
Macroalgal Defense against Competitors and Herbivores

 , , , , ,
, , , , ,
Abstract
:1. Introduction
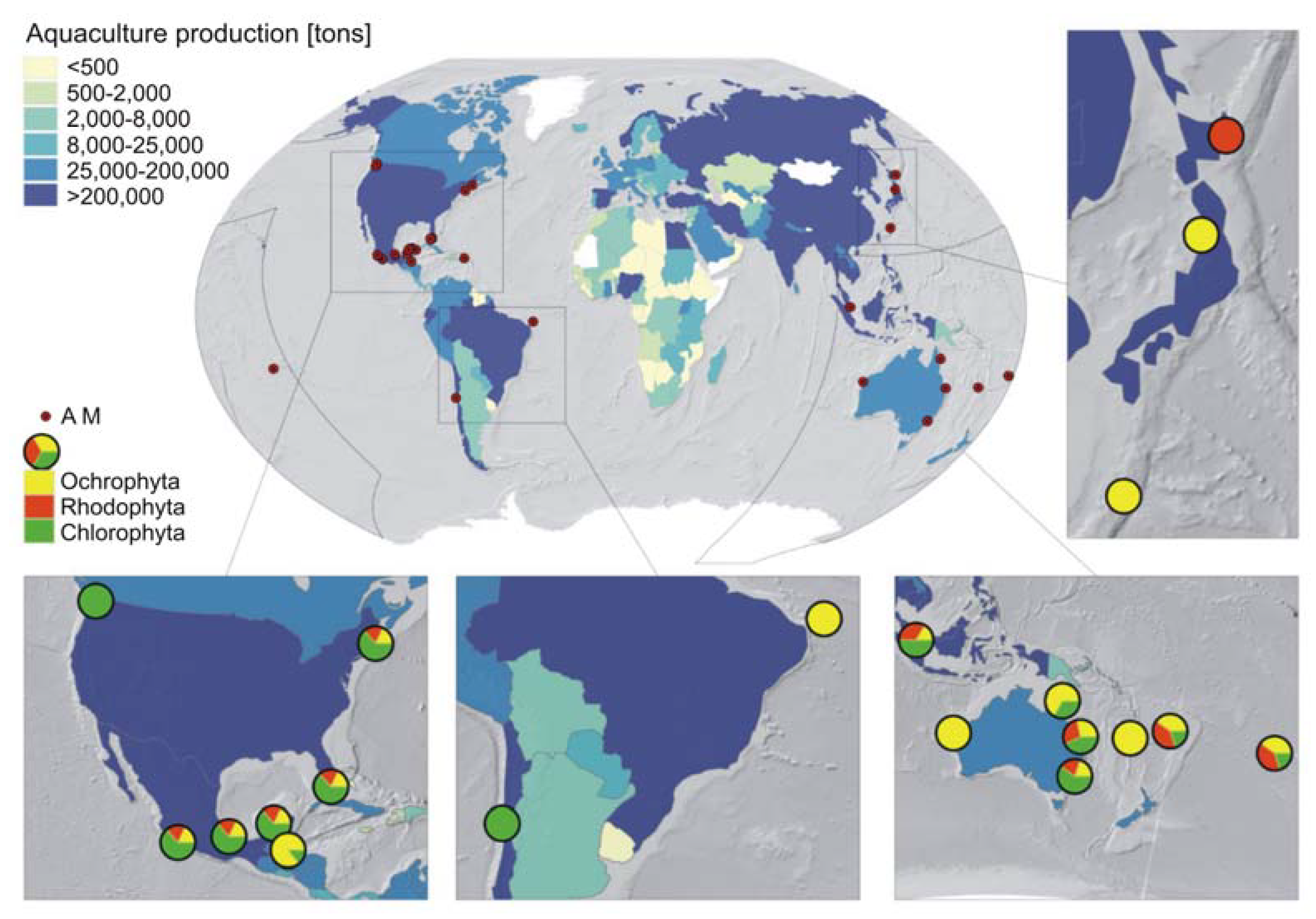
2. Distribution of Allelopathic Macroalgae and Aquaculture Locations
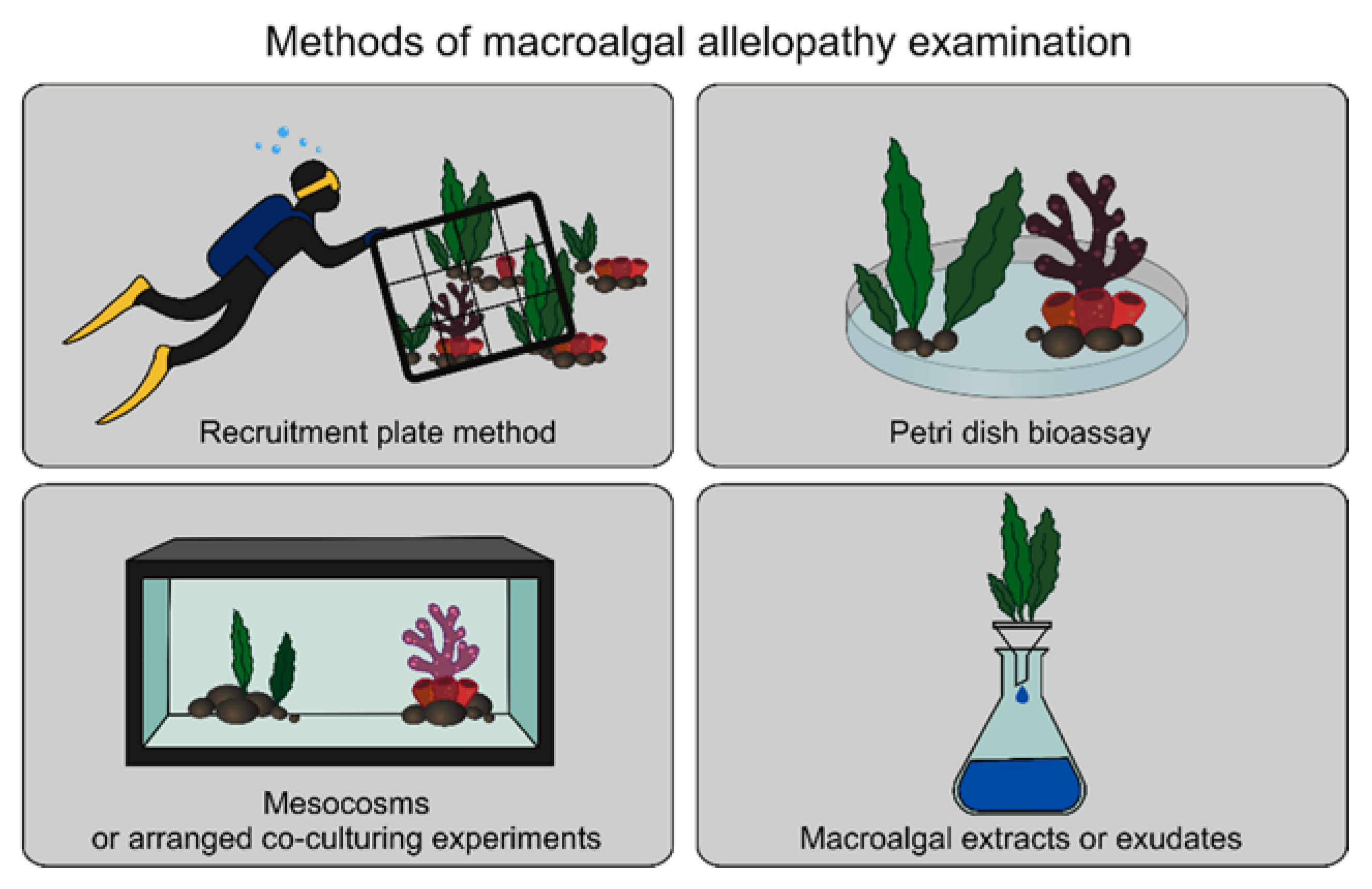
3. Methods for Macroalgal Allelopathy Examination
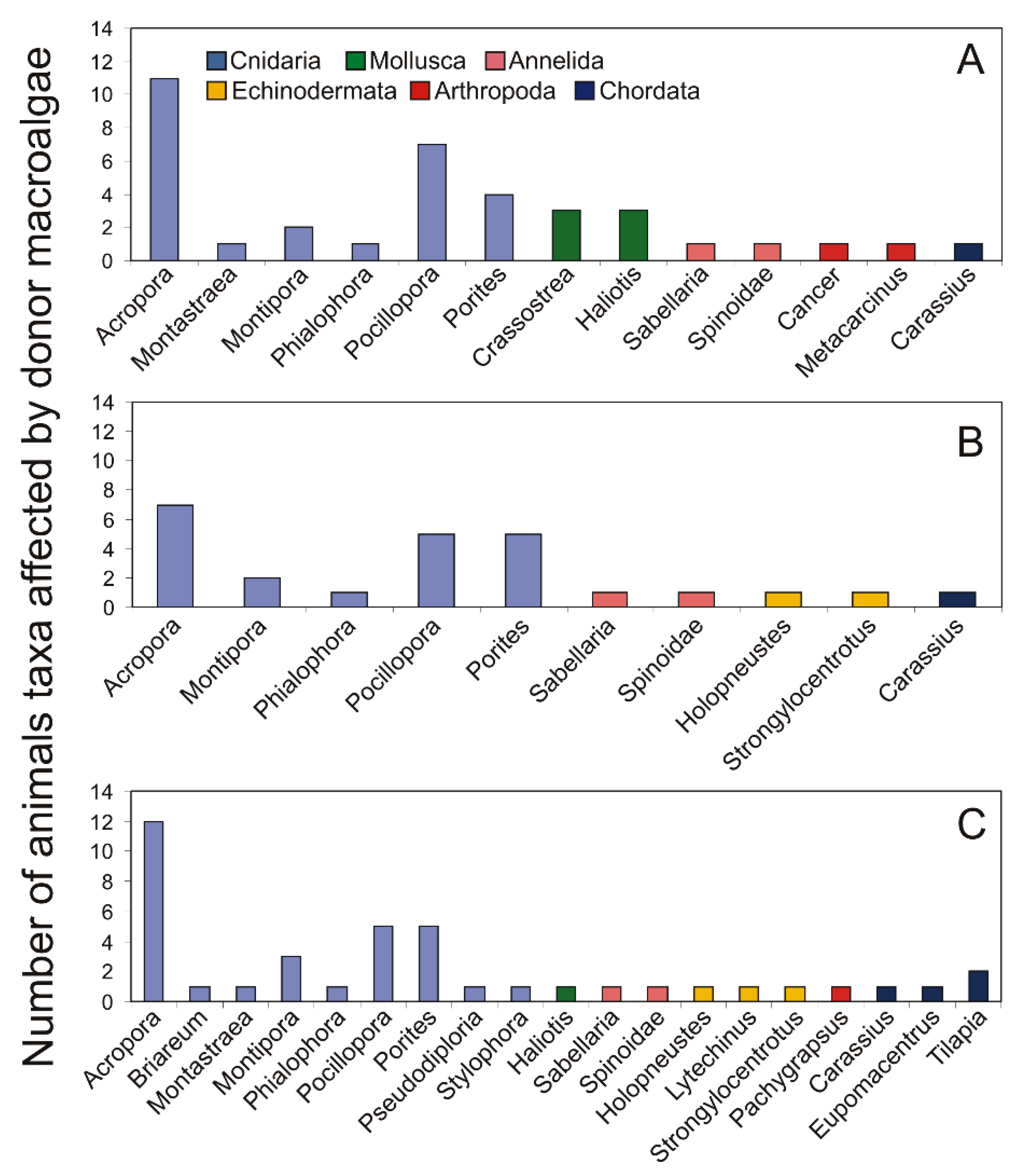
4. Taxonomic Position of Allelopathic Macroalgae and Their Impact on Coexisting Competitors and Herbivores
4.1. The Allelopathic Activity of Green Algae
4.2. The Allelopathic Activity of Red Algae
4.3. The Allelopathic Activity of Brown Algae
5. Allelopathic Compounds Produced by Macroalgae

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phylum/Species | Habitat | Compound | Activity | References |
|---|---|---|---|---|
| Green Algae (Chlorophyta) | ||||
| Avrainvillea nigricans | marine | Nigricanosides A–B | Antimitotic agent | Williams et al. [65] |
| Avrainvillea nigricans | marine | Hydroxyisoavrainvilleol | Protein tyrosine phosphate 1B inhibitors (PTP1B) | Colon et al. [66] |
| Avrainvillea rawsonii | marine | Isorawsonol | Cytotoxic and immunosuppressive activities | Chen et al. [67] |
| Bryopsis sp. | marine | Kahalalide F | Cytotoxic and immunosuppressive activities | Hamann and Scheuer [68] |
| Bryopsis sp. | marine | Kahalalide P | Cytotoxic and immunosuppressive activities | Dmitrenok et al. [69] |
| Caulerpa racemosa | marine | Sulfoquinovosyldiacylglycerol | Antiviral activity | Wang et al. [70] |
| Caulerpa taxifolia | marine | Caulerpals A–B | Anti-fungal activity | Aguilar-Santos [71] |
| Chara globularis | freshwater | Charamin | Antibiotic activity | Anthoni et al. [63] |
| Chara globularis | freshwater | Dithiolane, Trithiane | Antialgal activity | Wium-Andersen et al. [61] |
| Cladophora glomerata | freshwater | Gallic acid, Chlorogenic acid, Syringic acid, p-coumaric acid, Myricetin, 3,4-dihydroxybenzoic acid, Vanillic acid, 4-hydroxybenzoic acid, Rutin | Unknown | Korzeniowska et al. [64] |
| Codium iyengarii | marine | Lyengaroside | Antibacterial activity | Ali et al. [72] |
| Cymopolia barbata | marine | Cymobarbatol, 4-isocymobarbatol | Antimutagenic activity | Wall et al. [73] |
| Halimedatuna, Halimedaopuntia, Halimedaincrassata, Halimedasimulans, Halimedascabra, Halimedacopiosa | marine | Halimedatrial | Cytotoxic and antimicrobial activities | Paul and Fenical [60] |
| Halimeda tuna | marine | Halitunal | Antibacterial activity | Koehn et al. [74] |
| Halimeda sp. | marine | Halimedatrial | Antimicrobial and cytotoxic properties | Paul and Fenical [75] |
| Penicillus capitatus | marine | Capisterones A–B | Anti-fungal activity | Puglisi et al. [76] |
| Tydemania expeditionis | marine | Cycloartenol disulfates | Cytotoxic and immunosuppressive activities | Govindan et al. [77] |
| Ulva (Enteromorpha) intestinals | marine | Penostatins A–H | Cytotoxic and immunosuppressive activities | Takahashi et al. [78], Iwamoto et al. [79,80] |
| Ulva (Enteromorpha) intestinalis | marine | Cytochalasans, penochalasins A–H | Cytotoxic activity | Numata et al. [81] |
| Ulva (Enteromorpha) intestinalis | marine | Chaetoglobosin | Cytotoxic activity | Iwamoto et al. [82] |
| Ulva (Enteromorpha) intestinals | marine | Communesins A–B | Cytotoxic and immunosuppressive activities | Numata et al. [83] |
| Ulva lactuca | marine | 3-0-β-d-glucopyranosy-lstigmasta-5,25-diene | Anti-inflammatory substances | Awad et al. [84] |
| Ulvaria obscura | marine | Dopamine | Feeding-deterrent substances | Tocher and Craigie [59], Van Alstyne et al. [46] |
| Red Algae (Rhodophyta) | ||||
| Beckerella (Gelidium) subcostatum | marine | Bromo- beckerelide, epimer, chlorobeckerelide | Antimicrobial activity | Ohta [85] |
| Callophycus serratus | marine | Bromophycolides A–B | Cytotoxic activity | Kubanek et al. [86] |
| Callophycus serratus | marine | Bromophycolides C–I | Cytotoxic activity | Kubanek et al. [87] |
| Callophycus serratus | marine | Callophycoic acids A–H, diterpene-phenols, callophycols A–B | Antibacterial, antimalarial, anti-tumor and antifungal activity | Lane et al. [88] |
| Chondria armata | marine | Isodomic acid A–C | Insecticidal activity | Maeda et al. [89] |
| Chondria atropurpurea | marine | Chondriamide C, 3-indolacrylamide | Anthelmintic activity | Davyt et al. [90] |
| Chondria oppositiclada | marine | Cycloeudesmol | Antibacterial activity | Fenical and Sims [91] |
| Delisea pulchra | marine | Floridoside-isethionic acid complex | Induction of animal metamorphosis | Williamson et al. [33] |
| Digenea simplex | marine | α-alko-kainic acid | Neurophysiological activity | Biscoe et al. [92], Ferkany and Coyle [93] |
| Gracilaria asiatica | marine | Cerebroside gracilarioside, ceramides gracilamides A–B | Cytotoxic activity | Sun et al. [94] |
| Gigartina tenella | marine | Sulquinovosyldiacylglycerol: KM043 | Antiviral activity | Ohata et al. [95] |
| Jania rubens | marine | Deoxyparguerol-7-acetate | Anthelmintic activity | Awad [96] |
| Laurencia brongniartii | marine | Polybromoindoles | Antimicrobial activity, cytotoxic activity | Carter et al. [97], El Gamal et al. [98] |
| Laurencia brongniartii | marine | Brominated indoles | Antibacterial activities | Carter et al. [97] |
| Laurencia elata | marine | Elatol | Antibacterial activities | Sims [99] |
| Laurencia obtusa | marine | Teurilene, thyrsiferyl 23-acetate | Cytotoxic activity | Suzuki et al. [100] |
| Laurencia obtusa | marine | 3,7-dihydroxydihydrolaurene, perforenol B | Cytotoxic activity | Kladi et al. [101] |
| Laurencia obtusa | marine | Neorogioldiol B, prevezol B–D | Cytotoxic activity | IIopoulou et al. [102] |
| Laurencia obtusa | marine | Iso-obtusol | Antibacterial activities | Gonzalez et al. [103,104] |
| Laurencia obtusa | marine | Sesquiterpene | Antimalarial activity | Topeu et al. [105] |
| Laurencia pinnatifida | marine | Dehydrothyrsiferol, thyresenol A and B | Cytotoxic activity | Norte et al. [106], Pec et al. [107] |
| Laurancia pinnata | marine | Laurepinacine, isolaurepinnacin | Insecticidal activity | Fukuzawa and Masamune [108] |
| Laurencia mariannensis | marine | Brominated diterpene, 10-hydroxykahukuene B, 9-deoxyelatol, isoda-ctyloxene A, C15-acetogenin, laurenmariallene, sesquiterpenes | Antibacterial activities | Gonzalez et al. [109] |
| Laurencia nidifica | marine | Laurinterol, isolaurinterol, aplysin, α-bromocuparene | Insecticidal and repellent activities | Ishii et al. [110] |
| Laurencia nipponica | marine | (Z)-Laureatin, (Z)-isolaureatin, deoxyprepacifenol | Insecticidal activity | Watanabe et al. [111], El Sayed et al. [112] |
| Laurencia okamurae | marine | Laurinterol | Cytotoxic activity | Moon-Moo et al. [113] |
| Laurencia scoparia | marine | β-bisabolene sesquiterpenes | Anthelmintic activity | Davyt et al. [114] |
| Laurencia tristicha | marine | Cholest-5-en-3β,7α-diol Debromoepiaplysinol | Cytotoxic activity | Sun et al. [115] |
| Laurencia venusta | marine | Venustatriol | Antiviral activity | Sakemi et al. [116] |
