
Biocontrol of Biofilm Formation: Jamming of Sessile-Associated Rhizobial Communication by Rhodococcal Quorum-Quenching

 ,
,  , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
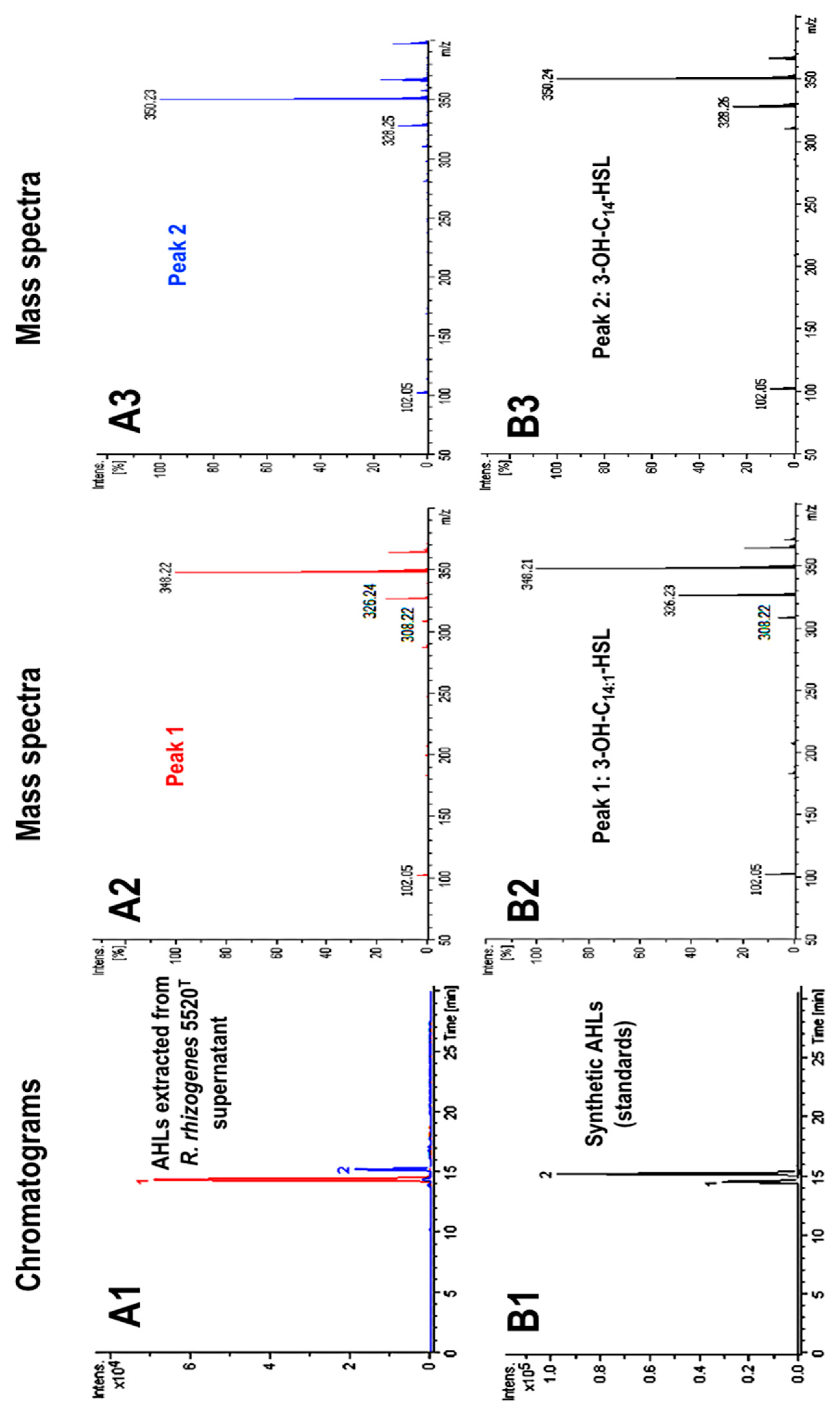
2.1. Characterization of the AHL Signaling Molecules Produced by R. rhizogenes
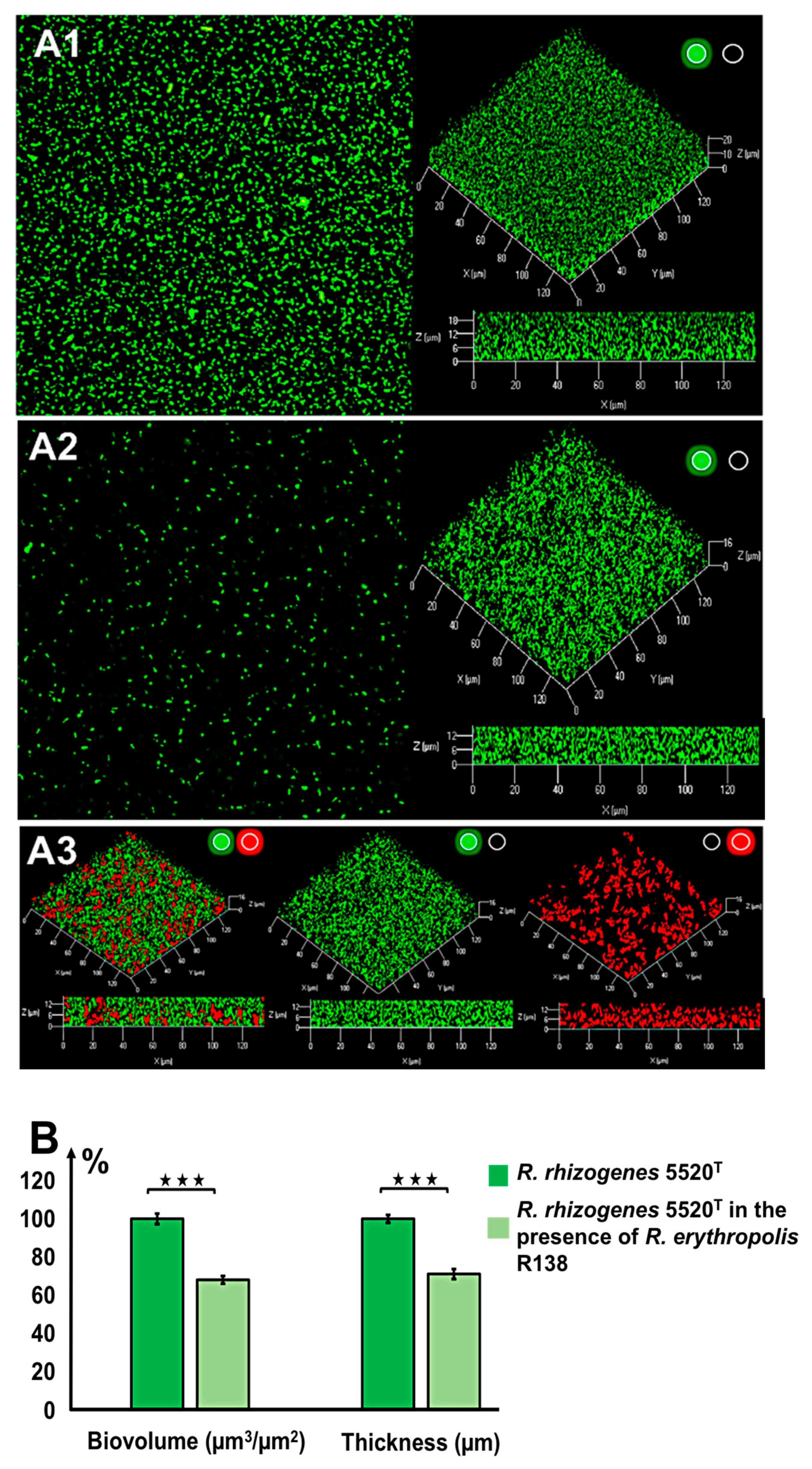
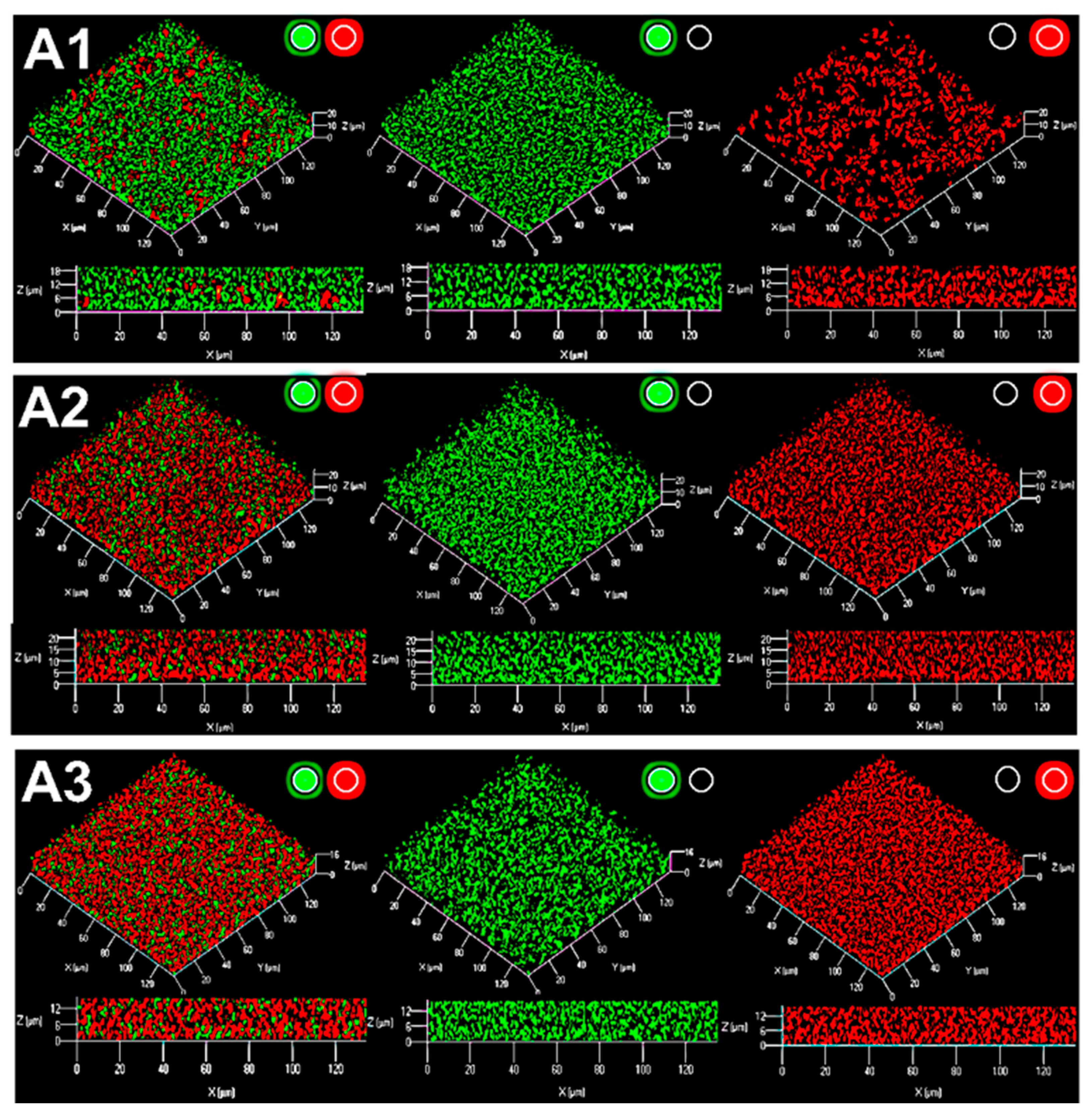
2.2. Characterization of the Structure of the Biofilm Generated by R. rhizogenes and Inhibition by the Biocontrol Agent R. erythropolis R138
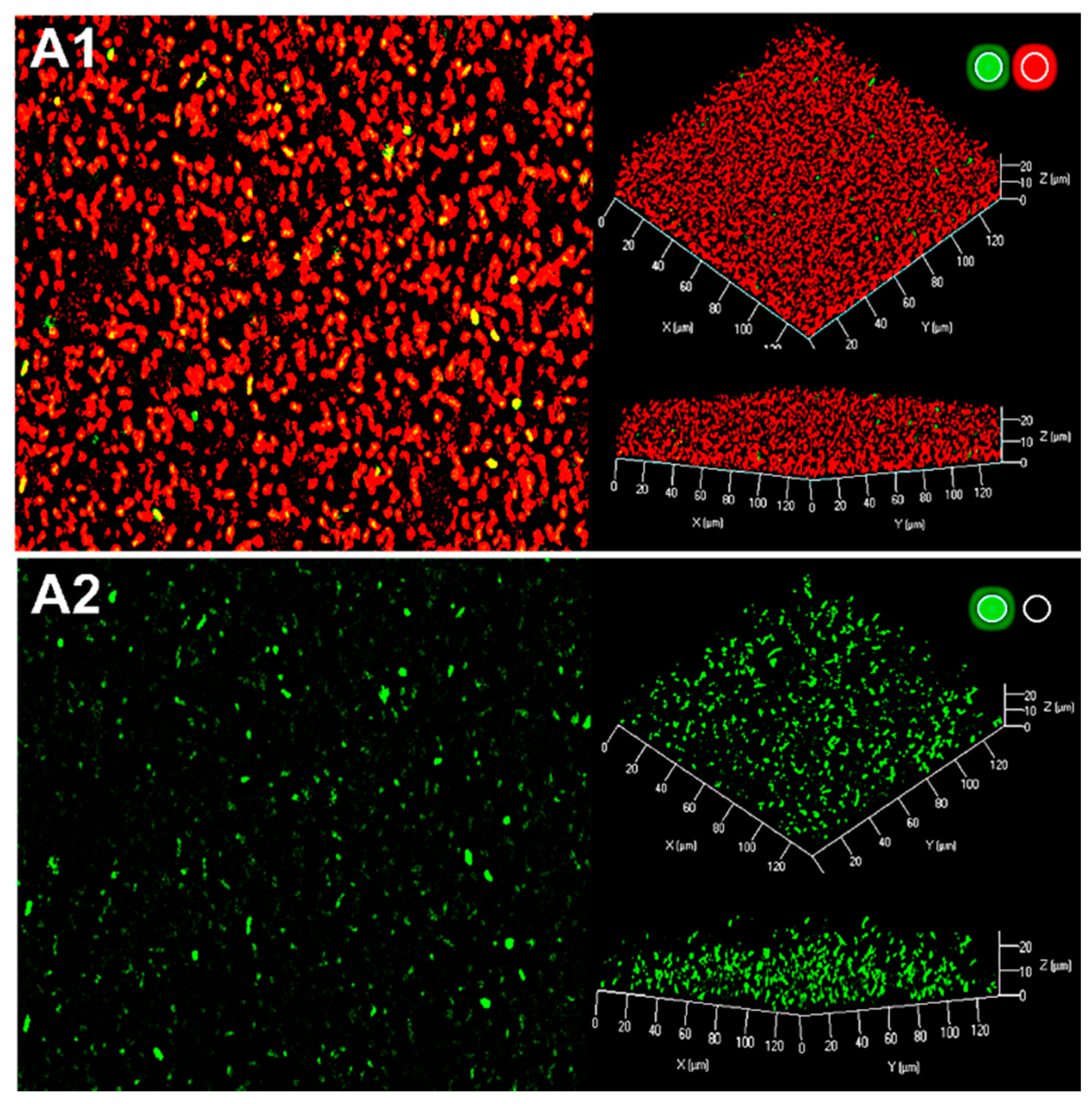
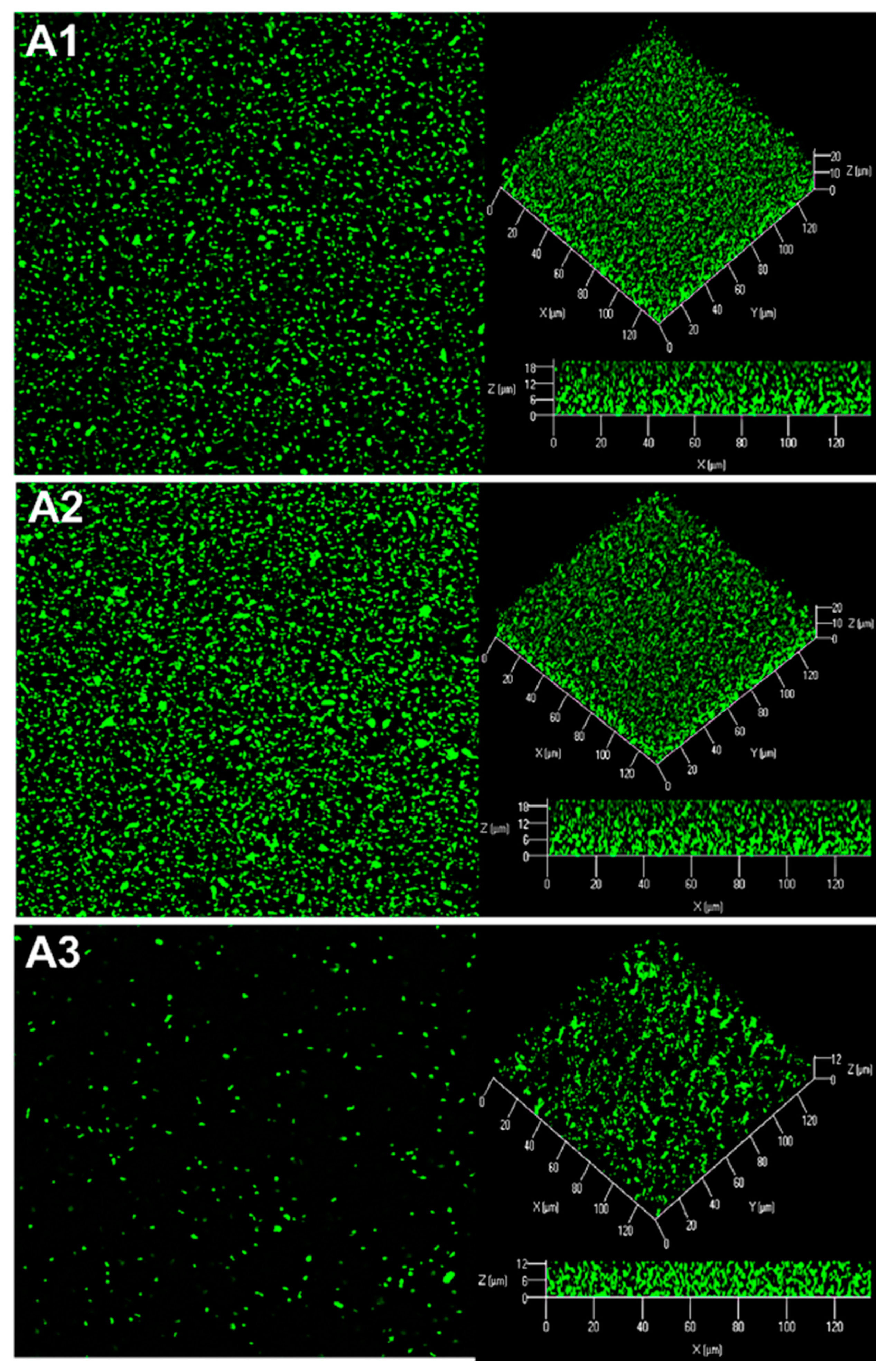
2.3. Production of AHL Signaling Molecules by R. rhizogenes within the Biofilm
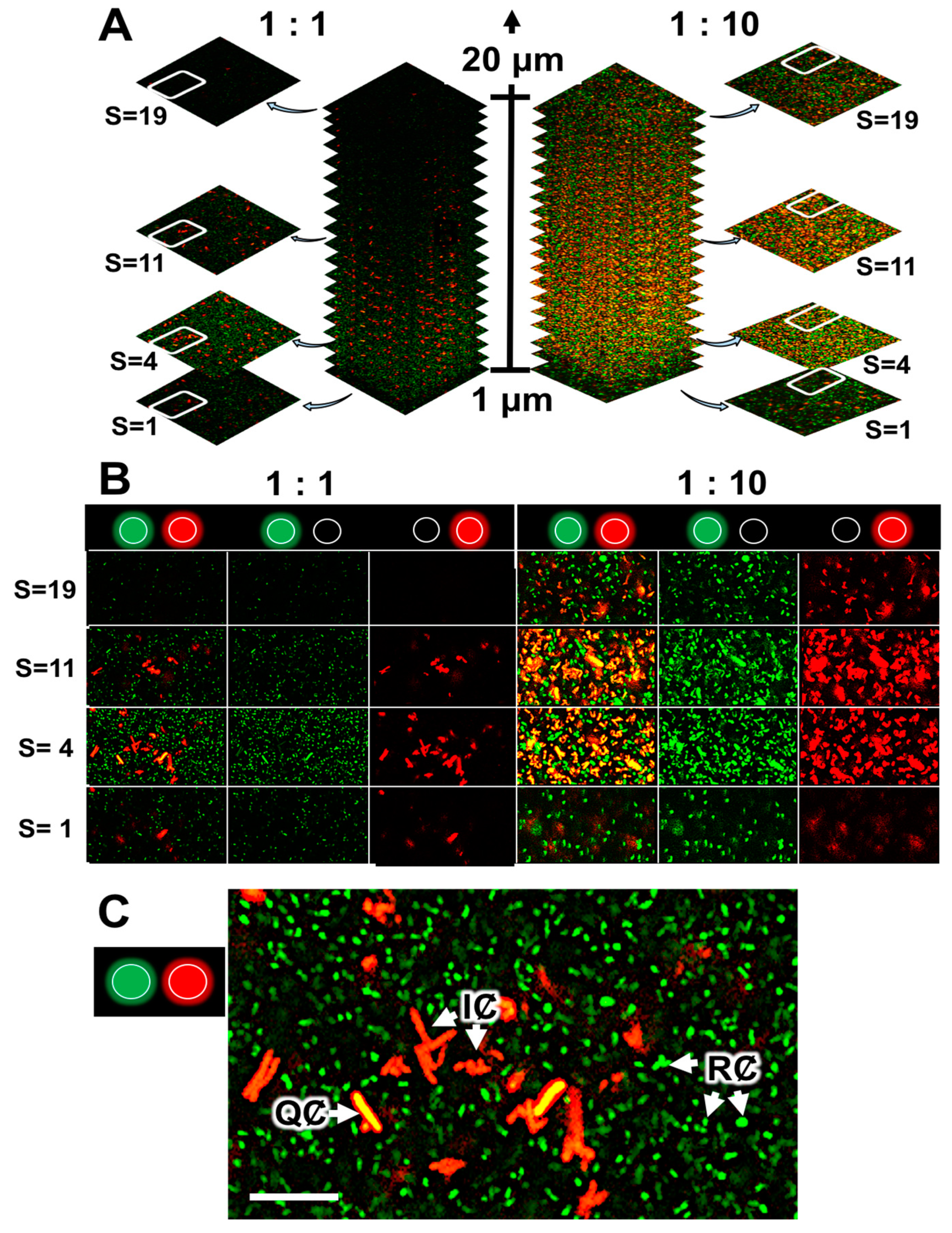
2.4. Quenching of R. rhizogenes AHL-Based Communication within the Biofilm by the Biocontrol Agent R. erythropolis R138
2.5. Rhodococcal Quorum-Quenching of Rhizobial AHL Communication Was Responsible for the Observed Changes in the Biofilm
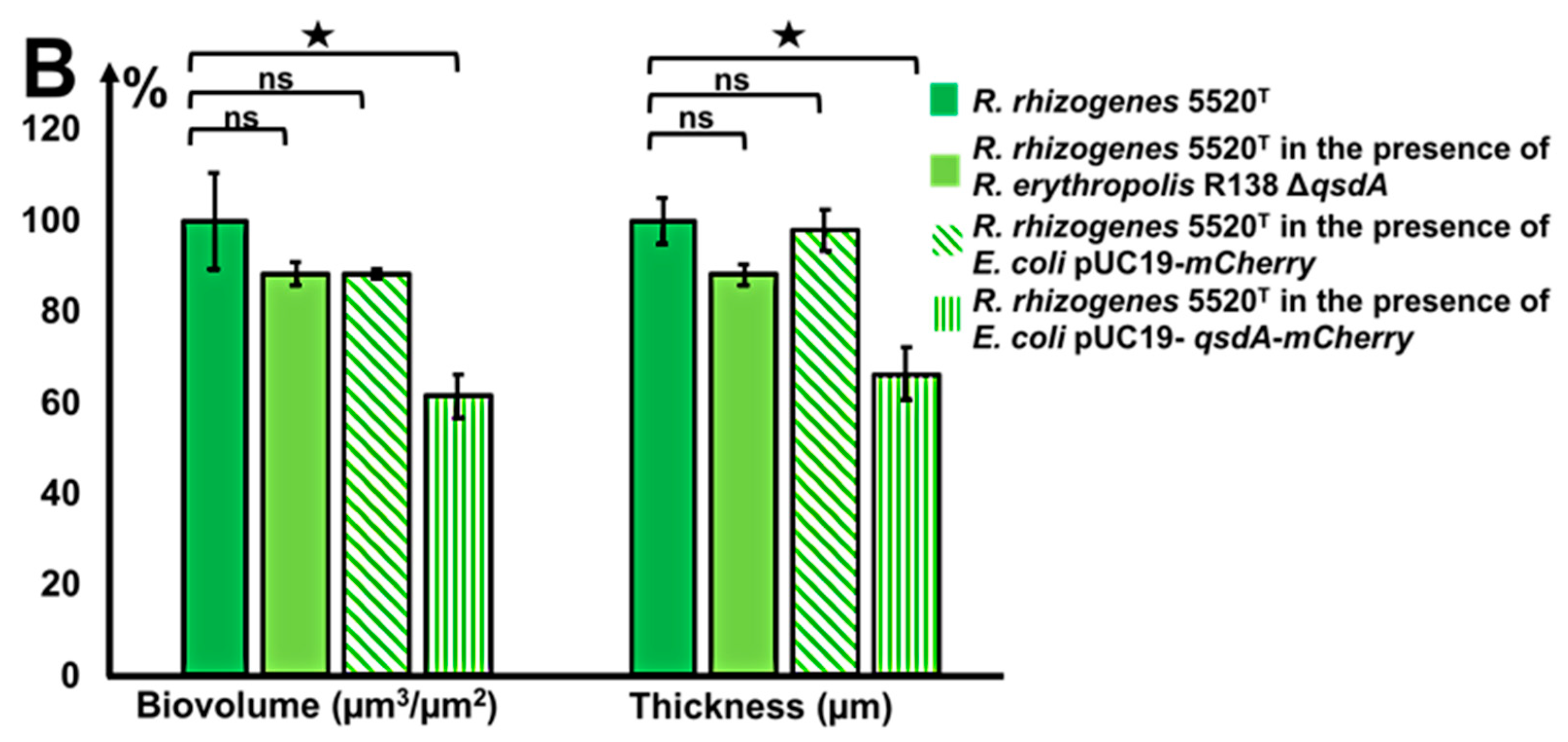
2.5.1. A Rhodococcus Mutant with A Deletion of the AHL-QsdA Lactonase Gene Lost Its Full Ability to Inhibit the Formation of Rhizobial Biofilms
2.5.2. A Heterologous Host Carrying the Gene Encoding the Rhodococcal QsdA Lactonase Acquired the Ability to Inhibit Formation of the Rhizobial Biofilm
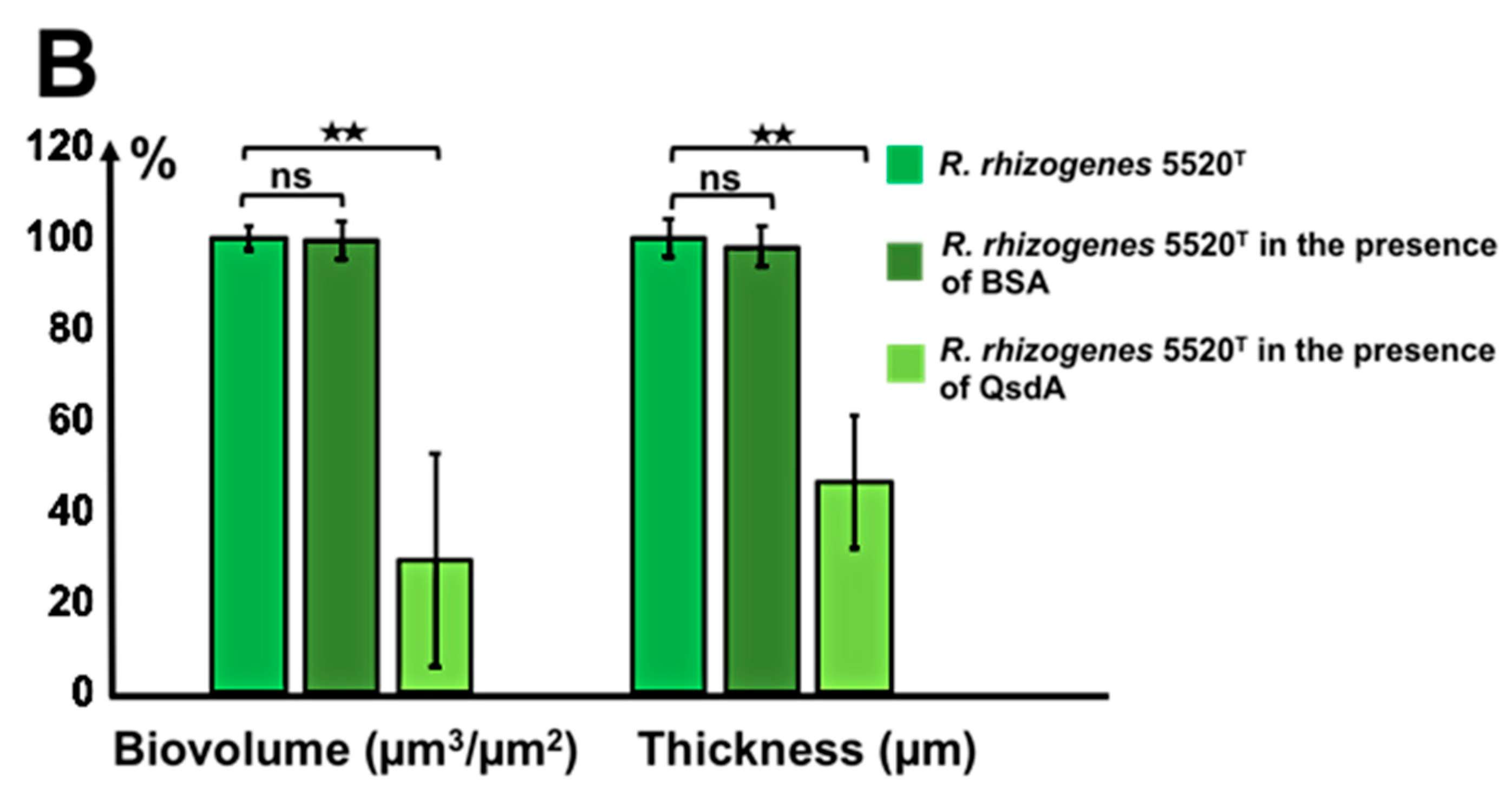
2.5.3. The Activity of the Rhodococcal Lactonase QsdA Was Sufficient to Inhibit Formation of the Rhizobial Biofilm
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Culture Conditions
4.2. Extraction and Characterization of the AHL Signaling Molecules Produced by R. rhizogenes 5520T
4.3. Construction of mCherry-Tagged E. coli Strains
4.4. Extraction and Purification of the QsdA Lactonase Enzyme
4.5. Biofilm Culture
4.6. Confocal Laser Scanning Microscopy (CLSM)
4.7. Antibiosis Test
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Costerton, J.W.; Stewart, P.S.; Greenberg, E.P. Bacterial biofilms: A common cause of persistent infections. Science 1999, 284, 1318–1322. [Google Scholar] [CrossRef] [Green Version]
- Flemming, H.C.; Wuertz, S. Bacteria and archaea on Earth and their abundance in biofilms. Nat. Rev. Microbiol. 2019, 17, 247–260. [Google Scholar] [CrossRef]
- Morris, C.E.; Monier, J.M. The ecological significance of biofilm formation by plant-associated bacteria. Ann. Rev. Phytopathol. 2003, 41, 236–241. [Google Scholar] [CrossRef]
- Flemming, H.C.; Wingender, J. The biofilm matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef] [PubMed]
- O’Toole, G.; Kaplan, H.B.; Kolter, R. Biofilm formation as microbial development. Annu. Rev. Microbiol. 2000, 54, 49–79. [Google Scholar] [CrossRef] [PubMed]
- Stoodley, P.; Sauer, K.; Davies, D.G.; Costerton, J.W. Biofilms as complex differentiated communities. Annu. Rev. Microbiol. 2002, 56, 187–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paluch, E.; Rewak-Soroczyńska, J.; Jędrusik, I.; Mazurkiewicz, E.; Jermakow, K. Prevention of biofilm formation by quorum quenching. Appl. Microbiol. Biotechnol. 2020, 104, 1871–1881. [Google Scholar] [CrossRef] [Green Version]
- Parsek, M.R.; Greenberg, E.P. Sociomicrobiology: The connections between quorum sensing and biofilms. Trends Microbiol. 2005, 13, 27–33. [Google Scholar] [CrossRef]
- Stewart, P.S.; Franklin, M.J. Physiological heterogeneity in biofilms. Nat. Rev. Microbiol. 2008, 6, 199–210. [Google Scholar] [CrossRef]
- Dieltjens, L.; Appermans, K.; Lissens, M.; Lories, B.; Kim, W.; Van de Eycken, E.V.; Foster, K.R.; Steenaker, H.P. Inhibiting bacterial cooperation is an evolutionarily robust anti-biofilm strategy. Nat. Commun. 2020, 11, 107. [Google Scholar] [CrossRef]
