
Overexpression of OsERF83, a Vascular Tissue-Specific Transcription Factor Gene, Confers Drought Tolerance in Rice

 , ,
, ,
Abstract
:1. Introduction
2. Results
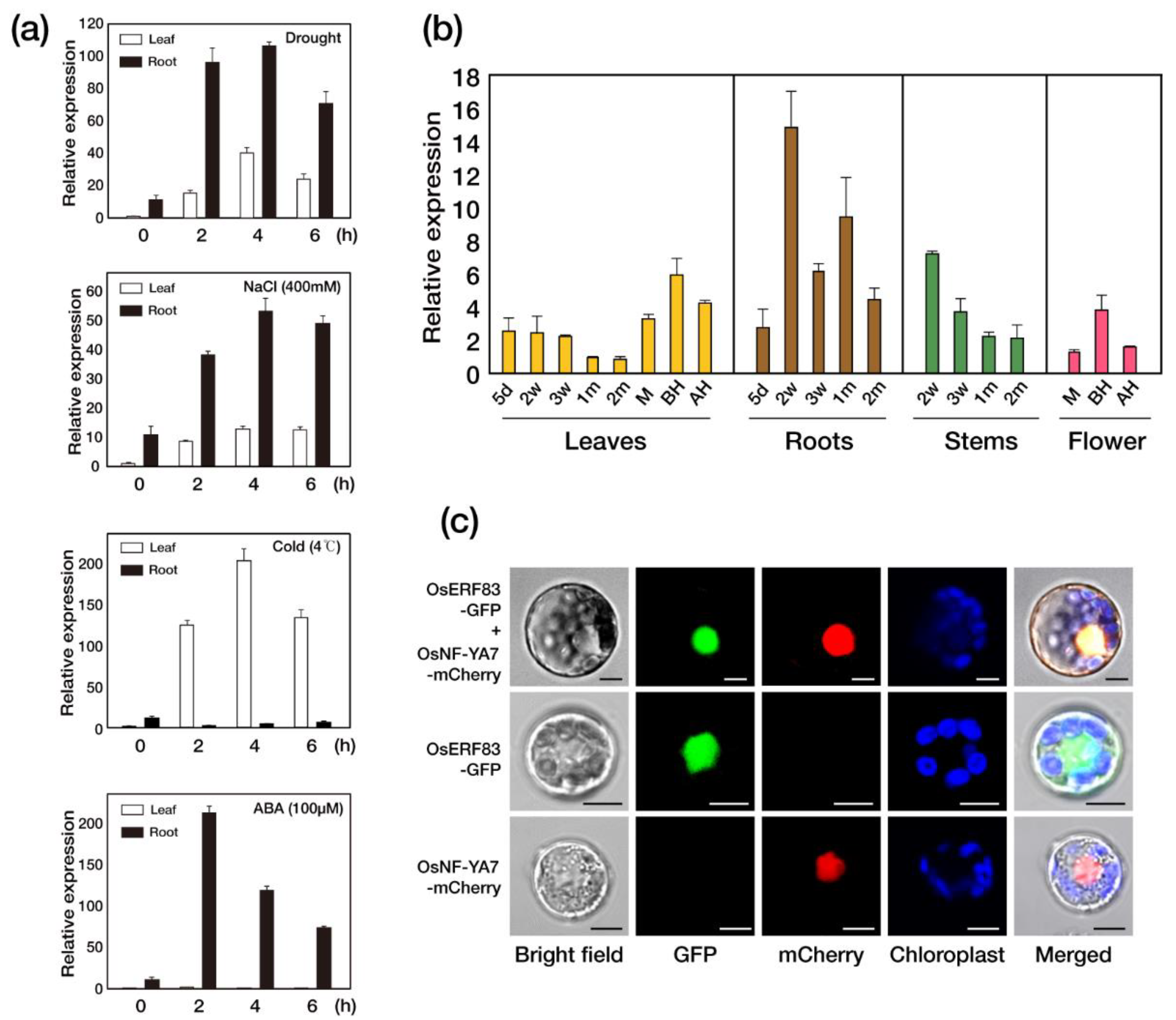
2.1. OsERF83 Is a Drought-Inducible Transcription Factor
2.2. Overexpression of OsERF83 in Rice Confers Drought Tolerance at the Vegetative Stage
2.3. Overexpression of OsERF83 in Rice Leads to Growth Retardation and Affects Grain Yield under Normal Conditions
2.4. CRISPR/Cas9-Mediated Loss-of-Function Study Analysis of OsERF83
2.5. OsERF83 Is Expressed Predominantly in the Vascular Tissue
2.6. Identification of Genes Involved in the OsERF83-Mediated Drought Tolerance Pathway
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Stress Treatments
4.3. Subcellular Localization of OsERF83
4.4. RNA Extraction, and Quantitative Real-Time PCR (qRT-PCR) Analysis
4.5. Histochemical GUS Assay
4.6. RNA-Sequencing Analysis
4.7. Drought Stress Treatment and Measurement of Chlorophyll Fluorescence
4.8. Statistical Analysis
4.9. Accession Numbers
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zandalinas, S.I.; Mittler, R.; Balfagón, D.; Arbona, V.; Gómez-Cadenas, A. Plant adaptations to the combination of drought and high temperatures. Physiol. Plant. 2017, 162, 2–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mickelbart, M.V.; Hasegawa, P.M.; Bailey-Serres, J. Genetic mechanisms of abiotic stress tolerance that translate to crop yield stability. Nat. Rev. Genet. 2015, 16, 237–251. [Google Scholar] [CrossRef]
- Lesk, C.; Rowhani, P.; Ramankutty, N. Influence of extreme weather disasters on global crop production. Nat. Cell Biol. 2016, 529, 84–87. [Google Scholar] [CrossRef]
- Barnabás, B.; Jäger, K.; Fehér, A. The effect of drought and heat stress on reproductive processes in cereals. Plant Cell Environ. 2007, 31, 11–38. [Google Scholar] [CrossRef]
- Hirayama, T.; Shinozaki, K. Research on plant abiotic stress responses in the post-genome era: Past, present and future. Plant. J. 2010, 61, 1041–1052. [Google Scholar] [CrossRef]
- Atkinson, N.J.; Urwin, P.E. The interaction of plant biotic and abiotic stresses: From genes to the field. J. Exp. Bot. 2012, 63, 3523–3543. [Google Scholar] [CrossRef] [Green Version]
- Nakano, T.; Suzuki, K.; Fujimura, T.; Shinshi, H. Genome-Wide Analysis of the ERF Gene Family in Arabidopsis and Rice. Plant. Physiol. 2006, 140, 411–432. [Google Scholar] [CrossRef] [Green Version]
- Sakuma, Y.; Liu, Q.; Dubouzet, J.G.; Abe, H.; Shinozaki, K.; Yamaguchi-Shinozaki, K. DNA-binding specificity of the ERF/AP2 domain of Arabidopsis DREBs, transcription factors involved in dehydration- and cold-inducible gene expression. Biochem. Bioph. Res. Co. 2002, 290, 998–1009. [Google Scholar] [CrossRef]
- Oh, S.-J.; Kim, Y.S.; Kwon, C.-W.; Park, H.K.; Jeong, J.S.; Kim, J.-K. Overexpression of the transcription factor AP37 in rice improves grain yield under drought conditions. Plant. Physiol. 2009, 150, 1368–1379. [Google Scholar] [CrossRef] [Green Version]
- Mizoi, J.; Shinozaki, K.; Yamaguchi-Shinozaki, K. AP2/ERF family transcription factors in plant abiotic stress responses. Biochim. Biophys. Acta Bioenerg. 2012, 1819, 86–96. [Google Scholar] [CrossRef]
