
Keratin 7 Is a Constituent of the Keratin Network in Mouse Pancreatic Islets and Is Upregulated in Experimental Diabetes

 and
and 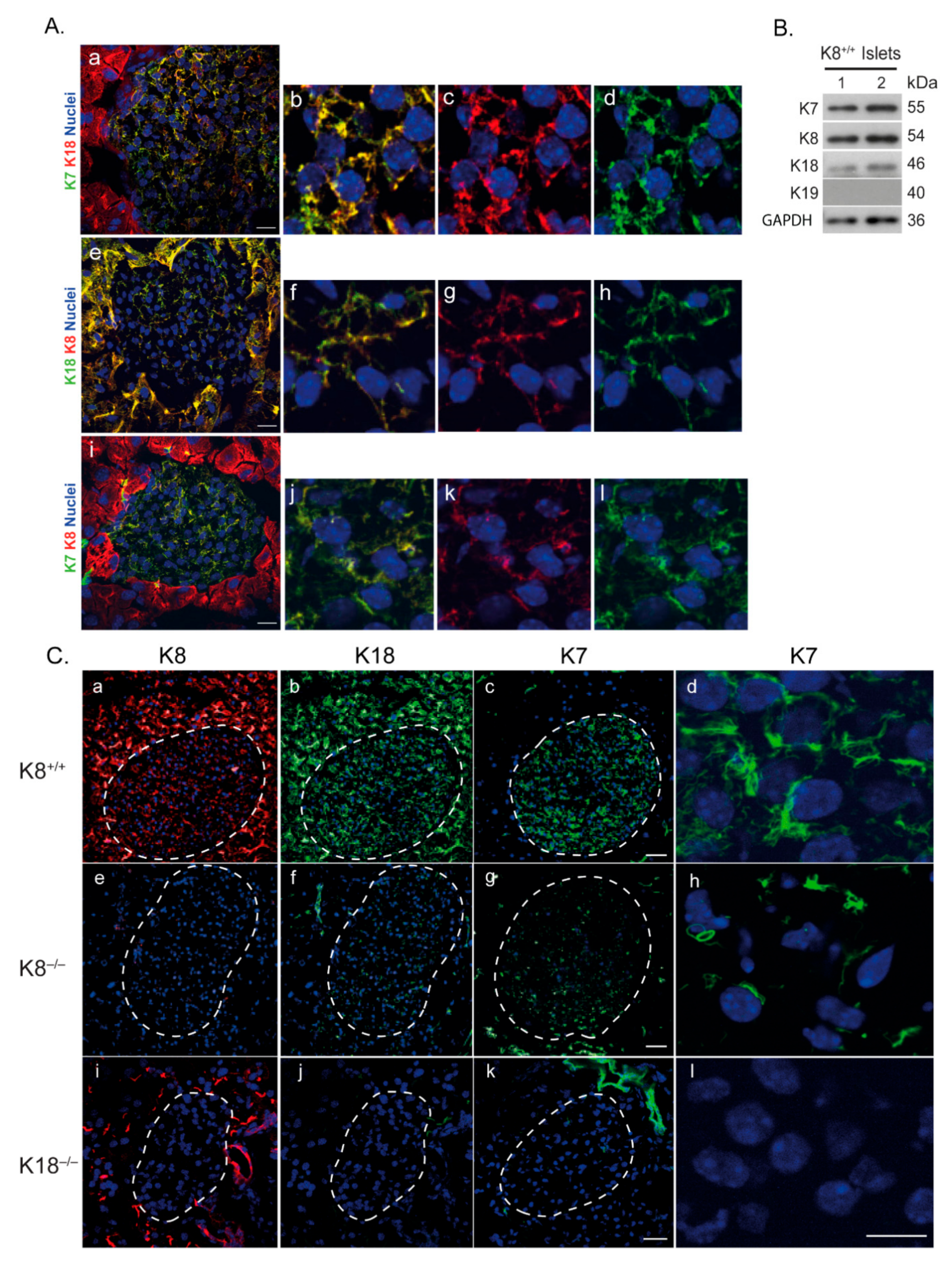{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Type II K7 Is Expressed in Murine Islets In Vivo Together with Type II K8 and Type I K18, and Its Expression Is Dependent on the Presence and Levels of K18
2.2. Keratin Expression in Islets Is Enriched Near the β-Cell Vertices and Lateral Edges
2.3. Over-Expression of Human K18 in Mice Induces K7 Upregulation
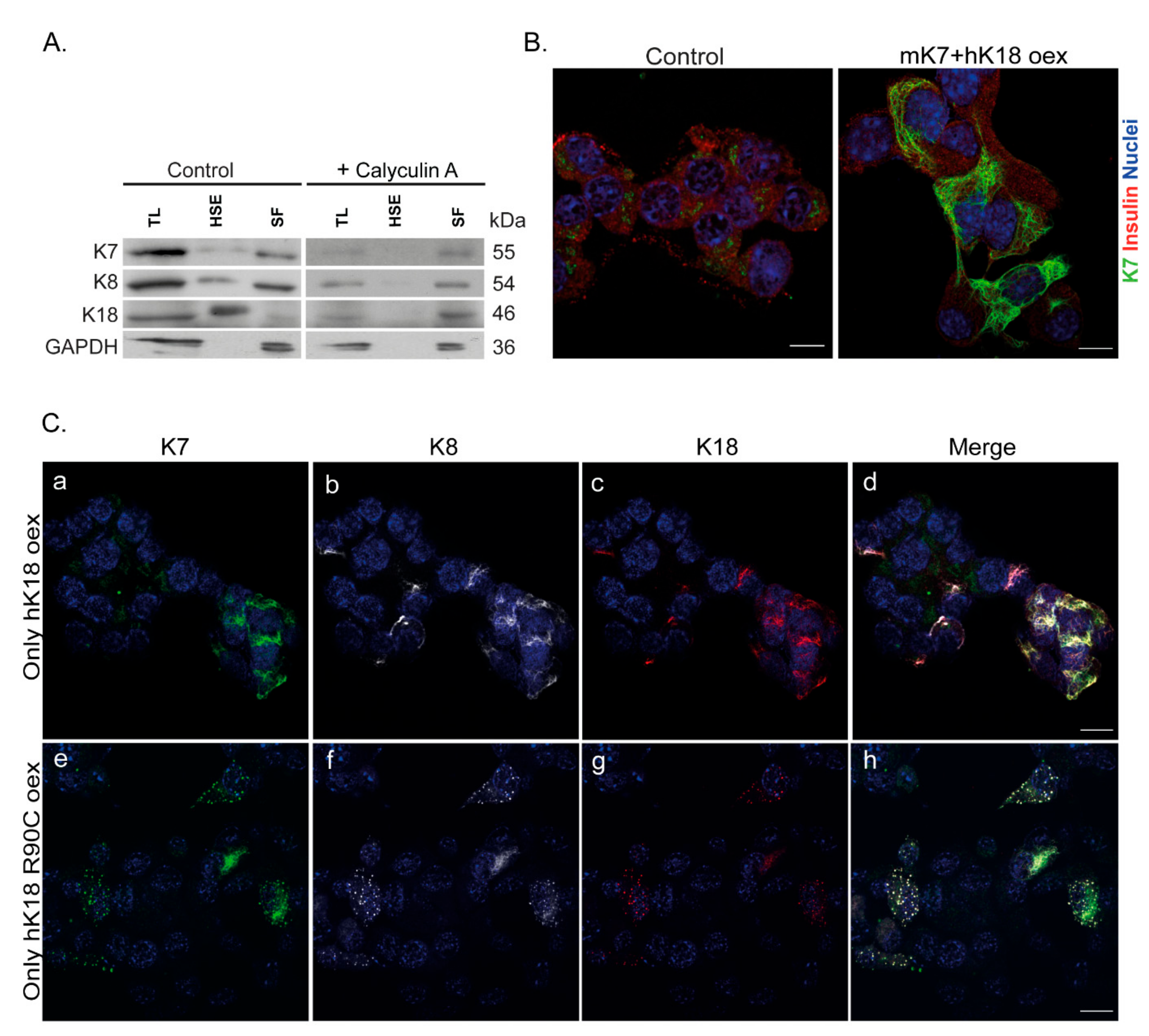
2.4. MIN6 β-Cells Contain Low Levels of Soluble Non-Filamentous K7 and Over-Expression of K18 Restores the K7 Filament Networks
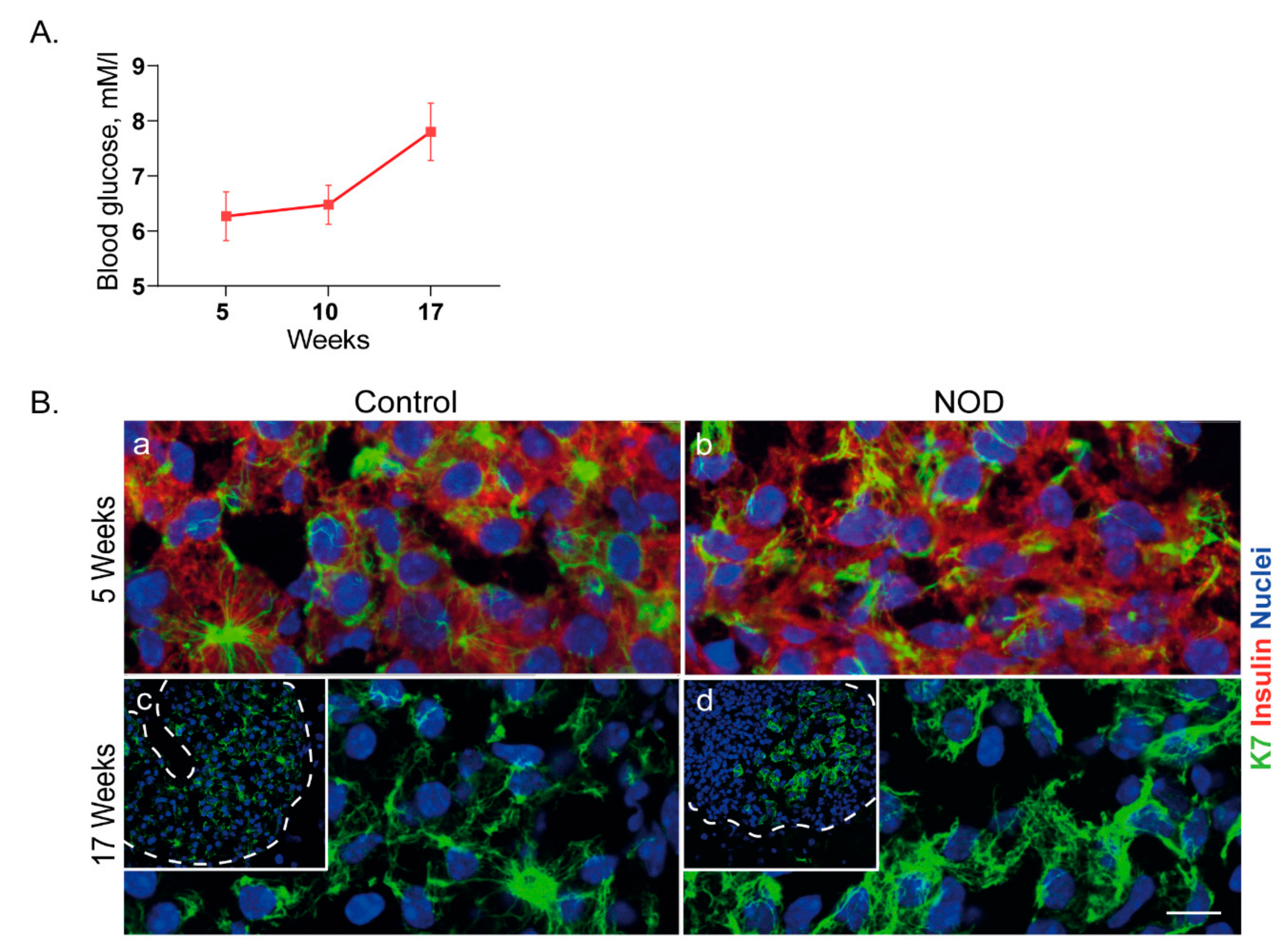
2.5. Islet K7 Becomes Upregulated and Broadly Distributed as Cytoplasmic Filaments in Prediabetic and Diabetic Mice
3. Discussion
4. Materials and Methods
4.1. Experimental Animals and In Vivo Experiments
4.2. Cell Culture and Transfection
4.3. Isolation of Islets of Langerhans
4.4. High-Salt Extraction
4.5. Immunofluorescence Staining and Analysis
4.6. SDS-PAGE and Immunoblotting
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Statistical Analysis
References
- Miettinen, M.; Fetsch, J.F. Distribution of keratins in normal endothelial cells and a spectrum of vascular tumors: Implications in tumor diagnosis. Hum. Pathol. 2000, 31, 1062–1067. [Google Scholar] [CrossRef]
- Ramaekers, F.; Huysmans, A.; Schaart, G.; Moesker, O.; Vooijs, P. Tissue distribution of keratin 7 as monitored by a monoclonal antibody. Exp. Cell Res. 1987, 170, 235–249. [Google Scholar] [CrossRef]
- Smith, F.J.; Porter, R.M.; Corden, L.D.; Lunny, D.P.; Lane, E.B.; McLean, W.H. Cloning of human, murine, and marsupial keratin 7 and a survey of K7 expression in the mouse. Biochem. Biophys. Res. Commun. 2002, 297, 818–827. [Google Scholar] [CrossRef]
