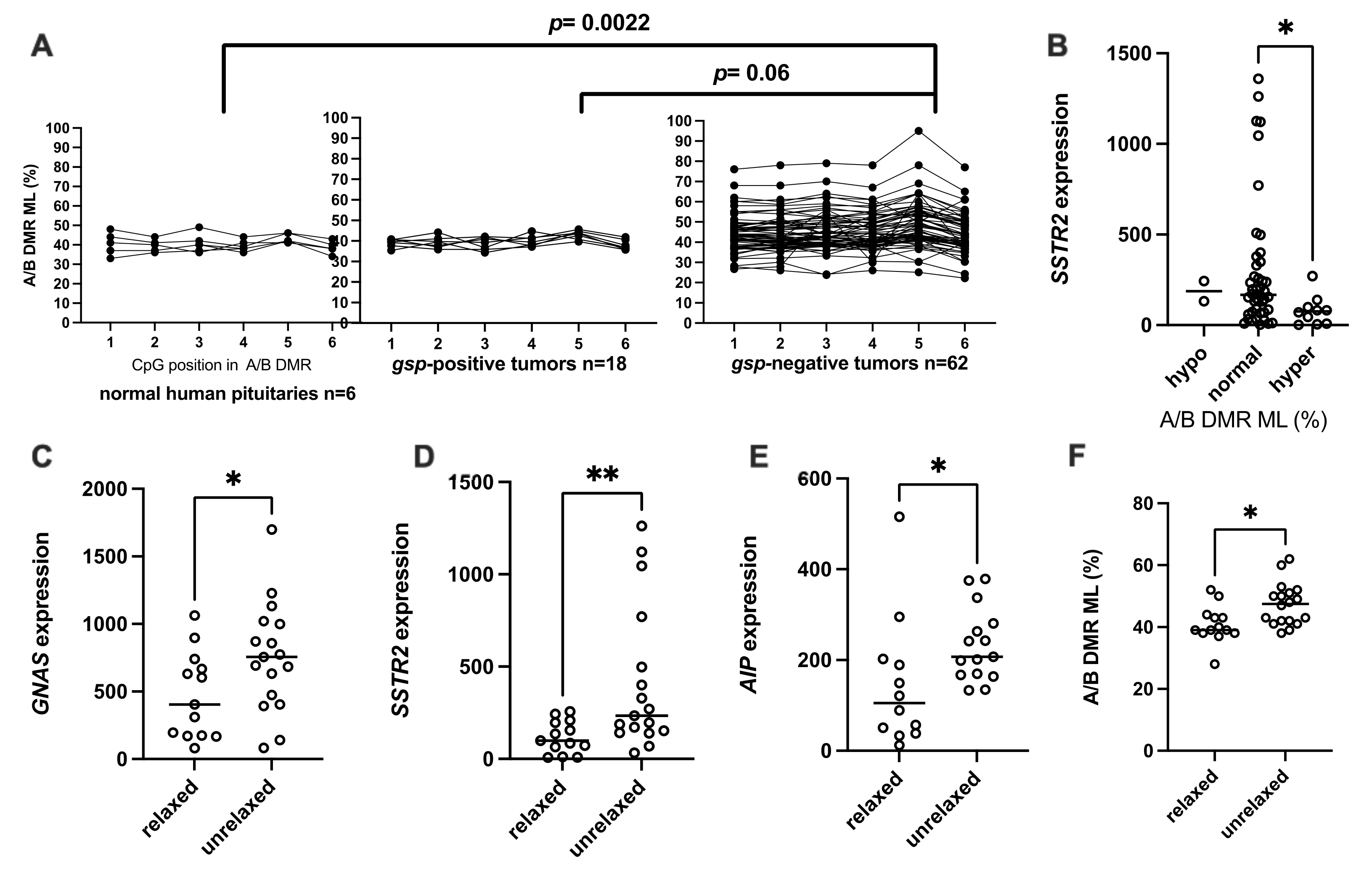
3.2.2. Methylation Levels of A/B DMR (A/B ML)
3.2.3. Relaxation of GNAS Imprinting
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ludecke, D.K.; Abe, T. Transsphenoidal microsurgery for newly diagnosed acromegaly: A personal view after more than 1,000 operations. Neuroendocrinology 2006, 83, 230–239. [Google Scholar] [CrossRef] [PubMed]
- Vélez, E.J.; Unniappan, S. A Comparative Update on the Neuroendocrine Regulation of Growth Hormone in Vertebrates. Front. Endocrinol. Lausanne 2021, 11, 1174. [Google Scholar] [CrossRef] [PubMed]
- Plöckinger, U.; Albrecht, S.; Mawrin, C.; Saeger, W.; Buchfelder, M.; Petersenn, S.; Schulz, S. Selective loss of somatostatin receptor 2 in octreotide-resistant growth hormone-secreting adenomas. J. Clin. Endocrinol. Metab. 2008, 93, 1203–1210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landis, C.A.; Masters, S.B.; Spada, A.; Pace, A.M.; Bourne, H.R.; Vallar, L. GTPase inhibiting mutations activate the alpha chain of Gs and stimulate adenylyl cyclase in human pituitary tumours. Nature 1989, 340, 692–696. [Google Scholar] [CrossRef]
- Lyons, J.; Landis, C.; Harsh, G.; Vallar, L.; Grunewald, K.; Feichtinger, H.; Duh, Q.; Clark, O.; Kawasaki, E.; Bourne, H.; et al. Two G protein oncogenes in human endocrine tumors. Science 1990, 249, 655–659. [Google Scholar] [CrossRef]
- Romano, D.; Magalon, K.; Pertuit, M.; Rasolonjanahary, R.; Barlier, A.; Enjalbert, A.; Gerard, C. Conditional Overexpression of the Wild-Type Gs α as the gsp Oncogene Initiates Chronic Extracellularly Regulated Kinase 1/2 Activation and Hormone Hypersecretion in Pituitary Cell Lines. Endocrinology 2007, 148, 2973–2983. [Google Scholar] [CrossRef] [Green Version]
- Pertuit, M.; Barlier, A.; Enjalbert, A.; Gérard, C. Signalling pathway alterations in pituitary adenomas: Involvement of Gsalpha, cAMP and mitogen-activated protein kinases. J. Neuroendocrinol. 2009, 21, 869–877. [Google Scholar] [CrossRef]
- Hayward, B.E.; Barlier, A.; Korbonits, M.; Grossman, A.B.; Jacquet, P.; Enjalbert, A.; Bonthron, D.T. Imprinting of the G(s)alpha gene GNAS1 in the pathogenesis of acromegaly. J. Clin. Investig. 2001, 107, R31–R36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barlier, A.; Pellegrini-Bouiller, I.; Gunz, G.; Zamora, A.J.; Jaquet, P.; Enjalbert, A. Impact of gsp oncogene on the expression of genes coding for G(s)α, Pit-1, G(i)2α, and somatostatin receptor 2 in human somatotroph adenomas: Involvement in octreotide sensitivity. J. Clin. Endocrinol. Metab. 1999, 84, 2759–2765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pertuit, M.; Romano, D.; Zeiller, C.; Barlier, A.; Enjalbert, A.; Gerard, C. The gsp oncogene disrupts Ras/ERK-dependent prolactin gene regulation in gsp inducible somatotroph cell line. Endocrinology 2011, 152, 1234–1243. [Google Scholar] [CrossRef] [Green Version]
