
The Presynaptic Scaffold Protein Bassoon in Forebrain Excitatory Neurons Mediates Hippocampal Circuit Maturation: Potential Involvement of TrkB Signalling

 , and
, and 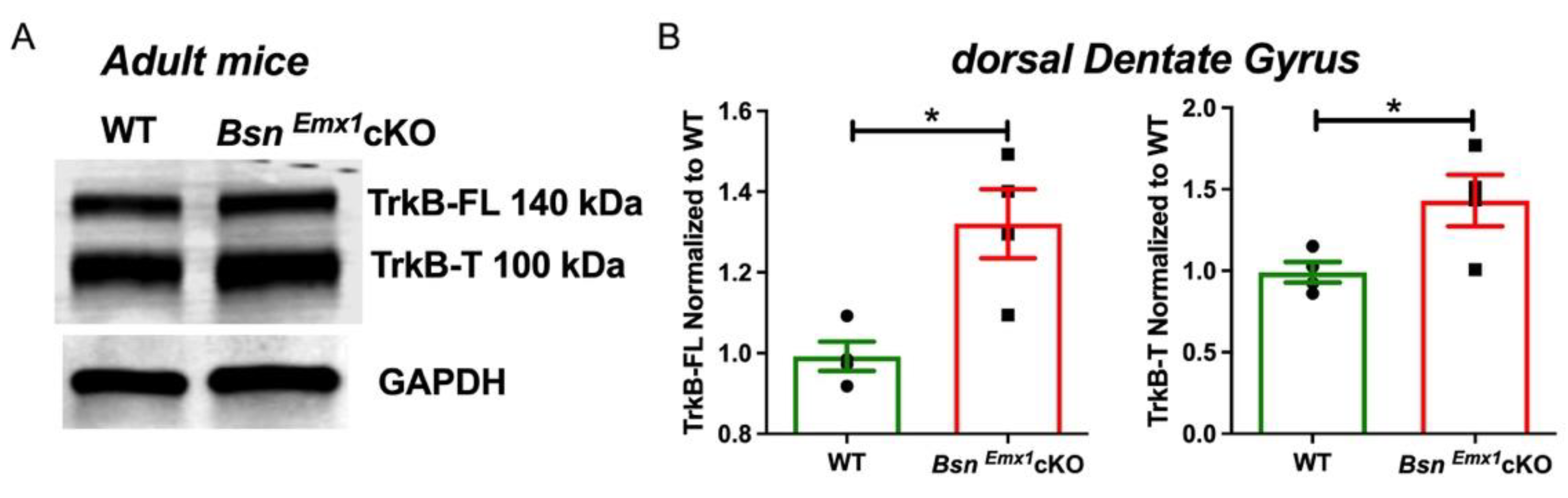{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
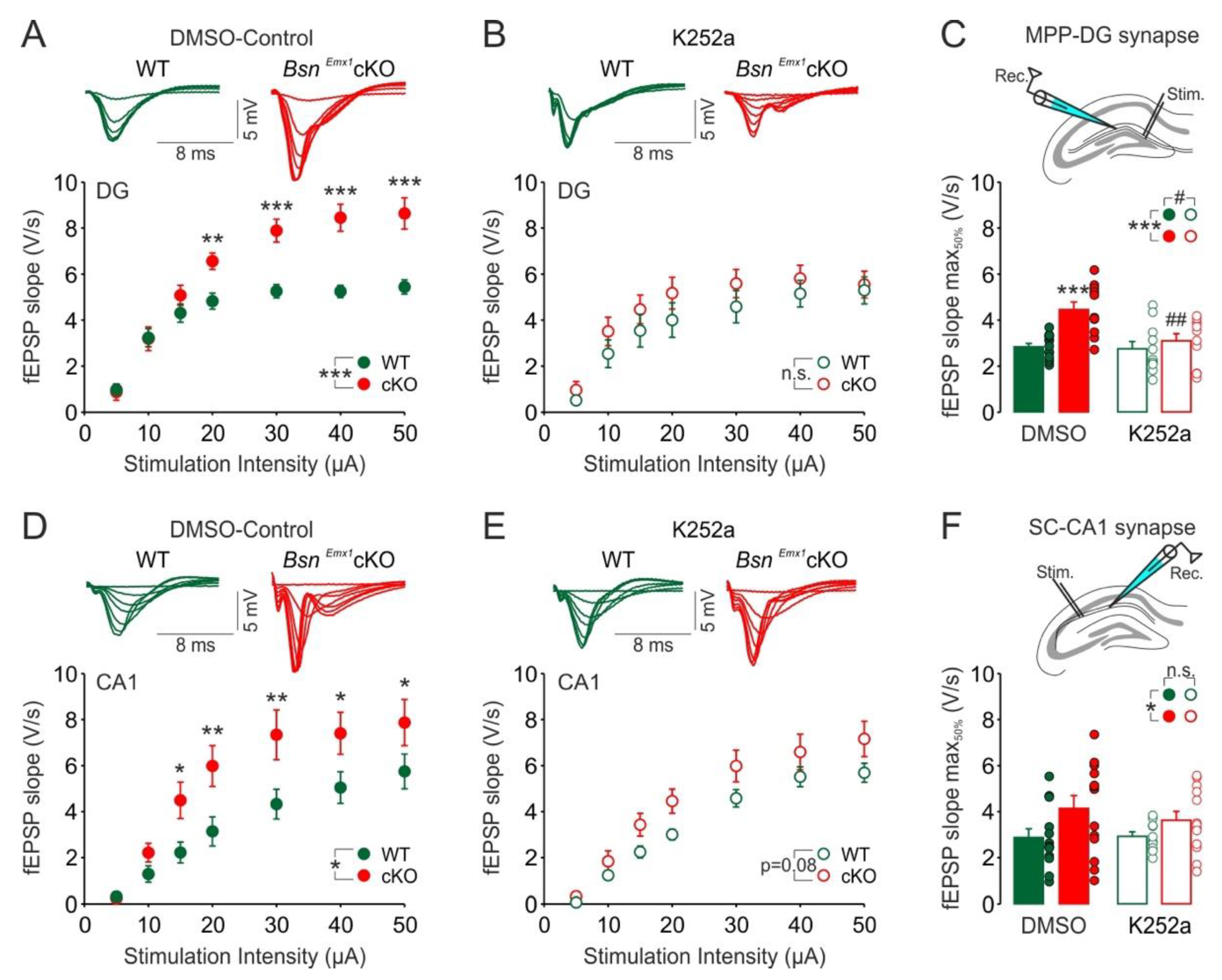
2.1. Increased MPP-DG Synapse Excitability Is Associated with Elevated TrkB Expression and Can Be Normalized by Acute TrkB Blockade in BsnEmx1cKO Mice at Adult Age
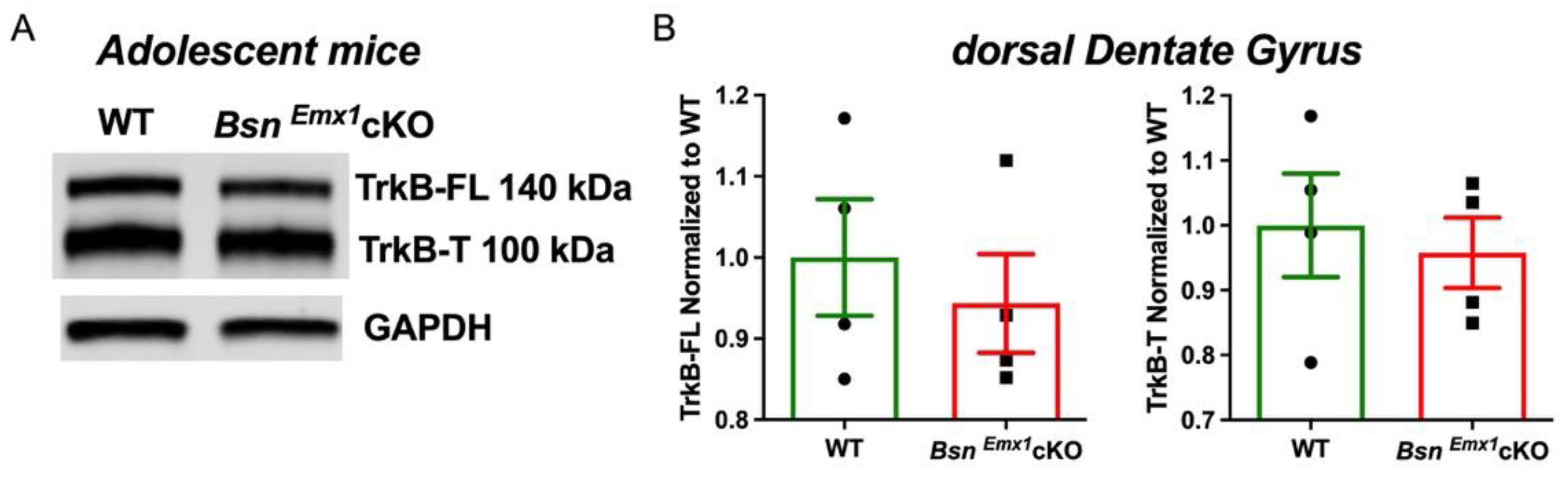
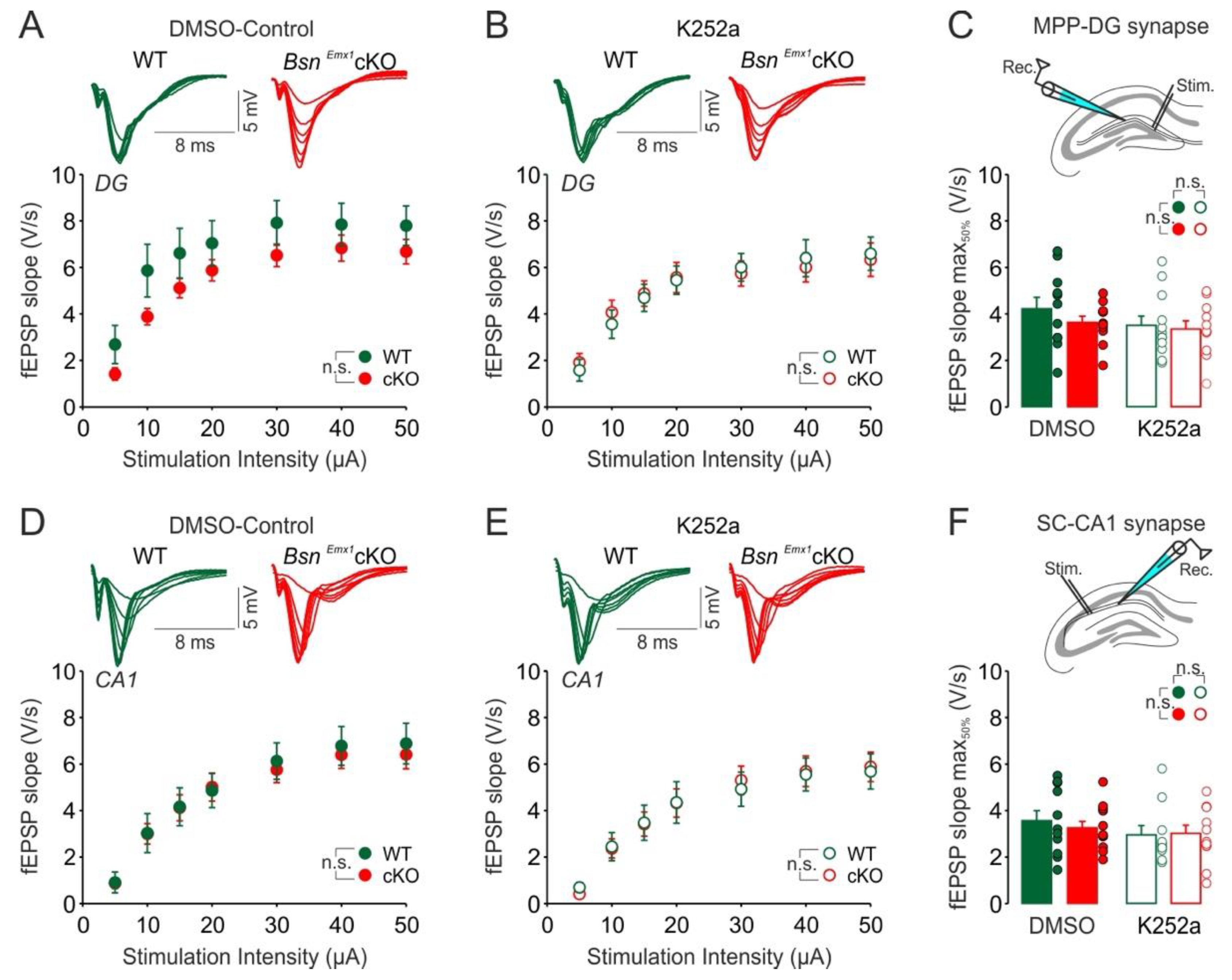
2.2. Enhanced Hippocampal Excitability and Its Modulation by TrkB-Mediated Signalling in the BsnEmx1cKO Mice Are Not Evident during Adolescence
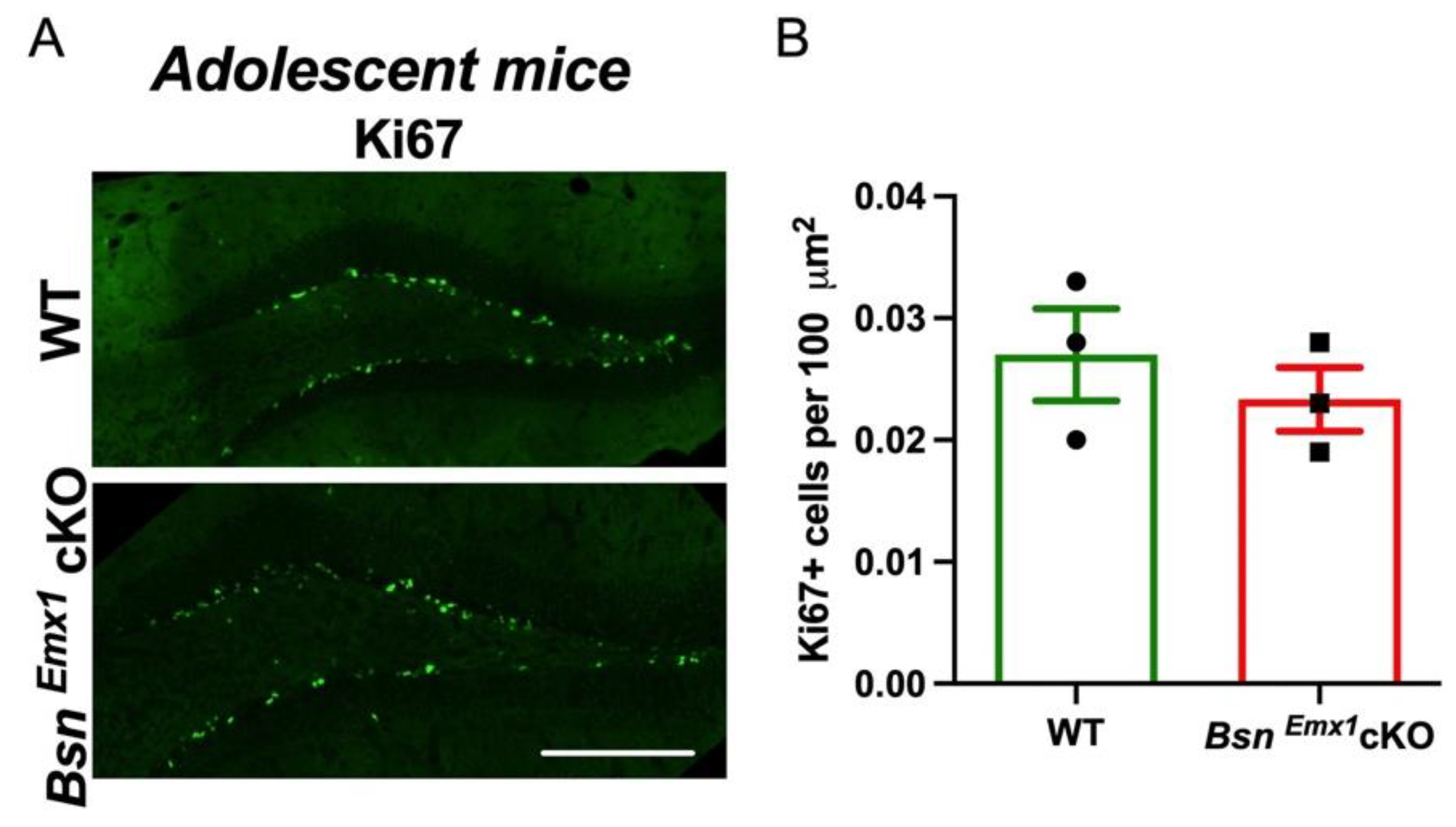
2.3. Unaltered Neurogenesis in Adolescent BsnEmx1cKO Mice
3. Discussion
4. Materials and Methods
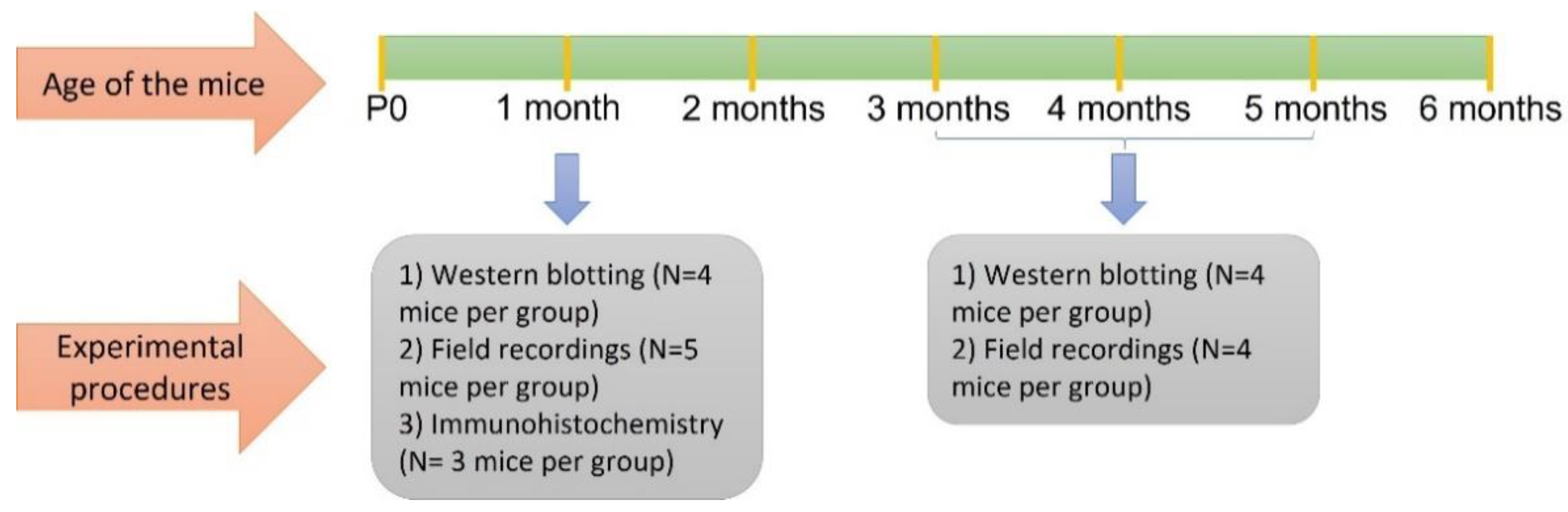
4.1. Animals
4.2. Immunohistochemistry and Microscopy
4.3. Tissue Preparation for Western Blot Analyses
4.4. Slice Preparation for Electrophysiology
4.5. Extracellular Field Potential Recordings
4.6. Data Analysis and Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tang, G.; Gudsnuk, K.; Kuo, S.-H.; Cotrina, M.L.; Rosoklija, G.; Sosunov, A.; Sonders, M.S.; Kanter, E.; Castagna, C.; Yamamoto, A.; et al. Loss of mTOR-Dependent Macroautophagy Causes Autistic-like Synaptic Pruning Deficits. Neuron 2014, 83, 1131–1143. [Google Scholar] [CrossRef] [Green Version]
- Hutsler, J.J.; Zhang, H. Increased dendritic spine densities on cortical projection neurons in autism spectrum disorders. Brain Res. 2010, 1309, 83–94. [Google Scholar] [CrossRef]
- Van Spronsen, M.; Hoogenraad, C.C. Synapse Pathology in Psychiatric and Neurologic Disease. Curr. Neurol. Neurosci. Rep. 2010, 10, 207–214. [Google Scholar] [CrossRef] [Green Version]
- Albrecht, A.; Müller, I.; Ardi, Z.; Çalışkan, G.; Gruber, D.; Ivens, S.; Segal, M.; Behr, J.; Heinemann, U.; Stork, O.; et al. Neurobiological consequences of juvenile stress: A GABAergic perspective on risk and resilience. Neurosci. Biobehav. Rev. 2017, 74 Pt A, 21–43. [Google Scholar] [CrossRef]
