
An Overview of Adipose Tissue ACE2 Modulation by Diet and Obesity. Potential Implications in COVID-19 Infection and Severity

 ,
,  ,
, 
Abstract
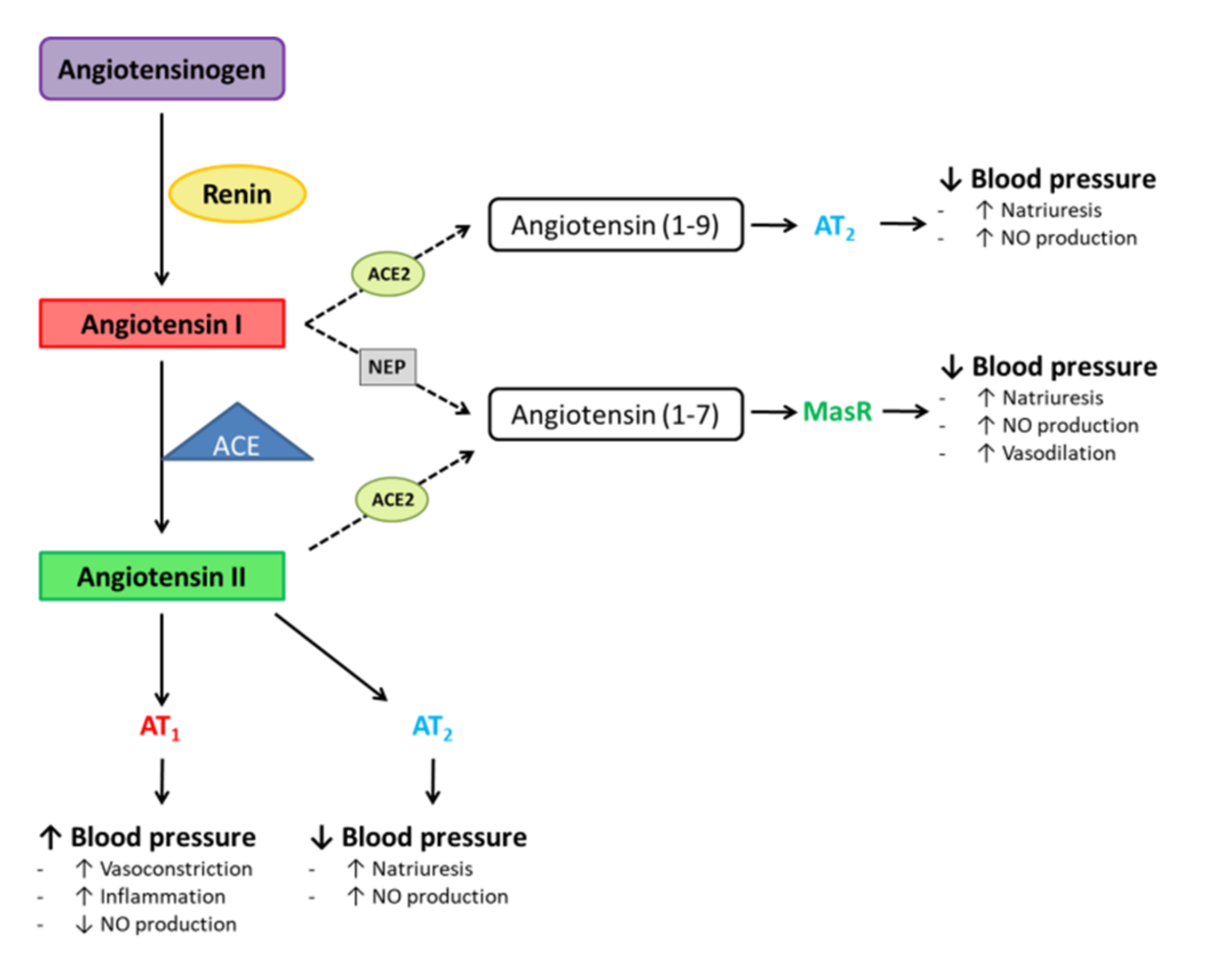
:1. Introduction
2. Influence of Different Dietary Patterns and Weight-Loss Strategies in Adipose Tissue ACE2 Levels. Evidence from Preclinical Studies
2.1. Preclinical Studies Carried Out in Dietary Rodent Models
2.1.1. Dietary Composition

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reference | Sample | Experimental Design | Observations on RAS |
|---|---|---|---|
| Gupte et al., 2008 [27] | Male C57BL/6 mice (8-week-old) | Mice fed with a LFD or a HFD (10 or 60% of energy as fat) for 1 week or 4 months. | ↑ circulating levels of angiotensin peptides I, II and IV were found after 4 months of HFD feeding. ↑ gene expression of angiotensinogen (epididymal AT) and Ace2 (epididymal and subcutaneous AT) in HFD fed mice after 1 week or 4 months. ↑ ACE2 protein expression in epididymal AT after 1 week of HFD feeding. ↓ ACE2 protein expression in epididymal AT after 4 months of HFD feeding. |
| Coelho 2010 [36] | Male Wistar rats | Rats fed with a standard diet for 1 months. A group of rats received sucrose in drinking water (20% solution). | ↑ plasma angiotensin I and II levels in sucrose fed animals. ↑ plasma renin and ACE enzyme activities in sucrose fed animals. ↑ angiotensin I, II and (1-7) levels in epididymal AT of sucrose fed rats. ↓ protein expression of ACE2 in epididymal AT of rats receiving sucrose. ↑ ACE protein expression and activity in epididymal AT of rats receiving sucrose. ↓ Agt gene expression in epididymal AT of rats receiving sucrose. ↑ AT1 and AT2 protein expression in epididymal AT of sucrose fed rats. |
