
The Impact of Early Life Exposure to Cannabis: The Role of the Endocannabinoid System

 , and
, and 
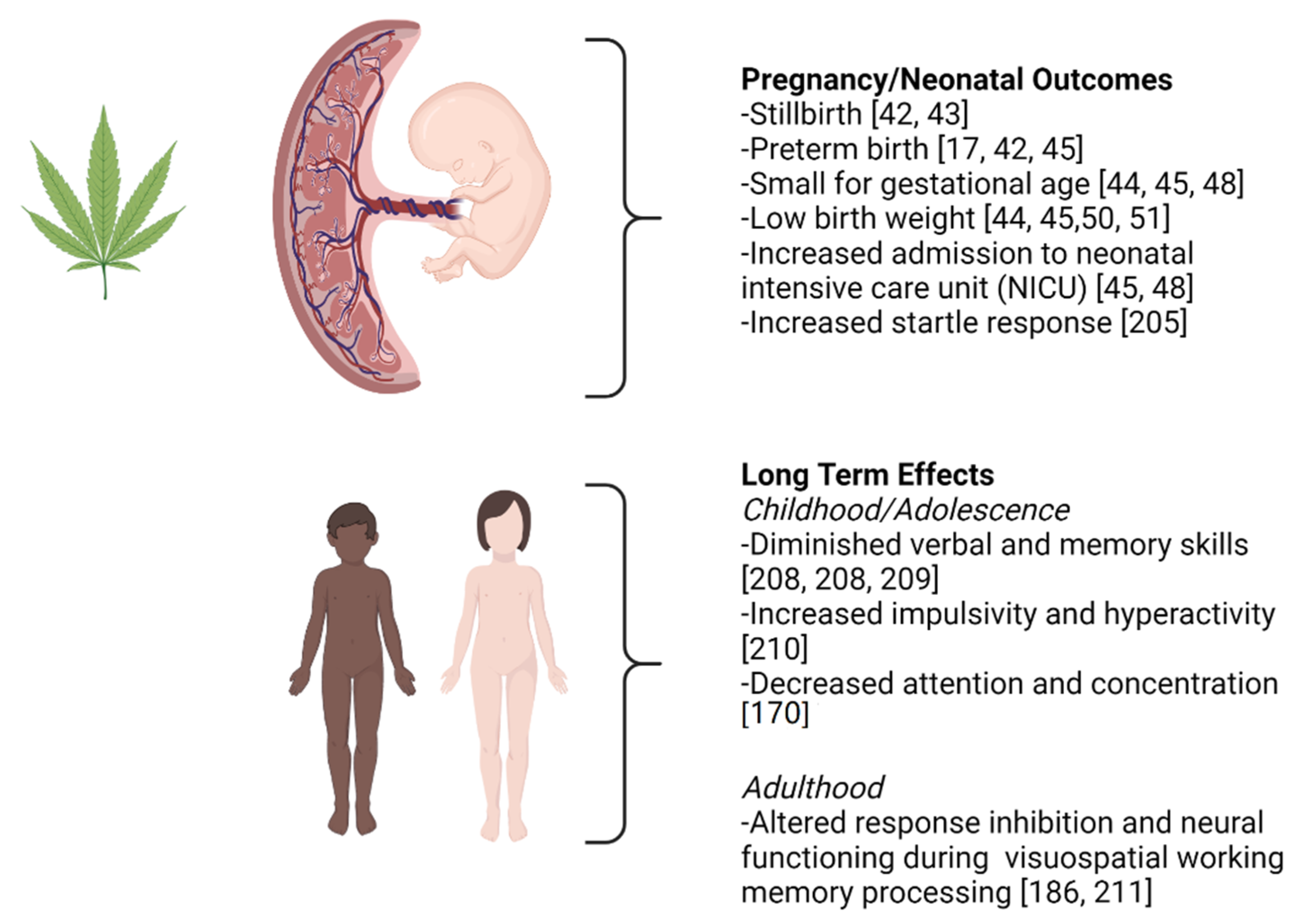{kind=link}
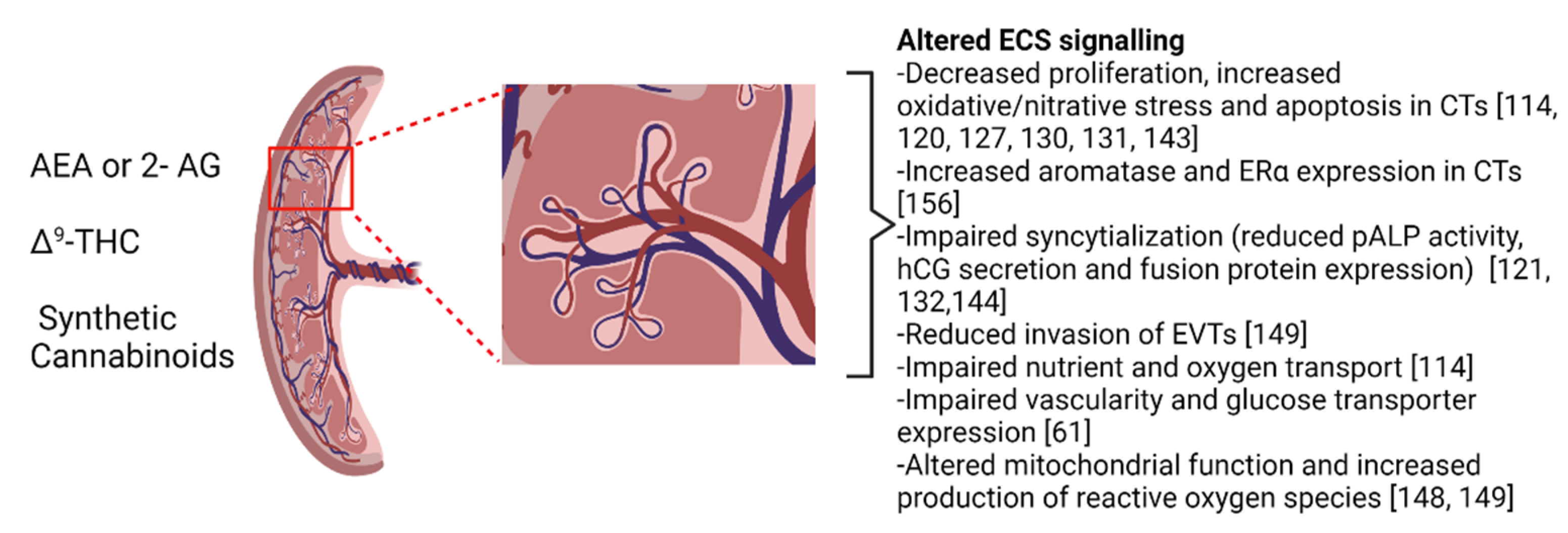{kind=link}
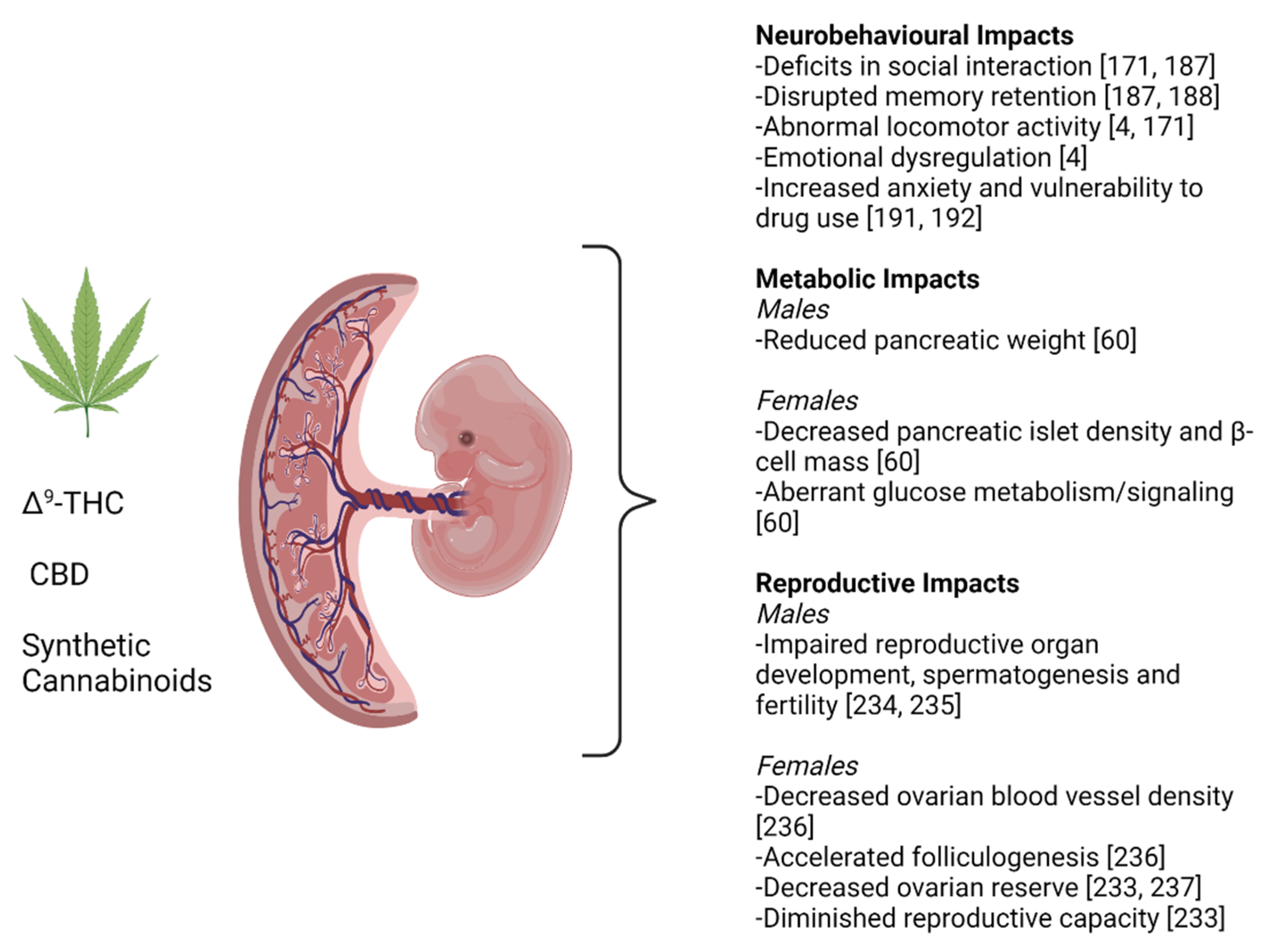{kind=link}
Abstract
:1. Introduction
2. The Endocannabinoid System
2.1. ECS Ligands
2.2. ECS Signaling
3. The ECS and the Placenta
3.1. Altered ECS Signaling in the Placenta
3.2. Impact of Exogenous Cannabinoids on the Placenta and ECS Signaling
4. Altered ECS Signaling during Fetal Development
4.1. Neurological Impacts
4.2. Metabolic Impacts
4.3. Reproductive Impacts



5. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- World Health Organization. Cannabis. Available online: https://www.who.int/teams/mental-health-and-substance-use/alcohol-drugs-and-addictive-behaviours/drugs-psychoactive/cannabis (accessed on 5 June 2021).
- Statistics Canada. National Cannabis Survey, Third Quarter 2019. Available online: https://www150.statcan.gc.ca/n1/daily-quotidien/191030/dq191030a-eng.htm (accessed on 5 June 2021).
- Alpar, A.; Di Marzo, V.; Harkany, T. At the tip of an iceberg: Prenatal marijuana and Its possible relation to neuropsychiatric outcome in the offspring. Biol. Psychiatry 2016, 79, e33–e45. [Google Scholar] [CrossRef] [PubMed]