- Fareri, D.S.; Tottenham, N. Effects of early life stress on amygdala and striatal development. Dev. Cogn. Neurosci. 2016, 19, 233–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolb, B.; Harker, A.; Mychasiuk, R.; de Melo, S.R.; Gibb, R. Stress and prefrontal cortical plasticity in the developing brain. Cogn. Dev. 2017, 42, 15–26. [Google Scholar] [CrossRef]
- Lanore, F.; Blanchet, C.; Fejtova, A.; Pinheiro, P.; Richter, K.; Balschun, D.; Gundelfinger, E.; Mulle, C. Impaired development of hippocampal mossy fibre synapses in mouse mutants for the presynaptic scaffold protein Bassoon. J. Physiol. 2010, 588 Pt 12, 2133–2145. [Google Scholar] [CrossRef]
- Annamneedi, A.; Caliskan, G.; Muller, S.; Montag, D.; Budinger, E.; Angenstein, F.; Fejtova, A.; Tischmeyer, W.; Gundelfinger, E.D.; Stork, O.; et al. Ablation of the presynaptic organizer Bassoon in excitatory neurons retards dentate gyrus maturation and enhances learning performance. Brain Struct. Funct. 2018, 223, 3423–3445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sudhof, T.C. The presynaptic active zone. Neuron 2012, 75, 11–25. [Google Scholar] [CrossRef] [Green Version]
- Gundelfinger, E.D.; Fejtova, A. Molecular organization and plasticity of the cytomatrix at the active zone. Curr. Opin. Neurobiol. 2012, 22, 423–430. [Google Scholar] [CrossRef] [PubMed]
- Gundelfinger, E.D.; Reissner, C.; Garner, C.C. Role of Bassoon and Piccolo in Assembly and Molecular Organization of the Active Zone. Front. Synaptic Neurosci. 2016, 7, 19. [Google Scholar] [CrossRef] [Green Version]
- Okerlund, N.D.; Schneider, K.; Leal-Ortiz, S.; Montenegro-Venegas, C.; Kim, S.A.; Garner, L.C.; Waites, C.L.; Gundelfinger, E.D.; Reimer, R.J.; Garner, C.C.; et al. Bassoon Controls Presynaptic Autophagy through Atg5. Neuron 2017, 93, 897–913.e7. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann-Conaway, S.; Brockmann, M.M.; Schneider, K.; Annamneedi, A.; Rahman, K.A.; Bruns, C.; Textoris-Taube, K.; Trimbuch, T.; Smalla, K.H.; Rosenmund, C.; et al. Parkin contributes to synaptic vesicle autophagy in Bassoon-deficient mice. eLife 2020, 9, e56590. [Google Scholar] [CrossRef]