| Gupte et al., 2012 [33] | Male and female C57BL/6 mice (8-week-old) | Mice fed with a LFD or a HFD (10 or 60% of energy as fat) for 4 months. | ↑ plasma angiotensin II levels in male mice fed the HFD compared to LFD-fed counterparts. ↓ plasma angiotensin (1-7) levels in LFD fed females compared to HFD fed males. ↓ plasma angiotensin (1-7) levels in HFD fed males compared to LFD fed males. ↑ plasma angiotensin (1-7) levels in HFD fed females compared to LFD fed males. ↑ AT ACE2 activity in HFD-fed males and females compared with LFD fed males. |
| Pinho 2013 [34] | Male FVB/N mice (8-week-old) | Mice fed with a STD, an AIN-93M, a HG, a HP or a HL diet for 2 months. | ↑ Ace gene expression in the epididymal AT of mice fed the HL diet compared to the groups fed with the STD and AIN-93M diets. ↓ Ace2 gene expression in epididymal AT of mice fed the HL diet compared to the animals fed the AIN-93M diet. |
| Zhang et al., 2014 [35] | Male Sprague-Dawley rats (8-week-old) | Rats fed with a STD or HFD (mainly lard) for 6 months | ↑ Ace2 gene and protein expressions in epididymal AT in HFD-fed rats. |
| De Almeida Pinheiro 2017 [37] | Male Swiss mice (4-week-old) | Mice fed with a STD or a HFD (61% of energy as fat) for 2 months. Animals fed with the two diets had free access to food or were submitted to 20, 40 or 60% of food restriction. | ↑ At1 gene expression in ad libitum HFD-fed mice compared to STD-fed controls. ↓ Agt and Ace gene expression in STD-fed animals submitted to energy restriction compared to ad libitum STD-fed counterparts. ↓ Ace gene expression in HFD-fed animals submitted to energy restriction compared to ad libitum HFD-fed counterparts. |
