
Unraveling Axon Guidance during Axotomy and Regeneration


Abstract
:1. Introduction
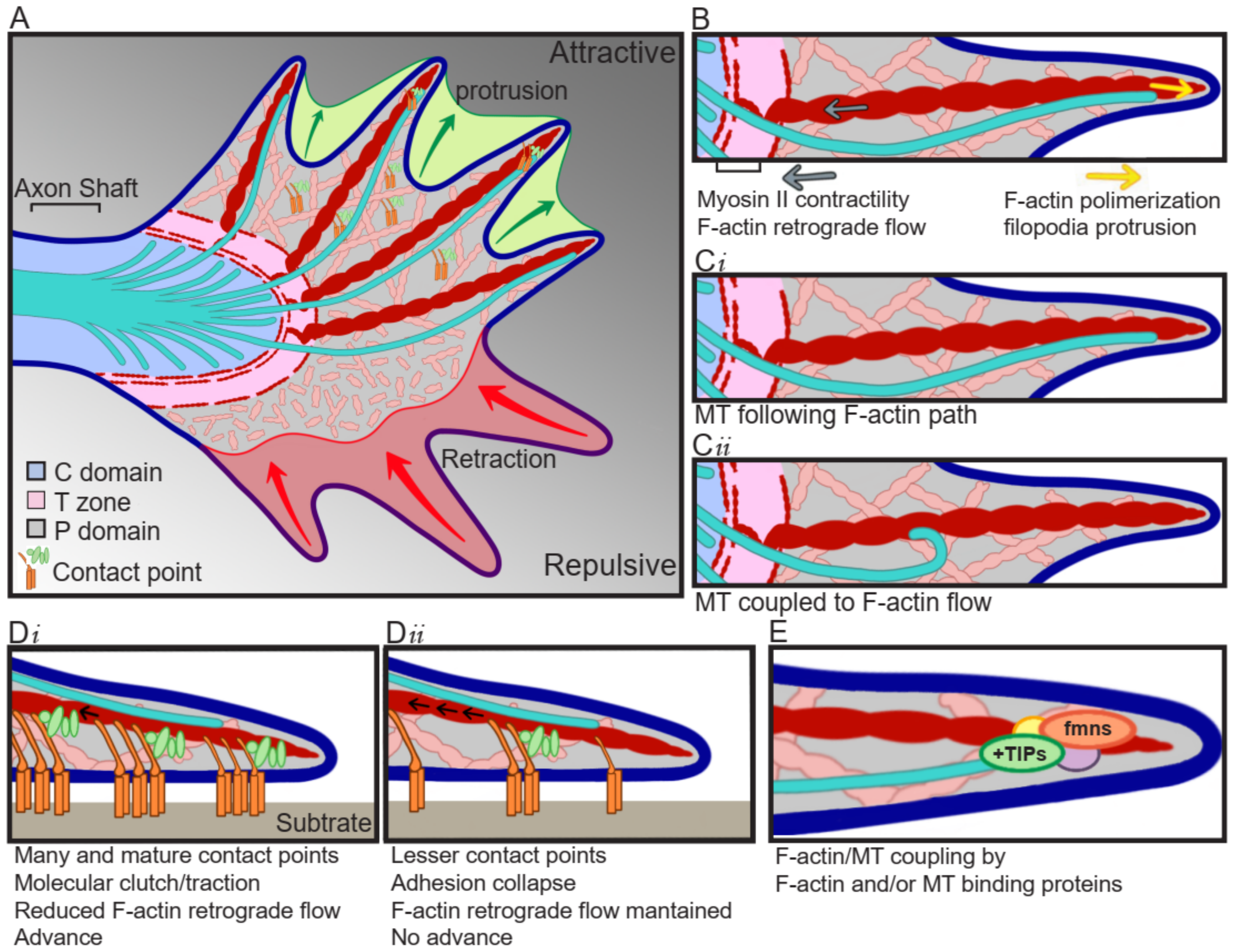
2. Growth Cone: Leading the Way
3. F-Actin and MT Cytoskeleton and Crosstalk
3.1. Actin Cytoskeleton
3.2. MT Cytoskeleton
3.3. Actin/MT Crosstalk
4. Canonical Guidance Cues: What Do We Know?
5. Guidance Cues in Concert
5.1. Guidance Cues in Embryonic Versus Adult Nervous System
5.2. Guidance Cues and Spinal Cord Development
5.3. Guidance Cues and Cytoskeleton Regulation
6. Guidance Cues in Axotomy and Axon Regeneration
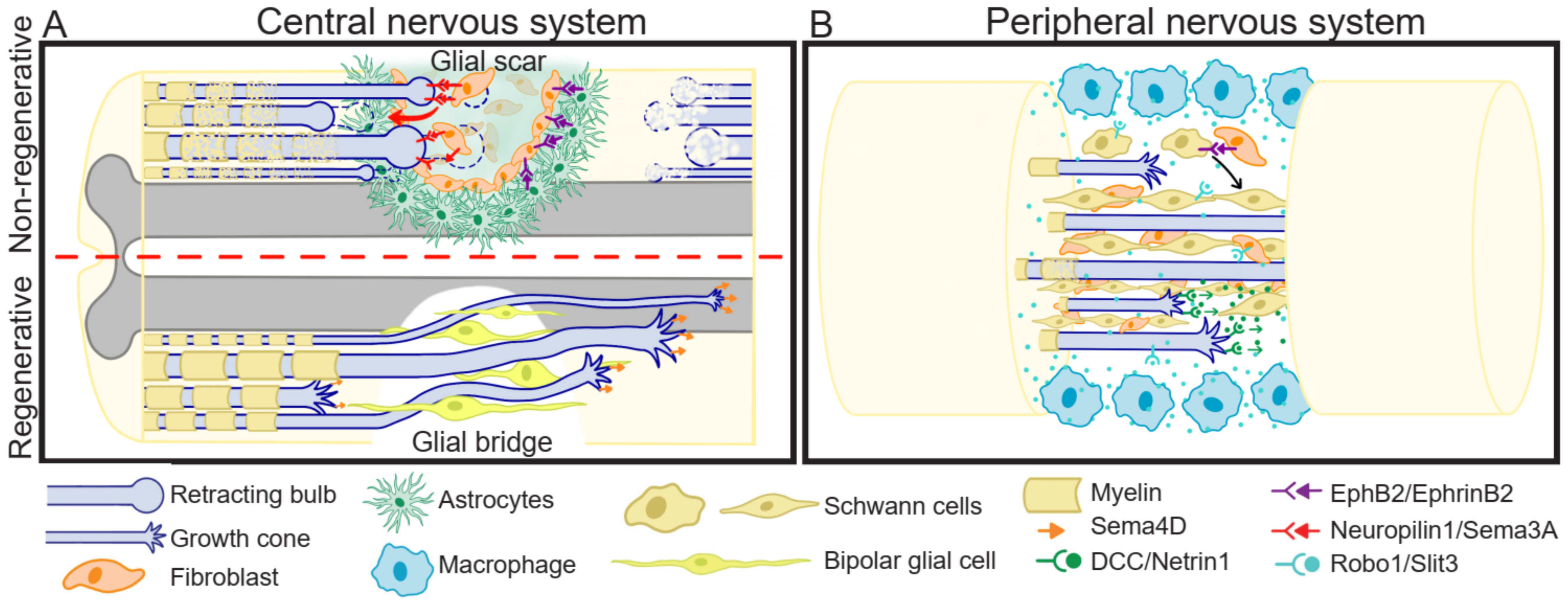
6.1. Spinal Cord Injury
6.1.1. Growth Cone Formation
6.1.2. Immune Response: Pro- and Anti-Inflammatory Environment
6.1.3. Glial Scar
6.1.4. Glial Bridge
6.2. Differential Expression of Guidance Cues after Nervous System Damage
6.2.1. Differential Expression of Guidance Cues after Different SCI Paradigms
6.2.2. Differential Expression of Guidance Cues after Different PNS Injury Paradigms
6.2.3. Differential Expression of Guidance Cues after Nervous System Injuries in Regenerative Animal Models
6.2.4. Differential Expression of Guidance Cues Detected by Functional Studies

{kind=link}
{kind=link}
| Guidance Cues | Injury | Expression Levels | mRNA/Protein | Comments | Ref. |
|---|---|---|---|---|---|
| Sema3A | Contusion, transection and motoneurons axotomy, in rats | Upregulated | mRNA/protein | In fibroblast and motoneurons in the lesion center | [149,151,173,174] |
| Sema3F | Xenopus spinal cord transection | Upregulated | mRNA | Caudal portion of NR stages spinal cord | [69] |
| Sema4F/ Neuropilin2 | Rat intraspinal motoneurons axotomy | Upregulated | mRNA | In affected motoneurons | [174] |
| Sema4B/ PlexinA3 | Xenopus spinal cord transection | Downregulated | mRNA | Caudal portion of NR stages spinal cord | [69] |
| Netrin-1 | Mouse dorsal hemisection, rat sagittal myelotomy, and rat spinal cord punch | Upregulated/ Downregulated | mRNA/protein | Upregulation in activated microglia and macrophages in the lesion epicenter. Detected in neurons and oligodendrocytes from sparse tissue. | [160,177,178] |
| DCC | Rat sagittal myelotomy and Xenopus spinal cord transection | Upregulated/ Downregulated | mRNA/protein | Early and transient upregulation, followed by downregulation in lesion epicenter | [69,177] |
| UNC5 | Rat sagittal myelotomy, spinal cord dorsal hemisection, and transection | Downregulated | mRNA/protein | Downregulation of UNC5A-D in neurons and oligodendrocytes | [176,177,179] |
| Slit-1/3 | Mouse dorsal hemisection | Upregulated | mRNA | In macrophages and fibroblasts in the lesion epicenter | [160] |
| Slit-2 | Rat spinal cord punch and contusion | Upregulated | mRNA/protein | Early and transient upregulation in neurons | [178,180] |
| Robo-1 | Rat spinal cord contusion | Upregulated | mRNA/protein | In surviving neurons | [181] |
| Ephrin-B2 | Thoracic spinal cord transection, and dorsal hemisection in rat | Upregulated | mRNA/protein | Mainly in reactive astrocytes in the glial scar. One study shows an early and transient downregulation | [134,154,182] |
| EphrinB3 | Rat spinal cord crush and dorsal hemisection | Down- and upregulated | mRNA/protein | Downregulation is most probably due to cell death. And upregulation in astrocytes | [134,182] |
| EphrinB1 | Rat spinal cord dorsal hemisection | Upregulated | Protein | In astrocytes | [134] |
| EphrinA1 | Rat spinal cord contusion | Downregulated/ Upregulated | mRNA/protein | Expressed in reactive astrocytes and neurons | [183] |
| EphA4 | Rat spinal cord dorsal hemisection | Upregulated | mRNA/protein | In axons rostrally, and astrocytes in the injury site | [134,182] |
| Rat spinal cord contusion | Downregulated/ Upregulated | mRNA/protein | In neurons caudal to injury and astrocytes rostral and caudal to the lesion epicenter | [186,212] | |
| EphB3 | Rat spinal cord contusion and transection | Upregulated | mRNA/protein | In astrocytes in the epicenter and in neurons rostral and caudal to the injury site | [184,185] |
| EphB2 | Spinal cord transection and contusion | Downregulation/ Upregulation | mRNA/protein | Expressed in activated astrocytes and in fibroblasts invading the lesion site | [154,155] |
| Guidance Cues | Injury | Expression Levels | mRNA/Protein | Comments | Ref. |
|---|---|---|---|---|---|
| Sema3A | Rat sciatic nerve transection and crush, and femoral nerve transection | Upregulated | mRNA | In ipsilateral spinal cord motor neurons | [187,188] |
| Rat dorsal root transection | Downregulated | mRNA | In DRG neurons | [189] | |
| Sema3F | Rat sciatic nerve transection, crush, and contusion | Upregulated | mRNA | In epineurial fibroblast and perineurium | [189,190] |
| Rat DRG transection | Downregulated | mRNA | In neurons | [189] | |
| Neuropilin-1/2 | Rat DRG and sciatic nerve transection | Upregulated | mRNA | DRG neurons | [189] |
| Neuropilin-2 | Rat sciatic nerve transection | Upregulated | mRNA/protein | In neuron caudal to injury, and in Schwann cells in the perineurium and epineurium | [190] |
| Netrin-1 | Rat sciatic nerve transection and crush, and experimental autoimmune neuritis | Upregulated | mRNA/protein | In Schwann cells, macrophages, ECM, and some axons | [191,192,193] |
| DCC | Rat sciatic nerve crush and transection | Upregulated | mRNA/protein | In DRG neurons, and Schwann cells | [193,194] |
| UNC5B | Rat sciatic nerve, and mice median nerve transection | Downregulated/ Upregulated | mRNA/protein | Downregulation in DRG neurons, followed by an increase in the distal nerve segment | [194,195] |
| EphA4 | Rat sciatic nerve crush | Upregulated/ Downregulated | protein | In Schwann cells at the injury site | [164] |
| Slit1 | Rat and mouse sciatic nerve transection and rat sciatic nerve crush | Down- and upregulated | mRNA/protein | Downregulated in mouse DRG neurons, and upregulated in rat DRG neurons and non-neuronal cells | [165,197,198] |
| Slit2 | Rat and mouse sciatic nerve transection | Down- and upregulated | mRNA | Downregulated in DRG neurons and in the injury site, and upregulated in Schwann cells and proximal stump | [165,196] |
| Slit3 | Mouse sciatic nerve transection | Down- and upregulated | mRNA | Downregulated in neurons and upregulated in macrophages surrounding the nerve bridge | [195] |
| Robo1 | Mouse and rat sciatic nerve transection | Downregulated/ Upregulated | mRNA | Early downregulation in DRG neurons, and upregulation in Schwann cells in the distal stump and in the nerve bridge | [165,196] |
| Robo 2 | Mouse and rat sciatic nerve transection | Down- and upregulated | mRNA/protein | In mouse a transient downregulation and in rats a transient upregulation | [165,196,197] |
| srGAP1/3 | Mouse sciatic nerve transection | Upregulated | mRNA/protein | In the ipsilateral of DRG neurons | [199] |
| Glypican-1 | Rat sciatic nerve transection | Upregulated | mRNA/protein | In the ipsilateral of DRG neurons and in neighboring non-neuronal cells | [200] |
| Guidance Cues | Injury | Expression Levels | mRNA/Protein | Comments | Ref. |
|---|---|---|---|---|---|
| Sema3 | Larval lampreys spinal cord transection | Upregulated | mRNA | In microglia/macrophages and in some neurons near the lesion site | [201] |
| Sema4 | Larval lampreys spinal cord transection | Downregulated | mRNA | Downregulated in dorsal cells, and present rostral and caudal to injury, but absent from scar | [201] |
| Sema4B/ PlexinA3 | Xenopus spinal cord transection | Upregulated | mRNA | The caudal portion of R stages spinal cord | [69] |
| Netrin | Larval lampreys spinal cord transection | Downregulated | mRNA | In neurons close to the injury site | [201] |
| DCC | Xenopus spinal cord transection | Upregulated | mRNA | The caudal portion of R stages spinal cord | [69] |
| UNC-5 | Larval lampreys spinal cord transection | Downregulated/ Upregulated | mRNA | Upregulation in neurons with poor regenerative capacity, and absent from the ones with good capacity | [202] |
| Ephrin | Xenopus spinal cord transection | Upregulated | mRNA | The caudal portion of R stages spinal cord | [69] |
| Slit1 | Zebrafish peripheral motor nerves transection | Upregulated | mRNA | Ventral and ventrolateral to the injury | [168] |
7. Treatments: Guidance Cues Regulation
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
List of Abbreviations
| ADF | Actin depolymerizing factor |
| APC | Adenomatous polyposis coli |
| Arp2/3 | Actin-related protein 2/3 |
| BBB | Blood brain barrier |
| BDNF | Brain-derived neurotrophic factor |