- Montenegro-Venegas, C.; Fienko, S.; Anni, D.; Pina-Fernández, E.; Frischknecht, R.; Fejtova, A. Bassoon inhibits proteasome activity via interaction with PSMB4. Cell. Mol. Life Sci. 2021, 78, 1545–1563. [Google Scholar] [CrossRef]
- Dick, O.; tom Dieck, S.; Altrock, W.D.; Ammermuller, J.; Weiler, R.; Garner, C.C.; Gundelfinger, E.D.; Brandstatter, J.H. The presynaptic active zone protein bassoon is essential for photoreceptor ribbon synapse formation in the retina. Neuron 2003, 37, 775–786. [Google Scholar] [CrossRef] [Green Version]
- tom Dieck, S.; Altrock, W.D.; Kessels, M.M.; Qualmann, B.; Regus, H.; Brauner, D.; Fejtová, A.; Bracko, O.; Gundelfinger, E.D.; Brandstätter, J.H.; et al. Molecular dissection of the photoreceptor ribbon synapse: Physical interaction of Bassoon and RIBEYE is essential for the assembly of the ribbon complex. J. Cell Biol. 2005, 168, 825–836. [Google Scholar] [CrossRef] [Green Version]
- Khimich, D.; Nouvian, R.; Pujol, R.; Tom Dieck, S.; Egner, A.; Gundelfinger, E.D.; Moser, T. Hair cell synaptic ribbons are essential for synchronous auditory signalling. Nature 2005, 434, 889–894. [Google Scholar] [CrossRef]
- Frank, T.; Rutherford, M.A.; Strenzke, N.; Neef, A.; Pangrsic, T.; Khimich, D.; Fejtova, A.; Gundelfinger, E.D.; Liberman, M.C.; Harke, B.; et al. Bassoon and the synaptic ribbon organize Ca2+ channels and vesicles to add release sites and promote refilling. Neuron 2010, 68, 724–738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altrock, W.D.; tom Dieck, S.; Sokolov, M.; Meyer, A.C.; Sigler, A.; Brakebusch, C.; Fassler, R.; Richter, K.; Boeckers, T.M.; Potschka, H.; et al. Functional inactivation of a fraction of excitatory synapses in mice deficient for the active zone protein bassoon. Neuron 2003, 37, 787–800. [Google Scholar] [CrossRef] [Green Version]
- Sgobio, C.; Ghiglieri, V.; Costa, C.; Bagetta, V.; Siliquini, S.; Barone, I.; Di Filippo, M.; Gardoni, F.; Gundelfinger, E.D.; Di Luca, M.; et al. Hippocampal synaptic plasticity, memory, and epilepsy: Effects of long-term valproic acid treatment. Biol. Psychiatry 2010, 67, 567–574. [Google Scholar] [CrossRef] [PubMed]
