
Directed Recovery and Molecular Characterization of Antibiotic Resistance Plasmids from Cheese Bacteria




Abstract
:1. Introduction
2. Results
2.1. Diversity of Plasmid Profiles
2.2. Screening of Plasmids Providing Antibiotic Resistance
2.3. Antibiotic Susceptibility of Plasmid-Harboring Transformants
2.4. Detection of Tetracycline and Erythromycin Resistance Genes
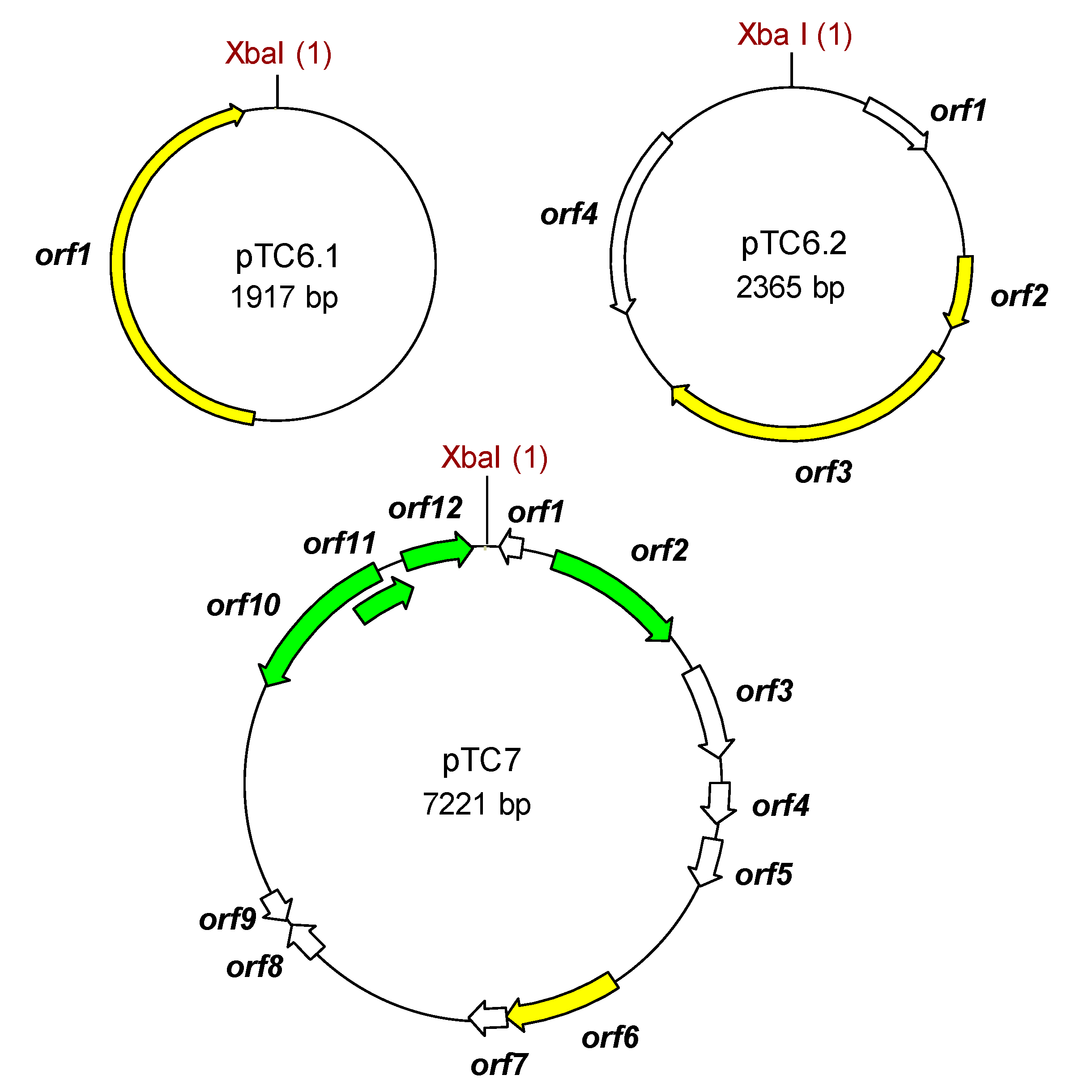
2.5. Sequencing, Assembly and Annotation of Plasmids
2.6. Tetracycline Resistance Plasmids
2.7. Erythromycin Resistance Plasmids
2.8. Nucleotide Sequence of tet(S) and Its Flanking Regions
3. Discussion
4. Materials and Methods
4.1. Lactic Acid Bacteria Strains and Culture Conditions
4.2. Cheese Sampling
4.3. Plasmid Isolation and Transformation
4.4. Minimum Inhibitory Concentrations of Antibiotics
4.5. PCR Detection of Tetracycline and Erythromycin Resistance Determinants
4.6. Whole Plasmid DNA Sequencing, Assembly and Annotation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- WHO. Antimicrobial Resistance: Global Report on Surveillance. 2014. Available online: https://www.who.int/drugresistance/documents/surveillancereport/en/ (accessed on 6 December 2019).