| Oliveira Andrade et al., 2014 [38] | Male FVB/N mice (4-week-old) | Mice fed with a STD or a HFD (11 or 61% of energy as fat) for 2 months. An additional group of animals fed with the HFD received 100 μg/kg bw/day of angiotensin (1-7). | ↓ Ace gene expression in epididymal AT of mice fed the HFD compared to mice fed the STD diet. ↓ Ace and Agt, and ↑ Ace2 gene expression in the epididymal AT of mice fed the HFD and treated with angiotensin (1-7), compared to the non-treated animals. |
| Crespo et al., 2017 [39] | Male Wistar rats (8-week-old) | Rats fed with a STD or a HFD (8 and 61% of energy as fat, respectively) for 2 months. Animals on each diet underwent L or SG. After surgical intervention, animals were fed the same diets for 1 additional month. | ↓ Gene expression of Agt in periepididymal AT in STD and HFD fed rats with SG compared to animals with L fed the STD and HFD. ↓ Ace gene expression in periepididymal AT in STD- and HFD-fed rats with SG compared to animals with L fed the HFD. ↑ Ace2 gene expression in periepididymal AT in HFD fed rats with SG compared to animals with L fed standard diet. |
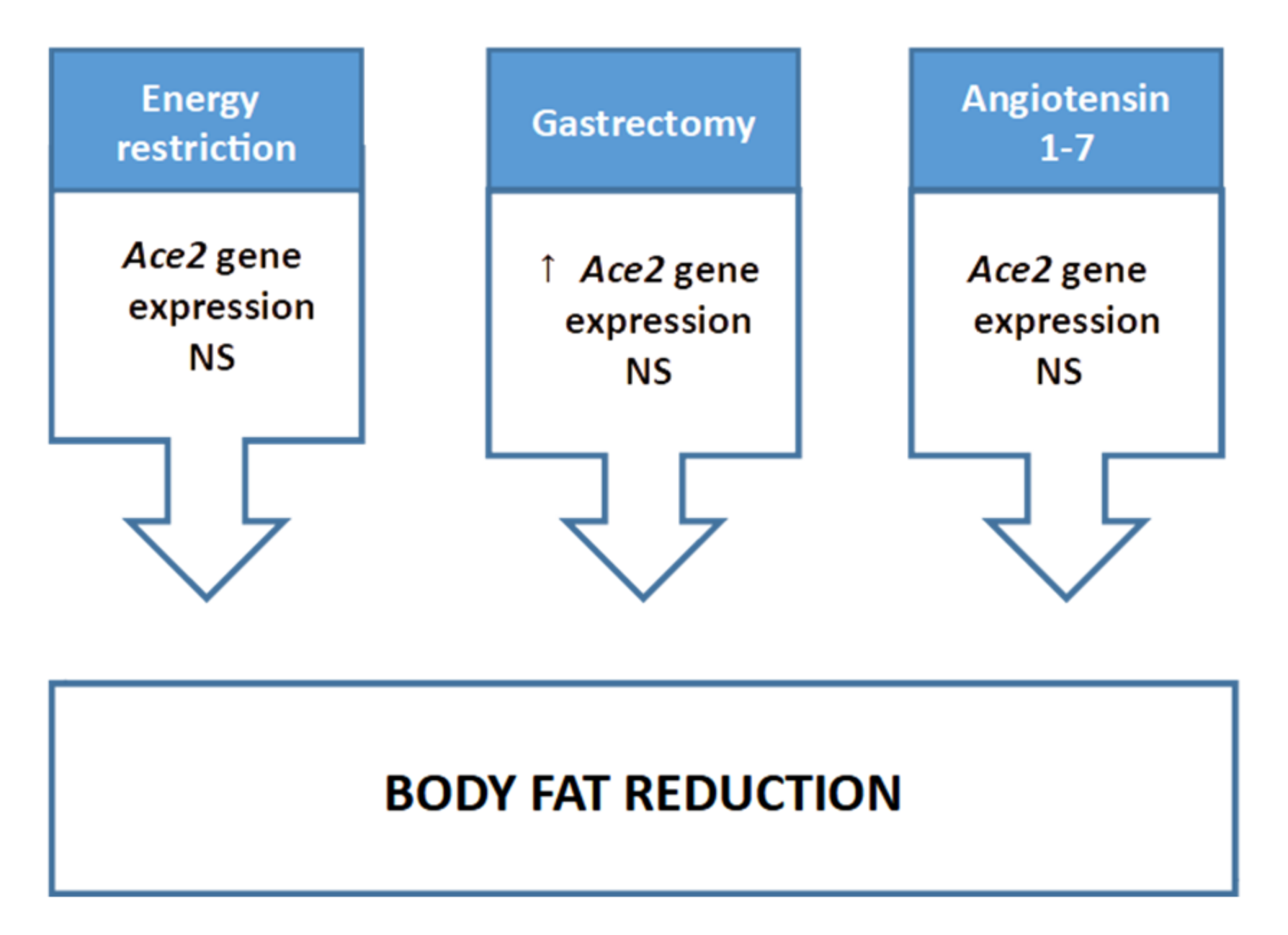
2.1.2. Energy Restriction
2.1.3. Non-Dietary Treatments Focused on Body Fat Reduction
2.1.4. Conclusions
2.2. Preclinical Studies Carried Out in Genetically Modified Rodent Models
3. Influence of Obesity and Diet in Adipose Tissue ACE2 Levels. Evidence from Studies Conducted in Humans
4. Influence of Obesity and ACE2 in COVID-19. beyond ACE2 Expression
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Available online: https://covid19.who.int/ (accessed on 5 June 2021).