- Campolongo, P.; Trezza, V.; Ratano, P.; Palmery, M.; Cuomo, V. Developmental consequences of perinatal cannabis exposure: Behavioral and neuroendocrine effects in adult rodents. Psychopharmacology 2011, 214, 5–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebrahim, S.H.; Gfroerer, J. Pregnancy-related substance use in the United States during 1996–1998. Obstet. Gynecol. 2003, 101, 374–379. [Google Scholar] [CrossRef]
- McCabe, J.E.; Arndt, S. Demographic and substance abuse trends among pregnant and non-pregnant women: Eleven years of treatment admission data. Matern. Child Health J. 2012, 16, 1696–1702. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.S.; Jew, C.P.; Lu, H.C. Lasting impacts of prenatal cannabis exposure and the role of endogenous cannabinoids in the developing brain. Future Neurol. 2011, 6, 459–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ko, J.Y.; Farr, S.L.; Tong, V.T.; Creanga, A.A.; Callaghan, W.M. Prevalence and patterns of marijuana use among pregnant and nonpregnant women of reproductive age. Am. J. Obstet. Gynecol. 2015, 213, 201.e1–201.e10. [Google Scholar] [CrossRef]
- Roberson, E.K.; Patrick, W.K.; Hurwitz, E.L. Marijuana use and maternal experiences of severe nausea during pregnancy in Hawai’i. Hawaii J. Med. Public Health 2014, 73, 283–287. [Google Scholar]
- Westfall, R.E.; Janssen, P.A.; Lucas, P.; Capler, R. Survey of medicinal cannabis use among childbearing women: Patterns of its use in pregnancy and retroactive self-assessment of its efficacy against ‘morning sickness’. Complement. Ther. Clin. Pract. 2006, 12, 27–33. [Google Scholar] [CrossRef]