- Martínez, J.L. Bottlenecks in the transferability of antibiotic resistance from natural ecosystems to human bacterial pathogens. Front. Microbiol. 2012, 2. [Google Scholar] [CrossRef] [Green Version]
- Walsh, T.R. A one-health approach to antimicrobial resistance. Nat. Microbiol. 2018, 3, 854–855. [Google Scholar] [CrossRef]
- Fletcher, S. Understanding the contribution of environmental factors in the spread of antimicrobial resistance. Environ. Health Prev. Med. 2015, 20, 243–252. [Google Scholar] [CrossRef] [Green Version]
- Prestinaci, F.; Pezzotti, P.; Pantosti, A. Antimicrobial resistance: A global multifaceted phenomenon. Pathog. Glob. Health 2015, 109, 309–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO. Global Action Plan on Antimicrobial Resistance. 2015. Available online: https://apps.who.int/iris/bitstream/handle/10665/193736/9789241509763_eng.pdf?sequence=1&isAllowed=y (accessed on 20 January 2021).
- Collignon, P. The importance of a One Health approach to preventing the development and spread of antibiotic resistance. Curr. Top. Microbiol. Immunol. 2013, 366, 19–36. [Google Scholar] [CrossRef] [PubMed]
- Shintani, M.; Sanchez, Z.K.; Kimbara, K. Genomics of microbial plasmids: Classification and identification based on replication and transfer systems and host taxonomy. Front. Microbiol. 2015, 6, 242. [Google Scholar] [CrossRef] [PubMed]
- San Millán, A.; MacLean, R.C. Fitness costs of plasmids: A limit to plasmid transmission. Microbiol. Spectr. 2017, 5. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Beltrán, J.; Dela Fuente, J.; León-Sampedro, R.; MacLean, R.C.; San Millán, Á. Beyond horizontal gene transfer: The role of plasmids in bacterial evolution. Nat. Rev. Microbiol. 2021, 19, 1–13. [Google Scholar] [CrossRef]
- Kelleher, P.; Mahony, J.; Bottacini, F.; Lugli, G.A.; Ventura, M.; Van Sinderen, D. The Lactococcus lactis pan-plasmidome. Front. Microbiol. 2019, 10, 707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanza, V.F.; Tedim, A.P.; Luís Martínez, J.; Baquero, F.; Coque, T.M. The plasmidome of Firmicutes: Impact on the emergence and the spread of resistance to antimicrobials. Microbiol. Spectr. 2015, 3. [Google Scholar] [CrossRef] [Green Version]
- Makarova, K.; Slesarev, A.; Wolf, Y.; Sorokin, A.; Mirkin, B.; Koonin, E.; Pavlov, A.; Pavlova, N.; Karamychev, V.; Polouchine, M.; et al. Comparative genomics of the lactic acid bacteria. Proc. Natl. Acad. Sci. USA 2006, 103, 15611–15616. [Google Scholar] [CrossRef] [Green Version]
- Sørensen, S.J.; Bailey, M.; Hansen, L.H.; Kroer, N.; Wuertz, S. Studying plasmid horizontal transfer in situ: A critical review. Nat. Rev. Microbiol. 2005, 3, 700–710. [Google Scholar] [CrossRef]
- Martínez, J.L.; Coque, T.M.; Baquero, F. What is a resistance gene? Ranking risk in resistomes. Nat. Rev. Microbiol. 2015, 13, 116–123. [Google Scholar] [CrossRef]
- Perry, J.A.; Wright, G.D.; Roberts, M.C.; Aminov, R.I.; Degroote, M.G. The antibiotic resistance “mobilome”: Searching for the link between environment and clinic. Front. Microbiol. 2013, 4, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.B.; Arnipalli, S.R.; Ziouzenkova, O. Antibiotics in food chain: The consequences for antibiotic resistance. Antibiotics 2020, 9, 688. [Google Scholar] [CrossRef] [PubMed]