- Lu, R.; Zhao, X.; Li, J.; Niu, P.; Yang, B.; Wu, H.; Wang, W.; Song, H.; Huang, B.; Zhu, N.; et al. Genomic characterisation and epidemiology of 2019 novel coronavirus: Implications for virus origins and receptor binding. Lancet 2020, 395, 565–574. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [Green Version]
- Qin, C.; Zhou, L.; Hu, Z.; Zhang, S.; Yang, S.; Tao, Y.; Xie, C.; Ma, K.; Shang, K.; Wang, W.; et al. Dysregulation of immune response in patients with COVID-19 in Wuhan, China. Clin. Infect. Dis. 2020, 71, 762–768. [Google Scholar] [CrossRef]
- Turner, R. Covid-19 and aerosol transmission: Up in the air. BMJ 2021, 372, n636. [Google Scholar] [CrossRef]
- Walls, A.C.; Park, Y.J.; Tortorici, M.A.; Wall, A.; McGuire, A.T.; Veesler, D. Structure, function, and antigenicity of the SARS-CoV-2 spike glycoprotein. Cell 2020, 181, 281–292. [Google Scholar] [CrossRef]
- Aksoy, H.; Karadag, A.S.; Wollina, U. Angiotensin II receptors: Impact for COVID-19 severity. Dermatol. Ther. 2020, 33, e13989. [Google Scholar] [CrossRef] [PubMed]
- Shang, J.; Ye, G.; Shi, K.; Wan, Y.; Luo, C.; Aihara, H.; Geng, Q.; Auerbach, A.; Li, F. Structural basis of receptor recognition by SARS-CoV-2. Nature 2020, 581, 221–224. [Google Scholar] [CrossRef] [Green Version]
- Krams, I.A.; Luoto, S.; Rantala, M.J.; Jõers, P.; Krama, T. Covid-19: Fat, obesity, inflammation, ethnicity, and sex differences. Pathogens 2020, 9, 887. [Google Scholar] [CrossRef]
- Spiegel, M.; Pichlmair, A.; Martínez-Sobrido, L.; Cros, J.; García-Sastre, A.; Haller, O.; Weber, F. Inhibition of beta interferon induction by severe acute respiratory syndrome coronavirus suggests a two-step model for activation of interferon regulatory factor 3. J. Virol. 2005, 79, 2079–2086. [Google Scholar] [CrossRef] [Green Version]
- Ferrario, C.M. Role of angiotensin II in cardiovascular disease—Therapeutic implications of more than a century of research. J. Renin Angiotensin Aldosterone Syst. 2006, 7, 3–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ocaranza, M.P.; Riquelme, J.A.; García, L.; Jalil, J.E.; Chiong, M.; Santos, R.A.S.; Lavandero, S. Counter-regulatory renin–angiotensin system in cardiovascular disease. Nat. Rev. Cardiol. 2020, 17, 116–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, H. Pulmonary Angiotensin-Converting Enzyme 2 (ACE2) and Inflammatory Lung Disease. Shock 2016, 46, 239–248. [Google Scholar] [CrossRef] [PubMed]
- Nie, W.; Zang, Y.; Chen, J.; Liu, T.; Xiao, L.; Xiu, Q. Angiotensin-converting enzyme I/D polymorphism is associated with pneumonia risk: A meta-analysis. J. Renin Angiotensin Aldosterone Syst. 2014, 15, 585–592. [Google Scholar] [CrossRef] [Green Version]
- Kuba, K.; Imai, Y.; Penninger, J.M. Angiotensin-converting enzyme 2 in lung diseases. Curr. Opin. Pharmacol. 2006, 6, 271–276. [Google Scholar] [CrossRef]
- Hrenak, J.; Simko, F. Renin-Angiotensin System: An Important Player in the Pathogenesis of Acute Respiratory Distress Syndrome. Int. J. Mol. Sci. 2020, 21, 8038. [Google Scholar] [CrossRef] [PubMed]
- Latil, M.; Camelo, S.; Veillet, S.; Lafont, R.; Dilda, P.J. Developing new drugs that activate the protective arm of the renin-angiotensin system as a potential treatment for repiratory failure in COVID-19 patients. Drug Discov. Today 2021. [Google Scholar] [CrossRef]