- Müller, M.; Munné-Bosch, S. Ethylene Response Factors: A Key Regulatory Hub in Hormone and Stress Signaling. Plant. Physiol. 2015, 169, 32–41. [Google Scholar] [CrossRef] [Green Version]
- Xie, Z.; Nolan, T.; Jiang, H.; Yin, Y. AP2/ERF Transcription Factor Regulatory Networks in Hormone and Abiotic Stress Responses in Arabidopsis. Front. Plant. Sci. 2019, 10, 228. [Google Scholar] [CrossRef] [Green Version]
- Cheng, M.-C.; Liao, P.-M.; Kuo, W.-W.; Lin, T.-P. The Arabidopsis ETHYLENE RESPONSE FACTOR1 Regulates Abiotic Stress-Responsive Gene Expression by Binding to Different cis-Acting Elements in Response to Different Stress Signals. Plant. Physiol. 2013, 162, 1566–1582. [Google Scholar] [CrossRef] [Green Version]
- Lorenzo, O.; Piqueras, R.; Serrano, J.J.S.; Solano, R. Ethylene Response FACTOR1 Integrates Signals from Ethylene and Jasmonate Pathways in Plant Defense. Plant. Cell 2003, 15, 165–178. [Google Scholar] [CrossRef] [Green Version]
- Jung, H.; Chung, P.J.; Park, S.-H.; Redillas, M.C.F.R.; Kim, Y.S.; Suh, J.-W.; Kim, J.-K. Overexpression of OsERF48 causes regulation of OsCML16, a calmodulin-like protein gene that enhances root growth and drought tolerance. Plant. Biotechnol. J. 2017, 15, 1295–1308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, D.K.; Jung, H.; Jang, G.; Jeong, J.S.; Kim, Y.S.; Ha, S.H.; Do Choi, Y.; Kim, J.K. Overexpression of the OsERF71 Tran-scription Factor Alters Rice Root Structure and Drought Resistance. Plant. Physiol. 2016, 172, 575–588. [Google Scholar] [CrossRef] [Green Version]
- Jisha, V.; Dampanaboina, L.; Vadassery, J.; Mithöfer, A.; Kappara, S.; Ramanan, R. Overexpression of an AP2/ERF Type Transcription Factor OsEREBP1 Confers Biotic and Abiotic Stress Tolerance in Rice. PLoS ONE 2015, 10, e0127831. [Google Scholar] [CrossRef] [PubMed]
- Scarpeci, T.E.; Frea, V.S.; Zanor, M.I.; Valle, E.M. Overexpression of AtERF019 delays plant growth and senescence, and improves drought tolerance in Arabidopsis. J. Exp. Bot. 2017, 68, 673–685. [Google Scholar] [PubMed] [Green Version]
- Berrocal-Lobo, M.; Molina, A.; Solano, R. Constitutive expression of ETHYLENE-RESPONSE-FACTOR1 in Arabidopsis con-fers resistance to several necrotrophic fungi. Plant. J. 2002, 29, 23–32. [Google Scholar] [CrossRef]
- Pré, M.; Atallah, M.; Champion, A.; De Vos, M.; Pieterse, C.; Memelink, J. The AP2/ERF Domain Transcription Factor ORA59 Integrates Jasmonic Acid and Ethylene Signals in Plant Defense. Plant. Physiol. 2008, 147, 1347–1357. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; Chen, X.; Liu, J.; Ye, J.; Guo, Z. The rice ERF transcription factor OsERF922 negatively regulates resistance to Magnaporthe oryzae and salt tolerance. J. Exp. Bot. 2012, 63, 3899–3911. [Google Scholar] [CrossRef] [Green Version]
- Tezuka, D.; Kawamata, A.; Kato, H.; Saburi, W.; Mori, H.; Imai, R. The rice ethylene response factor OsERF83 positively regulates disease resistance to Magnaporthe oryzae. Plant. Physiol. Biochem. 2019, 135, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.K.; Kim, H.I.; Jang, G.; Chung, P.J.; Jeong, J.S.; Kim, Y.S.; Bang, S.W.; Jung, H.; Choi, Y.D.; Kim, J.K. The NF-YA transcription factor OsNF-YA7 confers drought stress tolerance of rice in an abscisic acid independent manner. Plant. Sci. 2015, 241, 199–210. [Google Scholar] [CrossRef] [PubMed]