- Florez-Cuadrado, D.; Moreno, M.A.; Ugarte-Ruíz, M.; Domínguez, L. Antimicrobial resistance in the food chain in the European Union. Adv. Food Nutr. Res. 2018, 86, 115–136. [Google Scholar] [CrossRef]
- Capita, R.; Alonso-Calleja, C. Antibiotic-resistant bacteria: A challenge for the food industry. Crit. Rev. Food Sci. Nutr. 2013, 53, 11–48. [Google Scholar] [CrossRef]
- Soares-Santos, V.; Barreto, A.S.; Semedo-Lemsaddek, T. Characterization of enterococci from food and food-related settings. J. Food Prot. 2015, 78, 1320–1326. [Google Scholar] [CrossRef]
- Flórez, A.B.; Alegría, A.; Rossi, F.; Delgado, S.; Felis, G.E.; Torriani, S.; Mayo, B. Molecular identification and quantification of tetracycline and erythromycin resistance genes in Spanish and Italian retail cheeses. BioMed Res. Int. 2014, 2014. [Google Scholar] [CrossRef] [Green Version]
- Flórez, A.B.; Vázquez, L.; Mayo, B. A functional metagenomic analysis of tetracycline resistance in cheese bacteria. Front. Microbiol. 2017, 8, 907. [Google Scholar] [CrossRef]
- Flórez, A.B.; Campedelli, I.; Delgado, S.; Alegría, Á.; Salvetti, E.; Felis, G.E.; Mayo, B.; Torriani, S. Antibiotic susceptibility profiles of dairy Leuconostoc, analysis of the genetic basis of atypical resistances and transfer of genes in vitro and in a food matrix. PLoS ONE 2016, 11, e0145203. [Google Scholar] [CrossRef] [Green Version]
- Flórez, A.B.; Ammor, M.S.; Mayo, B. Identification of tet(M) in two Lactococcus lactis strains isolated from a Spanish traditional starter-free cheese made of raw milk and conjugative transfer of tetracycline resistance to lactococci and enterococci. Int. J. Food Microbiol. 2008, 121, 189–194. [Google Scholar] [CrossRef]
- Madec, J.Y.; Haenni, M. Antimicrobial resistance plasmid reservoir in food and food-producing animals. Plasmid 2018, 99, 72–81. [Google Scholar] [CrossRef]
- McMillan, E.A.; Gupta, S.K.; Williams, L.E.; Jové, T.; Hiott, L.M.; Woodley, T.A.; Barrett, J.B.; Jackson, C.R.; Wasilenko, J.L.; Simmons, M.; et al. Antimicrobial resistance genes, cassettes, and plasmids present in Salmonella enterica associated with United States food animals. Front. Microbiol. 2019, 10, 832. [Google Scholar] [CrossRef] [PubMed]
- Tarazanova, M.; Beerthuyzen, M.; Siezen, R.; Fernandez-Gutierrez, M.M.; de Jong, A.; van der Meulen, S.; Kok, J.; Bachmann, H. Plasmid complement of Lactococcus lactis NCDO712 reveals a novel pilus gene cluster. PLoS ONE 2016, 11, e0167970. [Google Scholar] [CrossRef] [Green Version]
- Ainsworth, S.; Stockdale, S.; Bottacini, F.; Mahony, J.; van Sinderen, D. The Lactococcus lactis plasmidome: Much learnt, yet still lots to discover. FEMS Microbiol. Rev. 2014, 38, 1066–1088. [Google Scholar] [CrossRef] [Green Version]
- Walsh, A.M.; Macori, G.; Kilcawley, K.N.; and Cotter, P.D. Meta-analysis of cheese microbiomes highlights contributions to multiple aspects of quality. Nat. Food 2020, 1, 500–510. [Google Scholar] [CrossRef]
- Peterson, E.; Kaur, P. Antibiotic resistance mechanisms in bacteria: Relationships between resistance determinants of antibiotic producers, environmental bacteria, and clinical pathogens. Front. Microbiol. 2018, 9, 2928. [Google Scholar] [CrossRef] [PubMed]
- Flórez, A.B.; Mayo, B. Antibiotic resistance-susceptibility profiles of Streptococcus thermophilus isolated from raw milk and genome analysis of the genetic basis of acquired resistances. Front. Microbiol. 2017, 8, 2608. [Google Scholar] [CrossRef] [PubMed]