- Steckelings, U.M.; Sumners, C. Correcting the imbalanced protective RAS in COVID-19 with angiotensin AT2-receptor agonists. Clin. Sci. 2020, 134, 2987–3006. [Google Scholar] [CrossRef]
- Coutard, B.; Valle, C.; de Lamballerie, X.; Canard, B.; Seidah, N.G.; Decroly, E. The spike glycoprotein of the new coronavirus 2019-nCoV contains a furin-like cleavage site absent in CoV of the same clade. Antiviral Res. 2020, 176, 104742. [Google Scholar] [CrossRef]
- Jamal, M.; Bangash, H.I.; Habiba, M.; Lei, Y.; Xie, T.; Sun, J.; Wei, Z.; Hong, Z.; Shao, L.; Zhang, Q. Immune dysregulation and system pathology in COVID-19. Virulence 2021, 12, 918–936. [Google Scholar] [CrossRef]
- Gao, M.; Piernas, C.; Astbury, N.M.; Hippisley-Cox, J.; O’Rahilly, S.; Aveyard, P.; Jebb, S.A. Associations between body-mass index and COVID-19 severity in 6·9 million people in England: A prospective, community-based, cohort study. Lancet Diabetes Endocrinol. 2021, 9, 350–359. [Google Scholar] [CrossRef]
- Ryan, P.M.; Caplice, N.M. Is adipose tissue a reservoir for viral spread, immune activation, and cytokine amplification in coronavirus disease 2019? Obesity 2020, 28, 1191–1194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohammad, S.; Aziz, R.; Al Mahri, S.; Malik, S.S.; Haji, E.; Khan, A.H.; Khatlani, T.S.; Bouchama, A. Obesity and COVID-19: What makes obese host so vulnerable? Immun. Ageing 2021, 18, 1–10. [Google Scholar] [CrossRef]
- Klang, E.; Kassim, G.; Soffer, S.; Freeman, R.; Levin, M.A.; Reich, D.L. Severe obesity as an independent risk factor for COVID-19 mortality in hospitalized patients younger than 50. Obesity 2020, 28, 1595–1599. [Google Scholar] [CrossRef]
- Chu, Y.; Yang, J.; Shi, J.; Zhang, P.; Wang, X. Obesity is associated with increased severity of disease in COVID-19 pneumonia: A systematic review and meta-analysis. Eur. J. Med. Res. 2020, 25, 64. [Google Scholar] [CrossRef] [PubMed]
- Jia, X.; Yin, C.; Lu, S.; Chen, Y.; Liu, Q.; Bai, J.; Lu, Y. Two Things about COVID-19 Might Need Attention. Preprints 2020. [Google Scholar] [CrossRef]
- Gupte, M.; Boustany-Kari, C.M.; Bharadwaj, K.; Police, S.; Thatcher, S.; Gong, M.C.; English, V.L.; Cassis, L.A. ACE2 is expressed in mouse adipocytes and regulated by a high-fat diet. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2008, 295, R781–R788. [Google Scholar] [CrossRef] [Green Version]
- Biscarini, S.; Colaneri, M.; Ludovisi, S.; Seminari, E.; Pieri, T.C.; Valsecchi, P.; Gallazzi, I.; Giusti, E.; Cammà, C.; Zuccaro, V.; et al. The obesity paradox: Analysis from the SMAtteo COvid-19 REgistry (SMACORE) cohort. Nutr. Metab. Cardiovasc. Dis. 2020, 30, 1920–1925. [Google Scholar] [CrossRef] [PubMed]
- Schmieder, R.E.; Hilgers, K.F.; Schlaich, M.P.; Schmidt, B.M. Renin-angiotensin system and cardiovascular risk. Lancet 2007, 369, 1208–1219. [Google Scholar] [CrossRef]
- Tikellis, C.; Thomas, M.C. Angiotensin-Converting Enzyme 2 (ACE2) Is a Key Modulator of the Renin Angiotensin System in Health and Disease. Int. J. Pept. 2012, 2012, 256294. [Google Scholar] [CrossRef]
- Santos, S.H.; Braga, J.F.; Mario, E.G.; Porto, L.C.; Rodrigues-Machado, M.d.G.; Murari, A.; Botion, L.M.; Alenina, N.; Bader, M.; Santos, R.A. Improved lipid and glucose metabolism in transgenic rats with increased circulating angiotensin-(1-7). Arterioscler. Thromb. Vasc. Biol. 2010, 30, 953–961. [Google Scholar] [CrossRef] [Green Version]