- de Kievit, T.R. Quorum sensing in Pseudomonas aeruginosa biofilms. Environ. Microbiol. 2009, 11, 279–288. [Google Scholar] [CrossRef] [PubMed]
- Latour, X. The Evanescent GacS Signal. Microorganisms 2020, 8, 1746. [Google Scholar] [CrossRef] [PubMed]
- Moradali, M.F.; Ghods, S.; Rehm, B.H. Pseudomonas aeruginosa lifestyle: A paradigm for adaptation, survival, and persistence. Front. Cell Infect. Microbiol. 2017, 7, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valentini, M.; Gonzalez, D.; Mavridou, D.A.; Filloux, A. Lifestyle transitions and adaptive pathogenesis of Pseudomonas aeruginosa. Curr. Opin. Microbiol. 2018, 41, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Heindl, J.E.; Wang, Y.; Heckel, B.C.; Mohari, B.; Feirer, N.; Fuqua, C. Mechanisms and regulation of surface interactions and biofilm formation in Agrobacterium. Front. Plant Sci. 2014, 5. [Google Scholar] [CrossRef] [Green Version]
- Solano, C.; Echeverz, M.; Lasa, I. Biofilm dispersion and quorum sensing. Curr. Opin. Microbiol. 2014, 18, 96–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papenfort, K.; Bassler, B. Quorum-sensing signal-response systems in Gram-negative bacteria. Nat. Rev. Microbiol. 2016, 14, 576–588. [Google Scholar] [CrossRef]
- LaSarre, B.; Federle, M.J. Exploiting quorum sensing to confuse bacterial pathogens. Microbiol. Mol. Biol. Rev. 2013, 77, 73–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Schaefer, A.L.; Dandekar, A.A.; Greenberg, E.P. Quorum sensing and policing of Pseudomonas aeruginosa social cheaters. Proc. Natl. Acad. Sci. USA 2015, 112, 2187–2191. [Google Scholar] [CrossRef] [Green Version]
- Heilmann, S.; Krishna, S.; Kerr, B. Why do bacteria regulate public goods by quorum sensing?-How the shapes of cost and benefit functions determine the form of optimal regulation. Front. Microbiol. 2015, 6, 767. [Google Scholar] [CrossRef]
- Chow, J.Y.; Yang, Y.; Tay, S.B.; Chua, K.L.; Yew, W.S. Disruption of biofilm formation by the human pathogen Acinetobacter baumannii using engineered quorum-quenching lactonases. Antimicrob. Agents Chemother. 2014, 58, 1802–1805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brackman, G.; Coenye, T. Quorum sensing inhibitors as anti-biofilm agents. Curr. Pharm. Des. 2015, 21, 5–11. [Google Scholar] [CrossRef]
- Hraiech, S.; Hiblot, J.; Lafleur, J.; Lepidi, H.; Papazian, L.; Rolain, J.M.; Raoult, D.; Elias, M.; Silby, M.W.; Bzdrenga, J.; et al. Inhaled lactonase reduces Pseudomonas aeruginosa quorum sensing and mortality in rat pneumonia. PLoS ONE 2014, 9, e107125. [Google Scholar] [CrossRef] [PubMed]
- Guendouze, A.; Plener, L.; Bzdrenga, J.; Jacquet, P.; Rémy, B.; Elias, M.; Lavigne, J.P.; Daudé, D.; Chabrière, E. Effect of quorum quenching lactonase in clinical isolates of Pseudomonas aeruginosa and comparison with quorum sensing inhibitors. Front. Microbiol. 2017, 8, 227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choo, K.-H.; Park, P.-K.; Oh, H.-S. Quorum sensing and quorum quenching in membrane bioreactors. In Current Developments in Biotechnology and Bioengineering; Ng, H.Y., Ng, T.C.A., Ngo, H.H., Mannina, G., Pandey, A., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 245–274. [Google Scholar] [CrossRef]
- Muras, A.; Parga, A.; Mayer, C.; Otero, A. Use of quorum sensing inhibition strategies to control microfouling. Mar. Drugs 2021, 19, 74. [Google Scholar] [CrossRef]
- Grandclément, C.; Tannières, M.; Moréra, S.; Dessaux, Y.; Faure, D. Quorum quenching: Role in nature and applied develop- ments. FEMS Microbiol. Rev. 2016, 40, 86–116. [Google Scholar] [CrossRef]
- Kim, S.-R.; Lee, K.-B.; Kim, J.-E.; Won, Y.-J.; Yeon, K.-M.; Lee, C.-H.; Lim, D.-J. Macroencapsulation of quorum quenching bacteria by polymeric membrane layer and its application to MBR for biofouling control. J. Membrane Sci. 2015, 473, 109–117. [Google Scholar] [CrossRef]
