
Angulin-1 (LSR) Affects Paracellular Water Transport, However Only in Tight Epithelial Cells





Abstract
:1. Introduction
2. Results
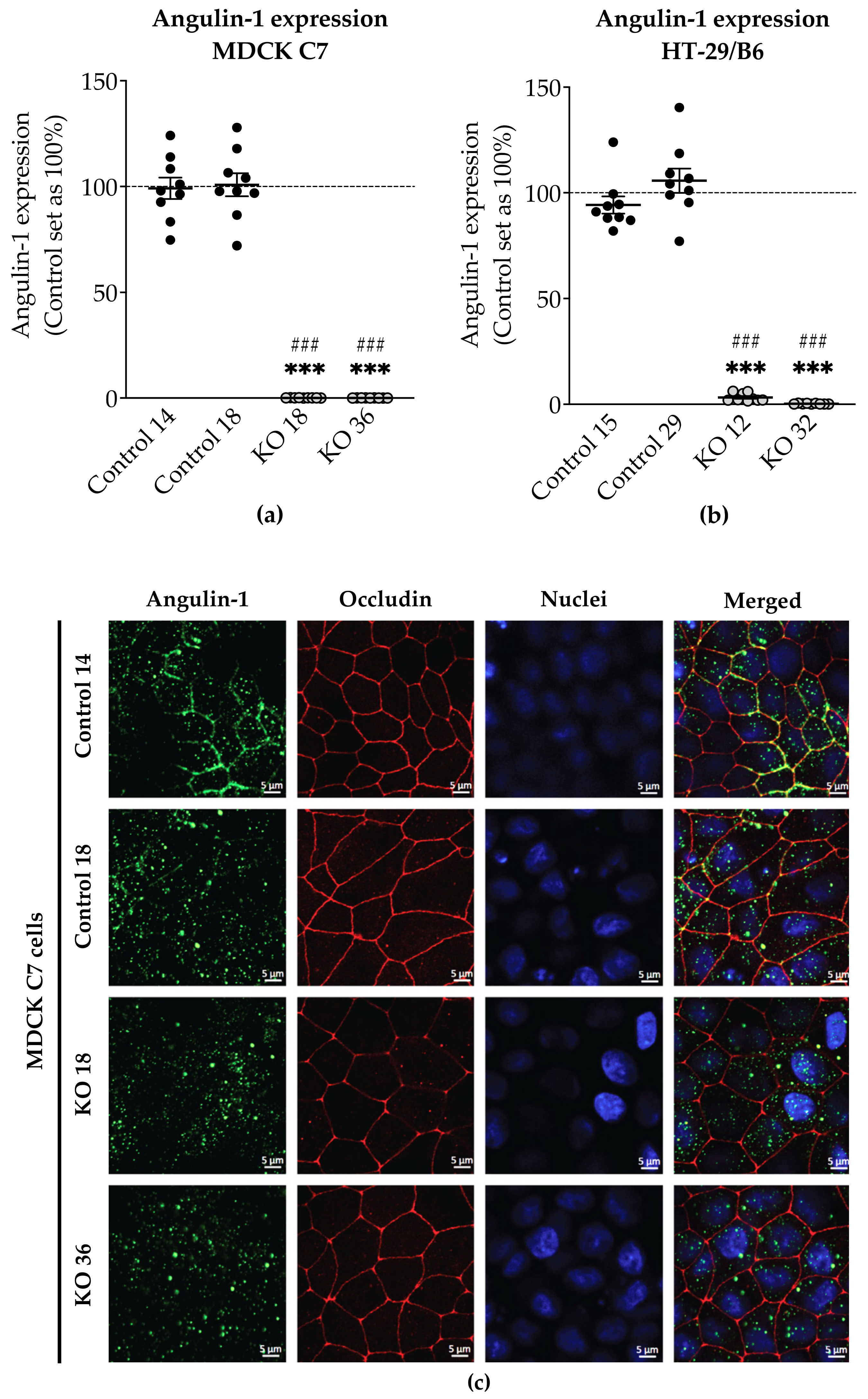
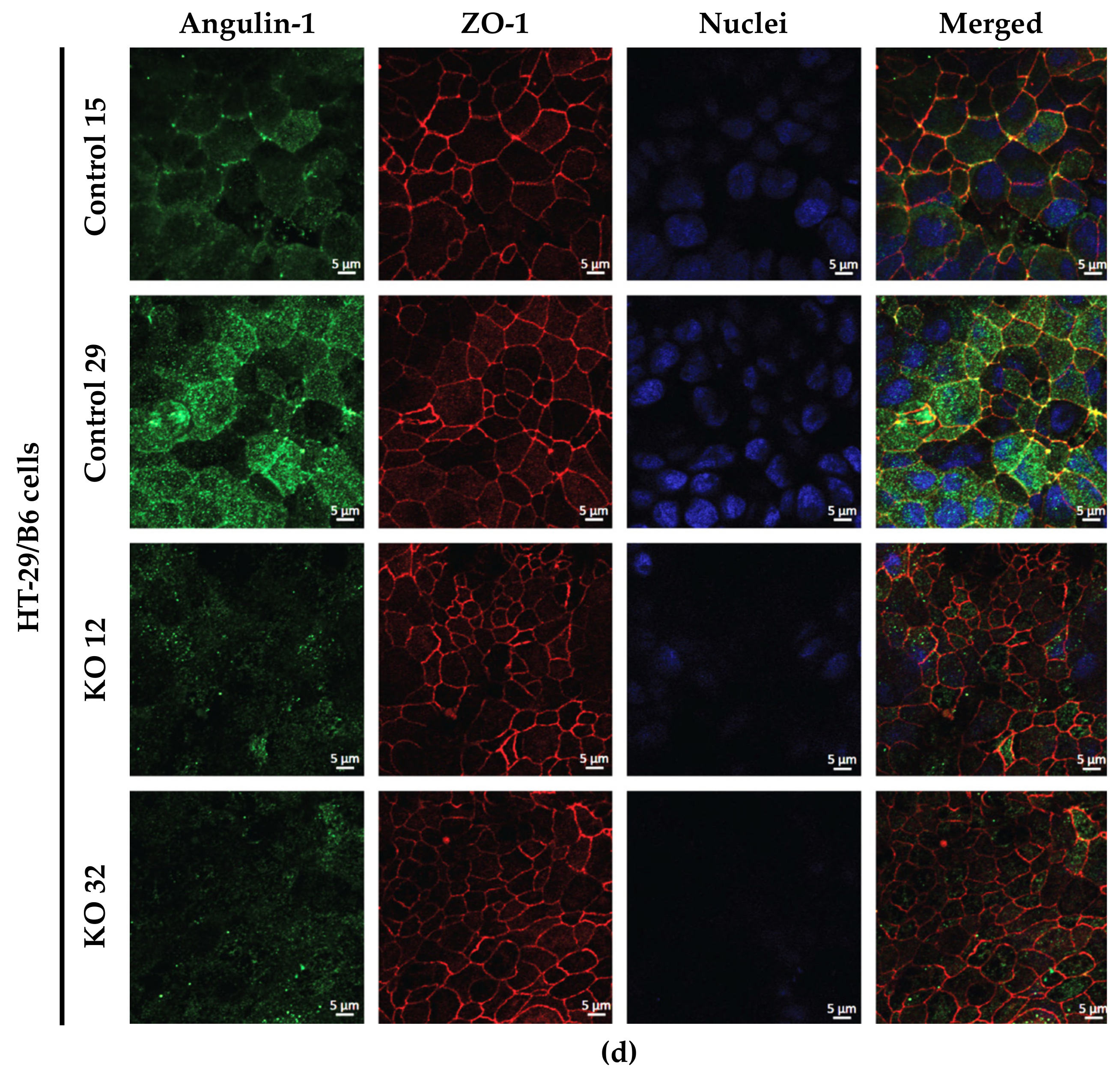
2.1. Establishment of Angulin-1 Knockout in MDCK C7 and HT-29/B6 Cells
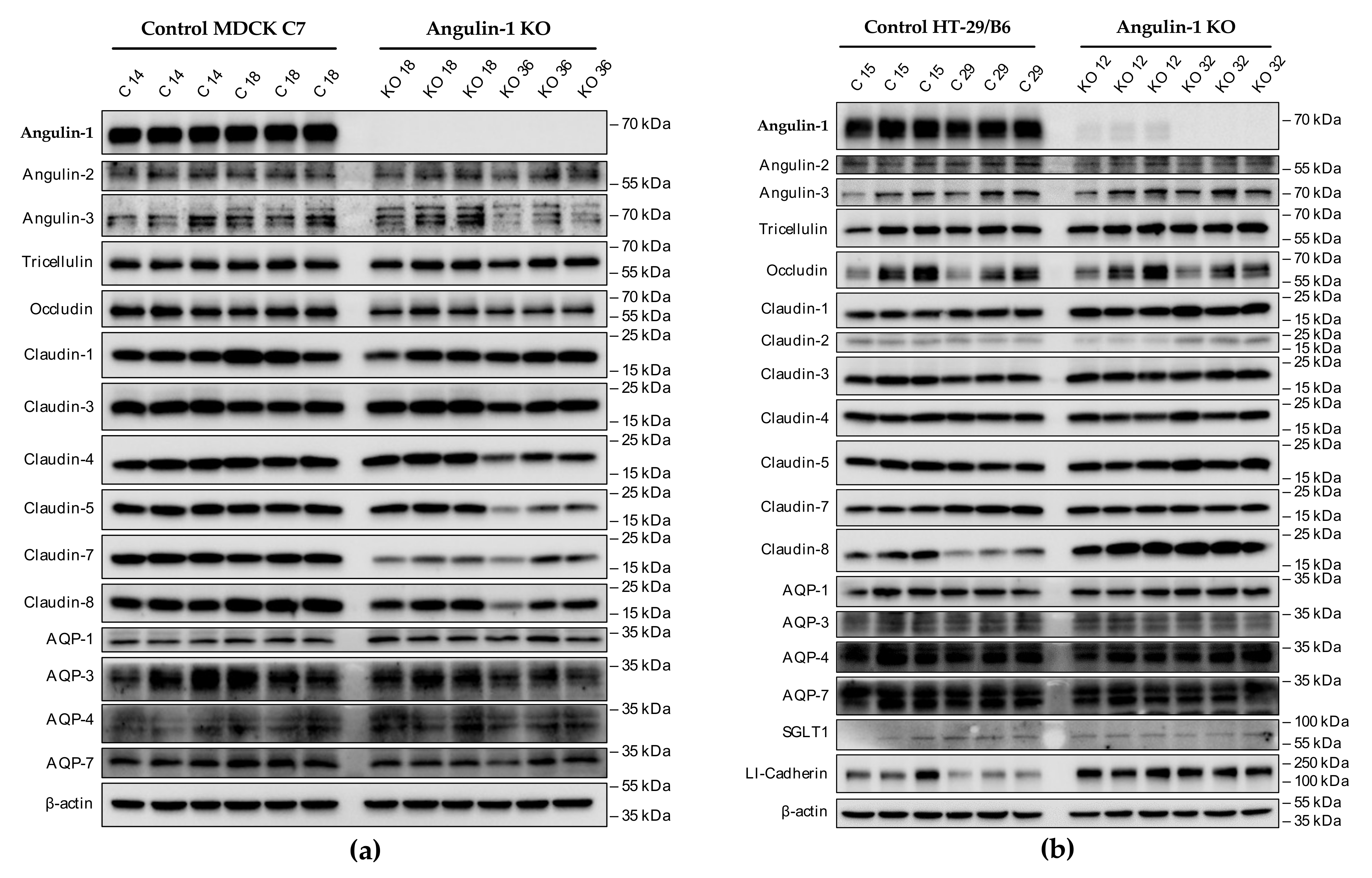
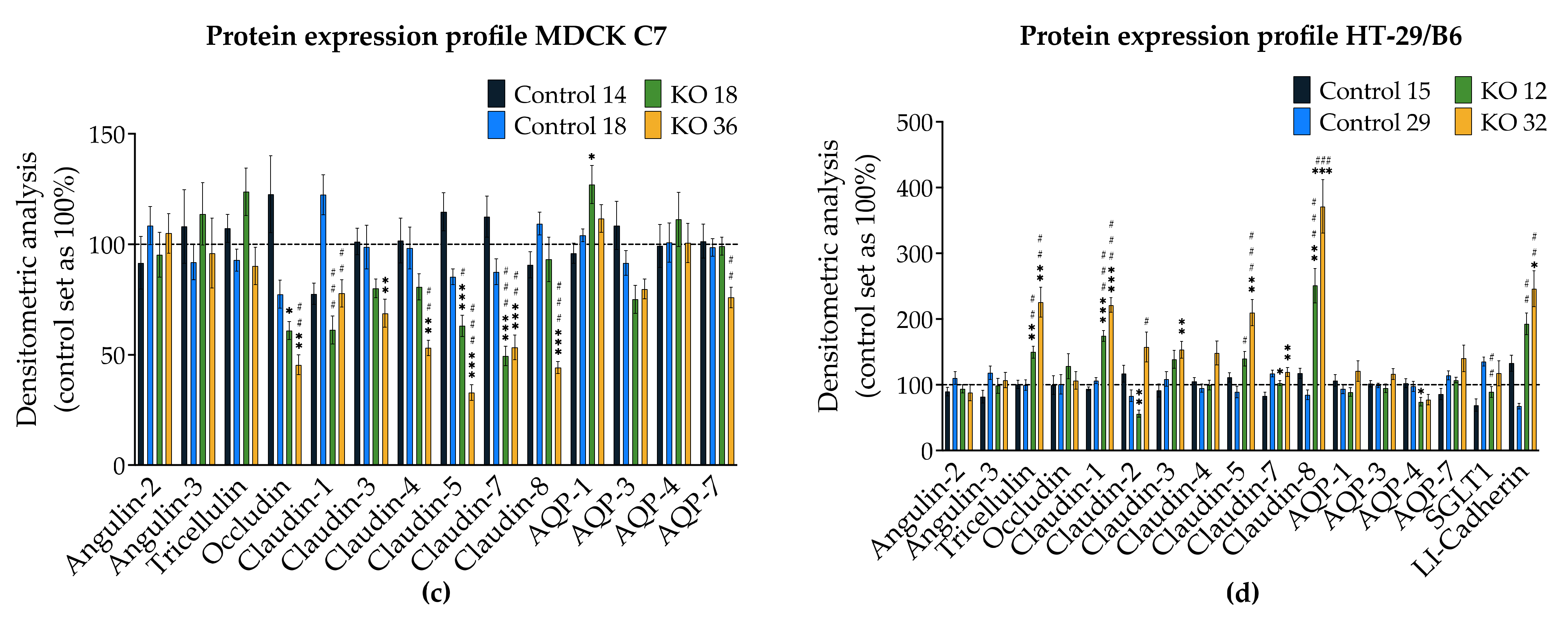
2.2. Effects of Angulin-1 Knockout on Endogenous Proteins of MDCK C7 and HT-29/B6 Cells
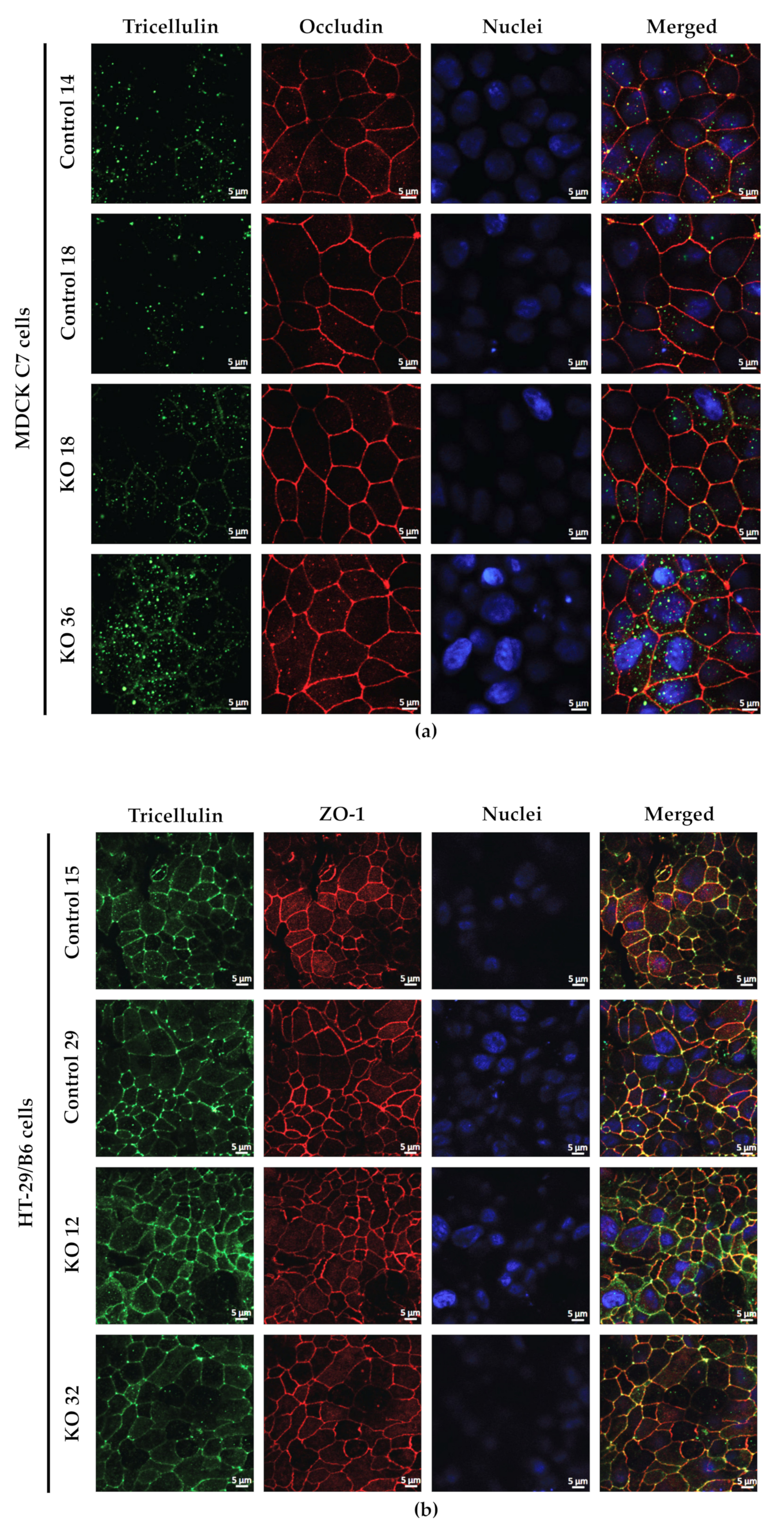
2.3. Effects of Angulin-1 Knockout on Tricellulin Localization in MDCK C7 and HT-29/B6 Cells
2.4. Effect of Angulin-1 Knockout on the TJ Ultrastructural Level in MDCK C7 and HT-29/B6 Cells
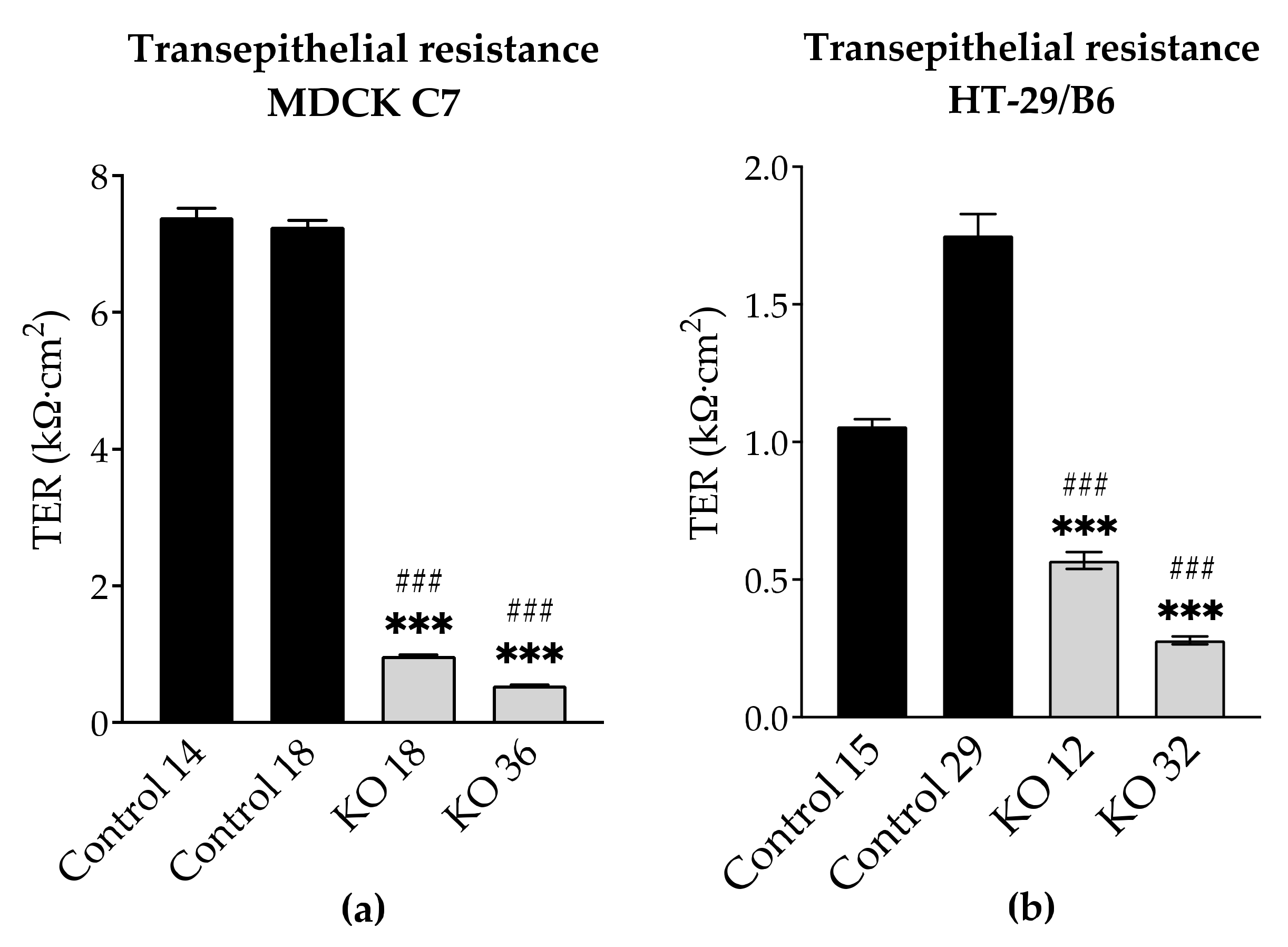
2.5. Effects of Angulin-1 Knockout on Ion Permeability in MDCK C7 and HT-29/B6 Cells
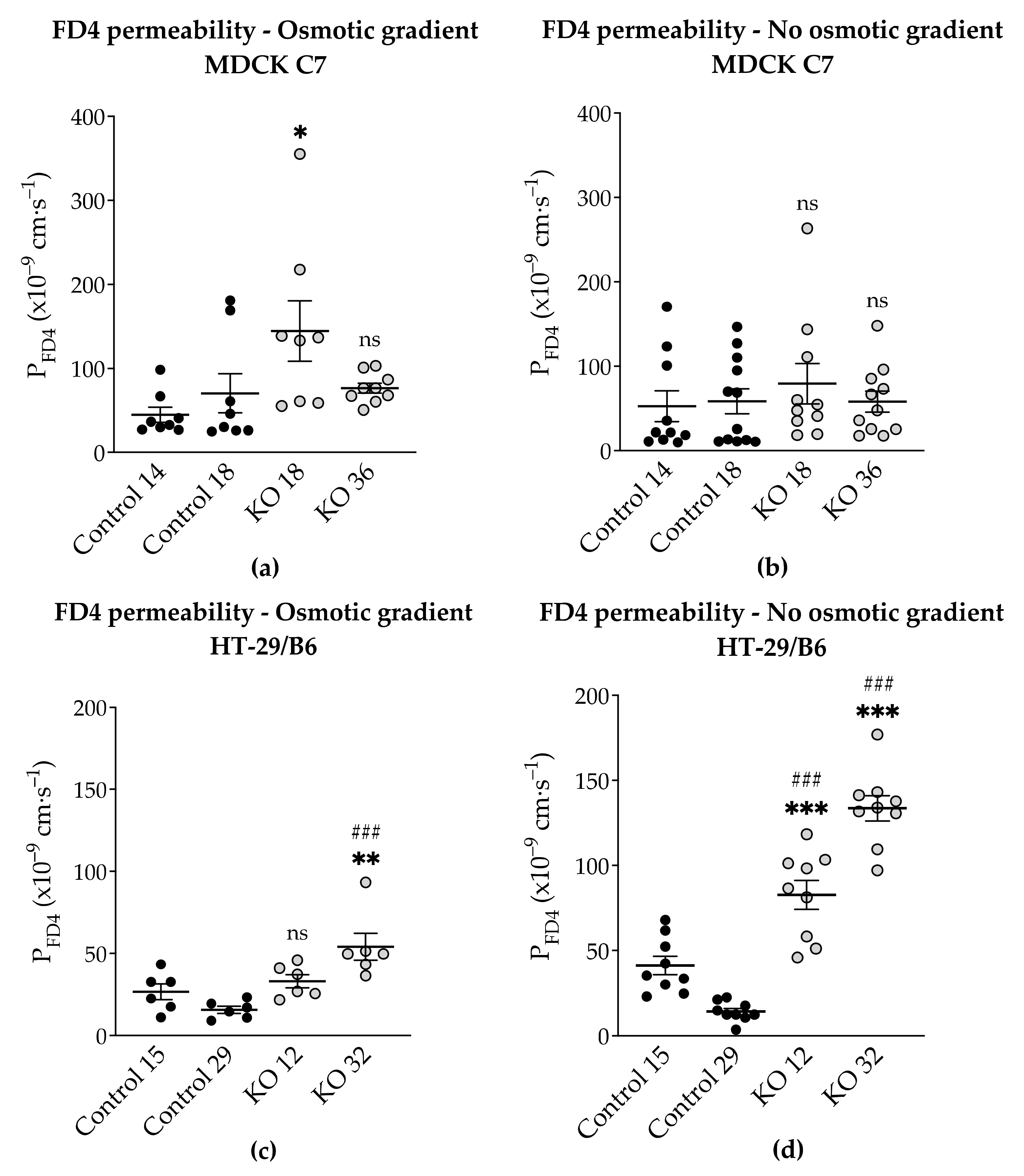
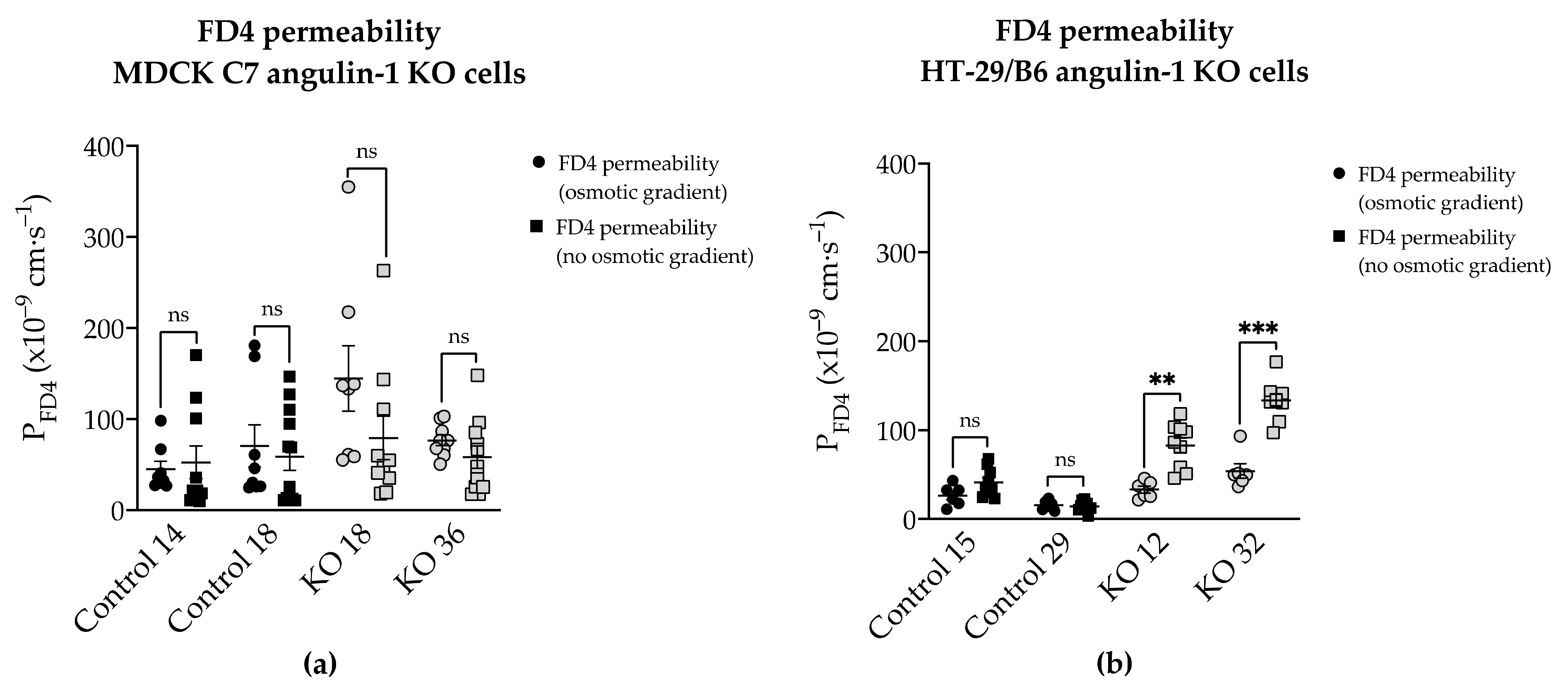
2.6. Effects of Angulin-1 Knockout on Macromolecule Permeability in MDCK C7 and HT-29/B6 Cells
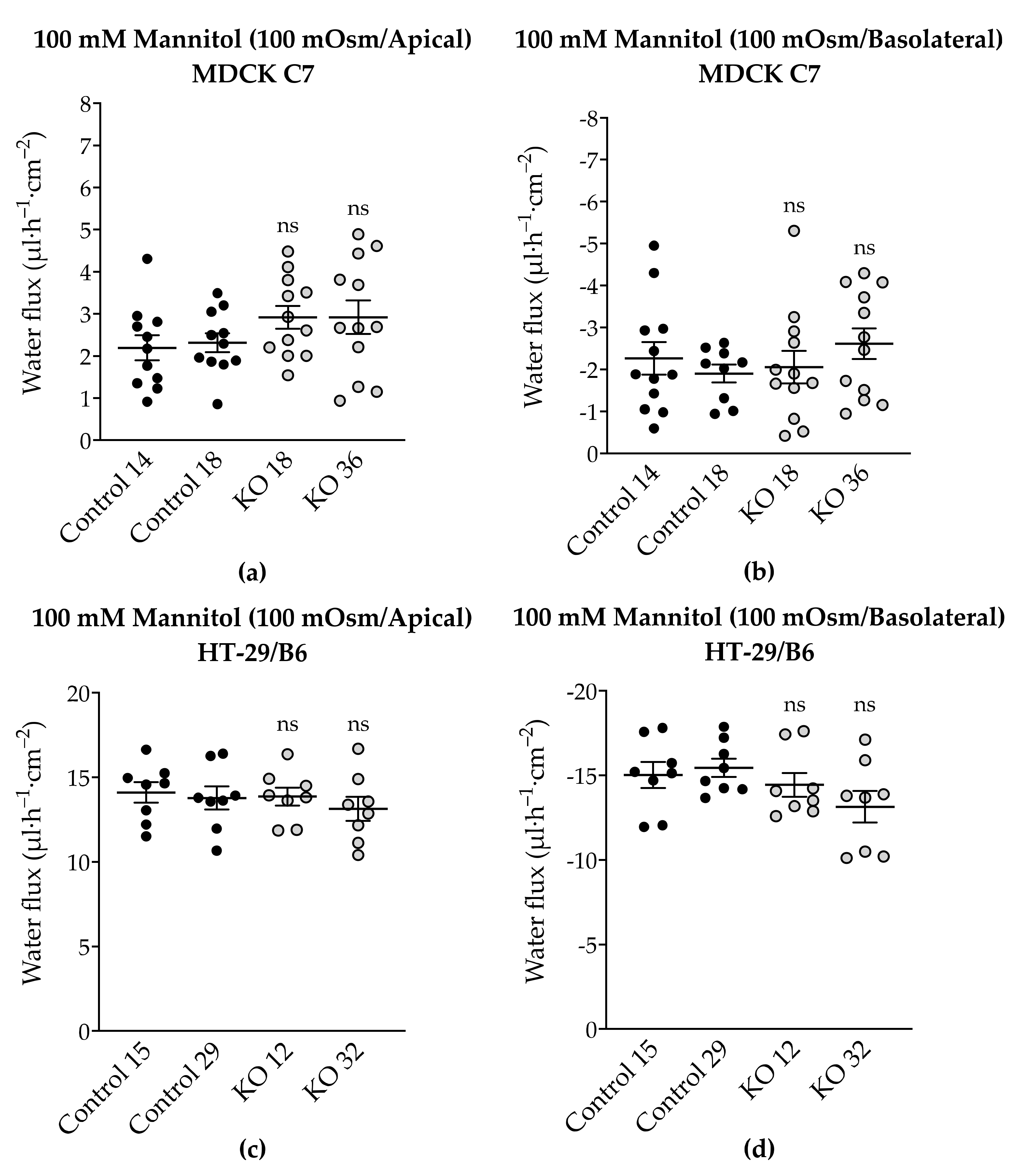
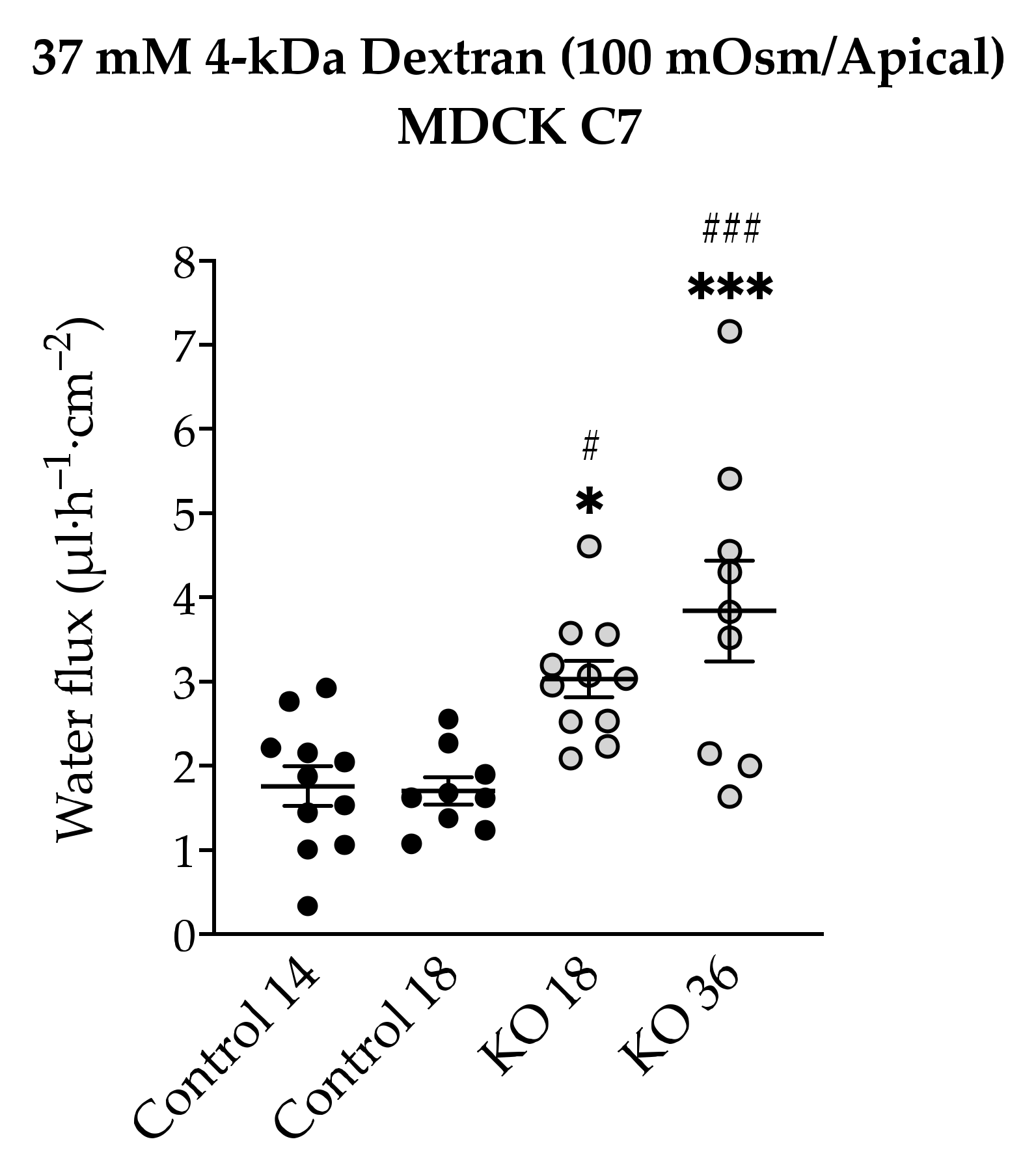
2.7. Effect of Angulin-1 Knockout on Transepithelial Water Transport in MDCK C7 and HT-29/B6 Cells
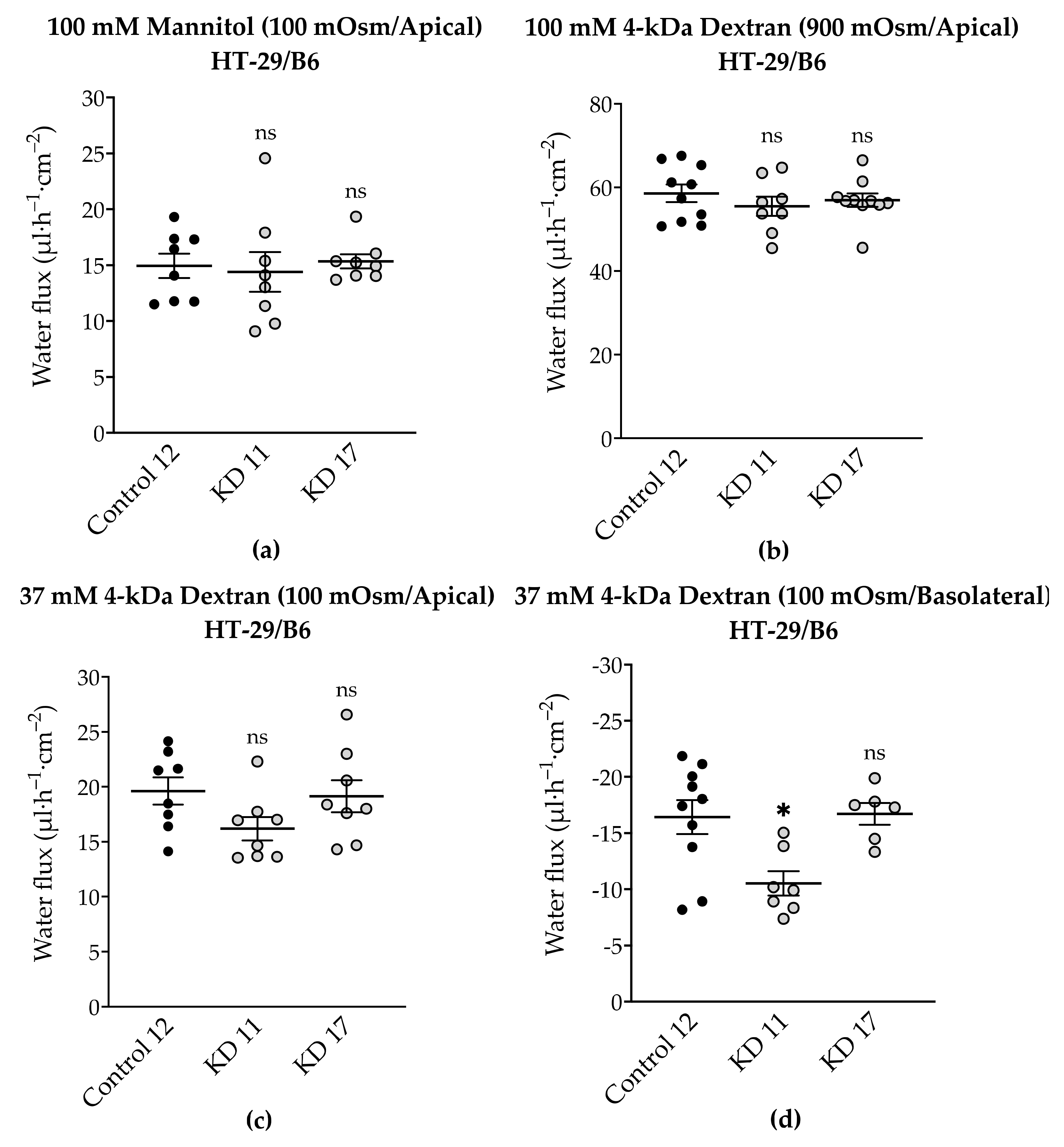
2.8. Effect of Tricellulin KD on Transepithelial Water Transport in HT-29/B6 Cells
3. Discussion
3.1. Angulin-1 Knockout Alters the Expression and Localization of Other Proteins in MDCK C7 and HT-29/B6 Cells
3.2. Angulin-1 Knockout Did Not Alter the Ultrastructure of the Bicellular Tight Junction in MDCK C7 and HT-29/B6 Cells
3.3. Angulin-1 Knockout Increases the Ion Permeability in MDCK C7 and HT-29/B6 Cells
3.4. Angulin-1 Knockout Increases the Macromolecule Permeability Only in HT-29/B6 Cells
3.5. Angulin-1 Knockout Increases Transepithelial Water Transport Only in MDCK C7 Cells
3.6. Tricellulin Knockdown Does Not Significantly Affect Paracellular Water Transport in the HT-29/B6 Cell Line
4. Materials and Methods
4.1. Cell Culture, Transfection and TER Measurement
4.2. Western Blot Analysis
4.3. Immunofluorescent Staining
4.4. Freeze Fracture Electron Microscopy
4.5. Measurement of 4-kDa FITC-Dextran Flux
4.6. Dilution Potential Measurements
4.7. Measurement of Transepithelial Water Transport