- Andreska, T.; Aufmkolk, S.; Sauer, M.; Blum, R. High abundance of BDNF within glutamatergic presynapses of cultured hippocampal neurons. Front. Cell. Neurosci. 2014, 8, 107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, P.Z.; Nusslock, R. Exercise-Mediated Neurogenesis in the Hippocampus via BDNF. Front. Neurosci. 2018, 12, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choo, M.; Miyazaki, T.; Yamazaki, M.; Kawamura, M.; Nakazawa, T.; Zhang, J.; Tanimura, A.; Uesaka, N.; Watanabe, M.; Sakimura, K.; et al. Retrograde BDNF to TrkB signaling promotes synapse elimination in the developing cerebellum. Nat. Commun. 2017, 8, 195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bosman, L.W.J.; Hartmann, J.; Barski, J.J.; Lepier, A.; Noll-Hussong, M.; Reichardt, L.F.; Konnerth, A. Requirement of TrkB for synapse elimination in developing cerebellar Purkinje cells. Brain Cell Biol. 2006, 35, 87–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez, A.; Alcantara, S.; Borrell, V.; Del Rio, J.A.; Blasi, J.; Otal, R.; Campos, N.; Boronat, A.; Barbacid, M.; Silos-Santiago, I.; et al. TrkB and TrkC signaling are required for maturation and synaptogenesis of hippocampal connections. J. Neurosci. 1998, 18, 7336–7350. [Google Scholar] [CrossRef] [Green Version]
- Louhivuori, V.; Vicario, A.; Uutela, M.; Rantamäki, T.; Louhivuori, L.M.; Castrén, E.; Tongiorgi, E.; Åkerman, K.E.; Castrén, M.L. BDNF and TrkB in neuronal differentiation of Fmr1-knockout mouse. Neurobiol. Dis. 2011, 41, 469–480. [Google Scholar] [CrossRef]
- Castrén, M.L.; Castrén, E. BDNF in fragile X syndrome. Neuropharmacology 2014, 76, 729–736. [Google Scholar] [CrossRef]
- Ray, M.T.; Shannon Weickert, C.; Webster, M.J. Decreased BDNF and TrkB mRNA expression in multiple cortical areas of patients with schizophrenia and mood disorders. Transl. Psychiatry 2014, 4, e389. [Google Scholar] [CrossRef]
- Wong, J.; Rothmond, D.A.; Webster, M.J.; Shannon Weickert, C. Increases in Two Truncated TrkB Isoforms in the Prefrontal Cortex of People with Schizophrenia. Schizophr. Bull. 2011, 39, 130–140. [Google Scholar] [CrossRef] [Green Version]