- Hayward, B.E.; Kamiya, M.; Strain, L.; Moran, V.; Campbell, R.; Hayashizaki, Y.; Bonthron, D.T. The human GNAS1 gene is imprinted and encodes distinct paternally and biallelically expressed G proteins. Proc. Natl. Acad. Sci. USA 1998, 95, 10038–10043. [Google Scholar] [CrossRef] [Green Version]
- Freson, K.; Jaeken, J.; Van Helvoirt, M.; de Zegher, F.; Wittevrongel, C.; Thys, C.; Hoylaerts, M.F.; Vermylen, J.; Van Geet, C. Functional polymorphisms in the paternally expressed XLαs and its cofactor ALEX decrease their mutual interaction and enhance receptor-mediated cAMP formation. Hum. Mol. Genet. 2003, 12, 1121–1130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Erlichman, B.; Weinstein, L.S. The stimulatory G protein alpha-subunit Gs alpha is imprinted in human thyroid glands: Implications for thyroid function in pseudohypoparathyroidism types 1A and 1B. J. Clin. Endocrinol. Metab. 2003, 88, 4336–4341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mantovani, G.; Ballare, E.; Giammona, E.; Beck-Peccoz, P.; Spada, A. The gsalpha gene: Predominant maternal origin of transcription in human thyroid gland and gonads. J. Clin. Endocrinol. Metab. 2002, 87, 4736–4740. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Yu, S.; Litman, D.; Chen, W.; Weinstein, L.S. Identification of a methylation imprint mark within the mouse Gnas locus. Mol. Cell. Biol. 2000, 20, 5808–5817. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Litman, D.; Rosenberg, M.J.; Yu, S.; Biesecker, L.G.; Weinstein, L.S. A GNAS1 imprinting defect in pseudohypoparathyroidism type IB. J. Clin. Investig. 2000, 106, 1167–1174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ballaré, E.; Mantovani, S.; Lania, A.; Di Blasio, A.M.; Vallar, L.; Spada, A. Activating mutations of the G(s)α gene are associated with low levels of Gsα protein in growth hormone-secreting tumors. J. Clin. Endocrinol. Metab. 1998, 83, 4386–4390. [Google Scholar] [CrossRef] [Green Version]
- Lania, A.; Mantovani, G.; Spada, A. Genetics of pituitary tumors: Focus on G-protein mutations. Exp. Biol. Med. 2003, 228, 1004–1017. [Google Scholar] [CrossRef] [PubMed]