| C domain | Central domain |
| CLASP | Cytoplasmic linker-associated protein |
| CLIP-170 | Cytoplasmic linker protein 170 |
| CM | Corticomotoneural |
| CNS | Central nervous system |
| CRMP | Collapsin response mediator protein |
| CSPG | Chondroitin sulfate proteoglycans |
| DAAM | Disheveled-associated activator of morphogenesis |
| DCC | Deleted in colorectal cancer |
| DCX | Doublecortin |
| Dpi | Days post-injury |
| DRG | Dorsal root ganglia |
| DSCAM | Down syndrome cell adhesion molecule |
| EAE | Experimental autoimmune enchephalomyelitis |
| EB1 | End-binding protein 1 |
| ECM | Extracellular matrix |
| Ena/VASP | Enabled/vasodilator-stimulated phosphoprotein |
| FAK | Focal adhesion kinase |
| FLRT3 | Fibronectin leucine rich transmembrane protein 3 |
| Fmn | Formin |
| GFAP | Glial fibrillary acidic protein |
| IFNγ | Interferon gamma |
| INF | N-terminally truncated Drf-like inverted formin |
| KD | Knock down |
| LMC | Lateral motor column |
| L1-CAM | L1 cell adhesion molecule |
| LPS | Lipopolysaccharide |
| MAP | Microtubules associated protein |
| MICAL | Molecule interacting with CasL |
| MMC | Medial motor column |
| MT | Microtubule |
| NAV1 | Navigator-1 |
| NGF | Nerve growth factor |
| NO | Nitric oxide |
| NR | Non-regenerative |
| P domain | Peripheral domain |
| PNS | Peripheral nevous system |
| +TIP | Plus end binding protein |
| R | Regenerative |
| RGC | Retinal ganglion cells |
| RGM | Repulsive guidance molecule |
| RTKs | Receptor tyrosine-kinase |
| Robo | Roundabout |
| ROCK | RhoA target Rho kinase |
| SCI | Spinal cord injury |
| Sema | Semaphorin |
| SlitC | Slit C-terminal fragment |
| SlitN | Slit N-terminal fragment |
| srGAP | Slit-Robo GAP |
| T zone | Transition zone |
| TACC3 | Transforming acidic coiled-coil protein 3 |
| TBI | Traumatic brain injury |
| TCA | Thalamocortical área |
| TG | Trigeminal ganglia |
| TGF-ß1 | Transforming growth factor ß1 |
| TNF | Tumor necrosis factor |
| UNC5 | Uncoordinated-5 |
| XMAP215 | Xenopus microtubule associated protein 215 |
References
- Schaefer, A.W.; Kabir, N.; Forscher, P. Filopodia and actin arcs guide the assembly and transport of two populations of microtubules with unique dynamic parameters in neuronal growth cones. J. Cell Biol. 2002, 158, 139–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medeiros, N.A.; Burnette, D.T.; Forscher, P. Myosin II functions in actin-bundle turnover in neuronal growth cones. Nat. Cell Biol. 2006, 8, 216–226. [Google Scholar] [CrossRef] [PubMed]
- Burnette, D.T.; Schaefer, A.W.; Ji, L.; Danuser, G.; Forscher, P. Filopodial actin bundles are not necessary for microtubule advance into the peripheral domain of Aplysia neuronal growth cones. Nat. Cell Biol. 2007, 9, 1360–1369. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, D.J.; Burmeister, D.W. Stages in axon formation: Observations of growth of Aplysia axons in culture using video-enhanced contrast-differential interference contrast microscopy. J. Cell Biol. 1986, 103, 1921–1931. [Google Scholar] [CrossRef] [Green Version]
- Nichol, R.H.; Hagen, K.M.; Lumbard, D.C.; Dent, E.W.; Gómez, T.M. Guidance of axons by local coupling of retrograde flow to point contact adhesions. J. Neurosci. 2016, 36, 2267–2282. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.H.; Forscher, P. Growth cone advance is inversely proportional to retrograde F-actin flow. Neuron 1995, 14, 763–771. [Google Scholar] [CrossRef] [Green Version]
- Schaefer, A.W.; Schoonderwoert, V.T.G.; Ji, L.; Mederios, N.; Danuser, G.; Forscher, P. Coordination of Actin Filament and Microtubule Dynamics during Neurite Outgrowth. Dev. Cell 2008, 15, 146–162. [Google Scholar] [CrossRef] [Green Version]
- Gomez, T.M.; Letourneau, P.C. Actin Dynamics in Growth Cone Motility and Navigation. J. Neurochem. 2014, 129, 221–234. [Google Scholar] [CrossRef]
- Lowery, L.A.; Van Vactor, D. The trip of the tip: Understanding the growth cone machinery. Nat. Rev. Mol. Cell Biol. 2009, 10, 332–343. [Google Scholar] [CrossRef]
- Lee, A.C.; Suter, D.M. Quantitative analysis of microtubule dynamics during adhesion-mediated growth cone guidance. Dev. Neurobiol. 2008, 68, 1363–1377. [Google Scholar] [CrossRef] [Green Version]
- Suter, D.M.; Schaefer, A.W.; Forscher, P. Microtubule Dynamics Are Necessary for Src Family Kinase-Dependent Growth Cone Steering. Curr. Biol. 2004, 14, 1194–1199. [Google Scholar] [CrossRef] [Green Version]
- Suter, D.M.; Forscher, P. Transmission of growth cone traction force through apCAM-cytoskeletal linkages is regulated by Src family tyrosine kinase activity. J. Cell Biol. 2001, 155, 427–438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robles, E.; Gomez, T.M. Focal adhesion kinase signaling at sites of integrin-mediated adhesion controls axon pathfinding. Nat. Neurosci. 2006, 9, 1274–1283. [Google Scholar] [CrossRef] [PubMed]
- Gomez, T.M.; Roche, F.K.; Letourneau, P.C. Chick sensory neuronal growth cones distinguish fibronectin from laminin by making substratum contacts that resemble focal contacts. J. Neurobiol. 1996, 29, 18–34. [Google Scholar] [CrossRef]
- Woo, S.; Gomez, T.M. Rac1 and RhoA promote neurite outgrowth through formation and stabilization of growth cone point contacts. J. Neurosci. 2006, 26, 1418–1428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chighizola, M.; Dini, T.; Lenardi, C.; Milani, P.; Podestà, A.; Schulte, C. Mechanotransduction in neuronal cell development and functioning. Biophys. Rev. 2019, 11, 701–720. [Google Scholar] [CrossRef] [PubMed]
- De Vincentiis, S.; Falconieri, A.; Scribano, V.; Ghignoli, S.; Raffa, V. Manipulation of axonal outgrowth via exogenous low forces. Int. J. Mol. Sci. 2020, 21, 8009. [Google Scholar] [CrossRef]
- Koser, D.E.; Thompson, A.J.; Foster, S.K.; Dwivedy, A.; Pillai, E.K.; Sheridan, G.K.; Svoboda, H.; Viana, M.; Costa, L.D.F.; Guck, J.; et al. Mechanosensing is critical for axon growth in the developing brain. Nat. Neurosci. 2016, 19, 1592–1598. [Google Scholar] [CrossRef] [Green Version]
- Green, H.J.; Brown, N.H. Integrin intracellular machinery in action. Exp. Cell Res. 2019, 378, 226–231. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Costell, M.; Fässler, R. Integrin activation by talin, kindlin and mechanical forces. Nat. Cell Biol. 2019, 21, 25–31. [Google Scholar] [CrossRef]
- Myers, J.P.; Gomez, T.M. Focal adhesion kinase promotes integrin adhesion dynamics necessary for chemotropic turning of nerve growth cones. J. Neurosci. 2011, 31, 13585–13595. [Google Scholar] [CrossRef] [Green Version]
- Kalil, K.; Szebenyi, G.; Dent, E.W. Common mechanisms underlying growth cone guidance and axon branching. J. Neurobiol. 2000, 44, 145–158. [Google Scholar] [CrossRef]
- Tanaka, E.; Ho, T.; Kirschner, M.W. The role of microtubule dynamics in growth cone motility and axonal growth. J. Cell Biol. 1995, 128, 139–155. [Google Scholar] [CrossRef] [PubMed]
- Szebenyi, G.; Callaway, J.L.; Dent, E.W.; Kalil, K. Interstitial branches develop from active regions of the axon demarcated by the primary growth cone during pausing behaviors. J. Neurosci. 1998, 18, 7930–7940. [Google Scholar] [CrossRef] [PubMed]
- Omotade, O.F.; Pollitt, S.L.; Zheng, J.Q. Actin-based growth cone motility and guidance. Mol. Cell. Neurosci. 2017, 84, 4–10. [Google Scholar] [CrossRef] [PubMed]
- Dent, E.W.; Gupton, S.L.; Gertler, F.B. The growth cone cytoskeleton in Axon outgrowth and guidance. Cold Spring Harb. Perspect. Biol. 2011, 3, a001800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, G.; Dwyer, T. Microtubule dynamics in axon guidance. Neurosci. Bull. 2014, 30, 569–583. [Google Scholar] [CrossRef] [PubMed]
- Biswas, S.; Kalil, K. The microtubule-associated protein tau mediates the organization of microtubules and their dynamic exploration of actin-rich lamellipodia and filopodia of cortical growth cones. J. Neurosci. 2018, 38, 291–307. [Google Scholar] [CrossRef]
- Del Río, J.A.; González-Billault, C.; Ureña, J.M.; Jiménez, E.M.; Barallobre, M.J.; Pascual, M.; Pujadas, L.; Simó, S.; La Torre, A.; Wandosell, F.; et al. MAP1B Is Required for Netrin 1 Signaling in Neuronal Migration and Axonal Guidance. Curr. Biol. 2004, 14, 840–850. [Google Scholar] [CrossRef] [Green Version]
- Bearce, E.A.; Erdogan, B.; Lowery, L.A. TIPsy tour guides: How microtubule plus-end tracking proteins (+TIPS) facilitate axon guidance. Front. Cell. Neurosci. 2015, 9, 241. [Google Scholar] [CrossRef] [Green Version]