- Correia, C.T.; Coutinho, A.M.; Sequeira, A.F.; Sousa, I.G.; Lourenço Venda, L.; Almeida, J.P.; Abreu, R.L.; Lobo, C.; Miguel, T.S.; Conroy, J.; et al. Increased BDNF levels and NTRK2 gene association suggest a disruption of BDNF/TrkB signaling in autism. Genes Brain Behav. 2010, 9, 841–848. [Google Scholar] [CrossRef] [Green Version]
- Fahnestock, M.; Nicolini, C. Bridging the Gap between Genes and Behavior: Brain-Derived NeurotrophicFactor and the mTOR Pathway in Idiopathic Autism. Autism Open Access 2015, 5, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Heyden, A.; Ionescu, M.C.; Romorini, S.; Kracht, B.; Ghiglieri, V.; Calabresi, P.; Seidenbecher, C.; Angenstein, F.; Gundelfinger, E.D. Hippocampal enlargement in Bassoon-mutant mice is associated with enhanced neurogenesis, reduced apoptosis, and abnormal BDNF levels. Cell Tissue Res. 2011, 346, 11–26. [Google Scholar] [CrossRef] [PubMed]
- Dieni, S.; Matsumoto, T.; Dekkers, M.; Rauskolb, S.; Ionescu, M.S.; Deogracias, R.; Gundelfinger, E.D.; Kojima, M.; Nestel, S.; Frotscher, M.; et al. BDNF and its pro-peptide are stored in presynaptic dense core vesicles in brain neurons. J. Cell Biol. 2012, 196, 775–788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angenstein, F.; Niessen, H.G.; Goldschmidt, J.; Lison, H.; Altrock, W.D.; Gundelfinger, E.D.; Scheich, H. Manganese-enhanced MRI reveals structural and functional changes in the cortex of Bassoon mutant mice. Cereb Cortex 2007, 17, 28–36. [Google Scholar] [CrossRef] [Green Version]
- Angenstein, F.; Hilfert, L.; Zuschratter, W.; Altrock, W.D.; Niessen, H.G.; Gundelfinger, E.D. Morphological and metabolic changes in the cortex of mice lacking the functional presynaptic active zone protein bassoon: A combined 1H-NMR spectroscopy and histochemical study. Cereb Cortex 2008, 18, 890–897. [Google Scholar] [CrossRef] [Green Version]
- Ghiglieri, V.; Sgobio, C.; Patassini, S.; Bagetta, V.; Fejtova, A.; Giampa, C.; Marinucci, S.; Heyden, A.; Gundelfinger, E.D.; Fusco, F.R.; et al. TrkB/BDNF-dependent striatal plasticity and behavior in a genetic model of epilepsy: Modulation by valproic acid. Neuropsychopharmacology 2010, 35, 1531–1540. [Google Scholar] [CrossRef]
- Kowianski, P.; Lietzau, G.; Czuba, E.; Waskow, M.; Steliga, A.; Morys, J. BDNF: A Key Factor with Multipotent Impact on Brain Signaling and Synaptic Plasticity. Cell. Mol. Neurobiol. 2018, 38, 579–593. [Google Scholar] [CrossRef]