- Volkow, N.D.; Compton, W.M.; Wargo, E.M. The risks of marijuana use during pregnancy. JAMA 2017, 317, 129–130. [Google Scholar] [CrossRef]
- Volkow, N.D.; Han, B.; Compton, W.M.; McCance-Katz, E.F. Self-reported medical and nonmedical cannabis use among pregnant women in the United States. JAMA 2019, 322, 167–169. [Google Scholar] [CrossRef]
- Metz, T.D.; Borgelt, L.M. Marijuana use in pregnancy and while breastfeeding. Obstet. Gynecol. 2018, 132, 1198–1210. [Google Scholar] [CrossRef]
- Moore, D.G.; Turner, J.D.; Parrott, A.C.; Goodwin, J.E.; Fulton, S.E.; Min, M.O.; Fox, H.C.; Braddick, F.M.; Axelsson, E.L.; Lynch, S.; et al. During pregnancy, recreational drug-using women stop taking ecstasy (3,4-methylenedioxy-N-methylamphetamine) and reduce alcohol consumption, but continue to smoke tobacco and cannabis: Initial findings from the Development and Infancy Study. J. Psychopharmacol. 2010, 24, 1403–1410. [Google Scholar] [CrossRef] [Green Version]
- Corsi, D.J.; Hsu, H.; Weiss, D.; Fell, D.B.; Walker, M. Trends and correlates of cannabis use in pregnancy: A population-based study in Ontario, Canada from 2012 to 2017. Can J. Public Health 2019, 110, 76–84. [Google Scholar] [CrossRef]
- Substance Abuse and Mental Health Services Administration. 2019 National Survey on Drug Use and Health: Women. Available online: https://www.samhsa.gov/data/report/2019-nsduh-women (accessed on 5 June 2021).