- Mishra, R.; Panda, A.K.; De Mandal, S.; Shakeel, M.; Bisht, S.S.; Khan, J. Natural anti-biofilm agents: Strategies to control biofilm-forming pathogens. Front. Microbiol. 2020, 11, 566325. [Google Scholar] [CrossRef]
- Vogel, J.; Quax, W.J. Enzymatic quorum quenching in biofilms. In Quorum Sensing: Molecular Mechanism and Biotechnological Application; Tommonaro, G., Ed.; Academic Press: Cambridge, MA, USA, 2019; pp. 173–193. [Google Scholar] [CrossRef]
- Dong, Y.H.; Wang, L.H.; Xu, J.L.; Zhang, H.B.; Zhang, X.F.; Zhang, L.H. Quenching quorum-sensing-dependent bacterial infec- tion by an N-acyl homoserine lactonase. Nature 2001, 411, 813–817. [Google Scholar] [CrossRef]
- Dong, Y.H.; Wang, L.H.; Zhang, L.H. Quorum-quenching microbial infections: Mechanisms and implications. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2007, 362, 1201–1211. [Google Scholar] [CrossRef]
- Vesuna, A.; Nerurkar, A.S. Enzymatic quorum quenching for virulence attenuation of phytopathogenic bacteria. In Biotechno-Logical Applications of Quorum Sensing Inhibitors; Kalia, V., Ed.; Springer: Singapore, 2018; pp. 447–473. [Google Scholar] [CrossRef]
- Bergonzi, C.; Schwab, M.; Naik, T.; Elias, M. The structural determinants accounting for the broad substrate specificity of the quorum quenching lactonase GcL. ChemBioChem 2019, 20, 1848–1855. [Google Scholar] [CrossRef]
- Fetzner, S. Quorum quenching enzymes. J. Biotechnol. 2015, 201, 2–14. [Google Scholar] [CrossRef] [PubMed]
- Ryu, D.H.; Lee, S.W.; Mikolaityte, V.; Kim, Y.W.; Jeong, H.Y.; Lee, S.J.; Lee, C.H.; Lee, J.K. Identification of a second type of AHL-lactonase from Rhodococcus sp. BH4, belonging to the α/β hydrolase superfamily. J. Microbiol. Biotechnol. 2020, 30, 937–945. [Google Scholar] [CrossRef]
- Crépin, A.; Barbey, C.; Cirou, A.; Tannières, M.; Orange, N.; Feuilloley, M.; Dessaux, Y.; Burini, J.-F.; Faure, D.; Latour, X. Biological control of pathogen communication in the rhizosphere: A novel approach applied to potato soft rot due to Pectobacterium atrosepticum. Plant Soil 2012, 358, 27–37. [Google Scholar] [CrossRef]
- Chane, A.; Barbey, C.; Robert, M.; Merieau, A.; Konto-Ghiorghi, Y.; Beury-Cirou, A.; Feuilloley, M.; Pátek, M.; Gobert, V.; Latour, X. Biocontrol of soft-rot: Confocal microscopy highlights virulent pectobacterial communication and its jamming by rhodococ- cal quorum-quenching. Mol. Plant Microbe Interact. 2019, 32, 802–812. [Google Scholar] [CrossRef]
- Vesuna, A.P.; Nerurkar, A.S. Biocontrol impact of AHL degrading actinobacteria on quorum sensing regulated virulence of phytopathogen Pectobacterium carotovorum subsp. carotovorum BR1. Plant Soil 2020, 453, 371–388. [Google Scholar] [CrossRef]
- Li, T.; Mei, Y.; He, B.; Sun, X.; Li, J. Reducing quorum sensing-mediated virulence factor expression and biofilm formation in Hafnia alvei by using the potential quorum sensing inhibitor L-carvone. Front. Microbiol. 2019, 9, 3324. [Google Scholar] [CrossRef] [PubMed]
- Schwab, M.; Bergonzi, C.; Sakkos, J.; Staley, C.; Zhang, Q.; Sadowsky, M.J.; Aksan, A.; Elias, M. Signal disruption leads to changes in bacterial community population. Front. Microbiol. 2019, 10, 611. [Google Scholar] [CrossRef] [PubMed]
- Abarca-Grau, A.M.; Penyalver, R.; López, M.M.; Marco-Noales, E. Pathogenic and non-pathogenic Agrobacterium tumefaciens, A. rhizogenes and A. vitis strains form biofilms on abiotic as well as on root surfaces. Plant Pathol. 2011, 60, 416–425. [Google Scholar] [CrossRef]