- Depater, B.S.; Vandermark, F.; Rueb, S.; Katagiri, F.; Chua, N.H.; Schilperoort, R.A.; Hensgens, L.A.M. The Promoter of the Rice Gene Gos2 Is Active in Various Different Monocot Tissues and Binds Rice Nuclear Factor Asf-1. Plant. J. 1992, 2, 837–844. [Google Scholar] [CrossRef]
- Bussotti, F.; Desotgiu, R.; Pollastrini, M.; Cascio, C. The JIP test: A tool to screen the capacity of plant adaptation to climate change. Scand. J. For. Res. 2010, 25, 43–50. [Google Scholar] [CrossRef]
- Chung, P.J.; Chung, H.; Oh, N.; Choi, J.; Bang, S.W.; Jung, S.E.; Jung, H.; Shim, J.S.; Kim, J.K. Efficiency of Recombinant CRISPR/rCas9-Mediated miRNA Gene Editing in Rice. Int. J. Mol. Sci. 2020, 21, 9606. [Google Scholar] [CrossRef] [PubMed]
- Kawahara, Y.; Oono, Y.; Wakimoto, H.; Ogata, J.; Kanamori, H.; Sasaki, H.; Mori, S.; Matsumoto, T.; Itoh, T. TENOR: Database for Comprehensive mRNA-Seq Experiments in Rice. Plant. Cell Physiol. 2016, 57, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, Y.; Namiki, N.; Takehisa, H.; Kamatsuki, K.; Minami, H.; Ikawa, H.; Ohyanagi, H.; Sugimoto, K.; Itoh, J.I.; Antonio, B.A.; et al. RiceFREND: A platform for retrieving coexpressed gene networks in rice. Nucl. Acids Res. 2013, 41, D1214–D1221. [Google Scholar] [CrossRef] [Green Version]
- Fujimoto, S.Y.; Ohta, M.; Usui, A.; Shinshi, H.; Ohme-Takagi, M. Arabidopsis ethylene-responsive element binding factors act as transcriptional activators or repressors of GCC box-mediated gene expression. Plant. Cell 2000, 12, 393–404. [Google Scholar]
- Kim, H.; Lee, K.; Hwang, H.; Bhatnagar, N.; Kim, D.Y.; Yoon, I.S.; Byun, M.O.; Kim, S.T.; Jung, K.H.; Kim, B.G. Overex-pression of PYL5 in rice enhances drought tolerance, inhibits growth, and modulates gene expression. J. Exp. Bot. 2014, 65, 453–464. [Google Scholar] [CrossRef] [Green Version]
- Ma, Z.; Wu, T.; Huang, K.; Jin, Y.-M.; Li, Z.; Chen, M.; Yun, S.; Zhang, H.; Yang, X.; Chen, H.; et al. A Novel AP2/ERF Transcription Factor, OsRPH1, Negatively Regulates Plant Height in Rice. Front. Plant. Sci. 2020, 11. [Google Scholar] [CrossRef]
- Lucas, W.J.; Groover, A.; Lichtenberger, R.; Furuta, K.; Yadav, S.-R.; Helariutta, Y.; He, X.-Q.; Fukuda, H.; Kang, J.; Brady, S.; et al. The Plant Vascular System: Evolution, Development and Functions. J. Integr. Plant. Biol. 2013, 55, 294–388. [Google Scholar] [CrossRef] [PubMed]
- Geng, D.; Chen, P.; Shen, X.; Zhang, Y.; Li, X.; Jiang, L.; Xie, Y.; Niu, C.; Zhang, J.; Huang, X.; et al. MdMYB88 and MdMYB124 Enhance Drought Tolerance by Modulating Root Vessels and Cell Walls in Apple. Plant. Physiol. 2018, 178, 1296–1309. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.S.; Zhang, H.; Srivastava, A.K.; Pan, Y.J.; Bai, J.J.; Fang, J.J.; Shi, H.Z.; Zhu, J.K. Knockdown of Rice MicroRNA166 Confers Drought Resistance by Causing Leaf Rolling and Altering Stem Xylem Development. Plant. Physiol. 2018, 176, 2082–2094. [Google Scholar] [CrossRef] [Green Version]