- Leemaqz, S.Y.; Dekker, G.A.; McCowan, L.M.; Kenny, L.C.; Myers, J.E.; Simpson, N.A.; Poston, L.; Roberts, C.T.; Consortium, S. Maternal marijuana use has independent effects on risk for spontaneous preterm birth but not other common late pregnancy complications. Reprod. Toxicol. 2016, 62, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Lozano, J.; Garcia-Algar, O.; Marchei, E.; Vall, O.; Monleon, T.; Giovannandrea, R.D.; Pichini, S. Prevalence of gestational exposure to cannabis in a Mediterranean city by meconium analysis. Acta Paediatr. 2007, 96, 1734–1737. [Google Scholar] [CrossRef] [PubMed]
- Metz, T.D.; Silver, R.M.; McMillin, G.A.; Allshouse, A.A.; Jensen, T.L.; Mansfield, C.; Heard, K.; Kinney, G.L.; Wymore, E.; Binswanger, I.A. Prenatal marijuana use by self-report and umbilical cord sampling in a state with marijuana legalization. Obstet. Gynecol. 2019, 133, 98–104. [Google Scholar] [CrossRef]
- PregnancyInfo. Available online: https://www.pregnancyinfo.ca/learn-more/ (accessed on 24 June 2021).
- Vastis, V.; Vincent, S.; Metz, T.D.; Shea, A.K. Are canadian cannabis dispensaries counselling pregnant women appropriately? J. Obstet. Gynaecol. Can. 2021, 43, 506–510. [Google Scholar] [CrossRef]
- Whiting, P.F.; Wolff, R.F.; Deshpande, S.; Di Nisio, M.; Duffy, S.; Hernandez, A.V.; Keurentjes, J.C.; Lang, S.; Misso, K.; Ryder, S.; et al. Cannabinoids for Medical Use: A Systematic Review and Meta-analysis. JAMA 2015, 313, 2456–2473. [Google Scholar] [CrossRef]
- Sarrafpour, S.; Urits, I.; Powell, J.; Nguyen, D.; Callan, J.; Orhurhu, V.; Simopoulos, T.; Viswanath, O.; Kaye, A.D.; Kaye, R.J.; et al. Considerations and implications of cannabidiol use during pregnancy. Curr. Pain Headache Rep. 2020, 24, 38. [Google Scholar] [CrossRef] [PubMed]
- Appendino, G.; Chianese, G.; Taglialatela-Scafati, O. Cannabinoids: Occurrence and medicinal chemistry. Curr. Med. Chem. 2011, 18, 1085–1099. [Google Scholar] [CrossRef] [PubMed]
- Elsohly, M.A.; Slade, D. Chemical constituents of marijuana: The complex mixture of natural cannabinoids. Life Sci. 2005, 78, 539–548. [Google Scholar] [CrossRef] [PubMed]
- Dewey, W.L. Cannabinoid pharmacology. Pharmacol. Rev. 1986, 38, 151–178. [Google Scholar] [CrossRef]
- Gaoni, Y.; Mechoulam, R. Isolation, structure, and partial synthesis of an active constituent of hashish. J. Am. Chem. Soc. 1964, 86, 1646–1647. [Google Scholar] [CrossRef]
- Hollister, L.E. Health aspects of cannabis. Pharmacol. Rev. 1986, 38, 1–20. [Google Scholar] [CrossRef]
- Burgdorf, J.R.; Kilmer, B.; Pacula, R.L. Heterogeneity in the composition of marijuana seized in California. Drug Alcohol Depend. 2011, 117, 59–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ElSohly, M.A.; Mehmedic, Z.; Foster, S.; Gon, C.; Chandra, S.; Church, J.C. Changes in cannabis potency over the last 2 decades (1995–2014): Analysis of current data in the United States. Biol. Psychiatry 2016, 79, 613–619. [Google Scholar] [CrossRef] [Green Version]
- ElSohly, M.A.; Ross, S.A.; Mehmedic, Z.; Arafat, R.; Yi, B.; Banahan, B.F. Potency trends of delta9-THC and other cannabinoids in confiscated marijuana from 1980–1997. J. Forensic Sci. 2000, 45, 24–30. [Google Scholar] [CrossRef] [Green Version]
- Mammen, G.; de Freitas, L.; Rehm, J.; Rueda, S. Cannabinoid concentrations in Canada’s regulated medical cannabis industry. Addiction 2017, 112, 730–732. [Google Scholar] [CrossRef]
- Mehmedic, Z.; Chandra, S.; Slade, D.; Denham, H.; Foster, S.; Patel, A.S.; Ross, S.A.; Khan, I.A.; ElSohly, M.A. Potency trends of Delta9-THC and other cannabinoids in confiscated cannabis preparations from 1993 to 2008. J. Forensic Sci. 2010, 55, 1209–1217. [Google Scholar] [CrossRef]
- Health Canada. About Cannabis. Available online: https://www.canada.ca/en/health-canada/services/drugs-medication/cannabis/about.html (accessed on 5 June 2021).