- Huang, E.J.; Reichardt, L.F. Neurotrophins: Roles in neuronal development and function. Annu. Rev. Neurosci. 2001, 24, 677–736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murray, P.S.; Holmes, P.V. An Overview of Brain-Derived Neurotrophic Factor and Implications for Excitotoxic Vulnerability in the Hippocampus. Int. J. Pept. 2011, 2011, 654085. [Google Scholar] [CrossRef] [Green Version]
- Tervonen, T.A.; Ajamian, F.; De Wit, J.; Verhaagen, J.; Castrén, E.; Castrén, M. Overexpression of a truncated TrkB isoform increases the proliferation of neural progenitors. Eur. J. Neurosci. 2006, 24, 1277–1285. [Google Scholar] [CrossRef] [PubMed]
- Minichiello, L.; Korte, M.; Wolfer, D.; Kuhn, R.; Unsicker, K.; Cestari, V.; Rossi-Arnaud, C.; Lipp, H.P.; Bonhoeffer, T.; Klein, R. Essential role for TrkB receptors in hippocampus-mediated learning. Neuron 1999, 24, 401–414. [Google Scholar] [CrossRef] [Green Version]
- Bekinschtein, P.; Kent, B.A.; Oomen, C.A.; Clemenson, G.D.; Gage, F.H.; Saksida, L.M.; Bussey, T.J. BDNF in the dentate gyrus is required for consolidation of “pattern-separated” memories. Cell Rep. 2013, 5, 759–768. [Google Scholar] [CrossRef] [Green Version]
- Bekinschtein, P.; Kent, B.A.; Oomen, C.A.; Clemenson, G.D.; Gage, F.H.; Saksida, L.M.; Bussey, T.J. Brain-derived neurotrophic factor interacts with adult-born immature cells in the dentate gyrus during consolidation of overlapping memories. Hippocampus 2014, 24, 905–911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, I.Y.; Lyons, W.E.; Mamounas, L.A.; Thompson, R.F. Brain-derived neurotrophic factor plays a critical role in contextual fear conditioning. J. Neurosci. 2004, 24, 7958–7963. [Google Scholar] [CrossRef] [PubMed]
- Koponen, E.; Võikar, V.; Riekki, R.; Saarelainen, T.; Rauramaa, T.; Rauvala, H.; Taira, T.; Castrén, E. Transgenic mice overexpressing the full-length neurotrophin receptor trkB exhibit increased activation of the trkB-PLCgamma pathway, reduced anxiety, and facilitated learning. Mol. Cell. Neurosci. 2004, 26, 166–181. [Google Scholar] [CrossRef]