- Asghar, M.N.; Silvander, J.S.; Helenius, T.O.; Lahdeniemi, I.A.; Alam, C.; Fortelius, L.E.; Holmsten, R.O.; Toivola, D.M. The amount of keratins matters for stress protection of the colonic epithelium. PLoS ONE 2015, 10, e0127436. [Google Scholar] [CrossRef] [PubMed]
- Bouwens, L. Cytokeratins and cell differentiation in the pancreas. J. Pathol. 1998, 184, 234–239. [Google Scholar] [CrossRef]
- Omary, M.B.; Ku, N.O.; Strnad, P.; Hanada, S. Toward unraveling the complexity of simple epithelial keratins in human disease. J. Clin. Investig. 2009, 119, 1794–1805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Omary, M.B.; Ku, N.O.; Toivola, D.M. Keratins: Guardians of the liver. Hepatology 2002, 35, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Toivola, D.M.; Baribault, H.; Magin, T.; Michie, S.A.; Omary, M.B. Simple epithelial keratins are dispensable for cytoprotection in two pancreatitis models. Am. J. Physiol. Gastrointest. Liver Physiol. 2000, 279, G1343–G1354. [Google Scholar] [CrossRef] [PubMed]
- Zhong, B.; Omary, M.B. Actin overexpression parallels severity of pancreatic injury. Exp. Cell Res. 2004, 299, 404–414. [Google Scholar] [CrossRef] [PubMed]
- Zhong, B.; Zhou, Q.; Toivola, D.M.; Tao, G.Z.; Resurreccion, E.Z.; Omary, M.B. Organ-specific stress induces mouse pancreatic keratin overexpression in association with NF-kappaB activation. J. Cell Sci. 2004, 117, 1709–1719. [Google Scholar] [CrossRef] [Green Version]
- Bouwens, L.; Braet, F.; Heimberg, H. Identification of rat pancreatic duct cells by their expression of cytokeratins 7, 19, and 20 in vivo and after isolation and culture. J. Histochem. Cytochem. 1995, 43, 245–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, K.H.; Quickel, R.R.; Tatarkiewicz, K.; Ulrich, T.R.; Hollister-Lock, J.; Trivedi, N.; Bonner-Weir, S.; Weir, G.C. Differentiation and expansion of beta cell mass in porcine neonatal pancreatic cell clusters transplanted into nude mice. Cell Transplant. 1999, 8, 673–689. [Google Scholar] [CrossRef] [PubMed]