- Fonseca, B.M.; Costa, M.A.; Almada, M.; Correia-da-Silva, G.; Teixeira, N.A. Endogenous cannabinoids revisited: A biochemistry perspective. Prostaglandins Other Lipid Mediat. 2013, 102–103, 13–30. [Google Scholar] [CrossRef]
- Komorowski, J.; Stepien, H. The role of the endocannabinoid system in the regulation of endocrine function and in the control of energy balance in humans. Postepy Hig. Med. Dosw. (Online) 2007, 61, 99–105. [Google Scholar]
- Habayeb, O.M.; Taylor, A.H.; Bell, S.C.; Taylor, D.J.; Konje, J.C. Expression of the endocannabinoid system in human first trimester placenta and its role in trophoblast proliferation. Endocrinology 2008, 149, 5052–5060. [Google Scholar] [CrossRef] [Green Version]
- Lewis, S.E.; Rapino, C.; Di Tommaso, M.; Pucci, M.; Battista, N.; Paro, R.; Simon, L.; Lutton, D.; Maccarrone, M. Differences in the endocannabinoid system of sperm from fertile and infertile men. PLoS ONE 2012, 7, e47704. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Dey, S.K. Endocannabinoid signaling in female reproduction. ACS Chem. Neurosci. 2012, 3, 349–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, A.H.; Ang, C.; Bell, S.C.; Konje, J.C. The role of the endocannabinoid system in gametogenesis, implantation and early pregnancy. Hum. Reprod. Update 2007, 13, 501–513. [Google Scholar] [CrossRef]
- Singh, S.; Filion, K.B.; Abenhaim, H.A.; Eisenberg, M.J. Prevalence and outcomes of prenatal recreational cannabis use in high-income countries: A scoping review. BJOG 2020, 127, 8–16. [Google Scholar] [CrossRef]
- Luke, S.; Hutcheon, J.; Kendall, T. Cannabis use in pregnancy in British Columbia and selected birth outcomes. J. Obstet. Gynaecol. Can. 2019, 41, 1311–1317. [Google Scholar] [CrossRef] [PubMed]
- Varner, M.W.; Silver, R.M.; Rowland Hogue, C.J.; Willinger, M.; Parker, C.B.; Thorsten, V.R.; Goldenberg, R.L.; Saade, G.R.; Dudley, D.J.; Coustan, D.; et al. Association between stillbirth and illicit drug use and smoking during pregnancy. Obstet. Gynecol. 2014, 123, 113–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatch, E.E.; Bracken, M.B. Effect of marijuana use in pregnancy on fetal growth. Am. J. Epidemiol. 1986, 124, 986–993. [Google Scholar] [CrossRef]
- Hayatbakhsh, M.R.; Flenady, V.J.; Gibbons, K.S.; Kingsbury, A.M.; Hurrion, E.; Mamun, A.A.; Najman, J.M. Birth outcomes associated with cannabis use before and during pregnancy. Pediatr. Res. 2012, 71, 215–219. [Google Scholar] [CrossRef] [Green Version]
- Metz, T.D.; Stickrath, E.H. Marijuana use in pregnancy and lactation: A review of the evidence. Am. J. Obstet. Gynecol. 2015, 213, 761–778. [Google Scholar] [CrossRef] [PubMed]
- Hurd, Y.L.; Wang, X.; Anderson, V.; Beck, O.; Minkoff, H.; Dow-Edwards, D. Marijuana impairs growth in mid-gestation fetuses. Neurotoxicol. Teratol. 2005, 27, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Warshak, C.R.; Regan, J.; Moore, B.; Magner, K.; Kritzer, S.; Van Hook, J. Association between marijuana use and adverse obstetrical and neonatal outcomes. J. Perinatol. 2015, 35, 991–995. [Google Scholar] [CrossRef] [PubMed]
- Campbell, E.E.; Gilliland, J.; Dworatzek, P.D.N.; De Vrijer, B.; Penava, D.; Seabrook, J.A. Socioeconomic status and adverse birth outcomes: A population-based Canadian sample. J. Biosoc. Sci. 2018, 50, 102–113. [Google Scholar] [CrossRef] [Green Version]
- El Marroun, H.; Tiemeier, H.; Steegers, E.A.; Jaddoe, V.W.; Hofman, A.; Verhulst, F.C.; van den Brink, W.; Huizink, A.C. Intrauterine cannabis exposure affects fetal growth trajectories: The Generation R Study. J. Am. Acad. Child Adolesc. Psychiatry 2009, 48, 1173–1181. [Google Scholar] [CrossRef] [Green Version]