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| angulin-1 | lipolysis-stimulated lipoprotein receptor (LSR) |
| angulin-2 | immunoglobulin-like domain containing receptor-1 (ILDR1) |
| angulin-3 | immunoglobulin-like domain containing receptor-2 (ILDR2) |
| bTJ | bicellular tight junction, located between two cells |
| CRISPR/Cas9 | Clustered Regularly Interspaced Short Palindromic Repeats/Cas9 |
| d(Å) | diameter (Å) |
| FD4 | 4-kDa FITC-dextran |
| HDR | homology-directed repair |
| KD | knockdown |
| KO | knockout |
| SEM | standard error of the mean |
| TJ | tight junction |
| TER | transepithelial resistance (Ω·cm2) |
| tTJ | tricellular tight junction, located at corners of three cells |
| P | Apparent permeability (cm·s−1) |
References
- Zeuthen, T. General models for water transport across leaky epithelia. Int. Rev. Cytol. 2002, 215, 285–317. [Google Scholar] [CrossRef]
- Marchiando, A.M.; Graham, W.V.; Turner, J.R. Epithelial barriers in homeostasis and disease. Annu. Rev. Pathol. 2010, 5, 119–144. [Google Scholar] [CrossRef]
- Agre, P.; Preston, G.M.; Smith, B.L.; Jung, J.S.; Raina, S.; Moon, C.; Guggino, W.B.; Nielsen, S. Aquaporin CHIP: The archetypal molecular water channel. Am. J. Physiol. 1993, 265, F463–F476. [Google Scholar] [CrossRef] [Green Version]
- Angelow, S.; Ahlstrom, R.; Yu, A.S. Biology of claudins. Am. J. Physiol. Renal Physiol. 2008, 295, F867–F876. [Google Scholar] [CrossRef] [PubMed]
- Ikenouchi, J.; Furuse, M.; Furuse, K.; Sasaki, H.; Tsukita, S.; Tsukita, S. Tricellulin constitutes a novel barrier at tricellular contacts of epithelial cells. J. Cell Biol. 2005, 171, 939–945. [Google Scholar] [CrossRef] [PubMed]
- Furuse, M.; Izumi, Y.; Oda, Y.; Higashi, T.; Iwamoto, N. Molecular organization of tricellular tight junctions. Tissue Barriers 2014, 2, e28960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varadarajan, S.; Stephenson, R.E.; Miller, A.L. Multiscale dynamics of tight junction remodeling. J. Cell Sci. 2019, 132. [Google Scholar] [CrossRef] [PubMed]
- Furuse, M. Molecular basis of the core structure of tight junctions. Cold Spring Harb. Perspect. Biol. 2010, 2, a002907. [Google Scholar] [CrossRef] [PubMed]