- Ammor, M.S.; Flórez, A.B.; van Hoek, A.H.; de Los Reyes-Gavilán, C.G.; Aarts, H.J.; Margolles, A.; Mayo, B. Molecular characterization of intrinsic and acquired antibiotic resistance in lactic acid bacteria and bifidobacteria. J. Mol. Microbiol. Biotechnol. 2008, 14, 6–15. [Google Scholar] [CrossRef]
- Maki, T.; Santos, M.D.; Kondo, H.; Hirono, I.; Aoki, T. A transferable 20-kilobase multiple drug resistance-conferring R plasmid (pKL0018) from a fish pathogen (Lactococcus garvieae) is highly homologous to a conjugative multiple drug resistance-conferring enterococcal plasmid. Appl. Environ. Microbiol. 2009, 75, 3370–3372. [Google Scholar] [CrossRef] [Green Version]
- Raha, A.R.; Ross, E.; Yusoff, K.; Manap, M.Y.; Ideris, A. Characterisation and molecular cloning of an erythromycin resistance plasmid of Lactococcus lactis isolated from chicken cecum. J. Biochem. Mol. Biol. Biophys. 2020, 6, 7–11. [Google Scholar] [CrossRef]
- Biggel, M.; Zurfluh, K.; Treier, A.; Nüesch-Inderbinen, M.; Stephan, R. Characteristics of fosA-carrying plasmids in E. coli and Klebsiella spp. isolates originating from food and environmental samples. J. Antimicrob. Chemother. 2021, 76, 2004–2011. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, Y.; Schwarz, S.; Li, Y.; Shen, Z.; Zhang, Q.; Wu, C.; Shen, J. Transferable multiresistance plasmids carrying cfr in Enterococcus spp. from swine and farm environment. Antimicrob. Agents Chemother. 2013, 57, 42–48. [Google Scholar] [CrossRef] [Green Version]
- Loftie-Eaton, W.; Crabtree, A.; Perry, D.; Millstein, J.; Baytosh, J.; Stalder, T.; Robison, B.D.; Forney, L.J.; Top, E.M. Contagious antibiotic resistance: Plasmid transfer among bacterial residents of the zebrafish gut. Appl. Environ. Microbiol. 2021, 87. [Google Scholar] [CrossRef] [PubMed]
- Chaslus-Dancla, E.; Gerbaud, G.; Lagorce, M.; Lafont, J.P.; Courvalin, P. Persistence of an antibiotic resistance plasmid in intestinal Escherichia coli of chickens in the absence of selective pressure. Antimicrob. Agents Chemother. 1987, 31, 784–788. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, W.; Yoshida, S.; Izumi, K.; Koide, S.; Soga, E.; Takizawa, S.; Arakawa, Y.; Nagano, Y.; Nagano, N. Genomic characterisation and epidemiology of nosocomial Serratia marcescens isolates resistant to ceftazidime and their plasmids mediating rare blaTEM-61. J. Glob. Antimicrob. Resist. 2021, 25, 124–131. [Google Scholar] [CrossRef]
- Pietsch, M.; Pfeifer, Y.; Fuchs, S.; Werner, G. Genome-based analyses of fitness effects and compensatory changes associated with acquisition of bla CMY¯, bla CTX-M¯, and bla OXA-48/VIM-1¯containing plasmids in Escherichia coli. Antibiotics 2021, 10, 90. [Google Scholar] [CrossRef] [PubMed]
- Tyson, G.H.; Sabo, J.L.; Hoffmann, M.; Hsu, C.H.; Mukherjee, S.; Hernandez, J.; Tillman, G.; Wasilenko, J.L.; Haro, J.; Simmons, M.; et al. Novel linezolid resistance plasmids in Enterococcus from food animals in the USA. J. Antimicrob. Chemother. 2018, 73, 3254–3258. [Google Scholar] [CrossRef]
- ESVAC. Sales of Veterinary Antimicrobial Agents in 31 European Countries in 2018. Tenth ESVAC Report. 2020. Available online: https://www.ema.europa.eu/en/documents/report/sales-veterinary-antimicrobial-agents-31-european-countries-2018-trends-2010-2018-tenth-esvac-report_en.pdf (accessed on 15 December 2020).