| Laurencia yonaguniensis | marine | Neoirietetraol | Cytotoxic activity | Takahashi et al. [117] |
| Lophocladia sp. | marine | Lophocladine B | Cytotoxic activity | Gross et al. [118] |
| Murrayella periclados | marine | 12S-hydroxyeicosapentaenoic acid | Lipooxygenase inhibitor | Bernari and Gerwick [119] |
| Odonthalia corymbifera | marine | Bromophenols | Inhibition of isocitrate lyase enzyme | Lee et al. [120] |
| Peyssonnelia sp. | marine | Avarol | Antiviral activity | Talpir et al. [121] |
| Plocamium corallorhiza | marine | Plocaralides B–C | Cytotoxic activity | Knott et al. [122] |
| Plocamium telfairiae | marine | Telfairine | Insecticidal activity | Watanabe et al. [123] |
| Ptilota filicina | marine | Ptiollodene | Lipo-oxygenase inhibitor | Lopez and Gerwick [124] |
| Symphyocladia latiuscula | marine | Tasipeptins A–B | Aldose reductase inhibitors activity | Wang et al. [125] |
| Vidalia obtusiloba | marine | Vidalols A–B | Anti-inflammatory activity | Wiemer et al. [126] |
| Brown Algae (Ochrophyta) | ||||
| Chondria oppositiclada | marine | Cycloeudesmol | Antibacterial activity | Fenical and Sims [91] |
| Cystoseira crinita | marine | Meroterpenoids | Free radical scavenger and antioxidant activities | Fisch et al. [127] |
| Cystoseira myrica | marine | Hydroazulene diterpenes | Cytotoxic activity | Ayyad et al. [128] |
| Cystoseira tamariscifolia | marine | Methoxybifurcarenone | Antifungal and antibacterial activity | Bennamara et al. [129] |
| Cystophora siliquosa | marine | Cystophorene | Sperm-attractants pheromone | Muller et al. [130] |
| Dictyopteris undulata | marine | Yahazunol | Antimicrobial activity | Ochi et al. [131] |
| Dictyopteris undulata | marine | Cyclozonarone | Feeding-deterrent activity | Kurata et al. [132] |
| Dictyopteris zonarioides | marine | Zonarol, isozonarol | Antifungal activity | Fenical et al. [133] |
| Dictyota pfaffi | marine | 10,18-diacetoxy—8-hydroxy 2,6-dollabeladiene (dolabellane 1) | Antiviral activity | Barbosa et al. [35,134] |
| Dictyota spinulosa | marine | Hydroxydictyodial | Feeding-deterrent substances | Tanaka and Higa [32] |
| Dictyota sp. | marine | Dolabellane diterpenes | Cytotoxic activity | Tringali et al. [135] |
| Dilophus okamurae | marine | Dictyterepenoids A–B | Antifeedent activity | Suzuki et al. [34] |
| Ecklonia cava | marine | Fucodiphlorethol G | Antioxidant activity | Ham et al. [136] |
| Ecklonia stolonifera | marine | Phloroglucinol, eckstolonol, eckol, phlorofucofuroeckol A, dieckol | Hepatoprotective activity | Kang et al. [137] |
| Giffordia mitchelliae | marine | Giffordene | Gamete-attracting pheromone | Boland et al. [138] |
| Hizikia fusiformis | marine | Arsenic-containing ribofuranosides | Cytotoxic activity | Edmonds et al. [139] |
| Hormosira banksii | marine | Hormosirene | Sperm-attractants pheromone | Muller et al. [130] |
| Leptosphaeria sp. | marine | Leptosins M, MI, N, N1 | Cytotoxic activity | Yamada et al. [140] |
| Lobophora variegata | marine | Lobophorolide | Antifungal activity | Kubanek et al. [141] |
| Notheia anomala | marine | cis dihydroxyte-trahydrofuran | Nematocidal activity | Capon et al. [142] |
| Osmundaria serrata | marine | Lanosol enol ether | Antifungal and antibacterial activity | Barreto and Meyer [143] |
| Perithalia caudata | marine | Caudoxirene | Gamete-releasing, gamete-attracting pheromone | Muller et al. [144] |
| Pelvetia siliquosa | marine | Fucosterol | Anti-diabetic activity | Lee et al. [145] |
| Sargassum siliquastrum | marine | Sargachromanols A–P | Antioxidant activity | Jang et al. [146] |
| Sargassum tortile | marine | Dihydroxysargaquinone | Cytotoxic activity | Numata et al. [147] |
| Sargassum tortile | marine | Hydroxysargaquinone, sargasal-I-II | Cytotoxic activity | Numata et al. [148] |
| Sargassum thunbergii | marine | Thunbergols A–B | Scavenging activities, antioxidant activity | Seo et al. [149] |
| Sargassum thunbergii | marine | Sargothunbergol A | Antioxidant activity | Seo et al. [150] |
| Sargassum thunbergii | marine | Diacylglycerols | Antifungal activity | Kim et al. [151] |
| Stypopodium flabelliforme | marine | Isoepitaondiol | Insecticidal activity | Rovirosa et al. [152] |
| Stypopodium zonale | marine | Stypolactone | Cytotoxic activity | Dorta et al. [153] |
| Stypopodium zonale | marine | Stypotriol, stypoldione | Ichthyotoxic activity | Gerwick et al. [30] |
| Stypopodium zonale | marine | Stypoquinonic acid, taondiol, atomaric acid | Antimicrobial activity | Wessels et al. [154] |
| Stypopodium zonale | marine | Stypoldione, stypotriol, stypodiol, epistypodiol, epitaondiol | Ichthyotoxic activity, cytotoxic activity | Gerwick and Fenical [31] |
| Taonia atomaria | marine | Taondiol | Antimicrobial activity, cytotoxic activity | Othmani et al. [155] |
| Taonia atomaria | marine | Tetraprenyl benzoquinone sargaquinone | Anti-inflammatory activity | Tziveleka et al. [156] |
| Taonia atomaria | marine | Meroditerpenes atomarianones A–B | Cytotoxic activity | Abatis et al. [157] |
| Turbinaria ornata | marine | Turbinaric acid | Cytotoxic activity | Asari et al. [158] |
6. The Limitation of Macroalgae-Herbivores Interactions
7. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Toufique, K.A.; Belton, B. Is Aquaculture Pro-Poor? Empirical Evidence of Impacts on Fish Consumption in Bangladesh. World Dev. 2014, 64, 609–620. [Google Scholar] [CrossRef] [Green Version]
- Subasinghe, R. World aquaculture 2015: A brief overview; Food and Agriculture Organization of the United Nations: Rome, Italy, 2017. [Google Scholar]
- Van Ginneken, V.; de Vries, E. Towards a Seaweed Based Economy: The Global Ten Billion People Issue at the Midst of the 21st Century. J. FisheriesSciences. Com 2016, 10, 001–011. [Google Scholar]
- Oyinlola, M.A.; Reygondeau, G.; Wabnitz, C.C.; Troell, M.; Cheung, W.W. Global estimation of areas with suitable environmental conditions for mariculture species. PLoS ONE 2018, 13, e0191086. [Google Scholar] [CrossRef]
- Ben-Ari, T.; Neori, A.; Ben-Ezra, D.; Shauli, L.; Odintsov, V.; Shpigel, M. Management of Ulva lactuca as a biofilter of mariculture effluents in IMTA system. Aquaculture 2014, 434, 493–498. [Google Scholar] [CrossRef]
- Hurd, C.L. Seaweed Ecology and Physiology, 2nd ed.; Cambridge University Press: Cambridge, UK, 2015. [Google Scholar]
- Carl, C.; de Nys, R.; Paul, N.A. The seeding and cultivation of a tropical species of filamentous Ulva for algal biomass production. PLoS ONE 2014, 9, e98700. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Hernández, S.; Lozano-Ramírez, C.; Rodríguez-Palacio, M. Influence of the Habitat on Marine Macroalgae Toxicity. Annu. Res. Rev. 2019, 33, 1–9. [Google Scholar] [CrossRef]
- Harlin, M.M.; Rice, E.L. Allelochemistry in marine macroalgae. Crit. Rev. Plant Sci. 1987, 5, 237–249. [Google Scholar] [CrossRef]
- Lewis, W.M., Jr. Evolutionary interpretations of allelochemical interactions in phytoplankton algae. Am. Nat. 1986, 127, 184–194. [Google Scholar] [CrossRef]