- Van Itallie, C.M.; Lidman, K.F.; Tietgens, A.J.; Anderson, J.M. Newly synthesized claudins but not occludin are added to the basal side of the tight junction. Mol. Biol. Cell 2019, 30, 1406–1424. [Google Scholar] [CrossRef]
- Schulzke, J.D.; Gitter, A.H.; Mankertz, J.; Spiegel, S.; Seidler, U.; Amasheh, S.; Saitou, M.; Tsukita, S.; Fromm, M. Epithelial transport and barrier function in occludin-deficient mice. Biochim. Biophys. Acta 2005, 1669, 34–42. [Google Scholar] [CrossRef] [Green Version]
- Günzel, D.; Fromm, M. Claudins and other tight junction proteins. Compr. Physiol. 2012, 2, 1819–1852. [Google Scholar] [CrossRef]
- Rosenthal, R.; Milatz, S.; Krug, S.M.; Oelrich, B.; Schulzke, J.D.; Amasheh, S.; Günzel, D.; Fromm, M. Claudin-2, a component of the tight junction, forms a paracellular water channel. J. Cell Sci. 2010, 123, 1913–1921. [Google Scholar] [CrossRef] [Green Version]
- Rosenthal, R.; Günzel, D.; Krug, S.M.; Schulzke, J.D.; Fromm, M.; Yu, A.S. Claudin-2-mediated cation and water transport share a common pore. Acta Physiol. 2017, 219, 521–536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenthal, R.; Günzel, D.; Piontek, J.; Krug, S.M.; Ayala-Torres, C.; Hempel, C.; Theune, D.; Fromm, M. Claudin-15 forms a water channel through the tight junction with distinct function compared to claudin-2. Acta Physiol. 2020, 228, e13334. [Google Scholar] [CrossRef]
- Schnermann, J.; Huang, Y.; Mizel, D. Fluid reabsorption in proximal convoluted tubules of mice with gene deletions of claudin-2 and/or aquaporin1. Am. J. Physiol. Renal Physiol. 2013, 305, F1352–F1364. [Google Scholar] [CrossRef] [Green Version]
- Robben, J.H.; Knoers, N.V.; Deen, P.M. Cell biological aspects of the vasopressin type-2 receptor and aquaporin 2 water channel in nephrogenic diabetes insipidus. Am. J. Physiol. Renal Physiol. 2006, 291, F257–F270. [Google Scholar] [CrossRef]
- Staehelin, L.A. Further observations on the fine structure of freeze-cleaved tight junctions. J. Cell Sci. 1973, 13, 763–786. [Google Scholar] [CrossRef]
- Higashi, T.; Miller, A.L. Tricellular junctions: How to build junctions at the TRICkiest points of epithelial cells. Mol. Biol. Cell 2017, 28, 2023–2034. [Google Scholar] [CrossRef] [PubMed]