- Santos, S.H.S.; Fernandes, L.R.; Pereira, C.S.; Guimarães, A.L.S.; de Paula, A.M.; Campagnole-Santos, M.J.; Alvarez-Leite, J.I.; Bader, M.; Santos, R.A.S. Increased circulating angiotensin-(1–7) protects white adipose tissue against development of a proinflammatory state stimulated by a high-fat diet. Regul. Pept. 2012, 178, 64–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupte, M.; Thatcher, S.E.; Boustany-Kari, C.M.; Shoemaker, R.; Yiannikouris, F.; Zhang, X.; Karounos, M.; Cassis, L.A. Angiotensin converting enzyme 2 contributes to sex differences in the development of obesity hypertension in C57BL/6 mice. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 1392–1399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Pinho, L.; Andrade, J.M.O.; Paraíso, A.; Filho, A.B.M.; Feltenberger, J.D.; Guimaraes, A.L.S.; de Paula, A.M.B.; Caldeira, A.P.; de Carvalho Botelho, A.C.C.; Campagnole-Santos, M.J.; et al. Diet composition modulates expression of sirtuins and renin-angiotensin system components in adipose tissue. Obesity 2013, 21, 1830–1835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Xu, Y.-Z.; Liu, B.; Wu, R.; Yang, Y.-Y.; Xiao, X.-Q.; Zhang, X. Pioglitazone Upregulates Angiotensin Converting Enzyme 2 Expression in Insulin-Sensitive Tissues in Rats with High-Fat Diet-Induced Nonalcoholic Steatohepatitis. Sci. World J. 2014, 2014, 603409. [Google Scholar] [CrossRef]
- Coelho, M.S.; Lopes, K.L.; de Aquino Freitas, R.; de Oliveira-Sales, E.B.; Bergasmaschi, C.T.; Campos, R.R.; Casarini, D.E.; Carmona, A.K.; Araújo, M.D.; Heimann, J.C.; et al. High sucrose intake in rats is associated with increased ACE2 and angiotensin-(1–7) levels in the adipose tissue. Regul. Pept. 2010, 162, 61–67. [Google Scholar] [CrossRef]
- Pinheiro, T.D.A.; Barcala-Jorge, A.S.; Andrade, J.M.O.; Ferreira, E.C.N.; Crespo, T.; Batista-Jorge, G.C.; Vieira, C.A.; Lelis, D.D.F.; Paraíso, A.F.; Pinheiro, U.B.; et al. Obesity and malnutrition similarly alter the renin–angiotensin system and inflammation in mice and human adipose. J. Nutr. Biochem. 2017, 48, 74–82. [Google Scholar] [CrossRef]
- Andrade, J.M.O.; Paraíso, A.F.; Garcia, Z.M.; Ferreira, A.V.M.; Sinisterra, R.D.; De Sousa, F.B.; Guimarães, A.L.S.; De Paula, A.M.B.; Santos, M.J.C.; Dos Santos, R.A.; et al. Cross talk between angiotensin-(1–7)/Mas axis and sirtuins in adipose tissue and metabolism of high-fat feed mice. Peptides 2014, 55, 158–165. [Google Scholar] [CrossRef]
- Crespo, T.S.; Andrade, J.M.O.; Paraiso, A.F.; De Farias Lelis, D.; Chagas, P.V.F.; Jorge, A.S.B.; Santos, S.H.S.; Sena, A.L.; Batista, A.M.; Paula, D.; et al. Effects of Sleeve Gastrectomy on the Metabolic Profile and on the Expression of Renin-Angiotensin System in Adipose Tissue of Obese Rats. Protein Pept. Lett. 2017, 24, 861–868. [Google Scholar] [CrossRef]
- Patel, V.B.; Mori, J.; McLean, B.A.; Basu, R.; Das, S.K.; Ramprasath, T.; Parajuli, N.; Penninger, J.M.; Grant, M.B.; Lopaschuk, G.D.; et al. ACE2 deficiency worsens epicardial adipose tissue inflammation and cardiac dysfunction in response to diet-induced obesity. Diabetes 2016, 65, 85–95. [Google Scholar] [CrossRef] [Green Version]