- Kersen, P. Red Seaweeds Furcellaria Lumbricalis and Coccotylus Truncatus: Community Structure, Dynamics and Growth in the Northern Baltic Sea. Ph.D. Thesis, Tallinn University, Tallinn, Estonia, 2013. [Google Scholar]
- Gross, E.M. Allelopathy of aquatic autotrophs. Crit. Rev. Plant Sci. 2003, 22, 313–339. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.; Dao, G.; Tao, Y.; Zhan, X.; Hu, H. A review on control of harmful algal blooms by plant-derived allelochemicals. J. Hazard. Mater. 2020, 401, 123403. [Google Scholar] [CrossRef]
- Rybak, A.S. Ecological preferences of freshwater Ulva flexuosa (Ulvales; Ulvophyceae): Development of macroalgal mats in a Tulce fishpond (Wielkopolska Region, Poland). Oceanol. Hydrobiol. Stud. 2016, 45, 100–111. [Google Scholar] [CrossRef]
- Rybak, A.S. Freshwater macroalga, Ulva pilifera (Ulvaceae, Chlorophyta) as an indicator of the trophic state of waters for small water bodies. Ecol. Indic. 2021, 121, 106951. [Google Scholar] [CrossRef]
- Rybak, A.S.; Gąbka, M. The influence of abiotic factors on the bloom-forming alga Ulva flexuosa (Ulvaceae, Chlorophyta): Possibilities for the control of the green tides in freshwater ecosystems. J. Appl. Phycol. 2018, 30, 1405–1416. [Google Scholar] [CrossRef] [Green Version]
- Budzałek, G.; Śliwińska-Wilczewska, S.; Latała, A. Allelopathic effect of Ulva intestinalis L. on the Baltic filamentous cyanobacterium Nostoc sp. AUPC Studia Naturae 2018, 262, 80–89. [Google Scholar]
- Złoch, I.; Śliwińska-Wilczewska, S.; Kucharska, M.; Kozłowska, W. Allelopathic effects of Chara species (C. aspera, C. baltica, and C. canescens) on the bloom-forming picocyanobacterium Synechococcus sp. Environ. Sci. Pollut. Res. 2018, 25, 36403–36411. [Google Scholar] [CrossRef]
- Guiry, M.D.; Guiry, G.M. AlgaeBase World-Wide Electronic Publication. National University of Ireland: Galway, Ireland. Available online: algaebase.org (accessed on 3 November 2020).
- Gomes, M.P.; Garcia, Q.S.; Barreto, L.C.; Pimenta, L.P.S.; Matheus, M.T.; Figueredo, C.C. Allelopathy: An overview from micro-to macroscopic organisms, from cells to environments, and the perspectives in a climate-changing world. Biologia 2017, 72, 113–129. [Google Scholar] [CrossRef]
- Tanner, J.E. Competition between scleractinian corals and macroalgae: An experimental investigation of coral growth, survival and reproduction. J. Exp. Mar. Biol. Ecol. 1995, 190, 151–168. [Google Scholar] [CrossRef]
- Rasher, D.B.; Stout, E.P.; Engel, S.; Kubanek, J.; Hay, M.E. Macroalgal terpenes function as allelopathic agents against reef corals. Proc. Natl. Acad. Sci. USA 2011, 108, 17726–17731. [Google Scholar] [CrossRef] [Green Version]
- Bonaldo, R.M.; Hay, M.E. Seaweed-coral interactions: Variance in seaweed allelopathy, coral susceptibility, and potential effects on coral resilience. PLoS ONE 2014, 9, e85786. [Google Scholar] [CrossRef] [PubMed]
- Ritson-Williams, R.; Ross, C.; Paul, V.J. Elevated temperature and allelopathy impact coral recruitment. PLoS ONE 2016, 11, e0166581. [Google Scholar] [CrossRef]
- Del Monaco, C.; Hay, M.E.; Gartrell, P.; Mumby, P.J.; Diaz-Pulido, G. Effects of ocean acidification on the potency of macroalgal allelopathy to a common coral. Sci. Rep. 2017, 7, 998–1008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Longo, G.O.; Hay, M.E. Seaweed allelopathy to corals: Are active compounds on, or in, seaweeds? Coral Reefs 2017, 36, 247–253. [Google Scholar] [CrossRef]
- Ritson-Williams, R.; Arnold, S.N.; Paul, V.J. The impact of macroalgae and cyanobacteria on larval survival and settlement of the scleractinian corals Acropora palmata, A. cervicornis and Pseudodiploria strigosa. Mar. Biol. 2020, 167, 31. [Google Scholar] [CrossRef]
- Lee, C.S.; Walford, J.; Goh, B.P.L. The effect of benthic macroalgae on coral settlement. In Contributions to Marine Science: A Commemorative Volume Celebrating 10 Years of Research on St John’s Island; National University of Singapore: Singapore, 2012; pp. 89–93. [Google Scholar]
- Webster, F.J.; Babcock, R.C.; Van Keulen, M.; Loneragan, N.R. Macroalgae inhibits larval settlement and increases recruit mortality at Ningaloo Reef, Western Australia. PLoS ONE 2015, 10, e0124162. [Google Scholar] [CrossRef]
- Gerwick, W.H.; Fenical, W.; Fritsch, N.; Clardy, J. Stypotriol and stypoldione; ichthyotoxins of mixed biogenesis from the marine alga Stypodium zonale. Tetrahedron Lett. 1979, 2, 145–148. [Google Scholar] [CrossRef]
- Gerwick, W.H.; Fenical, W. Ichthyotoxic and cytotoxic metabolites of the tropical brown alga Stypopodium zonale (Lamouroux) Papenfuss. J. Org. Chem. 1981, 46, 22–27. [Google Scholar] [CrossRef]
- Tanaka, I.; Higa, T. Hydroxydictyodial, a new antifeedant diterpene from the brown alga Dictyota spinulosa. Chem. Lett. 1984, 2, 231–232. [Google Scholar] [CrossRef]
- Williamson, J.E.; De Nys, R.; Kumar, N.; Carson, D.G.; Steinberg, P.D. Induction of metamorphosis in the sea urchin Holopneustes purpurascens by a metabolite complex from the algal host Delisea pulchra. Biol. Bull. 2000, 198, 332–345. [Google Scholar] [CrossRef]
- Suzuki, M.; Yamada, H.; Kurata, K. Dictyterpenoids A and B two novel diterpenoids with feeding-deterrent activity from the brown alga Dilophus okamurae. J. Nat. Prod. 2002, 65, 121–125. [Google Scholar] [CrossRef]
- Barbosa, J.P.; Teixeira, V.L.; Pereira, R.C. A dolabellane diterpene from the brown alga Dictyota pfaffii as chemical defense against herbivores. Bot. Marina. 2004, 47, 147–151. [Google Scholar] [CrossRef]
- Ishii, T.; Okino, T.; Suzuki, M.; Machiguchi, Y. Tichocarpols A and B, Two Novel Phenylpropanoids with Feeding-Deterrent Activity from the Red Alga Tichocarpus c rinitus. J. Nat. Prod. 2004, 67, 1764–1766. [Google Scholar] [CrossRef]
- Morrow, K.M.; Bromhall, K.; Motti, C.A.; Munn, C.B.; Bourne, D.G. Allelochemicals produced by brown macroalgae of the Lobophora genus are active against coral larvae and associated bacteria, supporting pathogenic shifts to Vibrio dominance. Appl. Environ. Microbiol. 2017, 83, e02391-16. [Google Scholar] [CrossRef] [Green Version]
- Green-Gavrielidis, L.A.; MacKechnie, F.; Thornber, C.S.; Gomez-Chiarri, M. Bloom-forming macroalgae (Ulva spp.) inhibit the growth of co-occurring macroalgae and decrease eastern oyster larval survival. Mar. Ecol. Prog. Ser. 2018, 595, 27–37. [Google Scholar] [CrossRef]
- Fong, J.; Lim, Z.W.; Bauman, A.G.; Valiyaveettil, S.; Liao, L.M.; Yip, Z.T.; Todd, P.A. Allelopathic effects of macroalgae on Pocillopora acuta coral larvae. Mar. Environ. Res. 2019, 151, 104745. [Google Scholar] [CrossRef]
- Birrell, C.L.; McCook, L.J.; Willis, B.L.; Harrington, L. Chemical effects of macroalgae on larval settlement of the broadcast spawning coral Acropora millepora. Mar. Ecol. Prog. Ser. 2008, 362, 129–137. [Google Scholar]
- Warkus, E.L.; Wagstaff, M.; Morello, S.; Etter, R. Do Macroalgae Use Allelochemicals to Outcompete Invertebrates for Space in the Gulf of Maine? (Conference Material). 2010. Available online: https://cbs.asu.edu/sites/default/files/warkus_poster.pdf (accessed on 19 June 2021).