- Krug, S.M.; Amasheh, S.; Richter, J.F.; Milatz, S.; Günzel, D.; Westphal, J.K.; Huber, O.; Schulzke, J.D.; Fromm, M. Tricellulin Forms a Barrier to Macromolecules in Tricellular Tight Junctions without Affecting Ion Permeability. Mol. Biol. Cell 2009, 20, 3713–3724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krug, S.M. Contribution of the tricellular tight junction to paracellular permeability in leaky and tight epithelia. Ann. N. Y. Acad. Sci. 2017, 1397, 219–230. [Google Scholar] [CrossRef] [PubMed]
- Ayala-Torres, C.; Krug, S.M.; Schulzke, J.D.; Rosenthal, R.; Fromm, M. Tricellulin Effect on Paracellular Water Transport. Int. J. Mol. Sci. 2019, 20, 5700. [Google Scholar] [CrossRef] [Green Version]
- Higashi, T.; Tokuda, S.; Kitajiri, S.; Masuda, S.; Nakamura, H.; Oda, Y.; Furuse, M. Analysis of the ’angulin’ proteins LSR, ILDR1 and ILDR2--tricellulin recruitment, epithelial barrier function and implication in deafness pathogenesis. J. Cell Sci. 2013, 126, 966–977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stenger, C.; Pincon, A.; Hanse, M.; Royer, L.; Comte, A.; Koziel, V.; Olivier, J.L.; Pillot, T.; Yen, F.T. Brain region-specific immunolocalization of the lipolysis-stimulated lipoprotein receptor (LSR) and altered cholesterol distribution in aged LSR+/− mice. J. Neurochem. 2012, 123, 467–476. [Google Scholar] [CrossRef]
- Nakatsu, D.; Kano, F.; Taguchi, Y.; Sugawara, T.; Nishizono, T.; Nishikawa, K.; Oda, Y.; Furuse, M.; Murata, M. JNK1/2-dependent phosphorylation of angulin-1/LSR is required for the exclusive localization of angulin-1/LSR and tricellulin at tricellular contacts in EpH4 epithelial sheet. Genes Cells 2014, 19, 565–581. [Google Scholar] [CrossRef]
- Masuda, S.; Oda, Y.; Sasaki, H.; Ikenouchi, J.; Higashi, T.; Akashi, M.; Nishi, E.; Furuse, M. LSR defines cell corners for tricellular tight junction formation in epithelial cells. J. Cell Sci. 2011, 124, 548–555. [Google Scholar] [CrossRef] [Green Version]
- Czulkies, B.A.; Mastroianni, J.; Lutz, L.; Lang, S.; Schwan, C.; Schmidt, G.; Lassmann, S.; Zeiser, R.; Aktories, K.; Papatheodorou, P. Loss of LSR affects epithelial barrier integrity and tumor xenograft growth of CaCo-2 cells. Oncotarget 2017, 8, 37009–37022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohno, T.; Konno, T.; Kojima, T. Role of Tricellular Tight Junction