- Bosmans, L.; De Bruijn, I.; Gerards, S.; Moerkens, R.; Van Looveren, L.; Wittemans, L.; Van Calenberge, B.; Paeleman, A.; Van Kerckhove, S.; De Mot, R.; et al. Potential for biocontrol of hairy root disease by a Paenibacillus clade. Front. Microbiol. 2017, 8, 447. [Google Scholar] [CrossRef]
- Bosmans, L.; Álvarez-Pérez, S.; Moerkens, R.; Wittemans, L.; Van Calenberge, B.; Kerckhove, S.V.; Paeleman, A.; De Mot, R.; Rediers, H.; Lievens, B. Assessment of the genetic and phenotypic diversity among rhizogenic Agrobacterium biovar 1 strains infecting solanaceous and cucurbit crops. FEMS Microbiol. Ecol. 2015, 91, fiv081. [Google Scholar] [CrossRef] [Green Version]
- Irigoyen, S.; Ramasamy, M.; Pant, S.; Niraula, P.; Bedre, R.; Gurung, M.; Rossi, D.; Laughlin, C.; Gorman, Z.; Achor, D.; et al. Plant hairy roots enable high throughput identification of antimicrobials against Candidatus Liberibacter spp. Nat. Commun. 2020, 11, 5802. [Google Scholar] [CrossRef]
- Ron, M.; Kajala, K.; Pauluzzi, G.; Wang, D.; Reynoso, M.A.; Zumstein, K.; Garcha, J.; Winte, S.; Masson, H.; Inagaki, S.; et al. Hairy root transformation using Agrobacterium rhizogenes as a tool for exploring cell type-specific gene expression and function using tomato as a model. Plant Physiol. 2014, 166, 455–469. [Google Scholar] [CrossRef] [PubMed]
- Ho-Plágaro, T.; Huertas, R.; Tamayo-Navarrete, M.I.; Ocampo, J.A.; Garcia-Garrido, J.M. An improved method for Agrobacterium rhizogenes-mediated transformation of tomato suitable for the study of arbuscular mycorrhizal symbiosis. Plant Methods 2018, 14, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bettini, P.; Baraldi, R.; Rapparini, F.; Melani, L.; Mauro, M.L.; Bindi, D.; Buiatti, M. The insertion of the Agrobacterium rhizogenes rolC gene in tomato (Solanum lycopersicum L.) affects plant architecture and endogenous auxin and abscisic acid levels. Sci. Hortic. 2010, 123, 323–328. [Google Scholar] [CrossRef]
- Sanchez-Contreras, M.; Bauer, W.D.; Gao, M.; Robinson, J.B.; Allan Downie, J. Quorum-sensing regulation in rhizobia and its role in symbiotic interactions with legumes. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2007, 362, 1149–1163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cha, C.; Gao, P.; Chen, Y.C.; Shaw, P.D.; Farrand, S.K. Production of acyl-homoserine lactone quorum-sensing signals by gram-negative plant-associated bacteria. Mol. Plant-Microbe Interact. 1998, 11, 1119–1129. [Google Scholar] [CrossRef] [Green Version]
- González, J.E.; Marketon, M.M. Quorum sensing in nitrogen-fixing rhizobia. Microbiol. Mol. Biol. Rev. 2003, 67, 574–592. [Google Scholar] [CrossRef] [Green Version]
- Wisniewski-Dye, F.; Downie, J.A. Quorum-sensing in Rhizobium. Antonie Van Leeuwenhoek 2002, 81, 397–407. [Google Scholar] [CrossRef] [PubMed]
- Barbey, C.; Chane, A.; Burini, J.-F.; Maillot, O.; Merieau, A.; Gallique, M.; Beury-Cirou, A.; Konto-Ghiorghi, Y.; Feuilloley, M.; Gobert, V.; et al. A rhodococcal transcriptional regulatory mechanism detects the common lactone ring of AHL quorum-sensing signals and triggers the quorum-quenching response. Front. Microbiol. 2018, 9, 2800. [Google Scholar] [CrossRef] [PubMed]
- Cirou, A.; Raffoux, A.; Diallo, S.; Latour, X.; Dessaux, Y.; Faure, D. Gamma-caprolactone stimulates growth of quorum-quench- ing Rhodococcus populations in a large-scale hydroponic system for culturing Solanum tuberosum. Res. Microbiol. 2011, 162, 945–950. [Google Scholar] [CrossRef] [PubMed]
- Cirou, A.; Mondy, S.; An, S.; Charrier, A.; Sarrazin, A.; Thoison, O.; DuBow, M.; Faure, D. Efficient biostimulation of native and introduced quorum-quenching Rhodococcus erythropolis populations is revealed by a combination of analytical chemistry, mi- crobiology, and pyrosequencing. Appl. Environ. Microbiol. 2012, 78, 481–492. [Google Scholar] [CrossRef] [Green Version]