- Kuffner, I.B.; Walters, L.J.; Becerro, M.A.; Paul, V.J.; Ritson-Williams, R.; Beach, K.S. Inhibition of coral recruitment by macroalgae and cyanobacteria. Mar. Ecol. Prog. Ser. 2006, 323, 107–117. [Google Scholar] [CrossRef] [Green Version]
- Rasher, D.B.; Hay, M.E. Seaweed allelopathy degrades the resilience and function of coral reefs. Commun. Integr. Biol. 2010, 3, 564–566. [Google Scholar] [CrossRef]
- Chadwick, N.E.; Morrow, K.M. Competition among sessile organisms on coral reefs. In Coral Reefs: An Ecosystem in Transition; Springer: Dordrecht, The Netherlands, 2011; pp. 327–371. [Google Scholar]
- Roff, G.; Mumby, P.J. Global disparity in the resilience of coral reefs. Trends Ecol. Evol. 2012, 27, 404–413. [Google Scholar] [CrossRef]
- Van Alstyne, K.L.; Harvey, E.L.; Cataldo, M. Effects of dopamine, a compound released by the green-tide macroalga Ulvaria obscura (Chlorophyta), on marine algae and invertebrate larvae and juveniles. Phycologia 2014, 53, 195–202. [Google Scholar] [CrossRef]
- Nelson, T.A.; Lee, D.J.; Smith, B.C. Are “green tides” harmful algal blooms? Toxic properties of water-soluble extracts from two bloom-forming macroalgae, Ulva fenestrata and Ulvaria obscura (Ulvophyceae). J. Phycol. 2003, 39, 874–879. [Google Scholar] [CrossRef] [Green Version]
- Nelson, T.A.; Gregg, B.C. Determination of EC50 for normal oyster larval development in extracts from bloom-forming green seaweeds. Nautilus 2013, 127, 156–159. [Google Scholar]
- Huggett, M.J.; De Nys, R.; Williamson, J.E.; Heasman, M.; Steinberg, P.D. Settlement of larval blacklip abalone, Haliotis rubra, in response to green and red macroalgae. Mar. Biol. 2005, 147, 1155–1163. [Google Scholar] [CrossRef]
- Muñoz, P.; Ambler, R.; Bulboa, C. Settlement, Survival, and Post-Larval Growth of Red Abalone, Haliotis rufescens, on Polycarbonate Plates Treated with Germlings of Ulva sp. J. World Aquac. Soc. 2012, 43, 890–895. [Google Scholar] [CrossRef]
- Andras, T.D.; Alexander, T.S.; Gahlena, A.; Parry, R.M.; Fernandez, F.M.; Kubanek, J.; Wang, M.D.; Hay, M.E. Seaweed allelopathy against coral: Surface distribution of a seaweed secondary metabolite by imaging mass spectrometry. J. Chem. Ecol. 2012, 38, 1203–1214. [Google Scholar] [CrossRef] [Green Version]
- Morrow, K.M.; Ritson-Williams, R.; Ross, C.; Liles, M.R.; Paul, V.J. Macroalgal extracts induce bacterial assemblage shifts and sublethal tissue stress in Caribbean corals. PLoS ONE 2012, 7, e44859. [Google Scholar] [CrossRef] [Green Version]
- Rybak, A.S. Species of Ulva (Ulvophyceae, Chlorophyta) as indicators of salinity. Ecol. Indic. 2018, 85, 253–261. [Google Scholar] [CrossRef]
- Rybak, A.S. The Ulva flexuosa complex (Ulvaceae, Chlorophyta): An updated identification key with special reference to the freshwater and hyperhaline taxa. Phytotaxa 2018, 345, 83–103. [Google Scholar] [CrossRef]
- Paul, V.J.; Kuffner, I.B.; Walters, L.J.; Ritson-Williams, R.; Beach, K.S.; Becerro, M.A. Chemically mediated interactions between macroalgae Dictyota spp. and multiple life-history stages of the coral Porites astreoides. Mar. Ecol. Prog. Ser. 2011, 426, 161–170. [Google Scholar] [CrossRef]
- Olsen, K.; Ritson-Williams, R.; Paul, V.J.; Ross, C. Combined effects of macroalgal presence and elevated temperature on the early life-history stages of a common Caribbean coral. Mar. Ecol. Prog. Ser. 2014, 509, 181–191. [Google Scholar] [CrossRef]
- Vieira, C.; Thomas, O.P.; Culioli, G.; Genta-Jouve, G.; Houlbreque, F.; Gaubert, J.; De Clerc, O.; Payri, C.E. Allelopathic interactions between the brown algal genus Lobophora (Dictyotales, Phaeophyceae) and scleractinian corals. Sci. Rep. 2016, 6, 18637. [Google Scholar] [CrossRef] [Green Version]
- El Gamal, A.A. Biological importance of marine algae. Saudi. Pharm. J. 2010, 18, 1–25. [Google Scholar] [CrossRef] [Green Version]
- Tocher, R.D.; Craigie, J.S. Enzymes of marine algae: II. Isolation and identification of 3-hydroxytyramine as the phenolase substrate in Monostroma fuscum. Can. J. Bot. 1966, 44, 605–608. [Google Scholar] [CrossRef]
- Paul, V.J.; Fenical, W. Isolation of halimedtrial: Chemical defense adaptation in the calcareous reef-building alga Halimeda. Science 1983, 221, 747–749. [Google Scholar] [CrossRef]
- Wium-Andersen, S.; Anthoni, U.; Christophersen, C.; Houen, G. Allelopathic effects on phytoplankton by substances isolated from aquatic macrophytes (Charales). Oikos 1982, 39, 187–190. [Google Scholar] [CrossRef]
- Wium-Andersen, S.; Anthoni, U.; Houen, G. Elemental sulphur, a possible allelopathic compound from Ceratophyllum demersum. Phytochemistry 1983, 22, 2613. [Google Scholar] [CrossRef]
- Anthoni, U.; Nielsen, P.H.; Smith-Hansen, L.; Wium-Andersen, S.; Christophersen, C. Charamin, a quaternary ammonium ion antibiotic from the green alga Chara globularis. J. Org. Chem. 1987, 52, 694–695. [Google Scholar] [CrossRef]
- Korzeniowska, K.; Łęska, B.; Wieczorek, P.P. Isolation and determination of phenolic compounds from freshwater Cladophora glomerata. Algal. Res. 2020, 48, 101912. [Google Scholar] [CrossRef]
- Williams, D.E.; Sturgeon, C.M.; Roberge, M.; Andersen, R.J. Nigricanosides A and B antimitotic glycolipids isolated from the green alga Avrainvillea nigricans collected in Dominica. J. Am. Chem. Soc. 2007, 129, 5822–5823. [Google Scholar] [CrossRef]
- Colon, M.; Guevara, P.; Gerwick, W.H.; Ballantine, D. 50-Hydroxyisoavrainvilleol, a new diphenylmethane derivative from the tropical green alga Avrainvillea nigricans. J. Nat. Prod. 1987, 50, 368–374. [Google Scholar] [CrossRef]
- Chen, I.L.; Gerwick, W.H.; Schatzman, R.; Laney, M. Isorawsonol and related IMO dehydrogenase inhibitors from the tropical alga Avrainvillea rawsoni. J. Nat. Prod. 1994, 57, 947–952. [Google Scholar] [CrossRef]
- Hamann, M.T.; Scheuer, P. J Kahalalide F: A bioactive depsipeptide from the sacoglossan mollusk Elysia rufescens and the green alga Bryopsis sp. J. Am. Chem. Soc. 1993, 115, 5825–5826. [Google Scholar] [CrossRef]
- Dmitrenok, A.; Iwashita, T.; Nakajima, T.; Sakamoto, B.; Namikoshi, M.; Nagai, H. New cyclic desipeptides from the green alga species; application of a carboxypeptidase hydrolysis reaction to the structure determination. Tetrahedron 2006, 62, 1301–1308. [Google Scholar] [CrossRef]
- Wang, H.; Li, Y.-L.; Shen, W.-Z.; Rui, W.; Ma, X.-J.; Cen, Y.-Z. Antiviral activity of a sulfoquinovosyldiacylglycerol (SQDG) compound isolated from the green alga Caulerpa racemosa. Bot. Marina. 2007, 50, 185–190. [Google Scholar] [CrossRef]
- Aguilar-Santos, G. Caulerpin, a new red pigment from green algae of the genus Caulerpa. J. Chem. Soc. C 1970, 6, 842–843. [Google Scholar] [CrossRef]
- Ali, M.S.; Saleem, M.; Yammdagni, R.; Ali, M.A. Steroid and antibacterial glycosides from marine green alga Codium iyengarii Borgesen. Nat. Prod. Lett. 2002, 16, 407–413. [Google Scholar] [CrossRef]
- Wall, M.E.; Wani, M.C.; Manikumar, G.; Taylor, H.; Gaetano, K.; Hughes, T.J.; Gerwick, W.H.; McPhail, A.T.; McPhail, D.R. Plant antimutagenic agents, structure and antimutagenic properties of cymobarbatol and 4-isocymobarbatol, new symbols from green alga Cymopolia barbata. J. Nat. Prod. 1989, 52, 1092–1099. [Google Scholar] [CrossRef]
- Koehn, F.E.; Gunasekera, S.P.; Niel, D.N.; Cross, S.S. Halitunal, an unusual diterpene Aldehyde from the marine alga Halimeda tuna. Tetrahedron Lett. 1991, 32, 169–172. [Google Scholar] [CrossRef]
- Paul, V.J.; Fenical, W. Novel bioactive diterpenoid metabolites from tropical marine algae of the genus Halimeda. Tetrahedron 1984, 40, 3053–3062. [Google Scholar] [CrossRef]
- Puglisi, M.P.; Tan, L.T.; Jensen, P.R.; Fenical, W. Capisterones A and B from the tropical green alga Penicillus capitatus: Unexpected anti-fungal defenses targeting the marine pathogen Lindra thallasiae. Tetrahedron 2004, 60, 7035–7039. [Google Scholar] [CrossRef]
- Govindan, M.; Abbas, S.A.; Schmitz, E.I.; Lee, R.H.; Papkoff, I.S.; Slate, D.L. New cycloartanol sulfates from the alga Tydemania expeditionis: Inhibitor of the protein tyrosin kinase pp60. J. Nat. Prod. 1994, 57, 74–78. [Google Scholar] [CrossRef]