- Koester, M.P.; Müller, O.; Pollerberg, G.E. Adenomatous polyposis coli is differentially distributed in growth cones and modulates their steering. J. Neurosci. 2007, 27, 12590–12600. [Google Scholar] [CrossRef] [Green Version]
- Shintani, T.; Ihara, M.; Tani, S.; Sakuraba, J.; Sakuta, H.; Noda, M. APC2 plays an essential role in axonal projections through the regulation of microtubule stability. J. Neurosci. 2009, 29, 11628–11640. [Google Scholar] [CrossRef]
- Lee, H.; Engel, U.; Rusch, J.; Scherrer, S.; Sheard, K.; Van Vactor, D. The microtubule plus end tracking protein orbit/MAST/CLASP acts downstream of the tyrosine kinase Abl in mediating axon guidance. Neuron 2004, 42, 913–926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engel, U.; Zhan, Y.; Long, J.B.; Boyle, S.N.; Ballif, B.A.; Dorey, K.; Gygi, S.P.; Koleske, A.J.; VanVactor, D. Abelson phosphorylation of CLASP2 modulates its association with microtubules and actin. Cytoskeleton 2014, 71, 195–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erdogan, B.; Cammarata, G.M.; Lee, E.J.; Pratt, B.C.; Francl, A.F.; Rutherford, E.L.; Lowery, L.A. The microtubule plus-end-tracking protein TACC3 promotes persistent axon outgrowth and mediates responses to axon guidance signals during development. Neural Dev. 2017, 12, 3. [Google Scholar] [CrossRef] [Green Version]
- Slater, P.G.; Cammarata, G.M.; Samuelson, A.G.; Magee, A.; Hu, Y.; Lowery, L.A. XMAP215 promotes microtubule–F-actin interactions to regulate growth cone microtubules during axon guidance in Xenopus laevis. J. Cell Sci. 2019, 132, jcs224311. [Google Scholar] [CrossRef] [Green Version]
- Erdogan, B.; St. Clair, R.M.; Cammarata, G.M.; Zaccaro, T.; Ballif, B.A.; Lowery, L.A. Investigating the impact of the phosphorylation status of tyrosine residues within the TACC domain of TACC3 on microtubule behavior during axon growth and guidance. Cytoskeleton 2020, 77, 277–291. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Huertas, C.; Bonhomme, M.; Falco, A.; Fagotto-Kaufmann, C.; Van Haren, J.; Jeanneteau, F.; Galjart, N.; Debant, A.; Boudeau, J. The +TIP Navigator-1 is an actin-microtubule crosslinker that regulates axonal growth cone motility. J. Cell Biol. 2020, 219, e201905199. [Google Scholar] [CrossRef] [PubMed]
- Coles, C.H.; Bradke, F. Coordinating Neuronal Actin-Microtubule Dynamics. Curr. Biol. 2015, 25, R677–R691. [Google Scholar] [CrossRef] [Green Version]
- Vitriol, E.A.; Zheng, J.Q. Growth Cone Travel in Space and Time: The Cellular Ensemble of Cytoskeleton, Adhesion, and Membrane. Neuron 2012, 73, 1068–1081. [Google Scholar] [CrossRef] [Green Version]
- Sahasrabudhe, A.; Ghate, K.; Mutalik, S.; Jacob, A.; Ghose, A. Formin 2 regulates the stabilization of filopodial tip adhesions in growth cones and affects neuronal outgrowth and pathfinding in vivo. Development 2016, 143, 449–460. [Google Scholar] [CrossRef] [Green Version]
- Ghate, K.; Mutalik, S.P.; Sthanam, L.K.; Sen, S.; Ghose, A. Fmn2 Regulates Growth Cone Motility by Mediating a Molecular Clutch to Generate Traction Forces. Neuroscience 2020, 448, 160–171. [Google Scholar] [CrossRef] [PubMed]
- Kundu, T.; Dutta, P.; Nagar, D.; Maiti, S.; Ghose, A. Coupling of dynamic microtubules to F-actin by Fmn2 regulates chemotaxis of neuronal growth cones. J. Cell Sci. 2021, 134, jcs252916. [Google Scholar] [CrossRef]
- Szikora, S.; Földi, I.; Tóth, K.; Migh, E.; Vig, A.; Bugyi, B.; Maléth, J.; Hegyi, P.; Kaltenecker, P.; Sanchez-Soriano, N.; et al. The formin DAAM is required for coordination of the actin and microtubule cytoskeleton in axonal growth cones. J. Cell Sci. 2017, 130, 2506–2519. [Google Scholar] [CrossRef] [Green Version]
- Cammarata, G.M.; Bearce, E.A.; Lowery, L.A. Cytoskeletal social networking in the growth cone: How +TIPs mediate microtubule-actin cross-linking to drive axon outgrowth and guidance. Cytoskeleton 2016, 73, 461–476. [Google Scholar] [CrossRef] [Green Version]
- Henty-Ridilla, J.L.; Rankova, A.; Eskin, J.A.; Kenny, K.; Goode, B.L. Accelerated actin filament polymerization from microtubule plus ends. Science 2016, 352, 1004–1010. [Google Scholar] [CrossRef] [Green Version]
- Lowery, L.A.; Stout, A.; Faris, A.E.; Ding, L.; Baird, M.A.; Davidson, M.W.; Danuser, G.; Van Vactor, D. Growth cone-specific functions of XMAP215 in restricting microtubule dynamics and promoting axonal outgrowth. Neural Dev. 2013, 8, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, B.L. Semaphorin 3A: From growth cone repellent to promoter of neuronal regeneration. Neural Regen. Res. 2018, 13, 795–796. [Google Scholar] [CrossRef]
- Sharma, A.; Verhaagen, J.; Harvey, A.R. Receptor complexes for each of the class 3 Semaphorins. Front. Cell. Neurosci. 2012, 6, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alto, L.T.; Terman, J.R. Semaphorins and their signaling mechanisms. Methods Mol. Biol. 2017, 1493, 1–25. [Google Scholar] [CrossRef] [Green Version]
- Dun, X.P.; Parkinson, D.B. Role of Netrin-1 signaling in nerve regeneration. Int. J. Mol. Sci. 2017, 18, 491. [Google Scholar] [CrossRef] [Green Version]
- Boyer, N.P.; Gupton, S.L. Revisiting netrin-1: One who guides (Axons). Front. Cell. Neurosci. 2018, 12, 221. [Google Scholar] [CrossRef]
- Xu, K.; Wu, Z.; Renier, N.; Antipenko, A.; Tzvetkova-Robev, D.; Xu, Y.; Minchenko, M.; Nardi-Dei, V.; Rajashankar, K.R.; Himanen, J.; et al. Structures of netrin-1 bound to two receptors provide insight into its axon guidance mechanism. Science 2014, 344, 1275–1279. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.; Shao, Q.; Qu, C.; Yang, T.; Dwyer, T.; Liu, G. Coordinated interaction of Down syndrome cell adhesion molecule and deleted in colorectal cancer with dynamic TUBB3 mediates Netrin-1-induced axon branching. Neuroscience 2015, 293, 109–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finci, L.I.; Krüger, N.; Sun, X.; Zhang, J.; Chegkazi, M.; Wu, Y.; Schenk, G.; Mertens, H.D.T.; Svergun, D.I.; Zhang, Y.; et al. The Crystal Structure of Netrin-1 in Complex with DCC Reveals the Bifunctionality of Netrin-1 As a Guidance Cue. Neuron 2014, 83, 839–849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Purohit, A.A.; Li, W.; Qu, C.; Dwyer, T.; Shao, Q.; Guan, K.L.; Liu, G. Down syndrome cell adhesion molecule (DSCAM) associates with uncoordinated-5C (UNC5C) in netrin-1-mediated growth cone collapse. J. Biol. Chem. 2012, 287, 27126–27138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qu, C.; Dwyer, T.; Shao, Q.; Yang, T.; Huang, H.; Liu, G. Direct binding of TUBB3 with DCC couples netrin-1 signaling to intracellular microtubule dynamics in axon outgrowth and guidance. J. Cell Sci. 2013, 126, 3070–3081. [Google Scholar] [CrossRef] [Green Version]
- Shao, Q.; Yang, T.; Huang, H.; Alarmanazi, F.; Liu, G. Uncoupling of UNC5C with polymerized TUBB3 in microtubules mediates netrin-1 repulsion. J. Neurosci. 2017, 37, 5620–5633. [Google Scholar] [CrossRef] [Green Version]
- Taylor, A.M.; Menon, S.; Gupton, S.L. Passive microfluidic chamber for long-term imaging of axon guidance in response to soluble gradients. Lab Chip 2015, 15, 2781–2789. [Google Scholar] [CrossRef] [Green Version]
- Baba, K.; Yoshida, W.; Toriyama, M.; Shimada, T.; Manning, C.F.; Saito, M.; Kohno, K.; Trimmer, J.S.; Watanabe, R.; Inagaki, N. Gradient-reading and mechano-effector machinery for netrin-1-induced axon guidance. eLife 2018, 7, e34593. [Google Scholar] [CrossRef]
- Kubo, Y.; Baba, K.; Toriyama, M.; Minegishi, T.; Sugiura, T.; Kozawa, S.; Ikeda, K.; Inagaki, N. Shootin1-cortactin interaction mediates signal-force transduction for axon outgrowth. J. Cell Biol. 2015, 210, 663–676. [Google Scholar] [CrossRef] [Green Version]
- Tong, M.; Jun, T.; Nie, Y.; Hao, J.; Fan, D. The role of the SLIT/Robo signaling pathway. J. Cancer 2019, 10, 2694–2705. [Google Scholar] [CrossRef] [Green Version]
- Leyva-Díaz, E.; Del Toro, D.; Menal, M.J.; Cambray, S.; Susín, R.; Tessier-Lavigne, M.; Klein, R.; Egea, J.; López-Bendito, G. FLRT3 Is a Robo1-interacting protein that determines netrin-1 attraction in developing axons. Curr. Biol. 2014, 24, 494–508. [Google Scholar] [CrossRef] [Green Version]
- Taylor, H.; Campbell, J.; Nobes, C.D. Ephs and ephrins. Curr. Biol. 2017, 27, R90–R95. [Google Scholar] [CrossRef] [Green Version]