- Heydorn, A.; Nielsen, A.T.; Hentzer, M.; Sternberg, C.; Givskov, M.; Ersbøll, B.K.; Molin, S. Quantification of biofilm structures by the novel computer program COMSTAT. Microbiology 2000, 146, 2395–2407. [Google Scholar] [CrossRef] [Green Version]
- Andersen, J.B.; Heydorn, A.; Hentzer, M.; Eberl, L.; Geisenberger, O.; Christensen, B.B.; Molin, S.; Givskov, M. Gfp-based N-acyl homoserine-lactone sensor systems for detection of bacterial communication. Appl. Environ. Microbiol. 2001, 67, 575–585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Latour, X.; Diallo, S.; Chevalier, S.; Morin, D.; Smadja, B.; Burini, J.-F.; Haras, D.; Orange, N. Thermoregulation of N-acyl homoserine lactone-based quorum sensing in the soft rot bacterium Pectobacterium atrosepticum. Appl. Environ. Microbiol. 2007, 73, 4078–4081. [Google Scholar] [CrossRef] [Green Version]
- Chane, A.; Bourigault, Y.; Bouteiller, M.; Konto-Ghiorghi, Y.; Merieau, A.; Barbey, C.; Latour, X. Close-up on a bacterial informational war in the geocaulosphere. Can. J. Microbiol. 2020, 66, 447–454. [Google Scholar] [CrossRef] [PubMed]
- Uroz, S.; Oger, P.M.; Chapelle, E.; Adeline, M.T.; Faure, D.; Dessaux, Y. A Rhodococcus qsdA-encoded enzyme defines a novel class of large-spectrum quorum-quenching lactonases. Appl. Environ. Microbiol. 2008, 74, 1357–1366. [Google Scholar] [CrossRef] [Green Version]
- Barbey, C.; Crépin, A.; Bergeau, D.; Ouchiha, A.; Mijouin, L.; Taupin, L.; Orange, N.; Feuilloley, M.; Dufour, A.; Burini, J.F.; et al. In Planta biocontrol of Pectobacterium atrosepticum by Rhodococcus erythropolis involves silencing of pathogen communication by the rhodococcal gamma-lactone catabolic pathway. PLoS ONE 2013, 8, e66642. [Google Scholar] [CrossRef]
- Ward, J.P.; King, J.R.; Koerber, A.J.; Croft, J.M.; Sockett, R.E.; Williams, P. Early development and quorum sensing in bacterial biofilms. J. Math. Biol. 2003, 47, 23–55. [Google Scholar] [CrossRef]
- Shrout, J.D.; Chopp, D.L.; Just, C.L.; Hentzer, M.; Givskov, M.; Parsek, M.R. The impact of quorum sensing and swarming motility on Pseudomonas aeruginosa biofilm formation is nutritionally conditional. Mol. Microbiol. 2006, 62, 1264–1277. [Google Scholar] [CrossRef]
- Nadell, C.D.; Xavier, J.B.; Levin, S.A.; Foster, K.R. The evolution of quorum sensing in bacterial biofilms. PLoS Biol. 2008, 6, e14. [Google Scholar] [CrossRef] [Green Version]
- Baltenneck, J.; Reverchon, S.; Hommais, F. Quorum sensing regulation in phytopathogenic bacteria. Microorganisms 2021, 9, 239. [Google Scholar] [CrossRef]
- Charoenpanich, P.; Meyer, S.; Becker, A.; McIntosh, M. Temporal expression program of quorum sensing-based transcription regulation in Sinorhizobium meliloti. J. Bacteriol. 2013, 195, 3224–3236. [Google Scholar] [CrossRef] [Green Version]
- Hoang, H.H.; Gurich, N.; González, J.E. Regulation of motility by the ExpR/Sin quorum-sensing system in Sinorhizobium meliloti. J. Bacteriol. 2008, 190, 861–871. [Google Scholar] [CrossRef] [Green Version]
- Wells, D.H.; Chen, E.J.; Fisher, R.F.; Long, S.R. ExoR is genetically coupled to the ExoS-ChvI two-component system and located in the periplasm of Sinorhizobium meliloti. Mol. Microbiol. 2007, 64, 647–664. [Google Scholar] [CrossRef]