- Shoemaker, R.; Tannock, L.R.; Su, W.; Gong, M.; Gurley, S.B.; Thatcher, S.E.; Yiannikouris, F.; Ensor, C.M.; Cassis, L.A. Adipocyte deficiency of ACE2 increases systolic blood pressures of obese female C57BL/6 mice. Biol. Sex Differ. 2019, 10, 45. [Google Scholar] [CrossRef] [Green Version]
- Al-Benna, S. Association of high level gene expression of ACE2 in adipose tissue with mortality of COVID-19 infection in obese patients. Obes. Med. 2020, 19, 100283. [Google Scholar] [CrossRef]
- Al Heialy, S.; Hachim, M.Y.; Senok, A.; Gaudet, M.; Abou Tayoun, A.; Hamoudi, R.; Alsheikh-Ali, A.; Hamid, Q. Regulation of angiotensin-converting enzyme 2 in obesity: Implications for COVID-19. Front. Physiol. 2020, 11, 555039. [Google Scholar] [CrossRef]
- Li, L.; Spranger, L.; Soll, D.; Beer, F.; Brachs, M.; Spranger, J.; Mai, K. Metabolic impact of weight loss induced reduction of adipose ACE-2-Potential implication in COVID-19 infections? Metabolism 2020, 113, 154401. [Google Scholar] [CrossRef]
- Kristem, L.; Recamonde-Mendoza, M.; Cigerza, G.C.; Khoraki, J.; Campos, G.M.; Mazzini, G.S. Roux-en-Y Gastric Bypass Downregulates Angiotensin-Converting Enzyme 2 (ACE2) Gene Expression in Subcutaneous White Adipose Tissue: A Putative Protective Mechanism Against Severe COVID-19. Obes. Surgery 2021, 31, 2831–2834. [Google Scholar] [CrossRef]
- El-Sayed Moustafa, J.S.; Jackson, A.U.; Brotman, S.M.; Guan, L.; Villicaňa, S.; Roberts, A.L.; Zito, A.; Bonnycastle, L.; Erdos, M.R.; Narisu, N.; et al. ACE2 expression in adipose tissue is associated with COVID-19 cardio-metabolic risk factors and cell type composition. medRxiv 2020. [Google Scholar] [CrossRef]
- Johns Hopkins University. Available online: https://coronavirus.jhu.edu/data/mortality (accessed on 14 July 2021).
- Tamama, K. Potential benefits of dietary seaweeds as protection against COVID-19. Nutr. Rev. 2020. [Google Scholar] [CrossRef]
- Trypsteen, W.; Van Cleemput, J.; Snippenberg, W.V.; Gerlo, S.; Vandekerckhove, L. On the whereabouts of SARS-CoV-2 in the human body: A systematic review. PLoS Pathog. 2020, 16, e1009037. [Google Scholar] [CrossRef]
- Gu, H.; Xie, Z.; Li, T.; Zhang, S.; Lai, C.; Zhu, P.; Wang, K.; Han, L.; Duan, Y.; Zhao, Z.; et al. Angiotensin-converting enzyme 2 inhibits lung injury induced by respiratory syncytial virus. Sci. Rep. 2016, 6, 19840. [Google Scholar] [CrossRef]
- Drucker, D.J. Diabetes, obesity, metabolism, and SARS-CoV-2 infection: The end of the beginning. Cell Metab. 2021, 33, 479–498. [Google Scholar] [CrossRef]
- Banerjee, M.; Gupta, S.; Sharma, P.; Shekhawat, J.; Gauba, K. Obesity and COVID-19: A Fatal Alliance. Indian J. Clin. Biochem. 2020, 35, 410–417. [Google Scholar] [CrossRef] [PubMed]
- Engin, A.B.; Engin, E.D.; Engin, A. Two important controversial risk factors in SARS-CoV-2 infection: Obesity and smoking. Environ. Toxicol. Pharmacol. 2020, 78, 103411. [Google Scholar] [CrossRef] [PubMed]
- Kruglikov, I.L.; Scherer, P.E. The Role of Adipocytes and Adipocyte-Like Cells in the Severity of COVID-19 Infections. Obesity 2020, 28, 1187–1190. [Google Scholar] [CrossRef] [PubMed]
- Longo, M.; Zatterale, F.; Naderi, J.; Parrillo, L.; Formisano, P.; Raciti, G.A.; Beguinot, F.; Miele, C. Adipose Tissue Dysfunction as Determinant of Obesity-Associated Metabolic Complications. Int. J. Mol. Sci. 2019, 20, 2358. [Google Scholar] [CrossRef] [Green Version]