- Gabrhelik, R.; Mahic, M.; Lund, I.O.; Bramness, J.; Selmer, R.; Skovlund, E.; Handal, M.; Skurtveit, S. Cannabis use during pregnancy and risk of adverse birth outcomes: A longitudinal cohort study. Eur. Addict. Res. 2021, 27, 131–141. [Google Scholar] [CrossRef]
- Gunn, J.K.; Rosales, C.B.; Center, K.E.; Nunez, A.; Gibson, S.J.; Christ, C.; Ehiri, J.E. Prenatal exposure to cannabis and maternal and child health outcomes: A systematic review and meta-analysis. BMJ Open 2016, 6, e009986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conner, S.N.; Carter, E.B.; Tuuli, M.G.; Macones, G.A.; Cahill, A.G. Maternal marijuana use and neonatal morbidity. Am. J. Obstet. Gynecol. 2015, 213, e421–e424. [Google Scholar] [CrossRef] [PubMed]
- Mark, K.; Desai, A.; Terplan, M. Marijuana use and pregnancy: Prevalence, associated characteristics, and birth outcomes. Arch. Womens Ment. Health 2016, 19, 105–111. [Google Scholar] [CrossRef] [PubMed]
- van Gelder, M.M.; Reefhuis, J.; Caton, A.R.; Werler, M.M.; Druschel, C.M.; Roeleveld, N.; National Birth Defects Prevention Study. Characteristics of pregnant illicit drug users and associations between cannabis use and perinatal outcome in a population-based study. Drug Alcohol Depend. 2010, 109, 243–247. [Google Scholar] [CrossRef] [Green Version]
- Government of Canada. Canadian Cannabis Survey 2020: Summary. Available online: https://www.canada.ca/en/health-canada/services/drugs-medication/cannabis/research-data/canadian-cannabis-survey-2020-summary.html (accessed on 24 July 2021).
- Abel, E.L.; Tan, S.E.; Subramanian, M. Effects of delta 9-tetrahydrocannabinol, phenobarbital, and their combination on pregnancy and offspring in rats. Teratology 1987, 36, 193–198. [Google Scholar] [CrossRef]
- Wenger, T.; Fragkakis, G.; Giannikou, P.; Yiannikakis, N. The effects of prenatally administered endogenous cannabinoid on rat offspring. Pharmacol. Biochem. Behav. 1997, 58, 537–544. [Google Scholar] [CrossRef]
- Benevenuto, S.G.; Domenico, M.D.; Martins, M.A.; Costa, N.S.; de Souza, A.R.; Costa, J.L.; Tavares, M.F.; Dolhnikoff, M.; Veras, M.M. Recreational use of marijuana during pregnancy and negative gestational and fetal outcomes: An experimental study in mice. Toxicology 2017, 376, 94–101. [Google Scholar] [CrossRef]
- Gillies, R.; Lee, K.; Vanin, S.; Laviolette, S.R.; Holloway, A.C.; Arany, E.; Hardy, D.B. Maternal exposure to Delta9-tetrahydrocannabinol impairs female offspring glucose homeostasis and endocrine pancreatic development in the rat. Reprod. Toxicol. 2020, 94, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Natale, B.V.; Gustin, K.N.; Lee, K.; Holloway, A.C.; Laviolette, S.R.; Natale, D.R.C.; Hardy, D.B. Delta9-tetrahydrocannabinol exposure during rat pregnancy leads to symmetrical fetal growth restriction and labyrinth-specific vascular defects in the placenta. Sci. Rep. 2020, 10, 544. [Google Scholar] [CrossRef]
- Richardson, K.A.; Hester, A.K.; McLemore, G.L. Prenatal cannabis exposure—The “first hit” to the endocannabinoid system. Neurotoxicol. Teratol. 2016, 58, 5–14. [Google Scholar] [CrossRef] [PubMed]
- Bellocchio, L.; Cervino, C.; Pasquali, R.; Pagotto, U. The endocannabinoid system and energy metabolism. J. Neuroendocrinol. 2008, 20, 850–857. [Google Scholar] [CrossRef]
- Burstein, S.H.; Zurier, R.B. Cannabinoids, endocannabinoids, and related analogs in inflammation. AAPS J. 2009, 11, 109–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomes, T.M.; Dias da Silva, D.; Carmo, H.; Carvalho, F.; Silva, J.P. Epigenetics and the endocannabinoid system signaling: An intricate interplay modulating neurodevelopment. Pharmacol. Res. 2020, 162, 105237. [Google Scholar] [CrossRef]
- Walker, O.S.; Holloway, A.C.; Raha, S. The role of the endocannabinoid system in female reproductive tissues. J. Ovarian Res. 2019, 12, 3. [Google Scholar] [CrossRef]