- Russo, D.M.; Williams, A.; Edwards, A.; Posadas, D.M.; Finnie, C.; Dankert, M.; Downie, J.A.; Zorreguieta, A. Proteins exported via the PrsD-PrsE type I secretion system and the acidic exopolysaccharide are involved in biofilm formation by Rhizobium leguminosarum. J. Bacteriol. 2006, 188, 4474–4486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rinaudi, L.V.; González, J.E. The low-molecular-weight fraction of exopolysaccharide II from Sinorhizobium meliloti is a crucial determinant of biofilm formation. J. Bacteriol. 2009, 191, 7216–7224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Byers, J.T.; Lucas, C.; Salmond, G.P.C.; Welch, M. Nonenzymatic turnover of an Erwinia carotovora quorum-sensing signaling molecule. J. Bacteriol. 2002, 184, 1163–1171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wirth, R.; Muscholl, A.; Wanner, G. The role of pheromones in bacterial interactions. Trends Microbiol. 1996, 4, 96–103. [Google Scholar] [CrossRef]
- Blosser-Middleton, R.S.; Gray, K.M. Multiple N-acyl homoserine lactone signals of Rhizobium leguminosarum are synthesized in a distinct temporal pattern. J. Bacteriol. 2001, 183, 6771–6777. [Google Scholar] [CrossRef] [Green Version]
- Velázquez, E.; Peix, A.; Zurdo-Piñeiro, J.L.; Palomo, J.L.; Mateos, P.F.; Rivas, R.; Muñoz-Adelantado, E.; Toro, N.; García-Benavides, P.; Martínez-Molina, E. The coexistence of symbiosis and pathogenicity-determining genes in Rhizobium rhizogenes strains enables them to induce nodules and tumors or hairy roots in plants. Mol. Plant-Microbe Interact. 2005, 18, 1325–1332. [Google Scholar] [CrossRef] [Green Version]
- Velázquez, E.; Palomo, J.L.; Rivas, R.; Guerra, H.; Peix, A.; Trujillo, M.E.; García-Benavides, P.; Mateos, P.F.; Wabiko, H.; Martinez- Molina, E. Analysis of core genes supports the reclassification of strains Agrobacterium radiobacter K84 and Agrobacterium tumefaciens AKE10 into the species Rhizobium rhizogenes. Syst. Appl. Microbiol. 2010, 33, 247–251. [Google Scholar] [CrossRef]
- Rodrigues, S.; Paillard, C.; Le Pennec, G.; Dufour, A.; Bazire, A. Vibrio tapetis, the causative agent of brown ring disease, forms biofilms with spherical components. Front. Microbiol. 2015, 6, 1384. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, S.; Paillard, C.; Van Dillen, S.; Tahrioui, A.; Berjeaud, J.M.; Dufour, A.; Bazire, A. Relation between biofilm and virulence in Vibrio tapetis: A transcriptomic study. Pathogens 2018, 7, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doghri, I.; Rodrigues, S.; Bazire, A.; Dufour, A.; Akbar, D.; Sopena, V.; Sablé, S.; Lanneluc, I. Marine bacteria from the French Atlantic coast displaying high forming-biofilm abilities and different biofilm 3D architectures. BMC Microbiol. 2015, 15, 231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desriac, F.; Clamens, T.; Rosay, T.; Rodrigues, S.; Tahrioui, A.; Enault, J.; Roquigny, L.; Racine, P.J.; Taupin, L.; Bazire, A.; et al. Different Dose-Dependent Modes of Action of C-Type Natriuretic Peptide on Pseudomonas aeruginosa Biofilm Formation. Pathogens 2018, 7, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlafer, S.; Meyer, R.L. Confocal microscopy imaging of the biofilm matrix. J. Microbiol. Methods. 2017, 138, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Chen, Y.P.; Qiu, J.H.; Dai, Y.Z.; Feng, B. Imaging the microprocesses in biofilm matrices. Trends Biotechnol. 2019, 37, 214–226. [Google Scholar] [CrossRef] [PubMed]
- Reichhardt, C.; Parsek, M.R. Confocal laser scanning microscopy for analysis of Pseudomonas aeruginosa biofilm architecture and matrix localization. Front. Microbiol. 2019, 10, 677. [Google Scholar] [CrossRef] [PubMed]
- Charlesworth, J.; Kimyon, O.; Manefield, M.; Burns, B.P. Detection and characterization of N-acyl-l-homoserine lactones using GFP-based biosensors in conjunction with thin-layer chromatography. J. Microbiol. Methods 2015, 118, 164–167. [Google Scholar] [CrossRef]
- Christensen, L.D.; van Gennip, M.; Jakobsen, T.H.; Givskov, M.; Bjarnsholt, T. Imaging N-acyl homoserine lactone quorum sensing in vivo. Methods Mol. Biol. 2011, 692, 147–157. [Google Scholar] [CrossRef]