- Robinson, R.A.; Griffiths, S.C.; van de Haar, L.L.; Malinauskas, T.; van Battum, E.Y.; Zelina, P.; Schwab, R.A.; Karia, D.; Malinauskaite, L.; Brignani, S.; et al. Simultaneous binding of Guidance Cues NET1 and RGM blocks extracellular NEO1 signaling. Cell 2021, 184, 2103–2120.e31. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zhou, Q.; Luo, Y.; Nguyen, T.; Rosenblatt, M.I.; Guaiquil, V.H. Semaphorin3A induces nerve regeneration in the adult cornea-A switch from its repulsive role in development. PLoS ONE 2018, 13, e0191962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bott, C.J.; Johnson, C.G.; Yap, C.C.; Dwyer, N.D.; Litwa, K.A.; Winckler, B. Nestin in immature embryonic neurons affects axon growth cone morphology and Semaphorin3a sensitivity. Mol. Biol. Cell 2019, 30, 1214–1229. [Google Scholar] [CrossRef]
- Bott, C.J.; McMahon, L.P.; Keil, J.M.; Choo Yap, C.; Kwan, K.Y.; Winckler, B. Nestin Selectively Facilitates the Phosphorylation of the Lissencephaly-Linked Protein Doublecortin (DCX) by cdk5/p35 to Regulate Growth Cone Morphology and Sema3a Sensitivity in Developing Neurons. J. Neurosci. 2020, 40, 3720–3740. [Google Scholar] [CrossRef] [PubMed]
- Lee-Liu, D.; Moreno, M.; Almonacid, L.I.; Tapia, V.S.; Muñoz, R.; von Marées, J.; Gaete, M.; Melo, F.; Larraín, J. Genome-wide expression profile of the response to spinal cord injury in Xenopus laevis reveals extensive differences between regenerative and non-regenerative stages. Neural Dev. 2014, 9, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Edwards-Faret, G.; González-Pinto, K.; Cebrián-Silla, A.; Peñailillo, J.; García-Verdugo, J.M.; Larraín, J. Cellular response to spinal cord injury in regenerative and non-regenerative stages in Xenopus laevis. Neural Dev. 2021, 16, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, R.; Edwards-Faret, G.; Moreno, M.; Zuñiga, N.; Cline, H.; Larraín, J. Regeneration of Xenopus laevis spinal cord requires Sox2/3 expressing cells. Dev. Biol. 2015, 408, 229–243. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Wang, W.; Ren, Y.; Zhang, W.; Fang, P.; Huang, L.; Wang, X.; Shi, P. Regeneration of cortical tissue from brain injury by implantation of defined molecular gradient of semaphorin 3A. Biomaterials 2018, 157, 125–135. [Google Scholar] [CrossRef]
- Chédotal, A. Roles of axon guidance molecules in neuronal wiring in the developing spinal cord. Nat. Rev. Neurosci. 2019, 20, 380–396. [Google Scholar] [CrossRef]
- Nawabi, H.; Briançon-Marjollet, A.; Clark, C.; Sanyas, I.; Takamatsu, H.; Okuno, T.; Kumanogoh, A.; Bozon, M.; Takeshima, K.; Yoshida, Y.; et al. A midline switch of receptor processing regulates commissural axon guidance in vertebrates. Genes Dev. 2010, 24, 396–410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mire, E.; Hocine, M.; Bazellières, E.; Jungas, T.; Davy, A.; Chauvet, S.; Mann, F. Developmental Upregulation of Ephrin-B1 Silences Sema3C/Neuropilin-1 Signaling during Post-crossing Navigation of Corpus Callosum Axons. Curr. Biol. 2018, 28, 1768–1782.e4. [Google Scholar] [CrossRef] [Green Version]
- Bonanomi, D. Axon pathfinding for locomotion. Semin. Cell Dev. Biol. 2019, 85, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, K.; Yamazaki, M.; Abe, M.; Sakimura, K.; Lickert, H.; Kawasaki, T.; Murakami, F.; Hirata, T. Netrin-1 Derived from the Ventricular Zone, but not the Floor Plate, Directs Hindbrain Commissural Axons to the Ventral Midline. Sci. Rep. 2017, 7, 11992. [Google Scholar] [CrossRef] [Green Version]
- Varadarajan, S.G.; Kong, J.H.; Phan, K.D.; Kao, T.J.; Panaitof, S.C.; Cardin, J.; Eltzschig, H.; Kania, A.; Novitch, B.G.; Butler, S.J. Netrin1 Produced by Neural Progenitors, Not Floor Plate Cells, Is Required for Axon Guidance in the Spinal Cord. Neuron 2017, 94, 790–799.e3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno-Bravo, J.A.; Puiggros, S.R.; Blockus, H.; Dominici, C.; Zelina, P.; Mehlen, P.; Chédotal, A. Commissural neurons transgress the cns/pns boundary in absence of ventricular zone-derived netrin 1. Development 2018, 145, dev159400. [Google Scholar] [CrossRef] [Green Version]
- Gu, Z.; Ueno, M.; Klinefelter, K.; Mamidi, M.; Yagi, T.; Yoshida, Y. Skilled movements in mice require inhibition of corticospinal axon collateral formation in the spinal cord by semaphorin signaling. J. Neurosci. 2019, 39, 8885–8899. [Google Scholar] [CrossRef]
- Gu, Z.; Kalamboglas, J.; Yoshioka, S.; Han, W.; Li, Z.; Kawasawa, Y.I.; Pochareddy, S.; Li, Z.; Liu, F.; Xu, X.; et al. Control of species-dependent cortico-motoneuronal connections underlying manual dexterity. Science 2017, 357, 400–404. [Google Scholar] [CrossRef] [Green Version]
- Gu, Z.; Koppel, N.; Kalamboglas, J.; Alexandrou, G.; Li, J.; Craig, C.; Simon, D.J.; Tessier-Lavigne, M.; Baccei, M.L.; Martin, J.H.; et al. Semaphorin-Mediated Corticospinal Axon Elimination Depends on the Activity-Induced Bax/Bak-Caspase Pathway. J. Neurosci. 2020, 40, 5402–5412. [Google Scholar] [CrossRef]
- Gu, Z.; Serradj, N.; Ueno, M.; Liang, M.; Li, J.; Baccei, M.L.; Martin, J.H.; Yoshida, Y. Skilled Movements Require Non-apoptotic Bax/Bak Pathway-Mediated Corticospinal Circuit Reorganization. Neuron 2017, 94, 626–641.e4. [Google Scholar] [CrossRef] [Green Version]
- Croteau, L.P.; Kao, T.J.; Kania, A. Ephrin-A5 potentiates netrin-1 axon guidance by enhancing Neogenin availability. Sci. Rep. 2019, 9, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Bonanomi, D.; Valenza, F.; Chivatakarn, O.; Sternfeld, M.J.; Driscoll, S.P.; Aslanian, A.; Lettieri, K.; Gullo, M.; Badaloni, A.; Lewcock, J.W.; et al. p190RhoGAP Filters Competing Signals to Resolve Axon Guidance Conflicts. Neuron 2019, 102, 602–620.e9. [Google Scholar] [CrossRef]
- Hung, R.J.; Terman, J.R. Extracellular inhibitors, repellents, and semaphorin/plexin/MICAL-mediated actin filament disassembly. Cytoskeleton 2011, 68, 415–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, E.; Strittmatter, S. The CRMP family of proteins and their role in Sema3A signaling. In Semaphorins: Receptor and Intracellular Signaling Mechanisms; Springer Science & Business Media: New York, NY, USA, 2007; pp. 1–11. [Google Scholar]
- Casazza, A.; Fazzari, P.; Tamagnone, L. Semaphorin signals in cell adhesion and cell migration: Functional role and molecular mechanisms. Adv. Exp. Med. Biol. 2007, 600, 90–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McConnell, R.E.; van Veen, J.E.; Vidaki, M.; Kwiatkowski, A.V.; Meyer, A.S.; Gertler, F.B. A requirement for filopodia extension toward Slit during Robo-mediated axon repulsion. J. Cell Biol. 2016, 213, 261–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menon, S.; Boyer, N.P.; Winkle, C.C.; McClain, L.M.; Hanlin, C.C.; Pandey, D.; Rothenfußer, S.; Taylor, A.M.; Gupton, S.L. The E3 Ubiquitin Ligase TRIM9 Is a Filopodia Off Switch Required for Netrin-Dependent Axon Guidance. Dev. Cell 2015, 35, 698–712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quadri, S.A.; Farooqui, M.; Ikram, A.; Zafar, A.; Khan, M.A.; Suriya, S.S.; Claus, C.F.; Fiani, B.; Rahman, M.; Ramachandran, A.; et al. Recent update on basic mechanisms of spinal cord injury. Neurosurg. Rev. 2020, 43, 425–441. [Google Scholar] [CrossRef]
- Wanner, I.B.; Anderson, M.A.; Song, B.; Levine, J.; Fernandez, A.; Gray-Thompson, Z.; Ao, Y.; Sofroniew, M.V. Glial scar borders are formed by newly proliferated, elongated astrocytes that interact to corral inflammatory and fibrotic cells via STAT3-dependent mechanisms after spinal cord injury. J. Neurosci. 2013, 33, 12870–12886. [Google Scholar] [CrossRef] [Green Version]
- Gu, Y.; Cheng, X.; Huang, X.; Yuan, Y.; Qin, S.; Tan, Z.; Wang, D.; Hu, X.; He, C.; Su, Z. Conditional ablation of reactive astrocytes to dissect their roles in spinal cord injury and repair. Brain. Behav. Immun. 2019, 80, 394–405. [Google Scholar] [CrossRef]
- Li, Y.; Raisman, G. Sprouts from Cut Corticospinal Axons Persist in the Presence of Astrocytic Scarring in Long-Term Lesions of the Adult Rat Spinal Cord. Exp. Neurol. 1995, 134, 102–111. [Google Scholar] [CrossRef] [PubMed]
- Hill, C.E.; Beattie, M.S.; Bresnahan, J.C. Degeneration and sprouting of identified descending supraspinal axons after contusive spinal cord injury in the rat. Exp. Neurol. 2001, 171, 153–169. [Google Scholar] [CrossRef]
- Kerschensteiner, M.; Schwab, M.E.; Lichtman, J.W.; Misgeld, T. In vivo imaging of axonal degeneration in the rat optic nerve. Nat. Med. 2005, 11, 360. [Google Scholar] [CrossRef]
- Tom, V.J.; Steinmetz, M.P.; Miller, J.H.; Doller, C.M.; Silver, J. Studies on the development and behavior of the dystrophic growth cone, the hallmark of regeneration failure, in an in vitro model of the glial scar and after spinal cord injury. J. Neurosci. 2004, 24, 6531–6539. [Google Scholar] [CrossRef] [Green Version]