- Takahashi, C.; Numata, A.; Yamada, T.; Minoura, K.; Enomoto, S.; Konishi, K.; Nakai, M.; Matsuda, C.; Nomoto, K. Penostatins, novel cytotoxic metabolites from a Penicillium species separated from a green alga. Tetrahedron Lett. 1997, 37, 655–658. [Google Scholar] [CrossRef]
- Iwamoto, C.; Minoura, K.; Hagishita, S.; Nomoto, K.; Numata, A. Penostatins F–I novel cytotoxic metabolites from a Penicillium species from an Enteromorpha marine alga. J. Chem. Soc. Perkin Trans. 1997, 3, 449–456. [Google Scholar] [CrossRef]
- Iwamoto, C.; Minoura, K.; Hagishita, S.; Oka, T.; Ohta, T.; Hagishita, S.; Numata, A. Absolute sterostructures of novel penostatins A–E from a Penicillium species from an Enteromorpha marine alga. Tetrahedron 1999, 55, 14353–14368. [Google Scholar] [CrossRef]
- Numata, A.; Takahashi, C.; Ito, Y.; Minoura, K.; Yamada, T.; Matsuda, C.; Nomoto, K. Penochalasins, a novel class of cytotoxic cytochalasans from a Penicillium species separated from a marine alga: Structure determination and solution conformation. J. Chem. Soc. Perkin Trans. 1996, 1, 239–245. [Google Scholar] [CrossRef]
- Iwamoto, C.; Yamada, T.; Ito, Y.; Minoura, K.; Numata, A. Cytotoxic cytochalasans from a Penicillium species separated from a marine alga. Tetrahedron 2001, 57, 2904–2997. [Google Scholar] [CrossRef]
- Numata, A.; Takahashi, C.; Ito, Y.; Takeda, T.; Kawai, K.; Osami, Y.; Matsumura, E.; Imachi, M.; Ito, T.; Hasegawa, T. Communesins, cytotoxic metabolites of a fungus isolated from a marine alga. Tetrahedron Lett. 1996, 34, 2355–2358. [Google Scholar] [CrossRef]
- Awad, N.E. Biologically active steroid from the green alga Ulva lactuca. Phytother. Res. 2000, 14, 641–643. [Google Scholar] [CrossRef]
- Ohta, K. Antimicrobial compounds in the marine red alga Beckerella subcostatum. Agric. Biol. Chem. 1977, 41, 2105–2106. [Google Scholar] [CrossRef] [Green Version]
- Kubanek, I.; Prusak, A.C.; Snell, T.W.; Giese, R.A.; Hardcastle, K.I.; Fairchild, C.R.; Aalbersberg, W.; Raventos-Suarez, C.; Hay, M.E. Antineoplastic diterpene-benzoate macrolides from the Fijian red alga Callophycus serratus. Org. Lett. 2005, 7, 261–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubanek, J.; Prusak, A.C.; Snell, T.W.; Giese, R.A.; Fairchild, C.R.; Aalbersberg, W.; Hay, M.E. Bromophycolides C-I from the Fijian red alga Callophycus serratus. J. Nat. Prod. 2006, 69, 731–735. [Google Scholar] [CrossRef] [Green Version]
- Lane, A.L.; Stout, E.P.; Hay, M.E.; Prusak, A.C.; Hardcastle, K.; Fairchild, C.R.; Franzblau, S.G.; Le Roch, K.; Prudhomme, J.; Aalbersberg, W.; et al. Callophycoic acids and callophycols from the Fijian red alga Callophycus serratus. J. Org. Chem. 2007, 72, 7343–7351. [Google Scholar] [CrossRef] [Green Version]
- Maeda, M.; Kodama, T.; Tanaka, T.; Yoshizumi, H.; Takemoto, T.; Nomoto, K.; Fujita, T. Structures of isodomic acids A, B and C novel insecticidal amino acids from the red alga Chonria armata. Chem. Pharm. Bull. 1986, 34, 4892–4895. [Google Scholar] [CrossRef] [Green Version]
- Davyt, D.; Entz, W.; Fernandez, R.; Mariezcurrena, R.; Mombru, A.W.; Saldana, I.; Dominguez, L.; Coil, J.; Manta, E. A new indol derivative from the red alga chondra atropurpurea isolation, structure determination, and anthelmintic activity. J. Nat. Prod. 1998, 61, 1560–1563. [Google Scholar] [CrossRef]
- Fenical, W.; Sims, J.J. Cycloeudesmol, an antibiotic cyclopropane conatinnin sequiterpene from the marine alga, Chondria oppositiclada Dawson. Tetrahedron Lett. 1974, 13, 1137–1140. [Google Scholar] [CrossRef]
- Biscoe, T.J.; Evans, R.H.; Headley, P.M.; Martin, M.; Watkins, J.C. Domic and quisqualic acids as potent amino acids excitants of frog and rat spinal neurons. Nature 1975, 255, 166–167. [Google Scholar] [CrossRef]
- Ferkany, J.W.; Coyle, J.T. Kainic acid selectively stimulates the release of endogenous excitatory acidic amino acids. J. Pharmacol. Exp. Therapeut. 1983, 225, 399–406. [Google Scholar]
- Sun, Y.; Xu, Y.; Liu, K.; Hua, H.; Zhu, H.; Pei, Y. Gracilarioside and gracilamides from the red alga Gracilaria asiatica. J. Nat. Prod. 2006, 69, 1488–1491. [Google Scholar] [CrossRef] [PubMed]
- Ohata, K.; Mizushina, Y.; Hirata, N.; Sugawara, F.; Mutsukage, A.; Yoshida, S.; Sakaguchi, K. Sulphoquinovosy-ldiacylglycerol, KM043 a new potent inhibitor of eukaryotic DNA polymerases and HIV-reverse transcriptase type from a marine red alga Gigartina tenella. Chem. Pharm. Bull. 1998, 46, 684–686. [Google Scholar] [CrossRef] [Green Version]
- Awad, N.E. Bioactive brominated diterpenes from the marine red alga Jania Rubens (L.) Lamx. Phytother Res. 2004, 18, 275–279. [Google Scholar] [CrossRef]
- Carter, G.T.; Rinehart, K.L., Jr.; Li, L.H.; Kuentzel, S.L. Brominated indoles from Laurencia brongniartii. Tetrahedron Lett. 1978, 19, 4479–4482. [Google Scholar] [CrossRef]
- El Gamal, A.A.; Wang, W.-L.; Duh, C.-Y. Sulfur-containing polybromoindoles from the Formosan red alga Laurencia brongniartii. J. Nat. Prod. 2005, 68, 815–817. [Google Scholar] [CrossRef]
- Sims, J.J.; Lin, G.H.Y.; Wing, R.M. Marine natural products, elatol, a halogenated sesquiterpene alcohol from the red alga Laurencia elata. Tetrahedron Lett. 1974, 39, 3487–3490. [Google Scholar] [CrossRef]
- Suzuki, T.; Furusaki, A.; Matsumoto, T.; Kato, A.; Lmanaka, Y.; Kurosawa, E. Teurilene and thyrsiferyl 23 acetate, meso and remarkably cytotoxic compounds from the marine red alga Laurencia obtuse. Tetrahedron Lett. 1985, 26, 1329–1332. [Google Scholar] [CrossRef]
- Kladi, M.; Xenaki, H.; Vagias, C.; Papazafiri, P.; Roussis, V. New cytotoxic sesquiterpenes from the red algae Laurencia obtusa and Laurencia microcladia. Tetrahedron 2006, 62, 182–189. [Google Scholar] [CrossRef]
- IIopoulou, D.; Mihopoulos, N.; Vigias, C.; Papazafiri, P.; Roussis, V. Novel cytotoxic brominated diterpenes from the red alga Laurencia obtuse. J. Org. Chem. 2003, 68, 7667–7674. [Google Scholar] [CrossRef]
- Gonzalez, A.G.; Darias, J.; Diaz, A.; Fournero, J.D.; Martin, J.D.; Perez, C. Evidence for the biogenesis of halogenated chamigrenes from the red alga Laurencia obtusa. Tetrahedron Lett. 1976, 17, 3051–3054. [Google Scholar] [CrossRef]
- Gonzalez, A.G.; Delgado, M.J.; Martin, V.S.; Martinez-Ripoll, M.; Fayos, J. X-ray study of sesquiterpene constituents of the alga Laurencia obtusa leads to structure revision. Tetrahedron Lett. 1979, 29, 2717–2718. [Google Scholar] [CrossRef]
- Topeu, G.; Aydogmus, Z.; Imre, S.; Goren, A.C.; Pezzuto, J.M.; Clement, J.A.; Kinghorn, D.G. Brominated sesquiterpenes from the red alga Laurencia obtusa. J. Nat. Prod. 2003, 66, 1505–1508. [Google Scholar]
- Norte, M.; Fernandez, J.J.; Saouto, M.L.; Gavin, J.A.; Garcia-Gravalos, M.D. Thyrsenols A and B two unusual polyether squalene derivatives. Tetrahedron 1997, 53, 3173–3178. [Google Scholar] [CrossRef]
- Pec, M.K.; Aguirre, A.; Moser-Their, K.; Fernandez, J.J.; Souto, M.L.; Dota, J.; Diaz-Gonzalez, F.; Villar, J. Induction of apoptosis in estrogen dependent and independent breast cancer cells by the marine terpenoid dehydrothyrsiferol. Biochem. Pharmacol. 2003, 65, 1451–1461. [Google Scholar] [CrossRef]
- Fukuzawa, A.; Masamune, T. Laurepinnacin and isolaurepinnacin: New acetylenic cyclic ethers from the marine alga Laurencia pinnata Yamada. Tetrahedron Lett. 1982, 22, 4081–4084. [Google Scholar] [CrossRef]
- Gonzalez, A.G.; Martin, J.D.; Martıin, V.S.; Norte, M.; Perez, R. Biomimetic approach to the synthesis of rhodolaureol and rhodolauradiol. Tetrahedron Lett. 1982, 23, 2395–2398. [Google Scholar] [CrossRef]
- Ishii, T.; Nagamine, T.; Nguyen, B.C.Q.; Tawata, S. Insecticidal and repellent activities of laurinterol from the Okinawan red alga Laurencia nidifica. Rec. Nat. Prod. 2017, 11, 63–68. [Google Scholar]
- Watanabe, K.; Umeda, K.; Miyakado, M. Isolation and identification of three insecticidal principles from the red alga Laurencia nipponica Yamada. Agric. Biol. Chem. 1989, 53, 2513–2515. [Google Scholar] [CrossRef]
- El Sayed, K.A.; Dunbar, D.C.; Perry, T.L.; Wilkins, S.P.; Hamann, M.T. Marine natural products as prototype insecticidal agents. J. Agric. Food Chem. 1997, 45, 2735–2739. [Google Scholar] [CrossRef]
- Moon-Moo, K.; Sang-Hoon, L.; Se-Kwon, K. Patent from PCT Int. Appl. WO 2009048195 A1 20090416, 2009. (Language: English, Database: CAPLUS).