- Devirgiliis, C.; Caravelli, A.; Coppola, D.; Barile, S.; Perozzi, G. Antibiotic resistance and microbial composition along the manufacturing process of Mozzarella di Bufala Campana. Int. J. Food Microbiol. 2008, 128, 378–384. [Google Scholar] [CrossRef]
- Ammor, M.S.; Flórez, A.B.; Mayo, B. Antibiotic resistance in non-enterococcal lactic acid bacteria and bifidobacteria. Food Microbiol. 2007, 24. [Google Scholar] [CrossRef]
- Van Hoek, A.H.A.M.; Mevius, D.; Guerra, B.; Mullany, P.; Roberts, A.P.; Aarts, H.J.M. Acquired antibiotic resistance genes: An overview. Front. Microbiol. 2011, 2, 203. [Google Scholar] [CrossRef] [Green Version]
- Rao Thumu, S.C.; Halami, P.M. Presence of erythromycin and tetracycline resistance genes in lactic acid bacteria from fermented foods of Indian origin. Antonie Leeuwenhoek 2012, 102, 541–551. [Google Scholar] [CrossRef]
- Pinto, U.M.; Pappas, K.M.; Winans, S.C. The ABCs of plasmid replication and segregation. Nat. Rev. Microbiol. 2021, 10, 755–765. [Google Scholar] [CrossRef] [PubMed]
- Sørvig, E.; Skaugen, M.; Naterstad, K.; Eijsink, V.G.H.; Axelsson, L. Plasmid p256 from Lactobacillus plantarum represents a new type of replicon in lactic acid bacteria, and contains a toxin-antitoxin-like plasmid maintenance system. Microbiology 2005, 151, 421–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Mastrigt, O.; Di Stefano, E.; Hartono, S.; Abee, T.; Smid, E.J. Large plasmidome of dairy Lactococcus lactis subsp. lactis biovar diacetylactis FM03P encodes technological functions and appears highly unstable. BMC Genom. 2018, 19, 620. [Google Scholar] [CrossRef]
- Siezen, R.J.; Renckens, B.; Van Swam, I.; Peters, S.; Van Kranenburg, R.; Kleerebezem, M.; De Vos, W.M. Complete sequences of four plasmids of Lactococcus lactis subsp. cremoris SK11 reveal extensive adaptation to the dairy environment. Appl. Environ. Microbiol. 2005, 71, 8371–8382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seegers, J.F.M.L.; Bron, S.; Franke, C.M.; Venema, G.; Kiewiet, R. The majority of lactococcal plasmids carry a highly related replicon. Microbiology 1994, 140, 1291–1300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Del Solar, G.; Giraldo, R.; Ruiz-Echevarría, M.J.; Espinosa, M.; Díaz-Orejas, R. Replication and control of circular bacterial plasmids. Microbiol. Mol. Biol. Rev. 1998, 62, 434–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flórez, A.B.; Mayo, B. Diversity and dynamics of antibiotic-resistant bacteria in cheese as determined by PCR denaturing gradient gel electrophoresis. Int. J. Food Microbiol. 2015, 214, 63–69. [Google Scholar] [CrossRef] [Green Version]
- Alexa Oniciuc, E.A.; Walsh, C.J.; Coughlan, L.M.; Awad, A.; Simon, C.A.; Ruiz, L.; Crispie, F.; Cotter, P.D.; Alvarez-Ordóñez, A. Dairy products and dairy-processing environments as a reservoir of antibiotic resistance and quorum-quenching determinants as revealed through functional metagenomics. mSystems 2020, 5, e00723-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russo, N.; Caggia, C.; Pino, A.; Coque, T.M.; Arioli, S.; Randazzo, C.L. Enterococcus spp. in Ragusano PDO and Pecorino Siciliano cheese types: A snapshot of their antibiotic resistance distribution. Food Chem. Toxicol. 2018, 120, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, M.X.; Silva, N.C.C.; Trevilin, J.H.; Cruzado, M.M.B.; Mui, T.S.; Duarte, F.R.S.; Castillo, C.J.C.; Canniatti-Brazaca, S.G.; Porto, E. Molecular characterization and antibiotic resistance of Staphylococcus spp. isolated from cheese processing plants. J. Dairy Sci. 2017, 100, 5167–5175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelly, W.J.; Ward, L.J.H.; Leahy, S.C. Chromosomal diversity in Lactococcus lactis and the origin of dairy starter cultures. Genome Biol. Evol. 2010, 2, 729–744. [Google Scholar] [CrossRef] [Green Version]