- Zheng, L.; Huang, F.; Narsai, R.; Wu, J.; Giraud, E.; He, F.; Cheng, L.; Wang, F.; Wu, P.; Whelan, J.; et al. Physiological and Transcriptome Analysis of Iron and Phosphorus Interaction in Rice Seedlings. Plant. Physiol. 2009, 151, 262–274. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Wang, C.; Zheng, L.; Wang, L.; Chen, Y.; Whelan, J.; Shou, H. Ethylene is involved in the regulation of iron homeostasis by regulating the expression of iron-acquisition-related genes in Oryza sativa. J. Exp. Bot. 2010, 62, 667–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsieh, P.-H.; Kan, C.-C.; Wu, H.-Y.; Yang, H.-C.; Hsieh, M.-H. Early molecular events associated with nitrogen deficiency in rice seedling roots. Sci. Rep. 2018, 8, 1–23. [Google Scholar] [CrossRef]
- Agarwal, P.K.; Jha, B. Transcription factors in plants and ABA dependent and independent abiotic stress signalling. Biol. Plant. 2010, 54, 201–212. [Google Scholar] [CrossRef]
- Zhang, L.; Yu, Z.; Xu, Y.; Yu, M.; Ren, Y.; Zhang, S.; Yang, G.; Huang, J.; Yan, K.; Zheng, C.; et al. Regulation of the stability and ABA import activity of NRT1.2/NPF4.6 by CEPR2-mediated phosphorylation in Arabidopsis. Mol. Plant. 2021, 14, 633–646. [Google Scholar] [CrossRef]
- Bang, S.W.; Lee, D.-K.; Jung, H.; Chung, P.J.; Kim, Y.S.; Choi, Y.D.; Suh, J.-W.; Kim, J.-K. Overexpression of OsTF1L, a rice HD-Zip transcription factor, promotes lignin biosynthesis and stomatal closure that improves drought tolerance. Plant. Biotechnol. J. 2018, 17, 118–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tu, M.; Wang, X.; Yin, W.; Wang, Y.; Li, Y.; Zhang, G.; Li, Z.; Song, J.; Wang, X. Grapevine VlbZIP30 improves drought re-sistance by directly activating VvNAC17 and promoting lignin biosynthesis through the regulation of three peroxidase genes. Hortic. Res. 2020, 7, 150. [Google Scholar] [CrossRef]
- Sun, J.; Cui, X.; Teng, S.; Kunnong, Z.; Wang, Y.; Chen, Z.; Sun, X.; Wu, J.; Ai, P.; Quick, W.P.; et al. HD-ZIP IV gene Roc8 regulates the size of bulliform cells and lignin content in rice. Plant. Biotechnol. J. 2020, 18, 2559–2572. [Google Scholar] [CrossRef]
- Li, W.Q.; Zhang, M.J.; Gan, P.F.; Qiao, L.; Yang, S.Q.; Miao, H.; Wang, G.F.; Zhang, M.M.; Liu, W.T.; Li, H.F.; et al. CLD1/SRL1 modulates leaf rolling by affecting cell wall formation, epidermis integrity and water homeostasis in rice. Plant. J. 2017, 92, 904–923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Q.; Luo, L.; Wang, X.; Shen, Z.; Zheng, L. Comprehensive Analysis of Rice Laccase Gene (OsLAC) Family and Ectopic Expression of OsLAC10 Enhances Tolerance to Copper Stress in Arabidopsis. Int. J. Mol. Sci. 2017, 18, 209. [Google Scholar] [CrossRef]
- Barakat, A.; Bagniewska-Zadworna, A.; Choi, A.; Plakkat, U.; DiLoreto, D.S.; Yellanki, P.; Carlson, J.E. The cinnamyl alcohol dehydrogenase gene family in Populus: Phylogeny, organization, and expression. BMC Plant. Biol. 2009, 9. [Google Scholar] [CrossRef] [Green Version]
- Ubaidillah, M.; Kim, K.-A.; Kim, Y.H.; Lee, I.-J.; Yun, B.-W.; Kim, D.H.; Loake, G. Identification of a drought-induced rice gene, OsSAP, that suppresses Bax-induced cell death in yeast. Mol. Biol. Rep. 2013, 40, 6113–6121. [Google Scholar] [CrossRef]
- Kaur, R.; Chakraborty, A.; Bhunia, R.K.; Sen, S.K.; Ghosh, A.K. Tolerance to soil water stress by Oryza sativa cv. IR20 was improved by expression of Wsi18 gene locus from Oryza nivara. Biol. Plant. 2017, 62, 129–139. [Google Scholar] [CrossRef]
- Giarola, V.; Jung, N.U.; Singh, A.; Satpathy, P.; Bartels, D. Analysis of pcC13-62 promoters predicts a link between cis-element variations and desiccation tolerance in Linderniaceae. J. Exp. Bot. 2018, 69, 3773–3784. [Google Scholar] [CrossRef]