- Davyt, D.; Fernandez, R.; Suescun, L.; Mombru, A.W.; Saldaña, J.; Domıínguez, L.; Fujii, M.T.; Manta, E. Bisabolanes from the red alga Laurencia scoparia. J. Nat. Prod. 2006, 69, 1113–1116. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Shi, D.Y.; Li, S.; Wang, S.J.; Han, L.J.; Fan, X.; Yang, Y.C.; Shi, J.G. Chemical constituents of the red alga Laurencia tristicha. J. Asian Nat. Prod. Res. 2007, 9, 725–734. [Google Scholar] [CrossRef]
- Sakemi, S.; Higa, T.; Jefford, C.W.; Bernardinelli, G. Venustatriol: A new antiviral triterpene tetracyclic ether from Laurencia venusta. Tetrahedron Lett. 1986, 27, 4287–4290. [Google Scholar] [CrossRef]
- Takahashi, Y.; Daitoh, M.; Suzuki, M.; Abe, T.; Masuda, M. Halogenated metabolites from the new Okinawan red alga Laurencia yonaguniensis. J. Nat. Prod. 2002, 65, 395–398. [Google Scholar] [CrossRef]
- Gross, H.; Goeger, D.E.; Hills, P.; Mooberry, S.L.; Ballantine, D.L.; Murray, T.F.; Valeriote, F.A.; Gerwick, W.H. Lophocladines, bioactive alkaloids from the red alga Lophocladia sp. J. Nat. Prod. 2006, 69, 640–644. [Google Scholar] [CrossRef] [Green Version]
- Bernari, M.W.; Gerwick, W.H. Eicosanoids from the tropical red alga Murrayella periclados. Phytochemistry 1994, 36, 1233–1240. [Google Scholar] [CrossRef]
- Lee, H.-S.; Lee, T.-H.; Lee, J.H.; Chae, C.-S.; Chung, S.-C.; Shin, D.-S.; Shin, J.; Oh, K.-B. Inhibition of the pathogenicity of Magnaporthe grisea by bromophenols, isocitrate lyase inhibitors, from the red alga Odonthalia corymbifera. J. Agric. Food Chem. 2007, 55, 6923–6928. [Google Scholar] [CrossRef] [PubMed]
- Talpir, R.; Rudi, A.; Kashman, Y.; Loya, Y.; Hizi, A. Three new sesquiterpene hydroquinones from marine origin. Tetrahedron 1994, 50, 4179–4184. [Google Scholar] [CrossRef]
- Knott, M.G.; Mkwanazi, H.; Arendse, C.E.; Hendricks, D.T.; Bolton, J.J.; Beukes, D.R. Plocoralides A–C, polyhalogenated monoterpenes from the marine alga Plocamium corallorhiza. Phytochemistry 2005, 66, 1108–1112. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, K.; Miyakado, M.; Ohno, N.; Okada, A.; Yanagi, K.; Moriguchi, K.A. A polyhalogenated isecticidal monoterepene from the red alga Plocamium telfairiae. Phytochemistry 1988, 28, 77–78. [Google Scholar] [CrossRef]
- Lopez, A.; Gerwick, H. Ptiollodene, a novel eicosanoid inhibitor of 5 lipoxygenase and Na+/K+ ATPase from the red marine alga Ptilota filicina. Tetrahedron Lett. 1988, 29, 1505–1506. [Google Scholar] [CrossRef]
- Wang, W.; Okada, Y.; Shi, H.; Wang, Y.; Okuyama, T. Tasipeptins A and B: Structures and aldose reductase inhibitory effects of bromophenols from the red alga Symphyocladia latiuscula. J. Nat. Prod. 2005, 68, 620–622. [Google Scholar] [CrossRef]
- Wiemer, D.E.; Idler, D.D.; Fenical, W. Vidalols A and B, new antiinflammatory bromophenols from the Caribbean marine red alga Vidalia obtusaloba. Experientia 1991, 47, 851–853. [Google Scholar] [CrossRef]
- Fisch, K.M.; Bohm, V.; Wrightand, A.D.; Konig, G.M. Antioxidative meroterpenoids from the brown alga Cystoseira crinita. J. Nat. Prod. 2003, 66, 968–975. [Google Scholar] [CrossRef]
- Ayyad, S.-E.N.; Abdel-Halim, O.B.; Shier, W.T.; Hoye, T.R. Cytotoxic hydroazulene diterpenes from the brown alga Cystoseira myrica. Z. Natuforsch. C. Biosci. 2003, 58, 33–38. [Google Scholar] [CrossRef]
- Bennamara, A.; Abourrichi, A.; Berrada, M.; Charrouf, M.H.; Chaib, N.; Boudouma, M.; Garneau, X.F. Methoxybifurcarenone: An antifungal and antibacterial meroditerpenoid from the brown alga Cystoseira tamariscifolia. Phytochemistry 1999, 52, 37–40. [Google Scholar] [CrossRef]
- Muller, D.G.; Clayton, M.N.; Gassmann, O.; Boland, W.; Marner, F.J.; Schottes, T.; Jaenicke, L. Cystophorene and hormosirene, sperm attractants in Australian brown algae. Naturwissenschatien 1985, 72, 97–99. [Google Scholar] [CrossRef]
- Ochi, M.; Kotsuki, H.; Muraoka, K.; Tokoroyama, T. The structure of yahazunol, a new sesquiterpene-substituted hydroquinone from the brown seaweed Dictyopteris undulata Okamura. Bull. Chem. Soc. Jpn. 1979, 52, 629–630. [Google Scholar] [CrossRef]
- Kurata, K.; Tanguchi, K.; Suzuki, M. Cyclozonarone, a sesquiterpene-substituted benzoquinone derivative from the brown alga Dictyopteris undulate. Phytochemistry 1996, 41, 749–752. [Google Scholar] [CrossRef]
- Fenical, W.; Sims, J.J.; Squatrito, D.; Wing, R.M.; Radlick, P. Marine natural products, VII Zonarol and isozonarol, fungitoxic hydroquinones from the brown seaweeds Dictyopteris zonarioides. J. Org. Chem. 1973, 38, 2383–2386. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, J.P.; Teixeira, V.L.; Villca, R.; Pereira, R.C.; Abrantes, J.L.; da Paixao Frugulhetti, I.C.P. A dolabellane diterpene from the Brazilian brown alga Dictyota pfaffii. J. Biochem. Syst. Ecol. 2003, 31, 1451–1453. [Google Scholar] [CrossRef]
- Tringali, C.; Prattellia, M.; Nicols, G. Structure and conformation of new diterpenes based on the dolabellane skeleton from Dictyota species. Tetrahedron 1984, 40, 703–799. [Google Scholar] [CrossRef]
- Ham, Y.M.; Baik, J.S.; Hyun, J.W.; Lee, N.H. Isolation of a new phlorotannin, fucodiphlorethol G, from a brown alga Ecklonia cava. Bull. Korean Chem. Soc. 2007, 28, 1595–1597. [Google Scholar]
- Kang, H.S.; Chang, H.Y.; Jung, J.H.; Son, B.W.; Choi, J.S. A new phlorotannin from the brown alga Ecklonia stolonifera. Chem. Pharm. Bull. 2003, 51, 1012–1014. [Google Scholar] [CrossRef] [Green Version]
- Boland, W.; Jaenicke, L.; Muller, D.G.; Gassmann, G. Giffordene, 2Z, 4Z, 6E, 8Z-undecatetraene, is the odoriferous principle of the marine brown alga Giffordia mitchellae. Experientia 1987, 43, 466–468. [Google Scholar] [CrossRef]
- Edmonds, S.l.; Morita, M.; Shibata, Y. Isolation and identification of Arsenic containing ribfurnaoside and inorganic Arsenic from Japanes edible seaweed Hizikia fusiforme. J. Chem. Soc. Perkin. Trans. I 1987, 577–580. [Google Scholar] [CrossRef]
- Yamada, T.; Iwamoto, C.; Yamagaki, N.; Yamanouchi, T.; Minoura, K.; Yamori, T.; Uehara, Y.; Andoh, T.; Umemura, K.; Numata, A. Leptosins M–N Cytotoxic metabolites from a Leptosphaeria species separated from a marine alga, structure determination and biological activities. Tetrahedron 2002, 58, 479–487. [Google Scholar] [CrossRef]
- Kubanek, J.; Jensen, P.R.; Keifer, P.A.; Sullards, M.C.; Collins, D.O.; Fenical, W. Seaweed resistance to microbial attack: A targeted chemical defense against marine fungi. Proc. Nat. Acad. Sci. USA 2003, 100, 6916–6921. [Google Scholar] [CrossRef] [Green Version]
- Capon, R.I.; Barrow, R.A.; Rochfort, S.; Jobling, M.; Skene, C.; Larcey, E.; Gill, I.H.; Friedel, T.; Wadsworth, D. Marine Nematodes: Tetrahydrofuran from a southern Australian brown alga, Notheia anomala. Tetrahdron 1998, 54, 2227–2242. [Google Scholar] [CrossRef]
- Barreto, M.; Meyer, J.J.M. Isolation and antimicrobial activity of a lanosol derivative from Osmundaria serrata (Rhodophyta) and a visual exploration of its biofilm covering. S. Afr. J. Bot. 2006, 72, 521–528. [Google Scholar] [CrossRef] [Green Version]
- Muller, D.G.; Boland, W.; Becker, U.; Wahl, T. Caudoxirene, the spermatozide-releasing and attracting factor in the marine brown alga Perithalia caudate (Phaeophyceae, Sporochnales). Biol. Chem. Hopper-Seyler 1988, 369, 655–659. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.S.; Shin, K.H.; Kim, B.K.; Lee, S. Anti-diabetic activities of fucosterol from Pelvetia siliquosa. Arch. Pharmacal. Res. 2004, 27, 1120–1122. [Google Scholar] [CrossRef]
- Jang, K.H.; Lee, H.B.; Choi, B.W.; Lee, H.-S.; Shin, J. Chromenes from the brown alga Sargassum siliquastrum. J. Nat. Prod. 2005, 68, 716–723. [Google Scholar] [CrossRef]
- Numata, A.; Kambara, S.; Takahashi, C.; Fujiki, R.; Yoneda, M.; Fujita, E.; Nabeshima, Y. Cytotoxic activity of marine algae and a cytotoxic principle of the brown alga Sargassum tortile. Chem. Pharm. Bull. 1991, 39, 2129–2131. [Google Scholar] [CrossRef] [Green Version]
- Numata, A.; Kanbara, S.; Takahashi, C.; Fujiki, R.; Yoneda, M.; Usami, Y.; Fujita, E. A cytotoxic principle of the brown alga Sargassum tortile and structures of chromenes. Phytochemistry 1992, 31, 1209–1213. [Google Scholar] [CrossRef]
- Seo, Y.; Park, K.E.; Kim, Y.A.; Lee, H.-I.; Yoo, I.-S.; Ahn, I.-W.; Lee, B.-J. Isolation of tetraprenyltoluquinols from the brown alga Sargassum thunbergii. Chem. Pharm. Bull. 2006, 54, 1730–1733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, Y.; Park, K.E.; Nam Bull, T.J. Isolation of a new chromene from the brown alga Sargassum thunbergii. Korean Chem. Soc. 2007, 28, 1831–1833. [Google Scholar]
- Kim, Y.H.; Kim, E.-H.; Lee, C.; Kim, M.-H.; Rho, J.-R. Two new monogalactosyl diacylglycerols from brown alga Sargassum thunbergii. Lipids 2007, 42, 395–399. [Google Scholar] [PubMed]
- Rovirosa, J.; Sepulveda, M.; Quezada, E.; San-Martin, A. Isoepitaondiol, a diterpenoid of Stypopodium flabelliforme and the insecticidal activity of stypotriol Epitaondiol and derivatives. Phytochemistry 1992, 31, 2679–2681. [Google Scholar] [CrossRef]
- Dorta, E.; Cueto, M.; Diaz-Marrero, A.R.; Darias, J. Stypolactone an interesting diterpenoid from the brown alga Stypopodium zonale. Tetrahedron Lett. 2002, 65, 9043–9046. [Google Scholar] [CrossRef]