- Porse, A.; Schønning, K.; Munck, C.; Sommer, M.O.A. Survival and evolution of a large multidrug resistance plasmid in new clinical bacterial hosts. Mol. Biol. Evol. 2016, 33, 2860–2873. [Google Scholar] [CrossRef] [PubMed]
- Flórez, A.B.; Mayo, B. The plasmid complement of the cheese isolate Lactococcus garvieae IPLA 31405 revealed adaptation to the dairy environment. PLoS ONE 2015, 10, e0126101. [Google Scholar] [CrossRef] [PubMed]
- Van Kranenburg, R.; Golic, N.; Bongers, R.; Leer, R.J.; De Vos, W.M.; Siezen, R.J.; Kleerebezem, M. Functional analysis of three plasmids from Lactobacillus plantarum. Appl. Environ. Microbiol. 2005, 71, 1223–1230. [Google Scholar] [CrossRef] [Green Version]
- Erkus, O.; De Jager, V.C.L.; Spus, M.; Van Alen-Boerrigter, I.J.; Van Rijswijck, I.M.H.; Hazelwood, L.; Janssen, P.W.M.; Van Hijum, S.A.F.T.; Kleerebezem, M.; Smid, E.J. Multifactorial diversity sustains microbial community stability. ISME J. 2013, 7, 2126–2136. [Google Scholar] [CrossRef]
- Lee, Y.; Nguyen, T.L.; Kim, A.; Kim, N.; Roh, H.J.; Han, H.J.; Jung, S.H.; Cho, M.Y.; Kang, H.Y.; Kim, D.H. Complete genome sequence of multiple-antibiotic-resistant Streptococcus parauberis strain SPOF3K, isolated from diseased olive flounder (Paralichthys olivaceus). Genome Announc. 2018, 6. [Google Scholar] [CrossRef] [Green Version]
- Teuber, M.; Meile, L.; Schwarz, F. Acquired antibiotic resistance in lactic acid bacteria from food. Antonie Leeuwenhoek 1999, 76, 115–137. [Google Scholar] [CrossRef]
- Chanchaithong, P.; Perreten, V.; Schwendener, S. Macrococcus canis contains recombinogenic methicillin resistance elements and the mecB plasmid found in Staphylococcus aureus. J. Antimicrob. Chemother. 2019, 74, 2531–2536. [Google Scholar] [CrossRef] [PubMed]
- Ferrand, A.; Vergalli, J.; Pagès, J.M.; Davin-Regli, A. An intertwined network of regulation controls membrane permeability including drug influx and efflux in Enterobacteriaceae. Microorganisms 2020, 8, 833. [Google Scholar] [CrossRef]
- Putman, M.; van Veen, H.W.; Konings, W.N. Molecular properties of bacterial multidrug transporters. Microbiol. Mol. Biol. Rev. 2000, 64, 672–693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nawaz, M.; Wang, J.; Zhou, A.; Ma, C.; Wu, X.; Moore, J.E.; Millar, B.C.; Xu, J. Characterization and transfer of antibiotic resistance in lactic acid bacteria from fermented food products. Curr. Microbiol. 2011, 62, 1081–1089. [Google Scholar] [CrossRef]
- Schwarz, F.V.; Perreten, V.; Teuber, M. Sequence of the 50-kb conjugative multiresistance plasmid pRE25 from Enterococcus faecalis RE25. Plasmid 2001, 46, 170–187. [Google Scholar] [CrossRef]
- Chantziaras, I.; Boyen, F.; Callens, B.; Dewulf, J. Correlation between veterinary antimicrobial use and antimicrobial resistance in food-producing animals: A report on seven countries. J. Antimicrob. Chemother. 2014, 69, 827–834. [Google Scholar] [CrossRef] [Green Version]
- Yazdankhah, S.; Rudi, K.; Bernhoft, A. Zinc and copper in animal feed-development of resistance and co-resistance to antimicrobial agents in bacteria of animal origin. Microb. Ecol. Health Dis. 2014, 25, 25862. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Fuentes, M.A.; Ortega Morente, E.; Abriouel, H.; Pérez Pulido, R.; Gálvez, A. Isolation and identification of bacteria from organic foods: Sensitivity to biocides and antibiotics. Food Control 2012, 26, 73–78. [Google Scholar] [CrossRef]