- Driedonks, N.; Xu, J.; Peters, J.; Park, S.; Rieu, I. Multi-Level Interactions Between Heat Shock Factors, Heat Shock Proteins, and the Redox System Regulate Acclimation to Heat. Front. Plant. Sci. 2015, 6, 999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ul Haq, S.; Khan, A.; Ali, M.; Khattak, A.M.; Gai, W.X.; Zhang, H.X.; Wei, A.M.; Gong, Z.H. Heat Shock Proteins: Dynamic Biomolecules to Counter Plant Biotic and Abiotic Stresses. Int. J. Molec. Sci. 2019, 20, 5321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, J.; Liu, C.; Liu, A.; Zou, D.; Chen, X. Overexpression of OsHsp17.0 and OsHsp23.7 enhances drought and salt tolerance in rice. J. Plant. Physiol. 2012, 169, 628–635. [Google Scholar] [CrossRef]
- Tamiru, M.; Undan, J.; Takagi, H.; Abe, A.; Yoshida, K.; Undan, J.Q.; Natsume, S.; Uemura, A.; Saitoh, H.; Matsumura, H.; et al. A cytochrome P450, OsDSS1, is involved in growth and drought stress responses in rice (Oryza sativa L.). Plant. Mol. Biol. 2015, 88, 85–99. [Google Scholar] [CrossRef]
- Pandian, B.A.; Sathishraj, R.; Djanaguiraman, M.; Prasad, P.V.; Jugulam, M. Role of Cytochrome P450 Enzymes in Plant Stress Response. Antioxidants 2020, 9, 454. [Google Scholar] [CrossRef]
- Wu, J.; Kim, S.G.; Kang, K.Y.; Kim, J.G.; Park, S.R.; Gupta, R.; Kim, Y.H.; Wang, Y.; Kim, S.T. Overexpression of a Path-ogenesis-Related Protein 10 Enhances Biotic and Abiotic Stress Tolerance in Rice. Plant. Pathol. J. 2016, 32, 552–562. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.-X.; Zhang, F.-C.; Zhang, W.-Z.; Song, L.-F.; Wu, W.-H.; Chen, Y.-F. Arabidopsis Di19 Functions as a Transcription Factor and Modulates PR1, PR2, and PR5 Expression in Response to Drought Stress. Mol. Plant. 2013, 6, 1487–1502. [Google Scholar] [CrossRef] [Green Version]
- Le Gall, H.; Philippe, F.; Domon, J.-M.; Gillet, F.; Pelloux, J.; Rayon, C. Cell Wall Metabolism in Response to Abiotic Stress. Plants 2015, 4, 112–166. [Google Scholar] [CrossRef]
- Houston, K.; Tucker, M.; Chowdhury, J.; Shirley, N.; Little, A. The Plant Cell Wall: A Complex and Dynamic Structure As Revealed by the Responses of Genes under Stress Conditions. Front. Plant. Sci. 2016, 7, 984. [Google Scholar] [CrossRef] [Green Version]
- Bang, S.W.; Park, S.H.; Kim, Y.S.; Do Choi, Y.; Kim, J.K. The activities of four constitutively expressed promoters in sin-gle-copy transgenic rice plants for two homozygous generations. Planta 2015, 241, 1529–1541. [Google Scholar] [CrossRef]
- Zhang, Y.; Su, J.B.; Duan, S.; Ao, Y.; Dai, J.R.; Liu, J.; Wang, P.; Li, Y.G.; Liu, B.; Feng, D.R.; et al. A highly efficient rice green tissue protoplast system for transient gene expression and studying light/chloroplast-related processes. Plant. Meth. 2011. [Google Scholar] [CrossRef] [Green Version]
- Redillas, M.C.F.R.; Bang, S.W.; Lee, D.; Kim, Y.S.; Jung, H.; Chung, P.J.; Suh, J.; Kim, J. Allantoin accumulation through overexpression of ureide permease1 improves rice growth under limited nitrogen conditions. Plant. Biotechnol. J. 2018, 17, 1289–1301. [Google Scholar] [CrossRef] [Green Version]