- Wessels, M.; Konig, G.M.; Wright, A.D. A new tyrosine kinase inhibitor from the marine brown alga Stypopodium zonale. J. Nat. Prod. 1999, 62, 927–930. [Google Scholar] [CrossRef]
- Othmani, A.; Bunet, R.; Bonnefont, J.L.; Briand, J.F.; Culioli, G. Settlement inhibition of marine biofilm bacteria and barnacle larvae by compounds isolated from the Mediterranean brown alga Taonia atomaria. J. Appl. Physiol. 2016, 28, 1975–1986. [Google Scholar] [CrossRef]
- Tziveleka, L.-A.; Abatis, D.; Paulus, K.; Bauer, R.; Vigias, C.; Roussis, V. Marine polyprenylated hydroquinones, quinones, and chromenols with inhibitory effects on leukotriene formation. Chem. Biol. 2005, 2, 901–909. [Google Scholar] [CrossRef]
- Abatis, D.; Vigias, C.; Galanakis, D.; Norris, J.N.; Moreau, D.; Roussakis, C.; Rousis, V. Atomarianones A and B: Two cytotoxic meroditerpenes from the brown alga Taonia atomaria. Tetrahedron Lett. 2005, 46, 8525–8529. [Google Scholar] [CrossRef]
- Asari, F.; Kusumi, T.; Kakisawa, H. Turbinaric acid, a cytotoxic secosqualene carboxylic acid from the brown alga Turbinaria ornate. J. Nat. Prod. 1989, 52, 1167–1169. [Google Scholar] [CrossRef] [PubMed]
- Céspedes, C.L.; Avila, J.G.; Martínez, A.; Serrato, B.; Calderón-Mugica, J.C.; Salgado-Garciglia, R. Antifungal and antibacterial activities of Mexican tarragon (Tagetes lucida). J. Agric. Food Chem. 2006, 54, 3521–3527. [Google Scholar] [CrossRef]
- Li, X.; Wang, J.; Huang, D.; Wang, L.; Wang, K. Allelopathic potential of Artemisia frigida and successional changes of plant communities in the northern China steppe. Plant Soil 2011, 341, 383–398. [Google Scholar] [CrossRef]
- Talukdar, D. Allelopathic effects of Lantana camara L. on Lathyrus sativus L.: Oxidative imbalance and cytogenetic consequences. Allelopath. J. 2013, 31, 71–90. [Google Scholar]
- Yu, Z.W.; Sun, W.H.; Guo, K.Q. Allelopathic effects of several aquatic plants on algae. Acta Hydrobiol. Sin. 1992, 16, 1–7. [Google Scholar]
- Shannon-Firestone, S.; Firestone, J. Allelopathic potential of invasive species is determined by plant and soil community context. Plant Ecol. 2015, 216, 491–502. [Google Scholar] [CrossRef]
- Abrahim, D.; Braguini, W.L.; Kelmer-Bracht, A.M.; Ishii- Iwamoto, E.L. Effects of four monoterpenes on germination, primary root growth, and mitochondrial respiration of maize. J. Chem. Ecol. 2000, 26, 611–624. [Google Scholar] [CrossRef]
- Śliwińska-Wilczewska, S.; Budzałek, G.; Kowalska, Z.; Klin, M.; Latała, A. Baltic macroalgae as a potential source for commercial applications–review. Ann. Univ. Paedagog. Crac. Studia Nat. 2020, 5, 220–237. [Google Scholar]
- Lubchenco, J.; Gaines, S.D. A unified approach to marine plant-herbivore interactions: I. Populations and communities. Annu. Rev. Ecol. Evol. Syst. 1981, 12, 405–437. [Google Scholar] [CrossRef]
- Hay, M.E.; Fenical, W. Marine plant-herbivore interactions: The ecology of chemical defense. Annu. Rev. Ecol. Evol. Syst. 1988, 19, 111–145. [Google Scholar] [CrossRef]
- Reigosa, M.J.; Sánchez-Moreiras, A.; González, L. Ecophysiological approach in allelopathy. Crit. Rev. Plant Sci. 1999, 18, 577–608. [Google Scholar] [CrossRef]
- Chao, L.; Levin, B.R. Structured habitats and the evolution of anticompetitor toxins in bacteria. Proc. Natl. Acad. Sci. USA 1981, 78, 6324–6328. [Google Scholar] [CrossRef] [Green Version]
- Antonelli, P.L.; Sammarco, P.W.; Coll, J.C. A model of allelochemical interactions between soft and scleractinian corals on the Great Barrier Reef. J. Biol. Syst. 1993, 1, 1–17. [Google Scholar] [CrossRef]
- Cheng, H.H. Characterization of the mechanisms of allelopathy-modeling and experimental approaches. ACS Sym. Ser. 1995, 582, 132–141. [Google Scholar]
- Tapaswi, P.K.; Mukhopadhyay, A. Effects of environmental fluctuation on plankton allelopathy. J. Math. Biol. 1999, 39, 39–58. [Google Scholar] [CrossRef]
- An, M.; Johnson, I.R.; Lovett, J.V. Mathematical modelling of residue allelopathy the effects of intrinsic and extrinsic factors. Plant Soil 2002, 246, 11–22. [Google Scholar] [CrossRef]
- An, M.; Liu, D.L.; Johnson, I.R.; Lovett, J.V. Mathematical modelling of allelopathy. II. The dynamics of allelochemicals from living plants in the environment. Ecol. Model. 2003, 161, 53–66. [Google Scholar] [CrossRef]




| Donor Chlorophyta | Target Organism—Cnidaria | Effect | References |
| Bryopsis corymbose | Pocillopora damicornis | − | Lee et al. [28] |
| Bryopsis sp. | Pocillopora acuta | − | Fong et al. [39] |
| Chlorodesmis fastigiata | Acropora millepora | +/0 | Birrell et al. [40] |
| Chlorodesmis fastigiata | Acropora aspera | − | Bonaldo and Hay [23] |
| Pocillopora damicornis | − | ||
| Porites cylindrica | − | ||
| Porites lobata | − | ||
| Chlorodesmis fastigiata | Acropora intermedia | − | Del Monaco et al. [25] |
| Chlorodesmis fastigiata | Phialophora verrucosa | − | Longo and Hay [26] |
| Chlorodesmis fastigiata | Acropora millepora | − | Rasher et al. [22] |
| Montipora digitata | − | ||
| Pocillopora damicornis | − | ||
| Chlorodesmis fastigiata | Acropora millepora | − | Ritson-Williams et al. [24] |
| Montipora digitata | − | ||
| Pocillopora damicornis | − | ||
| Chlorodesmis fastigiata | Acropora cuneata | − | Tanner [21] |
| Acropora hrueggemanni | − | ||
| Acropora pnlifera | − | ||
| Pocillopora damicornis | − | ||
| Halimeda opuntia | Pocillopora damicornis | − | Lee et al. [28] |
| Halimeda tuna | Montastraea faveolate | − | Morrow et al. [52] |
| Porites astreoides | 0 | ||
| Halimeda sp. | Acropora cuneata | − | Tanner [21] |
| Acropora hrueggemanni | − | ||
| Acropora pnlifera | − | ||
| Pocillopora damicornis | − | ||
| Rhiphilia pencilloides | Porites rus | − | Andras et al. [51] |
| Donor Chlorophyta | Target Organism—Mollusca | Effect | References |
| Ulva australis | Haliotis rubra | + | Huggett et al. [49] |
| Ulva compressa | Crassostrea virginica | − | Green-Gavrielidis et al. [38] |
| Ulva compressa | Haliotis rubra | + | Huggett et al. [49] |
| Ulva fenestrata | Crassostrea gigas | − | Nelson et al. [47] |
| Ulva lactuca | Crassostrea virginica | − | Green-Gavrielidis et al. [38] |
| Ulvaria lactuca | Crassostrea gigas | − | Nelson and Gregg [48] |
| Ulva lens | Crassostrea gigas | − | Nelson et al. [47] |
| Ulvaria obscura | Haliotis rubra | + | Huggett et al. [49] |
| Ulva obscura | Crassostrea virginica | − | Green-Gavrielidis et al. [38] |
| Ulvaria obscura | Crassostrea gigas | − | Nelson and Gregg [48] |
| Ulva sp. | Haliotis rufescens | + | Muñoz et al. [50] |
| Donor Chlorophyta | Target Organism—Annelida | Effect | References |
| Chaetomorpha sp. | Sabellaria cementarium | − | Warkus et al. [41] |
| Spinoidae sp. | − | ||
| Codium fragile | Sabellaria cementarium | − | Warkus et al. [41] |
| Spinoidae sp. | − | ||
| Ulva (Enteromorpha) sp. | Sabellaria cementarium | − | Warkus et al. [41] |
| Spinoidae sp. | − | ||
| Ulva lactuca | Sabellaria cementarium | − | Warkus et al. [41] |
| Spinoidae sp. | − | ||
| Donor Chlorophyta | Target Organism—Arthropoda | Effect | References |
| Ulvaria obscura | Cancer oregonensis | 0/− | Van Alstyne et al. [46] |
| Metacarcinus magister | 0/− | ||
| Donor Chlorophyta | Target Organism—Chordata | Effect | References |
| Anadyomene stellata | Carassius auratus auratus | 0/− | Alvarez-Hernández et al. [8] |
| Caulerpa cupressoides | Carassius auratus auratus | 0/− | Alvarez-Hernández et al. [8] |
| Caulerpa paspaloides | Carassius auratus auratus | 0/− | Alvarez-Hernández et al. [8] |
| Caulerpa racemosa | Carassius auratus auratus | 0/− | Alvarez-Hernández et al. [8] |
| Chaetomorpha antennina | Carassius auratus auratus | 0/− | Alvarez-Hernández et al. [8] |
| Penicillus capitatus | Carassius auratus auratus | 0/− | Alvarez-Hernández et al. [8] |
| Donor Rhodophyta | Target Organism—Cnidaria | Effect | References |
| Amphiroa crassa | Acropora millepora | − | Rasher et al. [22], Ritson-Williams et al. [24] |
| Montipora digitata | 0 | ||
| Pocillopora damicornis | − | ||
| Amansia glomerata | Acropora intermedia | − | Del Monaco et al. [25] |
| Amansia rhodantha | Phialophora verrucosa | − | Longo and Hay [26] |
| Asparagopsis taxiformis | Phialophora verrucosa | − | Longo and Hay [26] |
| Callophycus densus | Porites rus | − | Andras et al. [51] |
| Chondrophycus poiteaui | Porites astreoides | 0 | Kuffner et al. [42] |
| Endosiphonia horrida | Pocillopora acuta | − | Fong et al. [39] |
| Galaxaura filamentosa | Acropora millepora | − | Rasher et al. [22], Ritson-Williams et al. [24] |
| Montipora digitata | − | ||
| Pocillopora damicornis | − | ||
| Galaxaura filamentosa | Acropora aspera | − | Bonaldo and Hay [23] |
| Pocillopora damicornis | − | ||
| Porites cylindrica | − | ||
| Porites lobata | − | ||
| Hypnea pannosa | Pocillopora acuta | 0 | Fong et al. [39] |
| Liagora sp. | Acropora millepora | − | Rasher et al. [22], Ritson-Williams et al. [24] |
| Montipora digitata | 0 | ||
| Pocillopora damicornis | − | ||
| Phacelocarpus neurymenioides | Porites rus | − | Andras et al. [51] |
| Plocamium pacificum | Porites rus | − | Andras et al. [51] |