Protein Lipolysis-Stimulated Lipoprotein Receptor (LSR) in Cancer Cells. Int. J. Mol. Sci. 2019, 20, 3555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimada, H.; Satohisa, S.; Kohno, T.; Konno, T.; Takano, K.I.; Takahashi, S.; Hatakeyama, T.; Arimoto, C.; Saito, T.; Kojima, T. Downregulation of lipolysis-stimulated lipoprotein receptor promotes cell invasion via claudin-1-mediated matrix metalloproteinases in human endometrial cancer. Oncol. Lett. 2017, 14, 6776–6782. [Google Scholar] [CrossRef]
- Mesli, S.; Javorschi, S.; Berard, A.M.; Landry, M.; Priddle, H.; Kivlichan, D.; Smith, A.J.; Yen, F.T.; Bihain, B.E.; Darmon, M. Distribution of the lipolysis stimulated receptor in adult and embryonic murine tissues and lethality of LSR-/- embryos at 12.5 to 14.5 days of gestation. Eur. J. Biochem. 2004, 271, 3103–3114. [Google Scholar] [CrossRef] [PubMed]
- Gong, Y.; Himmerkus, N.; Sunq, A.; Milatz, S.; Merkel, C.; Bleich, M.; Hou, J. ILDR1 is important for paracellular water transport and urine concentration mechanism. Proc. Natl. Acad. Sci. USA 2017, 114, 5271–5276. [Google Scholar] [CrossRef] [Green Version]
- Hempstock, W.; Sugioka, S.; Ishizuka, N.; Sugawara, T.; Furuse, M.; Hayashi, H. Angulin-2/ILDR1, a tricellular tight junction protein, does not affect water transport in the mouse large intestine. Sci. Rep. 2020, 10, 10374. [Google Scholar] [CrossRef]
- Krug, S.M.; Fromm, M.; Günzel, D. Two-path impedance spectroscopy for measuring paracellular and transcellular epithelial resistance. Biophys. J. 2009, 97, 2202–2211. [Google Scholar] [CrossRef] [Green Version]
- Stevenson, B.R.; Siliciano, J.D.; Mooseker, M.S.; Goodenough, D.A. Identification of ZO-1: A high molecular weight polypeptide associated with the tight junction (zonula occludens) in a variety of epithelia. J. Cell Biol. 1986, 103, 755–766. [Google Scholar] [CrossRef] [Green Version]
- Tsukita, S.; Furuse, M. Occludin and claudins in tight-junction strands: Leading or supporting players? Trends Cell Biol. 1999, 9, 268–273. [Google Scholar] [CrossRef]