- Woodhams, S.G.; Sagar, D.R.; Burston, J.J.; Chapman, V. The role of the endocannabinoid system in pain. Handb. Exp. Pharmacol. 2015, 227, 119–143. [Google Scholar] [CrossRef]
- Lu, H.C.; Mackie, K. An introduction to the endogenous cannabinoid system. Biol. Psychiatry 2016, 79, 516–525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Battista, N.; Di Tommaso, M.; Bari, M.; Maccarrone, M. The endocannabinoid system: An overview. Front. Behav. Neurosci. 2012, 6, 9. [Google Scholar] [CrossRef] [Green Version]
- Devane, W.A.; Hanus, L.; Breuer, A.; Pertwee, R.G.; Stevenson, L.A.; Griffin, G.; Gibson, D.; Mandelbaum, A.; Etinger, A.; Mechoulam, R. Isolation and structure of a brain constituent that binds to the cannabinoid receptor. Science 1992, 258, 1946–1949. [Google Scholar] [CrossRef] [PubMed]
- Mechoulam, R.; Ben-Shabat, S.; Hanus, L.; Ligumsky, M.; Kaminski, N.E.; Schatz, A.R.; Gopher, A.; Almog, S.; Martin, B.R.; Compton, D.R.; et al. Identification of an endogenous 2-monoglyceride, present in canine gut, that binds to cannabinoid receptors. Biochem. Pharmacol. 1995, 50, 83–90. [Google Scholar] [CrossRef]
- Rodriguez de Fonseca, F.; Del Arco, I.; Bermudez-Silva, F.J.; Bilbao, A.; Cippitelli, A.; Navarro, M. The endocannabinoid system: Physiology and pharmacology. Alcohol Alcohol. 2005, 40, 2–14. [Google Scholar] [CrossRef]
- Pertwee, R.G.; Howlett, A.C.; Abood, M.E.; Alexander, S.P.; Di Marzo, V.; Elphick, M.R.; Greasley, P.J.; Hansen, H.S.; Kunos, G.; Mackie, K.; et al. International Union of Basic and Clinical Pharmacology. LXXIX. Cannabinoid receptors and their ligands: Beyond CB(1) and CB(2). Pharmacol. Rev. 2010, 62, 588–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maccarrone, M.; Dainese, E.; Oddi, S. Intracellular trafficking of anandamide: New concepts for signaling. Trends Biochem. Sci. 2010, 35, 601–608. [Google Scholar] [CrossRef] [PubMed]
- Oddi, S.; Fezza, F.; Pasquariello, N.; De Simone, C.; Rapino, C.; Dainese, E.; Finazzi-Agro, A.; Maccarrone, M. Evidence for the intracellular accumulation of anandamide in adiposomes. Cell. Mol. Life Sci. 2008, 65, 840–850. [Google Scholar] [CrossRef]
- Lucas, C.J.; Galettis, P.; Schneider, J. The pharmacokinetics and the pharmacodynamics of cannabinoids. Br. J. Clin. Pharmacol. 2018, 84, 2477–2482. [Google Scholar] [CrossRef] [Green Version]
- Breijyeh, Z.; Jubeh, B.; Bufo, S.A.; Karaman, R.; Scrano, L. Cannabis: A toxin-producing plant with potential therapeutic uses. Toxins 2021, 13, 117. [Google Scholar] [CrossRef]
- Kennedy, E.K.C.; Perono, G.A.; Nemez, D.B.; Holloway, A.C.; Thomas, P.J.; Letcher, R.; Marvin, C.; Stetefeld, J.; Stout, J.; Peters, O.; et al. Increasing cannabis use and importance as an environmental contaminant mixture and associated risks to exposed biota: A review. Crit. Rev. Environ. Sci. Technol. 2020, 1–37. [Google Scholar] [CrossRef]
- Laprairie, R.B.; Bagher, A.M.; Kelly, M.E.; Denovan-Wright, E.M. Cannabidiol is a negative allosteric modulator of the cannabinoid CB1 receptor. Br. J. Pharmacol. 2015, 172, 4790–4805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ujvary, I.; Hanus, L. Human metabolites of cannabidiol: A review on their formation, biological activity, and relevance in therapy. Cannabis Cannabinoid Res. 2016, 1, 90–101. [Google Scholar] [CrossRef] [Green Version]
- Hourani, W.; Alexander, S.P.H. Cannabinoid ligands, receptors and enzymes: Pharmacological tools and therapeutic potential. Brain Neurosci. Adv. 2018, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pertwee, R.G. Pharmacology of cannabinoid CB1 and CB2 receptors. Pharmacol. Ther. 1997, 74, 129–180. [Google Scholar] [CrossRef]