- Picard, C.; Silvy, M.; Gerard, C.; Buffat, C.; Lavaque, E.; Figarella-Branger, D.; Dufour, H.; Gabert, J.; Beckers, A.; Brue, T.; et al. Gs alpha overexpression and loss of Gs alpha imprinting in human somatotroph adenomas: Association with tumor size and response to pharmacologic treatment. Int. J. Cancer 2007, 121, 1245–1252. [Google Scholar] [CrossRef] [PubMed]
- Neou, M.; Villa, C.; Armignacco, R.; Jouinot, A.; Raffin-Sanson, M.L.; Septier, A.; Letourneur, F.; Diry, S.; Diedisheim, M.; Izac, B.; et al. Pangenomic Classification of Pituitary Neuroendocrine Tumors. Cancer Cell 2020, 37, 123–134. [Google Scholar] [CrossRef] [PubMed]
- Romanet, P.; Osei, L.; Netchine, I.; Pertuit, M.; Enjalbert, A.; Reynaud, R.; Barlier, A. Case report of GNAS epigenetic defect revealed by a congenital hypothyroidism. Pediatrics 2015, 135, e1079–e1083. [Google Scholar] [CrossRef] [Green Version]
- Romanet, P.; Guerin, C.; Pedini, P.; Essamet, W.; Castinetti, F.; Sebag, F.; Roche, P.; Cascon, A.; Tischler, A.S.; Pacak, K.; et al. Pathological and Genetic Characterization of Bilateral Adrenomedullary Hyperplasia in a Patient with Germline MAX Mutation. Endocr. Pathol. 2017, 28, 302–307. [Google Scholar] [CrossRef] [PubMed]
- Ronchi, C.L.; Peverelli, E.; Herterich, S.; Weigand, I.; Mantovani, G.; Schwarzmayr, T.; Sbiera, S.; Allolio, B.; Honegger, J.; Appenzeller, S.; et al. Landscape of somatic mutations in sporadic GH-secreting pituitary adenomas. Eur. J. Endocrinol. 2016, 174, 363–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaffrain-Rea, M.-L.; Angelini, M.; Gargano, D.; Tichomirowa, M.A.; Daly, A.F.; Vanbellinghen, J.-F.; D’Innocenzo, E.; Barlier, A.; Giangaspero, F.; Esposito, V.; et al. Expression of aryl hydrocarbon receptor (AHR) and AHR-interacting protein in pituitary adenomas: Pathological and clinical implications. Endocr. Relat. Cancer 2009, 16, 1029–1043. [Google Scholar] [CrossRef] [PubMed]
- Formosa, R.; Xuereb-Anastasi, A.; Vassallo, J. Aip regulates cAMP signalling and GH secretion in GH3 cells. Endocr. Relat. Cancer 2013, 20, 495–505. [Google Scholar] [CrossRef] [Green Version]
- Tuominen, I.; Heliövaara, E.; Raitila, A.; Rautiainen, M.-R.; Mehine, M.; Katainen, R.; Donner, I.; Aittomäki, V.; Lehtonen, H.J.; Ahlsten, M.; et al. AIP inactivation leads to pituitary tumorigenesis through defective Gαi-cAMP signaling. Oncogene 2014, 34, 1174–1184. [Google Scholar] [CrossRef]
- Jaffrain-Rea, M.-L.; Rotondi, S.; Turchi, A.; Occhi, G.; Barlier, A.; Peverelli, E.; Rostomyan, L.; Defilles, C.; Angelini, M.; Oliva, M.-A.; et al. Somatostatin analogues increase AIP expression in somatotropinomas, irrespective of Gsp mutations. Endocr. Relat. Cancer 2013, 20, 753–766. [Google Scholar] [CrossRef] [Green Version]