- Amasheh, S.; Meiri, N.; Gitter, A.H.; Schöneberg, T.; Mankertz, J.; Schulzke, J.D.; Fromm, M. Claudin-2 expression induces cation-selective channels in tight junctions of epithelial cells. J. Cell Sci. 2002, 115, 4969–4976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furuse, M.; Oda, Y.; Higashi, T.; Iwamoto, N.; Masuda, S. Lipolysis-stimulated lipoprotein receptor: A novel membrane protein of tricellular tight junctions. Ann. N. Y. Acad. Sci. 2012, 1257, 54–58. [Google Scholar] [CrossRef] [PubMed]
- Kirschner, N.; Rosenthal, R.; Furuse, M.; Moll, I.; Fromm, M.; Brandner, J.M. Contribution of tight junction proteins to ion, macromolecule, and water barrier in keratinocytes. J. Investig. Dermatol. 2013, 133, 1161–1169. [Google Scholar] [CrossRef] [Green Version]
- Ikenouchi, J.; Sasaki, H.; Tsukita, S.; Furuse, M.; Tsukita, S. Loss of occludin affects tricellular localization of tricellulin. Mol. Biol. Cell 2008, 19, 4687–4693. [Google Scholar] [CrossRef] [Green Version]
- Nakatsu, D.; Kano, F.; Shinozaki-Narikawa, N.; Murata, M. Pyk2-dependent phosphorylation of LSR enhances localization of LSR and tricellulin at tricellular tight junctions. PLoS ONE 2019, 14, e0223300. [Google Scholar] [CrossRef]
- Weth, A.; Dippl, C.; Striedner, Y.; Tiemann-Boege, I.; Vereshchaga, Y.; Golenhofen, N.; Bartelt-Kirbach, B.; Baumgartner, W. Water transport through the intestinal epithelial barrier under different osmotic conditions is dependent on LI-cadherin trans-interaction. Tissue Barriers 2017, 5, e1285390. [Google Scholar] [CrossRef] [Green Version]
- Vereshchaga, Y.; Arnold, N.; Baumgartner, W. Physiological relevance of epithelial geometry: New insights into the standing gradient model and the role of LI cadherin. PLoS ONE 2018, 13, e0208791. [Google Scholar] [CrossRef] [PubMed]
- Higashi, T.; Katsuno, T.; Kitajiri, S.; Furuse, M. Deficiency of angulin-2/ILDR1, a tricellular tight junction-associated membrane protein, causes deafness with cochlear hair cell degeneration in mice. PLoS ONE 2015, 10, e0120674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furuse, M.; Sasaki, H.; Tsukita, S. Manner of interaction of heterogeneous claudin species within and between tight junction strands. J. Cell Biol. 1999, 147, 891–903. [Google Scholar] [CrossRef] [PubMed]
- Krug, S.M.; Hayaishi, T.; Iguchi, D.; Watari, A.; Takahashi, A.; Fromm, M.; Nagahama, M.; Takeda, H.; Okada, Y.; Sawasaki, T.; et al. Angubindin-1, a novel paracellular absorption enhancer acting at the tricellular tight junction. J. Control. Release 2017, 260, 1–11. [Google Scholar] [CrossRef]