- Alam, C.M.; Silvander, J.S.; Daniel, E.N.; Tao, G.Z.; Kvarnstrom, S.M.; Alam, P.; Omary, M.B.; Hanninen, A.; Toivola, D.M. Keratin 8 modulates beta-cell stress responses and normoglycaemia. J. Cell Sci. 2013. [Google Scholar] [CrossRef] [Green Version]
- Silvander, J.S.G.; Kvarnstrom, S.M.; Kumari-Ilieva, A.; Shrestha, A.; Alam, C.M.; Toivola, D.M. Keratins regulate beta-cell mitochondrial morphology, motility, and homeostasis. FASEB J. 2017, 31, 4578–4587. [Google Scholar] [CrossRef] [Green Version]
- Petyuk, V.A.; Qian, W.J.; Hinault, C.; Gritsenko, M.A.; Singhal, M.; Monroe, M.E.; Camp, D.G., 2nd; Kulkarni, R.N.; Smith, R.D. Characterization of the mouse pancreatic islet proteome and comparative analysis with other mouse tissues. J. Proteome Res. 2008, 7, 3114–3126. [Google Scholar] [CrossRef] [Green Version]
- Gan, W.J.; Zavortink, M.; Ludick, C.; Templin, R.; Webb, R.; Webb, R.; Ma, W.; Poronnik, P.; Parton, R.G.; Gaisano, H.Y.; et al. Cell polarity defines three distinct domains in pancreatic beta-cells. J. Cell Sci. 2017, 130, 143–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geron, E.; Boura-Halfon, S.; Schejter, E.D.; Shilo, B.Z. The Edges of Pancreatic Islet beta Cells Constitute Adhesive and Signaling Microdomains. Cell Rep. 2015, 10, 317–325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alam, C.; Baghestani, S.; Toivola, D.M. Intermediate Filament Keratins in the Exocrine Pancreas. Pancreapedia Exocrine Pancreas Knowl. 2020. [Google Scholar] [CrossRef]
- Polari, L.; Alam, C.M.; Nystrom, J.H.; Heikkila, T.; Tayyab, M.; Baghestani, S.; Toivola, D.M. Keratin intermediate filaments in the colon: Guardians of epithelial homeostasis. Int. J. Biochem. Cell Biol. 2020, 129, 105878. [Google Scholar] [CrossRef] [PubMed]
- Honda, H. The world of epithelial sheets. Dev. Growth Differ. 2017, 59, 306–316. [Google Scholar] [CrossRef] [Green Version]