- Williamson, C.M.; Ball, S.T.; Nottingham, W.T.; Skinner, J.A.; Plagge, A.; Turner, M.D.; Powles, N.; Hough, T.; Papworth, D.; Fraser, W.D.; et al. A cis -acting control region is required exclusively for the tissue-specific imprinting of Gnas. Nat. Genet. 2004, 36, 894–899. [Google Scholar] [CrossRef]
- Liu, J.; Chen, M.; Deng, C.; Bourc’his, D.; Nealon, J.G.; Erlichman, B.; Bestor, T.H.; Weinstein, L.S. From the Cover: Identification of the control region for tissue-specific imprinting of the stimulatory G protein α-subunit. Proc. Natl. Acad. Sci. USA 2005, 102, 5513. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group 1 n = 16 | Group 2 n = 18 | Group 3 n = 14 | |
|---|---|---|---|
| GNAS mutational and imprinting status | gsp-positive | gsp-negative, unrelaxed | gsp-negative, relaxed |
| GNAS expression | Low * (p = 0.02) | High ** (p = 0.05) | Low * |
| AIP expression | Low * (ns) | High ** (p = 0.02) | Low ** |
| SSTR2 expression | Low * (ns) | High ** (p = 0.007) | Low ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Romanet, P.; Galluso, J.; Kamenicky, P.; Hage, M.; Theodoropoulou, M.; Roche, C.; Graillon, T.; Etchevers, H.C.; De Murat, D.; Mougel, G.; et al. Somatotroph Tumors and the Epigenetic Status of the GNAS Locus. Int. J. Mol. Sci. 2021, 22, 7570. https://doi.org/10.3390/ijms22147570
Romanet P, Galluso J, Kamenicky P, Hage M, Theodoropoulou M, Roche C, Graillon T, Etchevers HC, De Murat D, Mougel G, et al. Somatotroph Tumors and the Epigenetic Status of the GNAS Locus. International Journal of Molecular Sciences. 2021; 22(14):7570. https://doi.org/10.3390/ijms22147570
Chicago/Turabian StyleRomanet, Pauline, Justine Galluso, Peter Kamenicky, Mirella Hage, Marily Theodoropoulou, Catherine Roche, Thomas Graillon, Heather C. Etchevers, Daniel De Murat, Grégory Mougel, and et al. 2021. "Somatotroph Tumors and the Epigenetic Status of the GNAS Locus" International Journal of Molecular Sciences 22, no. 14: 7570. https://doi.org/10.3390/ijms22147570
APA StyleRomanet, P., Galluso, J., Kamenicky, P., Hage, M., Theodoropoulou, M., Roche, C., Graillon, T., Etchevers, H. C., De Murat, D., Mougel, G., Figarella-Branger, D., Dufour, H., Cuny, T., Assié, G., & Barlier, A. (2021). Somatotroph Tumors and the Epigenetic Status of the GNAS Locus. International Journal of Molecular Sciences, 22(14), 7570. https://doi.org/10.3390/ijms22147570