- Tolwani, R.J.; Buckmaster, P.S.; Varma, S.; Cosgaya, J.M.; Wu, Y.; Suri, C.; Shooter, E.M. BDNF overexpression increases dendrite complexity in hippocampal dentate gyrus. Neuroscience 2002, 114, 795–805. [Google Scholar] [CrossRef]
- Xu, B.; Zang, K.; Ruff, N.L.; Zhang, Y.A.; McConnell, S.K.; Stryker, M.P.; Reichardt, L.F. Cortical degeneration in the absence of neurotrophin signaling: Dendritic retraction and neuronal loss after removal of the receptor TrkB. Neuron 2000, 26, 233–245. [Google Scholar] [CrossRef] [Green Version]
- Donovan, M.H.; Yamaguchi, M.; Eisch, A.J. Dynamic expression of TrkB receptor protein on proliferating and maturing cells in the adult mouse dentate gyrus. Hippocampus 2008, 18, 435–439. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Luikart, B.W.; Birnbaum, S.; Chen, J.; Kwon, C.H.; Kernie, S.G.; Bassel-Duby, R.; Parada, L.F. TrkB regulates hippocampal neurogenesis and governs sensitivity to antidepressive treatment. Neuron 2008, 59, 399–412. [Google Scholar] [CrossRef] [Green Version]
- Binder, D.K.; Croll, S.D.; Gall, C.M.; Scharfman, H.E. BDNF and epilepsy: Too much of a good thing? Trends Neurosci. 2001, 24, 47–53. [Google Scholar] [CrossRef]
- Guyon, N.; Zacharias, L.R.; van Lunteren, J.A.; Immenschuh, J.; Fuzik, J.; Märtin, A.; Xuan, Y.; Zilberter, M.; Kim, H.; Meletis, K.; et al. Adult trkB Signaling in Parvalbumin Interneurons is Essential to Prefrontal Network Dynamics. J. Neurosci. 2021, 41, 3120–3141. [Google Scholar] [CrossRef]
- Trinchero, M.F.; Giacomini, D.; Schinder, A.F. Dynamic interplay between GABAergic networks and developing neurons in the adult hippocampus. Curr. Opin. Neurobiol. 2021, 69, 124–130. [Google Scholar] [CrossRef]
- Schattling, B.; Engler, J.B.; Volkmann, C.; Rothammer, N.; Woo, M.S.; Petersen, M.; Winkler, I.; Kaufmann, M.; Rosenkranz, S.C.; Fejtova, A.; et al. Bassoon proteinopathy drives neurodegeneration in multiple sclerosis. Nat. Neurosci. 2019, 22, 887–896. [Google Scholar] [CrossRef]
- Hagihara, H.; Toyama, K.; Yamasaki, N.; Miyakawa, T. Dissection of hippocampal dentate gyrus from adult mouse. J. Vis. Exp. 2009, 33, 1543. [Google Scholar] [CrossRef] [Green Version]