- Musch, A. The unique polarity phenotype of hepatocytes. Exp. Cell Res. 2014, 328, 276–283. [Google Scholar] [CrossRef] [Green Version]
- Weir, G.C.; Bonner-Weir, S. Islets of Langerhans: The puzzle of intraislet interactions and their relevance to diabetes. J. Clin. Investig. 1990, 85, 983–987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonner-Weir, S. Morphological evidence for pancreatic polarity of beta-cell within islets of Langerhans. Diabetes 1988, 37, 616–621. [Google Scholar] [CrossRef]
- Ku, N.O.; Soetikno, R.M.; Omary, M.B. Keratin mutation in transgenic mice predisposes to Fas but not TNF-induced apoptosis and massive liver injury. Hepatology 2003, 37, 1006–1014. [Google Scholar] [CrossRef] [PubMed]
- Toivola, D.M.; Ku, N.O.; Ghori, N.; Lowe, A.W.; Michie, S.A.; Omary, M.B. Effects of keratin filament disruption on exocrine pancreas-stimulated secretion and susceptibility to injury. Exp. Cell Res. 2000, 255, 156–170. [Google Scholar] [CrossRef]
- Toivola, D.M.; Omary, M.B.; Ku, N.O.; Peltola, O.; Baribault, H.; Eriksson, J.E. Protein phosphatase inhibition in normal and keratin 8/18 assembly-incompetent mouse strains supports a functional role of keratin intermediate filaments in preserving hepatocyte integrity. Hepatology 1998, 28, 116–128. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Toivola, D.M.; Feng, N.; Greenberg, H.B.; Franke, W.W.; Omary, M.B. Keratin 20 helps maintain intermediate filament organization in intestinal epithelia. Mol. Biol. Cell 2003, 14, 2959–2971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ku, N.O.; Michie, S.; Oshima, R.G.; Omary, M.B. Chronic hepatitis, hepatocyte fragility, and increased soluble phosphoglycokeratins in transgenic mice expressing a keratin 18 conserved arginine mutant. J. Cell Biol. 1995, 131, 1303–1314. [Google Scholar] [CrossRef]