- Seif, G.I.; Nomura, H.; Tator, C.H. Retrograde axonal degeneration (“dieback”) in the corticospinal tract after transection injury of the rat spinal cord: A confocal microscopy study. J. Neurotrauma 2007, 24, 1513–1528. [Google Scholar] [CrossRef] [PubMed]
- Ertürk, A.; Hellal, F.; Enes, J.; Bradke, F. Disorganized microtubules underlie the formation of retraction bulbs and the failure of axonal regeneration. J. Neurosci. 2007, 27, 9169–9180. [Google Scholar] [CrossRef] [Green Version]
- Hellal, F.; Hurtado, A.; Ruschel, J.; Flynn, K.C.; Laskowski, C.J.; Umlauf, M.; Kapitein, L.C.; Strikis, D.; Lemmon, V.; Bixby, J.; et al. Microtubule Stabilization Reduces Scarring and Causes Axon Regeneration after Spinal Cord Injury. Science 2011, 331, 928–931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghosh, S.; Hui, S.P. Axonal regeneration in zebrafish spinal cord. Regeneration 2018, 5, 43–60. [Google Scholar] [CrossRef]
- Bradke, F.; Fawcett, J.W.; Spira, M.E. Assembly of a new growth cone after axotomy: The precursor to axon regeneration. Nat. Rev. Neurosci. 2012, 13, 183–193. [Google Scholar] [CrossRef]
- Patrón, L.A.; Zinsmaier, K.E. Mitochondria on the Road to Power Axonal Regeneration. Neuron 2016, 92, 1152–1154. [Google Scholar] [CrossRef] [Green Version]
- Kiryu-Seo, S.; Kiyama, H. Mitochondrial behavior during axon regeneration/degeneration in vivo. Neurosci. Res. 2019, 139, 42–47. [Google Scholar] [CrossRef]
- Steketee, M.B.; Moysidis, S.N.; Weinstein, J.E.; Kreymerman, A.; Silva, J.P.; Iqbal, S.; Goldberg, J.L. Mitochondrial dynamics regulate growth cone motility, guidance, and neurite growth rate in perinatal retinal ganglion cells in vitro. Investig. Ophthalmol. Vis. Sci. 2012, 53, 7402–7411. [Google Scholar] [CrossRef] [PubMed]
- Sainath, R.; Armijo-Weingart, L.; Ketscheck, A.; Xu, Z.; Li, S.; Gallo, G. Chondroitin sulfate proteoglycans negatively regulate the positioning of mitochondria and endoplasmic reticulum to distal axons. Dev. Neurobiol. 2017, 77, 1351–1370. [Google Scholar] [CrossRef] [PubMed]
- Verburg, J.; Hollenbeck, P.J. Mitochondrial membrane potential in axons increases with local nerve growth factor or semaphorin signaling. J. Neurosci. 2008, 28, 8306–8315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Markham, A.; Cameron, I.; Franklin, P.; Spedding, M. BDNF increases rat brain mitochondrial respiratory coupling at complex I, but not complex II. Eur. J. Neurosci. 2004, 20, 1189–1196. [Google Scholar] [CrossRef] [PubMed]
- Cavallucci, V.; Bisicchia, E.; Cencioni, M.T.; Ferri, A.; Latini, L.; Nobili, A.; Biamonte, F.; Nazio, F.; Fanelli, F.; Moreno, S.; et al. Acute focal brain damage alters mitochondrial dynamics and autophagy in axotomized neurons. Cell Death Dis. 2014, 5, e1545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, B.; Yu, P.; Lin, M.Y.; Sun, T.; Chen, Y.; Sheng, Z.H. Facilitation of axon regeneration by enhancing mitochondrial transport and rescuing energy deficits. J. Cell Biol. 2016, 214, 103–119. [Google Scholar] [CrossRef] [Green Version]
- Han, Q.; Xie, Y.; Ordaz, J.D.; Huh, A.J.; Huang, N.; Wu, W.; Liu, N.; Chamberlain, K.A.; Sheng, Z.H.; Xu, X.M. Restoring Cellular Energetics Promotes Axonal Regeneration and Functional Recovery after Spinal Cord Injury. Cell Metab. 2020, 31, 623–641.e8. [Google Scholar] [CrossRef]
- Jha, M.K.; Lee, W.H.; Suk, K. Functional polarization of neuroglia: Implications in neuroinflammation and neurological disorders. Biochem. Pharmacol. 2016, 103, 1–16. [Google Scholar] [CrossRef]
- Kigerl, K.A.; Gensel, J.C.; Ankeny, D.P.; Alexander, J.K.; Donnelly, D.J.; Popovich, P.G. Identification of two distinct macrophage subsets with divergent effects causing either neurotoxicity or regeneration in the injured mouse spinal cord. J. Neurosci. 2009, 29, 13435–13444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zigmond, R.E.; Echevarria, F.D. Macrophage Biology in the Peripheral Nervous System after Injury. Prog. Neurobiol. 2019, 173, 102–121. [Google Scholar] [CrossRef] [PubMed]
- Tsarouchas, T.M.; Wehner, D.; Cavone, L.; Munir, T.; Keatinge, M.; Lambertus, M.; Underhill, A.; Barrett, T.; Kassapis, E.; Ogryzko, N.; et al. Dynamic control of proinflammatory cytokines Il-1β and Tnf-α by macrophages in zebrafish spinal cord regeneration. Nat. Commun. 2018, 9, 4670. [Google Scholar] [CrossRef] [Green Version]
- Feinstein, J.; Ramkhelawon, B. Netrins & Semaphorins: Novel regulators of the immune response. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 3183–3189. [Google Scholar] [CrossRef]
- Mirakaj, V.; Rosenberger, P. Immunomodulatory Functions of Neuronal Guidance Proteins. Trends Immunol. 2017, 38, 444–456. [Google Scholar] [CrossRef] [PubMed]
- Okuno, T.; Nakatsuji, Y.; Moriya, M.; Takamatsu, H.; Nojima, S.; Takegahara, N.; Toyofuku, T.; Nakagawa, Y.; Kang, S.; Friedel, R.H.; et al. Roles of Sema4D–Plexin-B1 Interactions in the Central Nervous System for Pathogenesis of Experimental Autoimmune Encephalomyelitis. J. Immunol. 2010, 184, 1499–1506. [Google Scholar] [CrossRef] [Green Version]
- Toguchi, M.; Gonzalez, D.; Furukawa, S.; Inagaki, S. Involvement of Sema4D in the control of microglia activation. Neurochem. Int. 2009, 55, 573–580. [Google Scholar] [CrossRef]
- Nakatsuji, Y.; Okuno, T.; Moriya, M.; Sugimoto, T.; Kinoshita, M.; Takamatsu, H.; Nojima, S.; Kimura, T.; Kang, S.; Ito, D.; et al. Elevation of Sema4A Implicates Th Cell Skewing and the Efficacy of IFN-β Therapy in Multiple Sclerosis. J. Immunol. 2012, 188, 4858–4865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koda, T.; Namba, A.; Kinoshita, M.; Nakatsuji, Y.; Sugimoto, T.; Sakakibara, K.; Tada, S.; Shimizu, M.; Yamashita, K.; Takata, K.; et al. Sema4A is implicated in the acceleration of Th17 cell-mediated neuroinflammation in the effector phase. J. Neuroinflammation 2020, 17, 82. [Google Scholar] [CrossRef]
- Ben-Gigi, L.; Sweetat, S.; Besser, E.; Fellig, Y.; Wiederhold, T.; Polakiewicz, R.D.; Behar, O. Astrogliosis induced by brain injury is regulated by sema4B phosphorylation. eNeuro 2015, 2, ENEURO.0078-14.2015. [Google Scholar] [CrossRef] [Green Version]
- Namavari, A.; Chaudhary, S.; Ozturk, O.; Chang, J.H.; Yco, L.; Sonawane, S.; Katam, N.; Khanolkar, V.; Hallak, J.; Sarkar, J.; et al. Semaphorin 7a links nerve regeneration and inflammation in the cornea. Investig. Ophthalmol. Vis. Sci. 2012, 53, 4575–4585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, Z.; Huang, L.; Enkhjargal, B.; Reis, C.; Wan, W.; Tang, J.; Cheng, Y.; Zhang, J.H. Recombinant Netrin-1 binding UNC5B receptor attenuates neuroinflammation and brain injury via PPARγ/NFκB signaling pathway after subarachnoid hemorrhage in rats. Brain. Behav. Immun. 2018, 69, 190–202. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Liu, Y.; Lin, X.; Yuan, F.; Long, D.; Zhang, Z.; Wang, Y.; Xuan, A.; Yang, G.Y. Netrin-1 attenuates brain injury after middle cerebral artery occlusion via downregulation of astrocyte activation in mice. J. Neuroinflammation 2018, 15, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mulero, P.; Córdova, C.; Hernández, M.; Martín, R.; Gutiérrez, B.; Muñoz, J.C.; Redondo, N.; Gallardo, I.; Téllez, N.; Nieto, M.L. Netrin-1 and multiple sclerosis: A new biomarker for neuroinflammation? Eur. J. Neurol. 2017, 24, 1108–1115. [Google Scholar] [CrossRef]
- Podjaski, C.; Alvarez, J.I.; Bourbonniere, L.; Larouche, S.; Terouz, S.; Bin, J.M.; Lécuyer, M.A.; Saint-Laurent, O.; Larochelle, C.; Darlington, P.J.; et al. Netrin 1 regulates blood-brain barrier function and neuroinflammation. Brain 2015, 138, 1598–1612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swidergall, M.; Solis, N.V.; Lionakis, M.S.; Filler, S.G. EphA2 is an epithelial cell pattern recognition receptor for fungal β-glucans. Nat. Microbiol. 2018, 3, 53–61. [Google Scholar] [CrossRef] [Green Version]
- Aaron, P.A.; Jamklang, M.; Uhrig, J.P.; Gelli, A. The blood–brain barrier internalises Cryptococcus neoformans via the EphA2-tyrosine kinase receptor. Cell. Microbiol. 2018, 20, e12811. [Google Scholar] [CrossRef] [Green Version]
- Tyzack, G.E.; Hall, C.E.; Sibley, C.R.; Cymes, T.; Forostyak, S.; Carlino, G.; Meyer, I.F.; Schiavo, G.; Zhang, S.C.; Gibbons, G.M.; et al. A neuroprotective astrocyte state is induced by neuronal signal EphB1 but fails in ALS models. Nat. Commun. 2017, 8, 1164. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, L.; Hua, F.; Zhuang, Y.; Liu, H.; Wang, S. EphA4 Obstructs Spinal Cord Neuron Regeneration by Promoting Excessive Activation of Astrocytes. Cell. Mol. Neurobiol. 2021, 1–12. [Google Scholar] [CrossRef]