- Albrecht, A.; Ivens, S.; Papageorgiou, I.E.; Çalışkan, G.; Saiepour, N.; Brück, W.; Richter-Levin, G.; Heinemann, U.; Stork, O. Shifts in excitatory/inhibitory balance by juvenile stress: A role for neuron-astrocyte interaction in the dentate gyrus. Glia 2016, 64, 911–922. [Google Scholar] [CrossRef]
- Petersen, R.P.; Moradpour, F.; Eadie, B.D.; Shin, J.D.; Kannangara, T.S.; Delaney, K.R.; Christie, B.R. Electrophysiological identification of medial and lateral perforant path inputs to the dentate gyrus. Neuroscience 2013, 252, 154–168. [Google Scholar] [CrossRef]
- Dahl, D.; Sarvey, J.M. Norepinephrine induces pathway-specific long-lasting potentiation and depression in the hippocampal dentate gyrus. Proc. Natl. Acad. Sci. USA 1989, 86, 4776–4780. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Annamneedi, A.; del Angel, M.; Gundelfinger, E.D.; Stork, O.; Çalışkan, G. The Presynaptic Scaffold Protein Bassoon in Forebrain Excitatory Neurons Mediates Hippocampal Circuit Maturation: Potential Involvement of TrkB Signalling. Int. J. Mol. Sci. 2021, 22, 7944. https://doi.org/10.3390/ijms22157944
Annamneedi A, del Angel M, Gundelfinger ED, Stork O, Çalışkan G. The Presynaptic Scaffold Protein Bassoon in Forebrain Excitatory Neurons Mediates Hippocampal Circuit Maturation: Potential Involvement of TrkB Signalling. International Journal of Molecular Sciences. 2021; 22(15):7944. https://doi.org/10.3390/ijms22157944
Chicago/Turabian StyleAnnamneedi, Anil, Miguel del Angel, Eckart D. Gundelfinger, Oliver Stork, and Gürsel Çalışkan. 2021. "The Presynaptic Scaffold Protein Bassoon in Forebrain Excitatory Neurons Mediates Hippocampal Circuit Maturation: Potential Involvement of TrkB Signalling" International Journal of Molecular Sciences 22, no. 15: 7944. https://doi.org/10.3390/ijms22157944
APA StyleAnnamneedi, A., del Angel, M., Gundelfinger, E. D., Stork, O., & Çalışkan, G. (2021). The Presynaptic Scaffold Protein Bassoon in Forebrain Excitatory Neurons Mediates Hippocampal Circuit Maturation: Potential Involvement of TrkB Signalling. International Journal of Molecular Sciences, 22(15), 7944. https://doi.org/10.3390/ijms22157944