- Ku, N.O.; Toivola, D.M.; Zhou, Q.; Tao, G.; Zhong, B.; Omary, M.B. Studying Simple Epithelial Keratins in Cells and Tissues. Methods Cell Biol. 2004, 78, 489–517. [Google Scholar] [CrossRef] [PubMed]
- Toivola, D.M.; Strnad, P.; Habtezion, A.; Omary, M.B. Intermediate filaments take the heat as stress proteins. Trends Cell Biol. 2010, 20, 79–91. [Google Scholar] [CrossRef] [Green Version]
- Ku, N.O.; Omary, M.B. Keratins turn over by ubiquitination in a phosphorylation-modulated fashion. J. Cell Biol. 2000, 149, 547–552. [Google Scholar] [CrossRef]
- Quinlan, R.A.; Cohlberg, J.A.; Schiller, D.L.; Hatzfeld, M.; Franke, W.W. Heterotypic tetramer (A2D2) complexes of non-epidermal keratins isolated from cytoskeletons of rat hepatocytes and hepatoma cells. J. Mol. Biol. 1984, 178, 365–388. [Google Scholar] [CrossRef]
- Magin, T.M.; Schroder, R.; Leitgeb, S.; Wanninger, F.; Zatloukal, K.; Grund, C.; Melton, D.W. Lessons from keratin 18 knockout mice: Formation of novel keratin filaments, secondary loss of keratin 7 and accumulation of liver-specific keratin 8-positive aggregates. J. Cell Biol. 1998, 140, 1441–1451. [Google Scholar] [CrossRef]
- Toivola, D.M.; Nakamichi, I.; Strnad, P.; Michie, S.A.; Ghori, N.; Harada, M.; Zeh, K.; Oshima, R.G.; Baribault, H.; Omary, M.B. Keratin overexpression levels correlate with the extent of spontaneous pancreatic injury. Am. J. Pathol. 2008, 172, 882–892. [Google Scholar] [CrossRef] [Green Version]
- Giudice, G.J.; Fuchs, E. The transfection of epidermal keratin genes into fibroblasts and simple epithelial cells: Evidence for inducing a type I keratin by a type II gene. Cell 1987, 48, 453–463. [Google Scholar] [CrossRef]
- Sandilands, A.; Smith, F.J.; Lunny, D.P.; Campbell, L.E.; Davidson, K.M.; MacCallum, S.F.; Corden, L.D.; Christie, L.; Fleming, S.; Lane, E.B.; et al. Generation and characterisation of keratin 7 (K7) knockout mice. PLoS ONE 2013, 