- Parmentier-Batteur, S.; Finger, E.N.; Krishnan, R.; Rajapakse, H.A.; Sanders, J.M.; Kandpal, G.; Zhu, H.; Moore, K.P.; Regan, C.P.; Sharma, S.; et al. Attenuation of scratch-induced reactive astrogliosis by novel EphA4 kinase inhibitors. J. Neurochem. 2011, 118, 1016–1031. [Google Scholar] [CrossRef]
- Munro, K.M.; Perreau, V.M.; Turnley, A.M. Differential gene expression in the epha4 knockout spinal cord and analysis of the inflammatory response following spinal cord injury. PLoS ONE 2012, 7, e37635. [Google Scholar] [CrossRef] [PubMed]
- Sherchan, P.; Huang, L.; Wang, Y.; Akyol, O.; Tang, J.; Zhang, J.H. Recombinant Slit2 attenuates neuroinflammation after surgical brain injury by inhibiting peripheral immune cell infiltration via Robo1-srGAP1 pathway in a rat model. Neurobiol. Dis. 2016, 85, 164–173. [Google Scholar] [CrossRef] [Green Version]
- Sherchan, P.; Huang, L.; Akyol, O.; Reis, C.; Tang, J.; Zhang, J.H. Recombinant Slit2 Reduces Surgical Brain Injury Induced Blood Brain Barrier Disruption via Robo4 Dependent Rac1 Activation in a Rodent Model. Sci. Rep. 2017, 7, 746. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; He, X.; Li, H.; Wu, Y.; Guan, Y.; Liu, S.; Jia, H.; Li, Y.; Wang, L.; Huang, R.; et al. Overexpression of Slit2 improves function of the paravascular pathway in the aging mouse brain. Int. J. Mol. Med. 2018, 42, 1935–1944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Y.; Liu, X.; Chen, Z. Glial Scar—a Promising Target for Improving Outcomes After CNS Injury. J. Mol. Neurosci. 2020, 70, 340–352. [Google Scholar] [CrossRef] [PubMed]
- Min, Q.; Parkinson, D.B.; Dun, X.P. Migrating Schwann cells direct axon regeneration within the peripheral nerve bridge. Glia 2021, 69, 235–254. [Google Scholar] [CrossRef]
- Goldshmit, Y.; Sztal, T.E.; Jusuf, P.R.; Hall, T.E.; Nguyen-Chi, M.; Currie, P.D. Fgf-dependent glial cell bridges facilitate spinal cord regeneration in Zebrafish. J. Neurosci. 2012, 32, 7477–7492. [Google Scholar] [CrossRef] [Green Version]
- Bradbury, E.J.; Burnside, E.R. Moving beyond the glial scar for spinal cord repair. Nat. Commun. 2019, 10, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, X.; Tian, J.; Shan, J.; Hu, Y.; Zhai, Y.; Guo, J. Talin promotes integrin activation accompanied by generation of tension in talin and an increase in osmotic pressure in neurite outgrowth. FASEB J. 2019, 33, 6311–6326. [Google Scholar] [CrossRef]
- Anderson, M.A.; Burda, J.E.; Ren, Y.; Ao, Y.; O’Shea, T.M.; Kawaguchi, R.; Coppola, G.; Khakh, B.S.; Deming, T.J.; Sofroniew, M.V. Astrocyte scar formation AIDS central nervous system axon regeneration. Nature 2016, 532, 195–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, M.A.; O’Shea, T.M.; Burda, J.E.; Ao, Y.; Barlatey, S.L.; Bernstein, A.M.; Kim, J.H.; James, N.D.; Rogers, A.; Kato, B.; et al. Required growth facilitators propel axon regeneration across complete spinal cord injury. Nature 2018, 561, 396–400. [Google Scholar] [CrossRef] [PubMed]
- Pasterkamp, R.J.; De Winter, F.; Giger, R.J.; Verhaagen, J. Role for semaphorin III and its receptor neuropilin-1 in neuronal regeneration and scar formation? Prog. Brain Res. 1998, 117, 151–170. [Google Scholar] [CrossRef]
- Pasterkamp, R.J.; Giger, R.J.; Ruitenberg, M.J.; Holtmaat, A.J.G.D.; De Wit, J.; De Winter, F.; Verhaagen, J. Expression of the gene encoding the chemorepellent semaphorin III is induced in the fibroblast component of neural scar tissue formed following injuries of adult but not neonatal CNS. Mol. Cell. Neurosci. 1999, 13, 143–166. [Google Scholar] [CrossRef] [PubMed]
- De Winter, F.; Oudega, M.; Lankhorst, A.J.; Hamers, F.P.; Blits, B.; Ruitenberg, M.J.; Pasterkamp, R.J.; Gispen, W.H.; Verhaagen, J. Injury-induced class 3 semaphorin expression in the rat spinal cord. Exp. Neurol. 2002, 175, 61–75. [Google Scholar] [CrossRef]
- Jeroen Pasterkamp, R.; Anderson, P.N.; Verhaagen, J. Peripheral nerve injury fails to induce growth of lesioned ascending dorsal column axons into spinal cord scar tissue expressing the axon repellent Semaphorin3A. Eur. J. Neurosci. 2001, 13, 457–471. [Google Scholar] [CrossRef]
- Kaneko, S.; Iwanami, A.; Nakamura, M.; Kishino, A.; Kikuchi, K.; Shibata, S.; Okano, H.J.; Ikegami, T.; Moriya, A.; Konishi, O.; et al. A selective Sema3A inhibitor enhances regenerative responses and functional recovery of the injured spinal cord. Nat. Med. 2006, 12, 1380–1389. [Google Scholar] [CrossRef] [PubMed]
- Hira, K.; Ueno, Y.; Tanaka, R.; Miyamoto, N.; Yamashiro, K.; Inaba, T.; Urabe, T.; Okano, H.; Hattori, N. Astrocyte-derived exosomes treated with a semaphorin 3A inhibitor enhance stroke recovery via prostaglandin D2 synthase. Stroke 2018, 49, 2483–2494. [Google Scholar] [CrossRef]
- Kopp, M.A.; Brommer, B.; Gatzemeier, N.; Schwab, J.M.; Prüss, H. Spinal cord injury induces differential expression of the profibrotic semaphorin 7A in the developing and mature glial scar. Glia 2010, 58, 1748–1756. [Google Scholar] [CrossRef]
- Bundesen, L.Q.; Scheel, T.A.; Bregman, B.S.; Kromer, L.F. Ephrin-B2 and EphB2 regulation of astrocyte-meningeal fibroblast interactions in response to spinal cord lesions in adult rats. J. Neurosci. 2003, 23, 7789–7800. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Lu, B.; Yang, R.; Chen, Y.; Chen, X.; Li, Y. EphB2 knockdown decreases the formation of astroglial-fibrotic scars to promote nerve regeneration after spinal cord injury in rats. CNS Neurosci. Ther. 2021, 27, 714–724. [Google Scholar] [CrossRef] [PubMed]
- Ren, Z.; Chen, X.; Yang, J.; Kress, B.T.; Tong, J.; Liu, H.; Takano, T.; Zhao, Y.; Nedergaard, M. Improved axonal regeneration after spinal cord injury in mice with conditional deletion of ephrin B2 under the GFAP promoter. Neuroscience 2013, 241, 89–99. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Chen, Y.; Tan, L.; Pan, J.Y.; Lin, W.W.; Wu, J.; Hu, W.; Chen, X.; Wang, X.D. RNAi-mediated ephrin-B2 silencing attenuates astroglial-fibrotic scar formation and improves spinal cord axon growth. CNS Neurosci. Ther. 2017, 23, 779–789. [Google Scholar] [CrossRef] [PubMed]
- Goldshmit, Y.; Galea, M.P.; Wise, G.; Bartlett, P.F.; Turnley, A.M. Axonal regeneration and lack of astrocytic gliosis in EphA4-deficient mice. J. Neurosci. 2004, 24, 10064–10073. [Google Scholar] [CrossRef] [Green Version]
- Goldshmit, Y.; Bourne, J. Upregulation of epha4 on astrocytes potentially mediates astrocytic gliosis after cortical lesion in the marmoset monkey. J. Neurotrauma 2010, 27, 1321–1332. [Google Scholar] [CrossRef]
- Zhou, X.; Wahane, S.; Friedl, M.S.; Kluge, M.; Friedel, C.C.; Avrampou, K.; Zachariou, V.; Guo, L.; Zhang, B.; He, X.; et al. Microglia and macrophages promote corralling, wound compaction and recovery after spinal cord injury via Plexin-B2. Nat. Neurosci. 2020, 23, 337–350. [Google Scholar] [CrossRef] [PubMed]
- Moreau-Fauvarque, C.; Kumanogoh, A.; Camand, E.; Jaillard, C.; Barbin, G.; Boquet, I.; Love, C.; Jones, E.Y.; Kikutani, H.; Lubetzki, C.; et al. The transmembrane semaphorin Sema4D/CD100, an inhibitor of axonal growth, is expressed on oligodendrocytes and upregulated after CNS lesion. J. Neurosci. 2003, 23, 9229–9239. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.L.; Wang, J.; Tang, L. Sema4D Knockdown in Oligodendrocytes Promotes Functional Recovery After Spinal Cord Injury. Cell Biochem. Biophys. 2014, 68, 489–496. [Google Scholar] [CrossRef]
- Wehrle, R.; Camand, E.; Chedotal, A.; Sotelo, C.; Dusart, I. Expression of netrin-1, slit-1 and slit-3 but not of slit-2 after cerebellar and spinal cord lesions. Eur. J. Neurosci. 2005, 22, 2134–2144. [Google Scholar] [CrossRef]
- Hagino, S.; Iseki, K.; Mori, T.; Zhang, Y.; Hikake, T.; Yokoya, S.; Takeuchi, M.; Hasimoto, H.; Kikuchi, S.; Wanaka, A. Slit and glypican-1 mRNAs are coexpressed in the reactive astrocytes of the injured adult brain. Glia 2003, 42, 130–138. [Google Scholar] [CrossRef]
- Dun, X.P.; Parkinson, D. Classic axon guidance molecules control correct nerve bridge tissue formation and precise axon regeneration. Neural Regen. Res. 2020, 15, 6–9. [Google Scholar] [CrossRef] [PubMed]
- Parrinello, S.; Napoli, I.; Ribeiro, S.; Digby, P.W.; Fedorova, M.; Parkinson, D.B.; Doddrell, R.D.S.; Nakayama, M.; Adams, R.H.; Lloyd, A.C. EphB signaling directs peripheral nerve regeneration through Sox2-dependent Schwann cell Sorting. Cell 2010, 143, 145–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, R.; Yang, X.; Zhang, B.; Wang, S.; Bao, S.; Gu, Y.; Li, S. EphA4 Negatively Regulates Myelination by Inhibiting Schwann Cell Differentiation in the Peripheral Nervous System. Front. Neurosci. 2019, 13, 1191. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Carr, L.; Dun, X.P. Dynamic expression of Slit1-3 and Robo1-2 in the mouse peripheral nervous system after injury. Neural Regen. Res. 2020, 15, 948–958. [Google Scholar] [CrossRef]