- Redillas, M.C.F.R.; Strasser, R.J.; Jeong, J.S.; Kim, Y.S.; Kim, J.-K. The use of JIP test to evaluate drought-tolerance of transgenic rice overexpressing OsNAC10. Plant. Biotechnol. Rep. 2011, 5, 169–175. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Descriptions | Gene ID | OX/NT Fc. | OX/NT Pval. | GCC Box |
|---|---|---|---|---|
| Transporters | ||||
| Similar to sugar transport protein 14 | Os09g0322000 | 12.80 | 0.000 | −1858, −1702, −902 |
| OsLTP2.7 | Os10g0504650 | 4.78 | 0.002 | |
| OsNPF8.10 | Os01g0142800 | 4.27 | 0.006 | |
| OsATL11 | Os02g0101000 | 3.95 | 0.003 | |
| ABC transporter-like domain-containing protein | Os08g0564300 | 3.14 | 0.021 | |
| NPF8.17 | Os10g0112500 | 2.53 | 0.040 | |
| OsACA1 | Os03g0203700 | 2.34 | 0.044 | −1439, −1052, −1049 |
| NPF5.1 | Os05g0336200 | 2.33 | 0.040 | |
| OsLHT1 | Os08g0127100 | 2.21 | 0.008 | −1281 |
| Transcription factors | ||||
| OsMSL38 | Os11g0282700 | 25.08 | 0.000 | −2906 |
| Zinc finger, RING-type domain-containing protein | Os06g0534800 | 11.73 | 0.002 | −2989, −2975 |
| OsC3H32 | Os04g0671800 | 10.06 | 0.000 | −2976, −2918, −2879, −2876, −2835, −1283, −1144 |
| Myb/SANT-like domain-containing protein | Os08g0496700 | 3.51 | 0.005 | −1455, −1191, −900 |
| OsC3H17, OsWD40–49 | Os02g0677700 | 3.44 | 0.015 | |
| OsbZIP89 | Os12g0634500 | 3.27 | 0.028 | −483 |
| OsABI5, OsbZIP10 | Os01g0859300 | 3.04 | 0.010 | −1554 |
| Zinc finger, RING/FYVE/PHD-type domain-containing protein | Os09g0504700 | 2.67 | 0.030 | −1835, −1803, −1740 |
| ONAC79 | Os04g0437000 | 2.63 | 0.038 | −2800, −506 |
| OsERF86 | Os07g0410700 | 2.62 | 0.029 | −2589, −2495, −657 |
| OsSPL1, OsDLN8 | Os01g0292900 | 2.47 | 0.000 | −2338, −2204, −1991 |
| OsMyb | Os01g0298400 | 2.45 | 0.011 | −1348, −1325, −1089, −889 |
| OsbHLH025 | Os01g0196300 | 2.41 | 0.046 | −1581, −1578 |
| Zinc finger, RING/FYVE/PHD-type domain-containing protein | Os07g0484300 | 2.16 | 0.045 | −2923, −2920, −888 |
| Abiotic stress-related genes | ||||
| OsDjA1 | Os02g0656500 | 26.99 | 0.000 | −2460, −2354, −2351 |
| OsSAP, OsTET13 | Os09g0425900 | 12.49 | 0.000 | −908 |
| OsTHIC | Os03g0679700 | 10.84 | 0.000 | −1881, −1878, −1875 |
| OsLEA14, wsi18 | Os01g0705200 | 9.95 | 0.000 | −2526 |
| Similar to sorting nexin 1 | Os01g0862300 | 6.86 | 0.000 | −2778, −1217, −982, −979, −690, −544, −487, −481, −470, −359, −329, −298, −278, −188, −29 |
| OsMPK14 | Os05g0143500 | 6.85 | 0.013 | |
| PCC13-62, OsEnS-64 | Os04g0404400 | 5.60 | 0.000 | −2980, −2977, −2974, −2223 |
| OsHSP23.7 | Os12g0569700 | 3.68 | 0.015 | −1065, −1018, −988, −957, −444, −441, −379, −174 |
| OsFAD2-3 | Os07g0416900 | 3.43 | 0.005 | −1476 |
| Similar to class III peroxidase 136 | Os12g0111800 | 3.29 | 0.001 | |
| OsBBX28 | Os09g0509700 | 2.65 | 0.009 | −2975 |
| OsNTP3 | Os01g0846500 | 2.63 | 0.040 | −1783, −1780, −1685, −1653 |
| OsPEX11−5 | Os06g0127000 | 2.49 | 0.019 | −2371, −2368, −2054, −2051, −2048 |
| Glycine-rich RNA-binding protein | Os04g0653700 | 2.37 | 0.032 | −2957, −2901, −2895, −2892, −2307, −2684, −2680, −1677, −1610, −1607, −1604, −1577, −1478, −1475, −1442, −1415, −1322, −1319, −1316, −1313, −710, −678 |
| OsSKIP | Os02g0759800 | 2.23 | 0.001 | −2986, −2831, −1142, −1139, −1136, −990, −906, −840, −837, −800 |
| OsRZFP34, OsRFP1 | Os01g0719100 | 2.21 | 0.027 | −1128, −1114, −346, −120 |
| OsVPE3 | Os02g0644000 | 2.20 | 0.000 | −2797, −2751 |