| Peyssonnelia sp. | Acropora cuneata | − | Tanner [21] |
| Acropora hrueggemanni | − | ||
| Acropora pnlifera | − | ||
| Pocillopora damicornis | 0/− | ||
| Donor Rhodophyta | Target Organism—Annelida | Effect | References |
| Grateloupia turu turu | Sabellaria cementarium | − | Warkus et al. [41] |
| Spinoidae sp. | − | ||
| Polysiphonia denudata | Sabellaria cementarium | − | Warkus et al. [41] |
| Spinoidae sp. | − | ||
| Donor Rhodophyta | Target Organism—Echinodermata | Effect | References |
| Delisea pulchra | Holopneustes purpurascens | + | Williamson et al. [33] |
| Tichocarpus crinitus | Strongylocentrotus intermedius | − | Ishii et al. [36] |
| Donor Rhodophyta | Target Organism—Chordata | Effect | References |
| Acanthophora spicifera | Carassius auratus auratus | 0/− | Alvarez-Hernández et al. [8] |
| Amphiroa beauvoisii | Carassius auratus auratus | 0/− | Alvarez-Hernández et al. [8] |
| Centroceras clavulatum | Carassius auratus auratus | 0/− | Alvarez-Hernández et al. [8] |
| Ceramium nitens | Carassius auratus auratus | 0/− | Alvarez-Hernández et al. [8] |
| Chondria littoralis | Carassius auratus auratus | 0 | Alvarez-Hernández et al. [8] |
| Chondriopsis dasyphylla f. pyrifera | Carassius auratus auratus | − | Alvarez-Hernández et al. [8] |
| Crassiphycus caudatus (Gracilaria caudata) | Carassius auratus auratus | 0 | Alvarez-Hernández et al. [8] |
| Dermonema virens | Carassius auratus auratus | 0 | Alvarez-Hernández et al. [8] |
| Digenea simplex | Carassius auratus auratus | 0 | Alvarez-Hernández et al. [8] |
| Gracilaria cervicornis | Carassius auratus auratus | 0/− | Alvarez-Hernández et al. [8] |
| Gracilaria tikvahiae | Carassius auratus auratus | 0/− | Alvarez-Hernández et al. [8] |
| Grateloupia filicina | Carassius auratus auratus | 0 | Alvarez-Hernández et al. [8] |
| Hypnea musciformis | Carassius auratus auratus | 0/− | Alvarez-Hernández et al. [8] |
| Hypnea spinella | Carassius auratus auratus | 0/− | Alvarez-Hernández et al. [8] |
| Laurencia obtusa | Carassius auratus auratus | 0/− | Alvarez-Hernández et al. [8] |
| Liagora ceranoides | Carassius auratus auratus | 0/− | Alvarez-Hernández et al. [8] |
| Tayloriella dictyurus | Carassius auratus auratus | 0 | Alvarez-Hernández et al. [8] |
| Yuzurua poiteaui var. gemmifera | Carassius auratus auratus | 0 | Alvarez-Hernández et al. [8] |
| Donor Ochrophyta | Target Organism—Cnidaria | Effect | References |
| Dictyota bartayresiana | Phialophora verrucosa | − | Longo and Hay [26] |
| Dictyota bartayresiana | Acropora millepora | − | Rasher et al. [22], Ritson-Williams et al. [24] |
| Montipora digitata | − | ||
| Pocillopora damicornis | − | ||
| Dictyota bartayresiana | Acropora cervicornis | 0 | Ritson-Williams et al. [27] |
| Acropora palmata | − | ||
| Pseudodiploria strigosa | 0 | ||
| Dictyota cervicornis | Acropora intermedia | − | Del Monaco et al. [25] |
| Dictyota menstrualis | Porites astreoides | − | Olsen et al. [56] |
| Dictyota pinnatifida | Porites astreoides | − | Paul et al. [55] |
| Dictyota pulchella | Porites astreoides | − | Paul et al. [55] |
| Dictyota pulchella | Acropora cervicornis | 0 | Ritson-Williams et al. [27] |
| Acropora palmata | − | ||
| Pseudodiploria strigosa | 0 | ||
| Dictyota sp. | Montastraea faveolate | 0/− | Morrow et al. [52] |
| Porites astreoides | 0/− | ||
| Dictyota sp. | Briareum asbestinum | − | Kuffner et al. [42] |
| Porites astreoides | − | ||
| Lobophora abscondita | Acropora muricate | − | Vieira et al. [57] |
| Montipora hirsute | 0 | ||
| Porites cylindrica | 0 | ||
| Stylophora pistillata | − | ||
| Lobophora crassa | Acropora muricate | − | Vieira et al. [57] |
| Montipora hirsute | 0 | ||
| Porites cylindrica | 0 | ||
| Stylophora pistillata | − | ||
| Lobophora dimorpha | Acropora muricate | − | Vieira et al. [57] |
| Montipora hirsute | 0 | ||
| Porites cylindrica | 0 | ||
| Stylophora pistillata | − | ||
| Lobophora hederacea | Acropora muricate | − | Vieira et al. [57] |
| Montipora hirsute | 0 | ||
| Porites cylindrica | 0 | ||
| Stylophora pistillata | − | ||
| Lobophora monticola | Acropora muricate | − | Vieira et al. [57] |
| Montipora hirsute | 0 | ||
| Porites cylindrica | 0 | ||
| Stylophora pistillata | − | ||
| Lobophora nigrescens | Acropora muricate | − | Vieira et al. [57] |
| Montipora hirsute | 0 | ||
| Porites cylindrica | 0 | ||
| Stylophora pistillata | − | ||
| Lobophora rosacea | Acropora muricate | − | Vieira et al. [57] |
| Montipora hirsute | 0 | ||
| Porites cylindrica | 0 | ||
| Stylophora pistillata | − | ||
| Lobophora undulata | Acropora muricate | − | Vieira et al. [57] |
| Montipora hirsute | 0 | ||
| Porites cylindrica | 0 | ||
| Stylophora pistillata | − | ||
| Lobophora variegata | Acropora millepora | + | Birrell et al. [40] |
| Lobophora variegata | Briareum asbestinum | − | Kuffner et al. [42] |
| Porites astreoides | − | ||
| Lobophora variegata | Montastraea faveolate | − | Morrow et al. [52] |
| Porites astreoides | − | ||
| Lobophora sp. | Pocillopora acuta | − | Fong et al. [39] |
| Lobophora sp. | Acropora cervicornis | − | Ritson-Williams et al. [27] |
| Acropora palmata | − | ||
| Pseudodiploria strigosa | 0 | ||
| Padina boryana | Acropora millepora | − | Rasher et al. [22], Ritson-Williams et al. [24] |
| Montipora digitata | 0 | ||
| Pocillopora damicornis | − | ||
| Padina minor | Pocillopora damicornis | 0 | Lee et al. [28] |
| Padina sp. | Acropora millepora | − | Birrell et al. [40] |
| Sargassum polycystum | Acropora millepora | − | Ritson-Williams et al. [24] |
| Montipora digitata | 0 | ||
| Pocillopora damicornis | − | ||
| Sargassum sp. | Pocillopora damicornis | − | Lee et al. [28] |
| Sphacelaria sp. | Acropora millepora | − | Webster et al. [29] |
| Turbinaria conoides | Acropora millepora | 0 | Rasher et al. [22] |
| Montipora digitata | 0 | ||
| Pocillopora damicornis | 0 | ||
| Turbinaria conoides | Acropora millepora | − | Ritson-Williams et al. [24] |
| Montipora digitata | 0 | ||
| Pocillopora damicornis | − | ||
| Turbinaria ornata | Phialophora verrucosa | 0 | Longo and Hay [26] |
| Turbinaria ornata | Acropora cuneata | − | Tanner [21] |
| Acropora hrueggemanni | − | ||
| Acropora pnlifera | − | ||
| Pocillopora damicornis | − | ||
| Donor Ochrophyta | Target Organism—Mollusca | Effect | References |
| Dilophus okamurae | Haliotis discus hannai | − | Suzuki et al. [34] |
| Donor Ochrophyta | Target Organism—Annelida | Effect | References |
| Desmarestia viridis | Sabellaria cementarium | − | Warkus et al. [41] |
| Spinoidae sp. | − | ||
| Laminaria sp. | Sabellaria cementarium | − | Warkus et al. [41] |
| Spinoidae sp. | − | ||
| Donor Ochrophyta | Target Organism—Echinodermata | Effect | References |
| Dictyota pfaffi | Lytechinus variegates | − | Barbosa et al. [35] |
| Ecklonia radiata | Holopneustes purpurascens | 0 | Williamson et al. [33] |
| Stypopodium zonale | Strongylocentrotus purpuratus | − | Gerwick and Fenical [31] |
| Donor Ochrophyta | Target Organism—Arthropoda | Effect | References |
| Dictyota pfaffii | Pachygrapsus transversus | 0 | Barbosa et al. [35] |
| Donor Ochrophyta | Target Organism—Chordata | Effect | References |
| Dictyopteris delicatula | Carassius auratus auratus | 0/− | Alvarez-Hernández et al. [8] |
| Dictyota bartayresiana | Carassius auratus auratus | 0/− | Alvarez-Hernández et al. [8] |
| Dictyota implexa | Carassius auratus auratus | 0/− | Alvarez-Hernández et al. [8] |
| Dictyota spinulosa | Tilapia mossambica | − | Tanaka and Higa [32] |
| Lobophora variegata | Carassius auratus auratus | 0/− | Alvarez-Hernández et al. [8] |
| Padina gymnospora | Carassius auratus auratus | 0/− | Alvarez-Hernández et al. [8] |
| Sargassum liebmannii | Carassius auratus auratus | 0/− | Alvarez-Hernández et al. [8] |
| Stypopodium zonale | Carassius auratus auratus | 0/− | Alvarez-Hernández et al. [8] |
| Stypopodium zonale | Eupomacentrus leucostictus | − | Gerwick and Fenical [31] |
| Stypopodium zonale | Eupomacentrus leucostictus | − | Gerwick et al. [30] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Budzałek, G.; Śliwińska-Wilczewska, S.; Wiśniewska, K.; Wochna, A.; Bubak, I.; Latała, A.; Wiktor, J.M. Macroalgal Defense against Competitors and Herbivores. Int. J. Mol. Sci. 2021, 22, 7865. https://doi.org/10.3390/ijms22157865
Budzałek G, Śliwińska-Wilczewska S, Wiśniewska K, Wochna A, Bubak I, Latała A, Wiktor JM. Macroalgal Defense against Competitors and Herbivores. International Journal of Molecular Sciences. 2021; 22(15):7865. https://doi.org/10.3390/ijms22157865
Chicago/Turabian StyleBudzałek, Gracjana, Sylwia Śliwińska-Wilczewska, Kinga Wiśniewska, Agnieszka Wochna, Iwona Bubak, Adam Latała, and Józef Maria Wiktor. 2021. "Macroalgal Defense against Competitors and Herbivores" International Journal of Molecular Sciences 22, no. 15: 7865. https://doi.org/10.3390/ijms22157865
APA StyleBudzałek, G., Śliwińska-Wilczewska, S., Wiśniewska, K., Wochna, A., Bubak, I., Latała, A., & Wiktor, J. M. (2021). Macroalgal Defense against Competitors and Herbivores. International Journal of Molecular Sciences, 22(15), 7865. https://doi.org/10.3390/ijms22157865