- Dun, X.P.; Carr, L.; Woodley, P.K.; Barry, R.W.; Drake, L.K.; Mindos, T.; Roberts, S.L.; Lloyd, A.C.; Parkinson, D.B. Macrophage-Derived Slit3 Controls Cell Migration and Axon Pathfinding in the Peripheral Nerve Bridge. Cell Rep. 2019, 26, 1458–1472.e4. [Google Scholar] [CrossRef] [Green Version]
- Rosenberg, A.F.; Isaacman-Beck, J.; Franzini-Armstrong, C.; Granato, M. Schwann cells and deleted in colorectal carcinoma direct regenerating motor axons towards their original path. J. Neurosci. 2014, 34, 14668–14681. [Google Scholar] [CrossRef] [Green Version]
- Isaacman-Beck, J.; Schneider, V.; Franzini-Armstrong, C.; Granato, M. The lh3 Glycosyltransferase Directs Target-Selective Peripheral Nerve Regeneration. Neuron 2015, 88, 691–703. [Google Scholar] [CrossRef] [Green Version]
- Cigliola, V.; Becker, C.J.; Poss, K.D. Building bridges, not walls: Spinal cord regeneration in zebrafish. DMM Dis. Model. Mech. 2020, 13, dmm044131. [Google Scholar] [CrossRef]
- Peng, S.X.; Yao, L.; Cui, C.; Zhao, H.D.; Liu, C.J.; Li, Y.H.; Wang, L.F.; Huang, S.B.; Shen, Y.Q. Semaphorin4D promotes axon regrowth and swimming ability during recovery following zebrafish spinal cord injury. Neuroscience 2017, 351, 36–46. [Google Scholar] [CrossRef]
- Edwards-Faret, G.; Cebrián-Silla, A.; Méndez-Olivos, E.E.; González-Pinto, K.; García-Verdugo, J.M.; Larraín, J. Cellular composition and organization of the spinal cord central canal during metamorphosis of the frog Xenopus laevis. J. Comp. Neurol. 2018, 526, 1712–1732. [Google Scholar] [CrossRef]
- Gaete, M.; Muñoz, R.; Sánchez, N.; Tampe, R.; Moreno, M.; Contreras, E.G.; Lee-Liu, D.; Larraín, J. Spinal cord regeneration in Xenopus tadpoles proceeds through activation of Sox2-positive cells. Neural Dev. 2012, 7, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashimoto, M.; Ino, H.; Koda, M.; Murakami, M.; Yoshinaga, K.; Yamazaki, M.; Moriya, H. Regulation of semaphorin 3A expression in neurons of the rat spinal cord and cerebral cortex after transection injury. Acta Neuropathol. 2004, 107, 250–256. [Google Scholar] [CrossRef] [PubMed]
- Lindholm, T.; Sköld, M.K.; Suneson, A.; Carlstedt, T.; Cullheim, S.; Risling, M. Semaphorin and neuropilin expression in motoneurons after intraspinal motoneuron axotomy. Neuroreport 2004, 15, 649–654. [Google Scholar] [CrossRef] [PubMed]
- Manitt, C.; Colicos, M.A.; Thompson, K.M.; Rousselle, E.; Peterson, A.C.; Kennedy, T.E. Widespread expression of netrin-1 by neurons and oligodendrocytes in the adult mammalian spinal cord. J. Neurosci. 2001, 21, 3911–3922. [Google Scholar] [CrossRef] [PubMed]
- Löw, K.; Culbertson, M.; Bradke, F.; Tessier-Lavigne, M.; Tuszynski, M.H. Netrin-1 is a novel myelin-associated inhibitor to axon growth. J. Neurosci. 2008, 28, 1099–1108. [Google Scholar] [CrossRef] [Green Version]
- Manitt, C.; Wang, D.; Kennedy, T.E.; Howland, D.R. Positioned to Inhibit: Netrin-1 and Netrin Receptor Expression After Spinal Cord Injury. J. Neurosci. Res. 2006, 84, 1808–1820. [Google Scholar] [CrossRef]
- Lu, Y.J.; Xu, N.W.; Yang, W.Q. Immunofluorescence laser confocal expression and localization study of rat nerve growth guidance cues Netrin-1 and Slit2 after spinal cord injury. Chin. J. Traumatol. Engl. Ed. 2008, 11, 98–103. [Google Scholar] [CrossRef] [Green Version]
- Ahn, K.J.; Seo, I.A.; Lee, H.K.; Choi, E.J.; Seo, E.H.; Lee, H.J.; Park, H.T. Down-regulation of UNC5 homologue expression after the spinal cord injury in the adult rat. Neurosci. Lett. 2007, 419, 43–48. [Google Scholar] [CrossRef]
- Liu, J.B.; Jiang, Y.Q.; Gong, A.H.; Zhang, Z.J.; Jiang, Q.; Chu, X.P. Expression of Slit2 and Robo1 after traumatic lesions of the rat spinal cord. Acta Histochem. 2011, 113, 43–48. [Google Scholar] [CrossRef]
- Li, Y.; Gao, Y.; Xu, X.; Shi, R.; Liu, J.; Yao, W.; Ke, C. Slit2/Robo1 promotes synaptogenesis and functional recovery of spinal cord injury. Neuroreport 2017, 28, 75–81. [Google Scholar] [CrossRef]
- Fabes, J.; Anderson, P.; Yáñez-Muñoz, R.J.; Thrasher, A.; Brennan, C.; Bolsover, S. Accumulation of the inhibitory receptor EphA4 may prevent regeneration of corticospinal tract axons following lesion. Eur. J. Neurosci. 2006, 23, 1721–1730. [Google Scholar] [CrossRef]
- Arocho, L.C.; Figueroa, J.D.; Torrado, A.I.; Santiago, J.M.; Vera, A.E.; Miranda, J.D. Expression profile and role of EphrinA1 ligand after spinal cord injury. Cell. Mol. Neurobiol. 2011, 31, 1057–1069. [Google Scholar] [CrossRef] [Green Version]
- Miranda, J.D.; White, L.A.; Marcillo, A.E.; Willson, C.A.; Jagid, J.; Whittemore, S.R. Induction of Eph B3 after spinal cord injury. Exp. Neurol. 1999, 156, 218–222. [Google Scholar] [CrossRef]
- Willson, C.A.; Miranda, J.D.; Foster, R.D.; Onifer, S.M.; Whittemore, S.R. Transection of the adult rat spinal cord upregulates EphB3 receptor and ligand expression. Cell Transplant. 2003, 12, 279–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cruz-Orengo, L.; Figueroa, J.D.; Velázquez, I.; Torrado, A.; Ortíz, C.; Hernández, C.; Puig, A.; Segarra, A.C.; Whittemore, S.R.; Miranda, J.D. Blocking EphA4 upregulation after spinal cord injury results in enhanced chronic pain. Exp. Neurol. 2006, 202, 421–433. [Google Scholar] [CrossRef]
- Pasterkamp, R.J.; Giger, R.J.; Verhaagen, J. Regulation of semaphorin III/collapsin-1 gene expression during peripheral nerve regeneration. Exp. Neurol. 1998, 153, 313–327. [Google Scholar] [CrossRef]
- He, Q.R.; Cong, M.; Chen, Q.Z.; Sheng, Y.F.; Li, J.; Zhang, Q.; Ding, F.; Gong, Y.P. Expression changes of nerve cell adhesion molecules l1 and semaphorin 3a after peripheral nerve injury. Neural Regen. Res. 2016, 11, 2025–2030. [Google Scholar] [CrossRef]
- Lindholm, T.; Risling, M.; Carlstedt, T.; Hammarberg, H.; Wallquist, W.; Cullheim, S.; Sköld, M.K. Expression of semaphorins, neuropilins, vegf, and tenascins in rat and human primary sensory neurons after a dorsal root injury. Front. Neurol. 2017, 8, 49. [Google Scholar] [CrossRef] [Green Version]
- Scarlato, M.; Ara, J.; Bannerman, P.; Scherer, S.; Pleasure, D. Induction of neuropilins-1 and -2 and their ligands, Sema3A, Sema3F, and VEGF, during Wallerian degeneration in the peripheral nervous system. Exp. Neurol. 2003, 183, 489–498. [Google Scholar] [CrossRef]
- Madison, R.D.; Zomorodi, A.; Robinson, G.A. Netrin-1 and peripheral nerve regeneration in the adult rat. Exp. Neurol. 2000, 161, 563–570. [Google Scholar] [CrossRef] [PubMed]
- Moon, C.; Kim, H.; Ahn, M.; Jin, J.K.; Wang, H.; Matsumoto, Y.; Shin, T. Enhanced expression of netrin-1 protein in the sciatic nerves of Lewis rats with experimental autoimmune neuritis: Possible role of the netrin-1/DCC binding pathway in an autoimmune PNS disorder. J. Neuroimmunol. 2006, 172, 66–72. [Google Scholar] [CrossRef]
- Wang, X.; Chen, Q.; Yi, S.; Liu, Q.; Zhang, R.; Wang, P.; Qian, T.; Li, S. The microRNAs let-7 and miR-9 down-regulate the axon-guidance genes Ntn1 and Dcc during peripheral nerve regeneration. J. Biol. Chem. 2019, 294, 3489–3500. [Google Scholar] [CrossRef] [Green Version]
- Webber, C.A.; Christie, K.J.; Cheng, C.; Martinez, J.A.; Singh, B.; Singh, V.; Thomas, D.; Zochodne, D.W. Schwann cells direct peripheral nerve regeneration through the Netrin-1 receptors, DCC and Unc5H2. Glia 2011, 59, 1503–1517. [Google Scholar] [CrossRef] [PubMed]
- Jaminet, P.; Köhler, D.; Rahmanian-schwarz, A.; Lotter, O.; Mager, A.; Fornaro, M.; Ronchi, G.; Geuna, S.; Rosenberger, P.; Schaller, H.-E. Expression patterns and functional evaluation of the UNC5b receptor during the early phase of peripheral nerve regeneration using the mouse median nerve model. Microsurgery 2013, 33, 216–222. [Google Scholar] [CrossRef] [Green Version]
- Tanno, T.; Fujiwara, A.; Takenaka, S.; Kuwamura, M.; Tsuyama, S. Expression of a chemorepellent factor, slit2, in peripheral nerve regeneration. Biosci. Biotechnol. Biochem. 2005, 69, 2431–2434. [Google Scholar] [CrossRef]
- Yi, X.N.; Zheng, L.F.; Zhang, J.W.; Zhang, L.Z.; Xu, Y.Z.; Luo, G.; Luo, X.G. Dynamic changes in Robo2 and Slit1 expression in adult rat dorsal root ganglion and sciatic nerve after peripheral and central axonal injury. Neurosci. Res. 2006, 56, 314–321. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Zhao, J.; Shen, J.; Zhang, X.; Ren, R.; Ma, Z.; He, Y.; Kang, Q.; Wang, Y.; Dong, X.; et al. The ATP-P2X7 Signaling Pathway Participates in the Regulation of Slit1 Expression in Satellite Glial Cells. Front. Cell. Neurosci. 2019, 13, 420. [Google Scholar] [CrossRef]