8, e64404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, M.G.; Maheshwari, R.R.; Anderson, S.J.; Berlinguer-Palmini, R.; Jones, C.; Richardson, S.J.; Rotti, P.G.; Armour, S.L.; Ding, Y.; Krasnogor, N.; et al. In Situ Analysis Reveals That CFTR Is Expressed in Only a Small Minority of beta-Cells in Normal Adult Human Pancreas. J. Clin. Endocrinol. Metab. 2020, 105. [Google Scholar] [CrossRef] [PubMed]
- An, Q.; Liu, T.; Wang, M.Y.; Yang, Y.J.; Zhang, Z.D.; Liu, Z.J.; Yang, B. KRT7 promotes epithelialmesenchymal transition in ovarian cancer via the TGFbeta/Smad2/3 signaling pathway. Oncol. Rep. 2021, 45, 481–492. [Google Scholar] [CrossRef]
- Jiang, F.X.; Georges-Labouesse, E.; Harrison, L.C. Regulation of laminin 1-induced pancreatic beta-cell differentiation by alpha6 integrin and alpha-dystroglycan. Mol. Med. 2001, 7, 107–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salas, P.J.; Forteza, R.; Mashukova, A. Multiple roles for keratin intermediate filaments in the regulation of epithelial barrier function and apico-basal polarity. Tissue Barriers 2016, 4, e1178368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moll, R.; Franke, W.W.; Schiller, D.L.; Geiger, B.; Krepler, R. The catalog of human cytokeratins: Patterns of expression in normal epithelia, tumors and cultured cells. Cell 1982, 31, 11–24. [Google Scholar] [CrossRef]
- Moll, R.; Divo, M.; Langbein, L. The human keratins: Biology and pathology. Histochem. Cell Biol. 2008, 129, 705–733. [Google Scholar] [CrossRef] [Green Version]
- Djudjaj, S.; Papasotiriou, M.; Bulow, R.D.; Wagnerova, A.; Lindenmeyer, M.T.; Cohen, C.D.; Strnad, P.; Goumenos, D.S.; Floege, J.; Boor, P. Keratins are novel markers of renal epithelial cell injury. Kidney Int. 2016, 89, 792–808. [Google Scholar] [CrossRef] [Green Version]
- Guldiken, N.; Kobazi Ensari, G.; Lahiri, P.; Couchy, G.; Preisinger, C.; Liedtke, C.; Zimmermann, H.W.; Ziol, M.; Boor, P.; Zucman-Rossi, J.; et al. Keratin 23 is a stress-inducible marker of mouse and human ductular reaction in liver disease. J. Hepatol. 2016, 65, 552–559. [Google Scholar] [CrossRef] [PubMed]