- Emilsson, V.; Gudmundsson, E.F.; Aspelund, T.; Jonsson, B.G.; Gudjonsson, A.; Launer, L.J.; Lamb, J.R.; Gudmundsdottir, V.; Jennings, L.L.; Gudnason, V. Serum levels of ACE2 are higher in patients with obesity and diabetes. Obes. Sci. Pract. 2021, 7, 239–243. [Google Scholar] [CrossRef] [PubMed]
- Emilsson, V.; Gudmundsson, E.F.; Aspelund, T.; Jonsson, B.G.; Gudjonsson, A.; Launer, L.J.; Lamb, J.R.; Gudmundsdottir, V.; Jennings, L.L.; Gudnason, V. ACE2 levels are altered in comorbidities linked to severe outcome in COVID-19. Medrxiv Prepr. Serv. Health Sci. 2020. [Google Scholar] [CrossRef]
- Van Lier, D.; Kox, M.; Santos, K.; van der Hoeven, H.; Pillay, J.; Pickkers, P. Increased blood angiotensin converting enzyme 2 activity in critically Ill COVID-19 patients. ERJ Open Res. 2021, 00848–02020. [Google Scholar] [CrossRef]
- Lundström, A.; Ziegler, L.; Havervall, S.; Rudberg, A.S.; Von Meijenfeldt, F.; Lisman, P.T.; Mackman, N.; Sandén, P.; Thålin, C. Soluble angiotensin-converting enzyme 2 is transiently elevated in COVID-19 and correlates with specific inflammatory and endothelial markers. J. Med. Virol. 2021. [Google Scholar] [CrossRef]
- Kragstrup, T.W.; Singh, H.S.; Grundberg, I.; Nielsen, A.L.L.; Rivellese, F.; Mehta, A.; Goldberg, M.B.; Filbin, M.R.; Qvist, P.; Bibby, B.M. Plasma ACE2 predicts outcome of COVID-19 in hospitalized patients. PLoS ONE 2021, 16, e0252799. [Google Scholar] [CrossRef]
- Ni, W.; Yang, X.; Yang, D.; Bao, J.; Li, R.; Xiao, Y.; Hou, C.; Wang, H.; Liu, J.; Yang, D.; et al. Role of angiotensin-converting enzyme 2 (ACE2) in COVID-19. Crit. Care 2020, 24. [Google Scholar] [CrossRef]
- Vardavas, C.I.; Nikitara, K. COVID-19 and smoking: A systematic review of the evidence. Tob. Induc. Dis. 2020, 18, 20. [Google Scholar] [CrossRef]
- Ghosh, S.; Klein, R.S. Sex Drives Dimorphic Immune Responses to Viral Infections. J. Immunol. 2017, 198, 1782–1790. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gómez-Zorita, S.; Milton-Laskibar, I.; García-Arellano, L.; González, M.; Portillo, M.P. An Overview of Adipose Tissue ACE2 Modulation by Diet and Obesity. Potential Implications in COVID-19 Infection and Severity. Int. J. Mol. Sci. 2021, 22, 7975. https://doi.org/10.3390/ijms22157975
Gómez-Zorita S, Milton-Laskibar I, García-Arellano L, González M, Portillo MP. An Overview of Adipose Tissue ACE2 Modulation by Diet and Obesity. Potential Implications in COVID-19 Infection and Severity. International Journal of Molecular Sciences. 2021; 22(15):7975. https://doi.org/10.3390/ijms22157975
Chicago/Turabian StyleGómez-Zorita, Saioa, Iñaki Milton-Laskibar, Laura García-Arellano, Marcela González, and María P. Portillo. 2021. "An Overview of Adipose Tissue ACE2 Modulation by Diet and Obesity. Potential Implications in COVID-19 Infection and Severity" International Journal of Molecular Sciences 22, no. 15: 7975. https://doi.org/10.3390/ijms22157975
APA StyleGómez-Zorita, S., Milton-Laskibar, I., García-Arellano, L., González, M., & Portillo, M. P. (2021). An Overview of Adipose Tissue ACE2 Modulation by Diet and Obesity. Potential Implications in COVID-19 Infection and Severity. International Journal of Molecular Sciences, 22(15), 7975. https://doi.org/10.3390/ijms22157975