- Amachawadi, R.G.; Shelton, N.W.; Shi, X.; Vinasco, J.; Dritz, S.S.; Tokach, M.D.; Nelssen, J.L.; Scott, H.M.; Nagaraja, T.G. Selection of fecal enterococci exhibiting tcrB-mediated copper resistance in pigs fed diets supplemented with copper. Appl. Environ. Microbiol. 2011, 77, 5597–5603. [Google Scholar] [CrossRef] [Green Version]
- Hasman, H.; Aarestrup, F.M. Relationship between copper, glycopeptide, and macrolide resistance among Enterococcus faecium strains isolated from pigs in Denmark between 1997 and 2003. Antimicrob. Agents Chemother. 2005, 49, 454–456. [Google Scholar] [CrossRef] [Green Version]
- Sachi, S.; Ferdous, J.; Sikder, M.H.; Azizul Karim Hussani, S.M. Antibiotic residues in milk: Past, present, and future. J. Adv. Vet. Anim. Res. 2019, 6, 315–332. [Google Scholar] [CrossRef] [PubMed]
- Zwierzchowski, G.; Ametaj, B.N. Minerals and heavy metals in the whole raw milk of dairy cows from different management systems and countries of origin: A meta-analytical study. J. Agric. Food Chem. 2018, 66, 6877–6888. [Google Scholar] [CrossRef] [PubMed]
- Schwaiger, K.; Harms, K.S.; Bischoff, M.; Preikschat, P.; Mölle, G.; Bauer-Unkauf, I.; Lindorfer, S.; Thalhammer, S.; Bauer, J.; Hölzel, C.S. Insusceptibility to disinfectants in bacteria from animals, food and humans-is there a link to antimicrobial resistance? Front. Microbiol. 2014, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Díaz-Orejas, R.; Espinosa, M.; Yeo, C.C. The importance of the expendable: Toxin-antitoxin genes in plasmids and chromosomes. Front. Microbiol. 2017, 8, 1479. [Google Scholar] [CrossRef] [PubMed]
- Gama, J.A.; Zilhão, R.; Dionisio, F. Impact of plasmid interactions with the chromosome and other plasmids on the spread of antibiotic resistance. Plasmid 2018, 99, 82–88. [Google Scholar] [CrossRef]
- Carroll, A.C.; Wong, A. Plasmid persistence: Costs, benefits, and the plasmid paradox. Can. J. Microbiol. 2018, 64, 293–304. [Google Scholar] [CrossRef]
- Holo, H.; Nes, I.F. High-frequency transformation, by electroporation, of Lactococcus lactis subsp. cremoris grown with glycine in osmotically stabilized media. Appl. Environ. Microbiol. 1989, 55, 3119–3123. [Google Scholar] [CrossRef] [Green Version]
- O’Sullivan, D.J.; Klaenhammer, T.R. Rapid mini-prep isolation of high-quality plasmid DNA from Lactococcus and Lactobacillus spp. Appl. Environ. Microbiol. 1993, 59, 2730–2733. [Google Scholar] [CrossRef] [Green Version]
- Clermont, D.; Chesneau, O.; De Cespédès, G.; Horaud, T. New tetracycline resistance determinants coding for ribosomal protection in streptococci and nucleotide sequence of tet(T) isolated from Streptococcus pyogenes A498. Antimicrob. Agents Chemother. 1997, 41, 112–116. [Google Scholar] [CrossRef] [Green Version]
- Barbosa, T.M.; Scott, K.P.; Flint, H.J. Evidence for recent intergeneric transfer of a new tetracycline resistance gene, tet(W), isolated from Butyrivibrio fibrisolvens, and the occurrence of tet(O) in ruminai bacteria. Environ. Microbiol. 1999, 1, 53–64. [Google Scholar] [CrossRef]
- Scott, K.P.; Melville, C.M.; Barbosa, T.M.; Flint, H.J. Occurrence of the new tetracycline resistance gene tet(W) in bacteria from the human gut. Antimicrob. Agents Chemother. 2000, 44, 775–777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gevers, D.; Danielsen, M.; Huys, G.; Swings, J. Molecular characterization of tet(M) genes in Lactobacillus isolates from different types of fermented dry sausage. Appl. Environ. Microbiol. 2003, 69, 1270–1275. [Google Scholar] [CrossRef] [Green Version]