- Helenius, T.O.; Antman, C.A.; Asghar, M.N.; Nystrom, J.H.; Toivola, D.M. Keratins Are Altered in Intestinal Disease-Related Stress Responses. Cells 2016, 5, 35. [Google Scholar] [CrossRef] [Green Version]
- Baribault, H.; Penner, J.; Iozzo, R.V.; Wilson-Heiner, M. Colorectal hyperplasia and inflammation in keratin 8-deficient FVB/N mice. Genes Dev. 1994, 8, 2964–2973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zmuda, E.J.; Powell, C.A.; Hai, T. A method for murine islet isolation and subcapsular kidney transplantation. J. Vis. Exp. 2011. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alam, C.M.; Baghestani, S.; Pajari, A.; Omary, M.B.; Toivola, D.M. Keratin 7 Is a Constituent of the Keratin Network in Mouse Pancreatic Islets and Is Upregulated in Experimental Diabetes. Int. J. Mol. Sci. 2021, 22, 7784. https://doi.org/10.3390/ijms22157784
Alam CM, Baghestani S, Pajari A, Omary MB, Toivola DM. Keratin 7 Is a Constituent of the Keratin Network in Mouse Pancreatic Islets and Is Upregulated in Experimental Diabetes. International Journal of Molecular Sciences. 2021; 22(15):7784. https://doi.org/10.3390/ijms22157784
Chicago/Turabian StyleAlam, Catharina M., Sarah Baghestani, Ada Pajari, M. Bishr Omary, and Diana M. Toivola. 2021. "Keratin 7 Is a Constituent of the Keratin Network in Mouse Pancreatic Islets and Is Upregulated in Experimental Diabetes" International Journal of Molecular Sciences 22, no. 15: 7784. https://doi.org/10.3390/ijms22157784
APA StyleAlam, C. M., Baghestani, S., Pajari, A., Omary, M. B., & Toivola, D. M. (2021). Keratin 7 Is a Constituent of the Keratin Network in Mouse Pancreatic Islets and Is Upregulated in Experimental Diabetes. International Journal of Molecular Sciences, 22(15), 7784. https://doi.org/10.3390/ijms22157784