| OsASMT1 | Os09g0344500 | 2.10 | 0.007 | |
| Cytochrome P450 family | ||||
| Oscyp71Z4 | Os10g0439800 | 5.79 | 0.001 | −2642, −2625 |
| Cytochrome P450 family protein | Os12g0582666 | 4.86 | 0.001 | |
| CYP76M10 | Os08g0507100 | 4.09 | 0.007 | −1515, −1512, −1354, −1297, −1008, −572 |
| CYP450-like Gene | Os08g0105700 | 4.09 | 0.000 | −2870, −2337, −2065. −1006, −998, −995, −974 |
| Similar to cytochrome P450 family protein, expressed | Os12g0640200 | 3.53 | 0.010 | −1064, −1061, −1050, −989, −940, −240, −190, −187 |
| OsCYP71AA2 | Os01g0957800 | 3.50 | 0.022 | −2440, −2437, −2434, −2410, −2224, −2029 |
| Cytochrome P450 family protein | Os02g0184700 | 3.01 | 0.047 | −1217 |
| Cytochrome P450 family protein | Os03g0594100 | 2.54 | 0.022 | −2178 |
| CYP76M8 | Os02g0569400 | 2.28 | 0.039 | |
| CYP99A2 | Os04g0180400 | 2.26 | 0.046 | −2113, −2010, −1980, −1959 |
| CYP76M2 | Os08g0508000 | 2.15 | 0.012 | −2928, −937 |
| Terpenoid-associated genes | ||||
| OsTPS33 | Os12g0491800 | 4.05 | 0.001 | −608 |
| OsTPS14 | Os03g0428200 | 2.83 | 0.024 | −1604 |
| OsTPS3 | Os02g0121700 | 2.80 | 0.027 | |
| OsCPS4 | Os04g0178300 | 2.68 | 0.000 | −1983, −1933, −1926, −1897, −1825, −1797, −1794, −1786 |
| OsTPS16 | Os04g0179700 | 2.61 | 0.036 | |
| Disease resistance-related genes | ||||
| OsCSN5A | Os04g0654700 | 25.76 | 0.000 | |
| Similar to blight-associated protein p12 | Os09g0472700 | 23.77 | 0.000 | −2580, −1500, −1353, −1308, −1305, −1302, −1069, −1066, −1063, −995, −992, −927, −924, −130, −98, −37 |
| OsLysM-RLK8 | Os11g0549300 | 13.44 | 0.000 | −2808, −2279, −2093, −2021, −1995, −1992, −1972 |
| OsCslE1 | Os09g0478100 | 10.69 | 0.000 | −2802 |
| eIF4Gtsv1 | Os07g0555200 | 9.14 | 0.000 | −1520 |
| OsAGP29 | Os01g0607100 | 7.42 | 0.000 | −2606 |
| OsWAK89b | Os09g0561450 | 6.55 | 0.005 | |
| OsTHI7 | Os06g0514100 | 5.48 | 0.004 | −1693, −995 |
| OsGLP4-1 | Os04g0617900 | 5.22 | 0.000 | −738 |
| Similar to blight-associated protein p12 precursor | Os09g0472900 | 4.91 | 0.000 | −2469, −2376, −178 |
| Similar to receptor-like kinase Xa21-binding protein 3. | Os05g0112000 | 4.86 | 0.016 | −2973 |
| Similar to herbicide safener binding protein | Os11g0306400 | 4.67 | 0.042 | |
| Leucine-rich repeat domain-containing protein | Os04g0122000 | 4.57 | 0.010 | −1725 |
| OsPR8 | Os10g0416500 | 4.50 | 0.000 | −2994, −2939, −871, −726 |
| OsGSL3 | Os01g0754200 | 4.45 | 0.018 | −2773, −2770, −584, 581, −578, −575, 572, −569 |
| OsEXO70B3 | Os05g0473500 | 4.31 | 0.003 | −2788, −2764, −2748, −508, −468, −465, −438, −413, −264, −159 |
| Leucine-rich repeat, typical subtype domain-containing protein | Os11g0568800 | 3.83 | 0.009 | −2429, −2426, −1239, −1210, −1185, −1130 |
| Leucine-rich repeat, N-terminal domain-containing protein | Os12g0211500 | 3.80 | 0.017 | |
| Similar to pathogen-related protein | Os01g0248300 | 3.80 | 0.000 | −2698, −2680, −2644, −2637, −1700, −1620, −1560, −1557, −1534, −1317 |
| OsDXS3 | Os07g0190000 | 3.28 | 0.000 | |
| Multi-antimicrobial extrusion protein MatE family protein | Os09g0524300 | 3.27 | 0.026 | |
| OsLOX9 | Os08g0508800 | 3.20 | 0.000 | −1626 |
| OsWAKL21.2 | Os12g0595800 | 2.93 | 0.016 | −959, −842, −839, −836, −833, −308, −305, −302, −299, −296, −183, −167, −164, −161, −92 |
| Similar to resistance protein candidate (fragment) | Os03g0258000 | 2.84 | 0.031 | −577, −478, −391 |