- Riedel, K.; Hentzer, M.; Geisenberger, O.; Huber, B.; Steidle, A.; Wu, H.; Høiby, N.; Givskov, M.; Molin, S.; Eberl, L. N-acylhomoserine-lactone-mediated communication between Pseudomonas aeruginosa and Burkholderia cepacia in mixed biofilms. Microbiology 2001, 147, 3249–3262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gantner, S.; Schmid, M.; Dürr, C.; Schuhegger, R.; Steidle, A.; Hutzler, P.; Langebartels, C.; Eberl, L.; Hartmann, A.; Dazzo, F.B. In Situ quantitation of the spatial scale of calling distances and population density-independent N-acylhomoserine lactone- mediated communication by rhizobacteria colonized on plant roots. FEMS Microbiol. Ecol. 2006, 56, 188–194. [Google Scholar] [CrossRef] [Green Version]
- Steidle, A.; Sigl, K.; Schuhegger, R.; Ihring, A.; Schmid, M.; Gantner, S.; Stoffels, M.; Riedel, K.; Givskov, M.; Hartmann, A.; et al. Visualization of N-acylhomoserine lactone- mediated cell-cell communication between bacteria colonizing the tomato rhizosphere. Appl. Environ. Microbiol. 2001, 67, 5761–5770. [Google Scholar] [CrossRef] [Green Version]
- Bourigault, Y.; Chane, A.; Barbey, C.; Jafra, S.; Czajkowski, R.; Latour, X. Biosensors used for epifluorescence and confocal laser scanning microscopies to study Dickeya and Pectobacterium virulence and biocontrol. Microorganisms 2021, 9, 295. [Google Scholar] [CrossRef]
- Uroz, S.; Dessaux, Y.; Oger, P. Quorum sensing and quorum quenching: The yin and yang of bacterial communication. ChemBioChem 2009, 10, 205–216. [Google Scholar] [CrossRef] [PubMed]
- Kalia, V.C. Quorum Sensing vs. Quorum Quenching: A Battle with No End in Sight; Kalia, V.C., Ed.; Springer: New Delhi, India, 2015; p. 324. [Google Scholar] [CrossRef]
- Cirou, A.; Diallo, S.; Kurt, C.; Latour, X.; Faure, D. Growth promotion of quorum-quenching bacteria in the rhizosphere of Solanum tuberosum. Environ. Microbiol. 2007, 9, 1511–1522. [Google Scholar] [CrossRef] [PubMed]
- Jafra, S.; Przysowa, J.; Czajkowski, R.; Michta, A.; Garbeva, P.; van der Wolf, J.M. Detection and characterization of bacteria from the potato rhizosphere degrading N-acyl-homoserine lactone. Can. J. Microbiol. 2006, 52, 1006–1015. [Google Scholar] [CrossRef]
- Achari, G.A.; Ramesh, R. Characterization of quorum quenching enzymes from endophytic and rhizosphere colonizing bacteria. Biocat. Agricult. Biotechnol. 2018, 13, 20–24. [Google Scholar] [CrossRef]
- Chane, A.; Barbey, C.; Bourigault, Y.; Beury-Cirou, A.; Bouteiller, M.; Merieau, A.; Konto-Ghiorghi, Y.; Feuilloley, M.; Gobert, V.; Latour, X. Informational war as therapy to silence blackleg and soft-rot diseases. J. Plant Pathol. 2019, 101, 869. [Google Scholar] [CrossRef]
- Singhi, D.; Goyal, A.; Gupta, G.; Yadav, A.; Srivastava, P. Rhodoccoccus erythropolis is different from other members of Actinobacteria: Monoploidy, overlapping replication cycle, and unique segregation pattern. J. Bacteriol. 2019, 201, e00320-19. [Google Scholar] [CrossRef] [PubMed]
- Singhi, D.; Jain, A.; Srivastava, P. Localization of low copy number plasmid pRC4 in replicating rod and non-replicating cocci cells of Rhodococcus erythropolis PR4. PLoS ONE 2016, 11, e0166491. [Google Scholar] [CrossRef] [PubMed]
- Abarca-Grau, A.M.; Burbank, L.P.; de Paz, H.D.; Crespo-Rivas, J.C.; Marco-Noales, E.; López, M.M.; Vinardell, J.M.; von Bodman, S.B.; Penyalver, R. Role for Rhizobium rhizogenes K84 cell envelope polysaccharides in surface interactions. Appl. Environ. Microbiol. 2012, 78, 1644–1651. [Google Scholar] [CrossRef] [Green Version]
- Latour, X.; Barbey, C.; Chane, A.; Groboillot, A.; Burini, J.-F. Rhodococcus erythropolis and its γ-lactone catabolic pathway: An unusual rhodococcal quorum-quenching pathway regulation biocontrol system that disrupts pathogen quorum sensing com- munication. Agronomy 2013, 3, 816–838. [Google Scholar] [CrossRef] [Green Version]