- Chen, Z.B.; Zhang, H.Y.; Zhao, J.H.; Zhao, W.; Zhao, D.; Zheng, L.F.; Zhang, X.F.; Liao, X.P.; Yi, X.N. Slit-Robo GTPase-Activating Proteins are Differentially Expressed in Murine Dorsal Root Ganglia: Modulation by Peripheral Nerve Injury. Anat. Rec. 2012, 295, 652–660. [Google Scholar] [CrossRef]
- Bloechlinger, S.; Karchewski, L.A.; Woolf, C.J. Dynamic changes in glypican-1 expression in dorsal root ganglion neurons after peripheral and central axonal injury. Eur. J. Neurosci. 2004, 19, 1119–1132. [Google Scholar] [CrossRef]
- Shifman, M.I.; Selzer, M.E. Differential Expression of Class 3 and 4 Semaphorins and Netrin in the Lamprey Spinal Cord during Regeneration. J. Comp. Neurol. 2007, 501, 631–646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shifman, M.I.; Selzer, M.E. Expression of the Netrin Receptor UNC-5 in Lamprey Brain: Modulation by Spinal Cord Transection. Neurorehabil. Neural Repair 2000, 14, 49–58. [Google Scholar] [CrossRef]
- Goulart, C.O.; Mendonça, H.R.; Oliveira, J.T.; Savoldi, L.M.; Dos Santos Heringer, L.; Dos Santos Rodrigues, A.; Paes-De-Carvalho, R.; Martinez, A.M.B. Repulsive environment attenuation during adult mouse optic nerve regeneration. Neural Plast. 2018, 2018, 5851914. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.K.; Chow, R.; Xie, F.; Chow, S.Y.; Tolentino, K.E.; Zheng, B. Combined genetic attenuation of myelin and semaphorin-mediated growth inhibition is insufficient to promote serotonergic axon regeneration. J. Neurosci. 2010, 30, 10899–10904. [Google Scholar] [CrossRef] [Green Version]
- Petrausch, B.; Jung, M.; Leppert, C.A.; Stuermer, C.A.O. Lesion-induced regulation of netrin receptors and modification of netrin-1 expression in the retina of fish and grafted rats. Mol. Cell. Neurosci. 2000, 16, 350–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cebrià, F.; Newmark, P.A. Planarian homologs of netrin and netrin receptor are required for proper regeneration of the central nervous system and the maintenance of nervous system architecture. Development 2005, 132, 3691–3703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Laramore, C.; Shifman, M.I. The expression of chemorepulsive guidance receptors and the regenerative abilities of spinal-projecting neurons after spinal cord injury. Neuroscience 2017, 341, 95–111. [Google Scholar] [CrossRef]
- Cebrià, F.; Guo, T.; Jopek, J.; Newmark, P.A. Regeneration and maintenance of the planarian midline is regulated by a slit orthologue. Dev. Biol. 2007, 307, 394–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasidharan, V.; Marepally, S.; Elliott, S.A.; Baid, S.; Lakshmanan, V.; Nayyar, N.; Bansal, D.; Sańchez Alvarado, A.; Vemula, P.K.; Palakodeti, D. The miR-124 family of microRNAs is crucial for regeneration of the brain and visual system in the planarian schmidtea mediterranea. Development 2017, 144, 3211–3223. [Google Scholar] [CrossRef] [Green Version]
- Duffy, P.; Wang, X.; Siegel, C.S.; Tu, N.; Henkemeyer, M.; Cafferty, W.B.J.; Strittmatter, S.M. Myelin-derived ephrinB3 restricts axonal regeneration and recovery after adult CNS injury. Proc. Natl. Acad. Sci. USA 2012, 109, 5063–5068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mladinic, M.; Lefèvre, C.; Del Bel, E.; Nicholls, J.; Digby, M. Developmental changes of gene expression after spinal cord injury in neonatal opossums. Brain Res. 2010, 1363, 20–39. [Google Scholar] [CrossRef]
- Cruz-Orengo, L.; Figueroa, J.D.; Torrado, A.; Puig, A.; Whittemore, S.R.; Miranda, J.D. Reduction of EphA4 receptor expression after spinal cord injury does not induce axonal regeneration or return of tcMMEP response. Neurosci. Lett. 2007, 418, 49–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shirvan, A.; Kimron, M.; Holdengreber, V.; Ziv, I.; Ben-Shaul, Y.; Melamed, S.; Melamed, E.; Barzilai, A.; Solomon, A.S. Anti-semaphorin 3A antibodies rescue retinal ganglion cells from cell death following optic nerve axotomy. J. Biol. Chem. 2002, 277, 49799–49807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanemaru, H.; Yamada, Y.; Ohazama, A.; Maeda, T.; Seo, K. Semaphorin 3A Inhibits Nerve Regeneration during Early Stage after Inferior Alveolar Nerve Transection. Sci. Rep. 2019, 9, 4245. [Google Scholar] [CrossRef]
- Zhang, L.; Kaneko, S.; Kikuchi, K.; Sano, A.; Maeda, M.; Kishino, A.; Shibata, S.; Mukaino, M.; Toyama, Y.; Liu, M.; et al. Rewiring of regenerated axons by combining treadmill training with semaphorin3A inhibition. Mol. Brain 2014, 7, 14. [Google Scholar] [CrossRef] [Green Version]
- Smith, E.S.; Jonason, A.; Reilly, C.; Veeraraghavan, J.; Fisher, T.; Doherty, M.; Klimatcheva, E.; Mallow, C.; Cornelius, C.; Leonard, J.E.; et al. SEMA4D compromises blood-brain barrier, activates microglia, and inhibits remyelination in neurodegenerative disease. Neurobiol. Dis. 2015, 73, 254–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- LaGanke, C.; Samkoff, L.; Edwards, K.; Henson, L.J.; Repovic, P.; Lynch, S.; Stone, L.; Mattson, D.; Galluzzi, A.; Fisher, T.L.; et al. Safety/tolerability of the anti-semaphorin 4D antibody VX15/2503 in a randomized phase 1 trial. Neurol. Neuroimmunol. NeuroInflamm. 2017, 4, e367. [Google Scholar] [CrossRef] [Green Version]
- Fabes, J.; Anderson, P.; Brennan, C.; Bolsover, S. Regeneration-enhancing effects of EphA4 blocking peptide following corticospinal tract injury in adult rat spinal cord. Eur. J. Neurosci. 2007, 26, 2496–2505. [Google Scholar] [CrossRef]
- Spanevello, M.D.; Tajouri, S.I.; Mirciov, C.; Kurniawan, N.; Pearse, M.J.; Fabri, L.J.; Owczarek, C.M.; Hardy, M.P.; Bradford, R.A.; Ramunno, M.L.; et al. Acute delivery of epha4-fc improves functional recovery after contusive spinal cord injury in rats. J. Neurotrauma 2013, 30, 1023–1024. [Google Scholar] [CrossRef] [Green Version]
- Ujigo, S.; Kamei, N.; Hadoush, H.; Fujioka, Y.; Miyaki, S.; Nakasa, T.; Tanaka, N.; Nakanishi, K.; Eguchi, A.; Sunagawa, T.; et al. Administration of MicroRNA-210 promotes spinal cord regeneration in mice. Spine 2014, 39, 1099–1107. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Yang, H.; Zhou, X.; Zhang, L.; Lu, X. MiR-93 Targeting EphA4 Promotes Neurite Outgrowth from Spinal Cord Neurons. J. Mol. Neurosci. 2016, 58, 517–524. [Google Scholar] [CrossRef]
- Hu, Y.W.; Jiang, J.J.; Yan-Gao; Wang, R.Y.; Tu, G.J. MicroRNA-210 promotes sensory axon regeneration of adult mice in vivo and in vitro. Neurosci. Lett. 2016, 622, 61–66. [Google Scholar] [CrossRef]
- Wang, J.; Zhai, W.; Yu, Z.; Sun, L.; Li, H.; Shen, H.; Li, X.; Liu, C.; Chen, G. Neuroprotection exerted by Netrin-1 and kinesin motor KIF1A in secondary brain injury following experimental intracerebral hemorrhage in rats. Front. Cell. Neurosci. 2018, 11, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Bähr, M.; Eschweiler, G.W.; Wolburg, H. Precrushed sciatic nerve grafts enhance the survival and axonal regrowth of retinal ganglion cells in adult rats. Exp. Neurol. 1992, 116, 13–22. [Google Scholar] [CrossRef]
- Ellezam, B.; Selles-Navarro, I.; Manitt, C.; Kennedy, T.E.; McKerracher, L. Expression of netrin-1 and its receptors DCC and UNC-5H2 after axotomy and during regeneration of adult rat retinal ganglion cells. Exp. Neurol. 2001, 168, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Symonds, A.C.E.; King, C.E.; Bartlett, C.A.; Sauvé, Y.; Lund, R.D.; Beazley, L.D.; Dunlop, S.A.; Rodger, J. EphA5 and ephrin-A2 expression during optic nerve regeneration: A “two-edged sword”. Eur. J. Neurosci. 2007, 25, 744–752. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Domínguez-Romero, M.E.; Slater, P.G. Unraveling Axon Guidance during Axotomy and Regeneration. Int. J. Mol. Sci. 2021, 22, 8344. https://doi.org/10.3390/ijms22158344
Domínguez-Romero ME, Slater PG. Unraveling Axon Guidance during Axotomy and Regeneration. International Journal of Molecular Sciences. 2021; 22(15):8344. https://doi.org/10.3390/ijms22158344
Chicago/Turabian StyleDomínguez-Romero, Miguel E., and Paula G. Slater. 2021. "Unraveling Axon Guidance during Axotomy and Regeneration" International Journal of Molecular Sciences 22, no. 15: 8344. https://doi.org/10.3390/ijms22158344
APA StyleDomínguez-Romero, M. E., & Slater, P. G. (2021). Unraveling Axon Guidance during Axotomy and Regeneration. International Journal of Molecular Sciences, 22(15), 8344. https://doi.org/10.3390/ijms22158344