| IAI2 | Os01g0132000 | 2.78 | 0.000 | −1174, −1167 |
| OsTHI9 | Os06g0514800 | 2.74 | 0.025 | −1718, −1715, −1697 |
| Leucine-rich repeat, N-terminal domain-containing protein | Os01g0891700 | 2.70 | 0.031 | |
| OsPR4d | Os11g0591800 | 2.26 | 0.001 | −2134, −993 |
| Allergen V5/Tpx-1 related family protein | Os07g0126401 | 2.24 | 0.014 | |
| Allergen V5/Tpx-1 related family protein | Os07g0125600 | 2.23 | 0.015 | −553, −386, −336, −333, −215 |
| OsPR1b | Os07g0125000 | 2.22 | 0.015 | |
| OsPR1-73 | Os07g0127600 | 2.15 | 0.018 | |
| Similar to thaumatin-like protein | Os12g0630500 | 2.15 | 0.000 | −2453 |
| PR5 | Os03g0663500 | 2.12 | 0.006 | |
| OsGLN1 | Os01g0946700 | 2.06 | 0.000 | −2549, −1263, −760, −669, −616, −457, −374, −323 |
| OsAOS2 | Os03g0225900 | 2.01 | 0.007 | −2843, −2202, −349 |
| Hormone-associated genes | ||||
| OsLUGL, OsWD40-17 | Os01g0607400 | 11.97 | 0.001 | −2980, −2906, −2903, −2900, −2897, −2836, −2833 |
| OsARF12 | Os04g0671900 | 10.33 | 0.000 | |
| OsJAZ8 | Os09g0401300 | 6.01 | 0.000 | −2541, −2538 |
| OsGA20ox4 | Os05g0421900 | 4.85 | 0.015 | |
| OsARF13, OsARF12 | Os04g0690600 | 2.74 | 0.030 | |
| OsOPR4 | Os06g0215900 | 2.46 | 0.000 | −2863, −1540, −1481, −1478, −1459, −1431, −1428, −1279, −1276, −1238, −1206, −1133, −1129, −1126, −1070, −1040, −1016, −1013 |
| Lignin biosynthesis-associated genes | ||||
| OsUGT707A5 | Os07g0503300 | 4.53 | 0.000 | −1544, −1507, −1470 |
| OsUGT93B | Os04g0556400 | 3.99 | 0.003 | −2364 |
| UDP-glucuronosyl/UDP-glucosyltransferase family protein | Os03g0804900 | 3.23 | 0.027 | −2977, −805, −777, −774, −740, −737, −690, −473, −470, −407, −404, −376 |
| OsLAC17 | Os10g0346300 | 3.02 | 0.010 | −1525, −1481, −1468, −151 |
| OsLAC10 | Os02g0749700 | 2.95 | 0.026 | −1963, −1907 |
| OsCAD8D | Os09g0400400 | 2.94 | 0.000 | −2137 |
| F-box protein | ||||
| OsFbox331 | Os07g0118900 | 8.11 | 0.013 | |
| OsFbox228 | Os04g0571300 | 4.07 | 0.003 | −1080 |
| OsFbox552 | Os10g0396400 | 3.34 | 0.036 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jung, S.E.; Bang, S.W.; Kim, S.H.; Seo, J.S.; Yoon, H.-B.; Kim, Y.S.; Kim, J.-K. Overexpression of OsERF83, a Vascular Tissue-Specific Transcription Factor Gene, Confers Drought Tolerance in Rice. Int. J. Mol. Sci. 2021, 22, 7656. https://doi.org/10.3390/ijms22147656
Jung SE, Bang SW, Kim SH, Seo JS, Yoon H-B, Kim YS, Kim J-K. Overexpression of OsERF83, a Vascular Tissue-Specific Transcription Factor Gene, Confers Drought Tolerance in Rice. International Journal of Molecular Sciences. 2021; 22(14):7656. https://doi.org/10.3390/ijms22147656
Chicago/Turabian StyleJung, Se Eun, Seung Woon Bang, Sung Hwan Kim, Jun Sung Seo, Ho-Bin Yoon, Youn Shic Kim, and Ju-Kon Kim. 2021. "Overexpression of OsERF83, a Vascular Tissue-Specific Transcription Factor Gene, Confers Drought Tolerance in Rice" International Journal of Molecular Sciences 22, no. 14: 7656. https://doi.org/10.3390/ijms22147656
APA StyleJung, S. E., Bang, S. W., Kim, S. H., Seo, J. S., Yoon, H.-B., Kim, Y. S., & Kim, J.-K. (2021). Overexpression of OsERF83, a Vascular Tissue-Specific Transcription Factor Gene, Confers Drought Tolerance in Rice. International Journal of Molecular Sciences, 22(14), 7656. https://doi.org/10.3390/ijms22147656