- Chane, A.; Barbey, C.; Bourigault, Y.; Maillot, O.; Rodrigues, S.; Bouteiller, M.; Merieau, A.; Konto-Ghiorghi, Y.; Beury-Cirou, A.; Gattin, R.; et al. A flavor lactone mimicking AHL quorum-sensing signals exploits the broad affinity of the QsdR regulator to stimulate transcription of the rhodococcal qsd operon involved in quorum-quenching and biocontrol activities. Front. Microbiol. 2019, 10, 786. [Google Scholar] [CrossRef] [PubMed]
- Uroz, S.; Chhabra, S.R.; Camara, M.; Williams, P.; Oger, P.; Dessaux, Y. N-acylhomoserine lactone quorum-sensing molecules are modified and degraded by Rhodococcus erythropolis W2 by both amidolytic and novel oxidoreductase activities. Microbiology 2005, 151, 3313–3322. [Google Scholar] [CrossRef] [Green Version]
- Park, S.Y.; Hwang, B.J.; Shin, M.H.; Kim, J.A.; Kim, H.K.; Lee, J.K. N-acylhomoserine lactonase producing Rhodococcus spp. with different AHL- degrading activities. FEMS Microbiol. Lett. 2006, 261, 102–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afriat, L.; Roodveldt, C.; Manco, G.; Tawfik, D.S. The latent promiscuity of newly identified microbial lactonases is linked to a recently diverged phosphotriesterase. Biochemistry 2006, 45, 13677–13686. [Google Scholar] [CrossRef]
- Hiblot, J.; Bzdrenga, J.; Champion, C.; Chabriere, E.; Elias, M. Crystal structure of VmoLac, a tentative quorum quenching lactonase from the extremophilic crenarchaeon Vulcanisaeta moutnovskia. Sci. Rep. 2015, 5, 8372. [Google Scholar] [CrossRef] [Green Version]
- Mersereau, M.; Pazour, G.J.; Das, A. Efficient transformation of Agrobacterium tumefaciens by electroporation. Gene 1990, 90, 149–151. [Google Scholar] [CrossRef]
- Barbey, C.; Crépin, A.; Cirou, A.; Budin-Verneuil, A.; Orange, N.; Feuilloley, M.; Faure, D.; Dessaux, Y.; Burini, J.-F.; Latour, X. Catabolic pathway of gamma-caprolactone in the biocontrol agent Rhodococcus erythropolis. J. Proteome Res. 2012, 11, 206–216. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bourigault, Y.; Rodrigues, S.; Crépin, A.; Chane, A.; Taupin, L.; Bouteiller, M.; Dupont, C.; Merieau, A.; Konto-Ghiorghi, Y.; Boukerb, A.M.; et al. Biocontrol of Biofilm Formation: Jamming of Sessile-Associated Rhizobial Communication by Rhodococcal Quorum-Quenching. Int. J. Mol. Sci. 2021, 22, 8241. https://doi.org/10.3390/ijms22158241
Bourigault Y, Rodrigues S, Crépin A, Chane A, Taupin L, Bouteiller M, Dupont C, Merieau A, Konto-Ghiorghi Y, Boukerb AM, et al. Biocontrol of Biofilm Formation: Jamming of Sessile-Associated Rhizobial Communication by Rhodococcal Quorum-Quenching. International Journal of Molecular Sciences. 2021; 22(15):8241. https://doi.org/10.3390/ijms22158241
Chicago/Turabian StyleBourigault, Yvann, Sophie Rodrigues, Alexandre Crépin, Andrea Chane, Laure Taupin, Mathilde Bouteiller, Charly Dupont, Annabelle Merieau, Yoan Konto-Ghiorghi, Amine M. Boukerb, and et al. 2021. "Biocontrol of Biofilm Formation: Jamming of Sessile-Associated Rhizobial Communication by Rhodococcal Quorum-Quenching" International Journal of Molecular Sciences 22, no. 15: 8241. https://doi.org/10.3390/ijms22158241
APA StyleBourigault, Y., Rodrigues, S., Crépin, A., Chane, A., Taupin, L., Bouteiller, M., Dupont, C., Merieau, A., Konto-Ghiorghi, Y., Boukerb, A. M., Turner, M., Hamon, C., Dufour, A., Barbey, C., & Latour, X. (2021). Biocontrol of Biofilm Formation: Jamming of Sessile-Associated Rhizobial Communication by Rhodococcal Quorum-Quenching. International Journal of Molecular Sciences, 22(15), 8241. https://doi.org/10.3390/ijms22158241