- Kreusel, K.M.; Fromm, M.; Schulzke, J.D.; Hegel, U. Cl- secretion in epithelial monolayers of mucus-forming human colon cells (HT-29/B6). Am. J. Physiol. 1991, 261, C574–C582. [Google Scholar] [CrossRef] [PubMed]
- Günzel, D.; Stuiver, M.; Kausalya, P.J.; Haisch, L.; Krug, S.M.; Rosenthal, R.; Meij, I.C.; Hunziker, W.; Fromm, M.; Müller, D. Claudin-10 exists in six alternatively spliced isoforms that exhibit distinct localization and function. J. Cell Sci. 2009, 122, 1507–1517. [Google Scholar] [CrossRef] [PubMed] [Green Version]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MDCK C7 Cells | HT-29/B6 Cells | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Control 14 (n = 29) | Control 18 (n = 26) | KO 18 (n = 18) | KO 36 (n = 28) | Control 15 (n = 17) | Control 29 (n = 20) | KO 12 (n = 11) | KO 32 (n = 27) | ||
| Number of strands | 3.62 ± 0.23 | 3.42 ± 0.20 | 4.17 ± 0.35 | 3.46 ± 0.23 | 4.53 ± 0.39 | 3.61 ± 0.34 | 3.91 ± 0.31 | 4.59 ± 0.41 | |
| Meshwork depth a (nm) | 151.03 ± 11.00 | 165.00 ± 22.42 | 171.11 ± 24.78 | 157.50 ± 16.22 | 276.47 ± 37.42 | 228.89 ± 36.71 | 219.09 ± 23.06 | 310.93 ± 39.08 | |
| Strand density b (1/pm) | 23.97 ± 2.32 | 20.75 ± 3.07 | 24.35 ± 4.09 | 22.00 ± 2.70 | 16.38 ± 2.64 | 15.78 ± 2.94 | 17.84 ± 2.36 | 14.77 ± 2.27 | |
| Number of breaks c per µm | 0.0 ± 0.0 | 0.01 ± 0.009 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.02 ± 0.015 | 0.0 ± 0.0 | 0.03 ± 0.029 | 0.0 ± 0.0 | |
| Strand appearance | Continuous (%) | 100 | 92 | 100 | 100 | 94 | 100 | 100 | 93 |
| Particle (%) | 0 | 8 | 0 | 0 | 6 | 0 | 0 | 7 | |
| Strand pattern | Straight (%) | 100 | 100 | 100 | 100 | 100 | 89 | 100 | 85 |
| Curved (%) | 0 | 0 | 0 | 0 | 0 | 11 | 0 | 15 | |
| Control 12 | KD 11 | KD 17 | ||
|---|---|---|---|---|
| Tricellulin expression (%) | 100.0 ± 8.9 (n = 11) | 64.6 ± 4.7 *** (n = 10) | 55.7 ± 6.3 *** (n = 10) | |
| TER (kΩ·cm2) | 1.44 ± 0.07 (n = 12) | 0.51 ± 0.03 *** (n = 12) | 0.54 ± 0.02 *** (n = 12) | |
| PFD4 (×10−9 cm·s−1) Osmotic gradient | 21.34 ± 3.80 (n = 7) | 42.96 ± 2.56 *** (n = 7) | 67.55 ± 6.54 *** (n = 7) | |