- Jensen, L.B.; Frimodt-Møller, N.; Aarestrup, F.M. Presence of erm gene classes in Gram-positive bacteria of animal and human origin in Denmark. FEMS Microbiol. Lett. 1999, 170, 151–158. [Google Scholar] [CrossRef]
- Roberts, M.C.; Chung, W.O.; Roe, D.; Xia, M.; Marquez, C.; Borthagaray, G.; Whittington, W.L.; Holmes, K.K. Erythromycin-resistant Neisseria gonorrhoeae and oral commensal Neisseria spp. carry known rRNA methylase genes. Antimicrob. Agents Chemother. 1999, 43, 1367–1372. [Google Scholar] [CrossRef] [Green Version]
- Luna, V.A.; Cousin, S., Jr.; Whittington, W.L.; Roberts, M.C. Identification of the conjugative mef gene in clinical Acinetobacter junii and Neisseria gonorrhoeae isolates. Antimicrob. Agents Chemother. 2000, 44, 2503–2506. [Google Scholar] [CrossRef] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Antipov, D.; Hartwick, N.; Shen, M.; Raiko, M.; Lapidus, A.; Pevzner, P.A. PlasmidSPAdes: Assembling plasmids from whole genome sequencing data. Bioinformatics 2016, 32, 3380–3387. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species/Strain | Minimum Inhibitory Concentration (μg mL−1) | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Gm | Km | Sm | Nm | Tc | Em | Cl | Cm | Am | Pc | Va | Q-da | Lz | Tm | Ci | Rif | |
| Lactococcus lactis NZ9000 | 1 | 4 | 16 | 4 | 1 | 0.12 | 0.12 | 4 | 0.25 | 0.5 | 0.5 | 4 | 2 | >64 | 16 | 64 |
| L. lactis pTC1 | 1 | 4 | 16 | 4 | 128 | 0.06 | 0.12 | 4 | 0.25 | 0.25 | 0.25 | 4 | 2 | >64 | 16 | 32 |
| L. lactis pTC2 | 0.5 | 4 | 16 | 2 | 128 | 0.06 | 0.06 | 4 | 0.25 | 0.25 | 0.5 | 4 | 2 | >64 | 16 | 32 |
| L. lactis pTC3 | 0.5 | 4 | 64 | 2 | 128 | 0.06 | 0.12 | 32 | 0.25 | 0.25 | 0.5 | 4 | 1 | >64 | 16 | 64 |
| L. lactis pTC4 | 1 | 4 | 16 | 4 | 128 | 0.06 | 0.12 | 4 | 0.25 | 0.25 | 0.25 | 4 | 2 | >64 | 16 | 32 |
| L. lactis pTC5 | 1 | 4 | 32 | 2 | 192 | 0.12 | 0.12 | 16 | 0.25 | 0.25 | 0.5 | 4 | 2 | >64 | 16 | 64 |
| L. lactis pEMR1 | 1 | 4 | >256 | 32 | >256 | >256 | >256 | 8 | 0.25 | 0.25 | 0.5 | >8 | 2 | >64 | 16 | 64 |
| Lacticaseibacillus casei BL23 | 2 | 16 | 8 | 4 | 2 | 0.06 | 0.03 | 4 | 1 | 0.5 | >128 | 0.5 | 1 | 1 | 1 | 0.12 |
| L. casei pTC6 | 1 | 32 | 16 | 2 | 32 | 0.25 | 0.06 | 8 | 0.5 | 1 | >128 | 2 | 4 | 0.5 | 32 | 8 |
| Lactiplantibacillus plantarum NC8 | 0.5 | 16 | 32 | 1 | 32 | 0.12 | 1 | 8 | 0.25 | 2 | >128 | 2 | 4 | 8 | 32 | 2 |
| L. plantarum pTC7 | 0.5 | 16 | 16 | 0.5 | 64 | 0.25 | 1 | 8 | 0.25 | 2 | >128 | 1 | 4 | 16 | 64 | 2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Flórez, A.B.; Vázquez, L.; Rodríguez, J.; Mayo, B. Directed Recovery and Molecular Characterization of Antibiotic Resistance Plasmids from Cheese Bacteria. Int. J. Mol. Sci. 2021, 22, 7801. https://doi.org/10.3390/ijms22157801
Flórez AB, Vázquez L, Rodríguez J, Mayo B. Directed Recovery and Molecular Characterization of Antibiotic Resistance Plasmids from Cheese Bacteria. International Journal of Molecular Sciences. 2021; 22(15):7801. https://doi.org/10.3390/ijms22157801
Chicago/Turabian StyleFlórez, Ana Belén, Lucía Vázquez, Javier Rodríguez, and Baltasar Mayo. 2021. "Directed Recovery and Molecular Characterization of Antibiotic Resistance Plasmids from Cheese Bacteria" International Journal of Molecular Sciences 22, no. 15: 7801. https://doi.org/10.3390/ijms22157801
APA StyleFlórez, A. B., Vázquez, L., Rodríguez, J., & Mayo, B. (2021). Directed Recovery and Molecular Characterization of Antibiotic Resistance Plasmids from Cheese Bacteria. International Journal of Molecular Sciences, 22(15), 7801. https://doi.org/10.3390/ijms22157801