| PNa (×10−6 cm·s−1) | 0.61 ± 0.10 (n = 10) | 1.50 ± 0.11 *** (n = 10) | 2.46 ± 0.30 *** (n = 11) | |
| PCl (×10−6 cm·s−1) | 0.80 ± 0.18 (n = 10) | 1.84 ± 0.17 ** (n = 10) | 2.98 ± 0.33 ** (n = 11) | |
| PNa/PCl | 0.91 ± 0.08 (n = 10) | 0.92 ± 0.06 (n = 10) | 0.90 ± 0.08 (n = 11) | |
| Water flux—apical side (µL·h−1·cm−2) | 100 mM mannitol (100 mOsm) | 14.9 ± 1.0 (n = 8) | 14.4 ± 1.7 (n = 8) | 15.3 ± 0.6 (n = 8) |
| 37 mM 4-kDa dextran (100 mOsm) | 19.6 ± 1.2 (n = 8) | 16.2 ± 1.0 (n = 8) | 19.1 ± 1.4 (n = 8) | |
| 100 mM 4-kDa dextran (900 mOsm) | 58.6 ± 2.0 (n = 10) | 54.0 ± 2.4 (n = 9) | 56.9 ± 1.6 (n = 10) | |
| Water flux—basolateral side (µL·h−1·cm−2) | 37 mM 4-kDa dextran (100 mOsm) | −16.4 ± 1.4 (n = 10) | −10.5 ± 1.0 * (n = 7) | −16.7 ± 0.9 (n = 6) |
| n | Concentration (mM) | Osmolality (mOsm/kg of Water) Mean ± SEM | ||
|---|---|---|---|---|
| Angulin-1 KO | HEPES only | 17 | - | 289.0 ± 13.7 |
| +Mannitol | 17 | 100 mM | 391.4 ± 14.6 | |
| +4-kDa dextran | 5 | 37 mM | 396.0 ± 4.5 | |
| Tricellulin KD | HEPES only | 10 | - | 308.0 ± 4.4 |
| +Mannitol | 10 | 100 mM | 410.3 ± 1.9 | |
| +4-kDa dextran | 10 | 37 mM | 396.2 ± 6.8 | |
| +4-kDa dextran | 10 | 100 mM | 877.8 ± 13.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ayala-Torres, C.; Krug, S.M.; Rosenthal, R.; Fromm, M. Angulin-1 (LSR) Affects Paracellular Water Transport, However Only in Tight Epithelial Cells. Int. J. Mol. Sci. 2021, 22, 7827. https://doi.org/10.3390/ijms22157827
Ayala-Torres C, Krug SM, Rosenthal R, Fromm M. Angulin-1 (LSR) Affects Paracellular Water Transport, However Only in Tight Epithelial Cells. International Journal of Molecular Sciences. 2021; 22(15):7827. https://doi.org/10.3390/ijms22157827
Chicago/Turabian StyleAyala-Torres, Carlos, Susanne M. Krug, Rita Rosenthal, and Michael Fromm. 2021. "Angulin-1 (LSR) Affects Paracellular Water Transport, However Only in Tight Epithelial Cells" International Journal of Molecular Sciences 22, no. 15: 7827. https://doi.org/10.3390/ijms22157827
APA StyleAyala-Torres, C., Krug, S. M., Rosenthal, R., & Fromm, M. (2021). Angulin-1 (LSR) Affects Paracellular Water Transport, However Only in Tight Epithelial Cells. International Journal of Molecular Sciences, 22(15), 7827. https://doi.org/10.3390/ijms22157827