- Chakravarti, B.; Ravi, J.; Ganju, R.K. Cannabinoids as therapeutic agents in cancer: Current status and future implications. Oncotarget 2014, 5, 5852–5872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrera, B.; Carracedo, A.; Diez-Zaera, M.; Gomez del Pulgar, T.; Guzman, M.; Velasco, G. The CB2 cannabinoid receptor signals apoptosis via ceramide-dependent activation of the mitochondrial intrinsic pathway. Exp. Cell Res. 2006, 312, 2121–2131. [Google Scholar] [CrossRef]
- Pacher, P.; Batkai, S.; Kunos, G. The endocannabinoid system as an emerging target of pharmacotherapy. Pharmacol. Rev. 2006, 58, 389–462. [Google Scholar] [CrossRef] [Green Version]
- Zou, S.; Kumar, U. Cannabinoid receptors and the endocannabinoid system: Signaling and function in the central nervous system. Int. J. Mol. Sci 2018, 19, 833. [Google Scholar] [CrossRef] [Green Version]
- Ryberg, E.; Larsson, N.; Sjogren, S.; Hjorth, S.; Hermansson, N.O.; Leonova, J.; Elebring, T.; Nilsson, K.; Drmota, T.; Greasley, P.J. The orphan receptor GPR55 is a novel cannabinoid receptor. Br. J. Pharmacol. 2007, 152, 1092–1101. [Google Scholar] [CrossRef]
- Sawzdargo, M.; Nguyen, T.; Lee, D.K.; Lynch, K.R.; Cheng, R.; Heng, H.H.; George, S.R.; O’Dowd, B.F. Identification and cloning of three novel human G protein-coupled receptor genes GPR52, PsiGPR53 and GPR55: GPR55 is extensively expressed in human brain. Mol. Brain Res. 1999, 64, 193–198. [Google Scholar] [CrossRef]
- Lauckner, J.E.; Jensen, J.B.; Chen, H.Y.; Lu, H.C.; Hille, B.; Mackie, K. GPR55 is a cannabinoid receptor that increases intracellular calcium and inhibits M current. Proc. Natl. Acad. Sci. USA 2008, 105, 2699–2704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marichal-Cancino, B.A.; Fajardo-Valdez, A.; Ruiz-Contreras, A.E.; Mendez-Diaz, M.; Prospero-Garcia, O. Advances in the physiology of GPR55 in the central nervous system. Curr. Neuropharmacol. 2017, 15, 771–778. [Google Scholar] [CrossRef] [Green Version]
- Romero-Zerbo, S.Y.; Rafacho, A.; Diaz-Arteaga, A.; Suarez, J.; Quesada, I.; Imbernon, M.; Ross, R.A.; Dieguez, C.; Rodriguez de Fonseca, F.; Nogueiras, R.; et al. A role for the putative cannabinoid receptor GPR55 in the islets of Langerhans. J. Endocrinol. 2011, 211, 177–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simcocks, A.C.; O′Keefe, L.; Jenkin, K.A.; Mathai, M.L.; Hryciw, D.H.; McAinch, A.J. A potential role for GPR55 in the regulation of energy homeostasis. Drug Discov. Today 2014, 19, 1145–1151. [Google Scholar] [CrossRef] [PubMed]
- Lowin, T.; Straub, R.H. Cannabinoid-based drugs targeting CB1 and TRPV1, the sympathetic nervous system, and arthritis. Arthritis Res. Ther. 2015, 17, 226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrosino, S.; Schiano Moriello, A.; Cerrato, S.; Fusco, M.; Puigdemont, A.; De Petrocellis, L.; Di Marzo, V. The anti-inflammatory mediator palmitoylethanolamide enhances the levels of 2-arachidonoyl-glycerol and potentiates its actions at TRPV1 cation channels. Br. J. Pharmacol. 2016, 173, 1154–1162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ross, R.A. Anandamide and vanilloid TRPV1 receptors. Br. J. Pharmacol. 2003, 140, 790–801. [Google Scholar] [CrossRef] [Green Version]
- Ahluwalia, J.; Urban, L.; Bevan, S.; Nagy, I. Anandamide regulates neuropeptide release from capsaicin-sensitive primary sensory neurons by activating both the cannabinoid 1 receptor and the vanilloid receptor 1 In Vitro. Eur. J. Neurosci. 2003, 17, 2611–2618. [Google Scholar] [CrossRef]
- Anand, U.; Otto, W.R.; Sanchez-Herrera, D.; Facer, P.; Yiangou, Y.; Korchev, Y.; Birch, R.; Benham, C.; Bountra, C.; Chessell, I.P.; et al. Cannabinoid receptor CB2 localisation and agonist-mediated inhibition of capsaicin responses in human sensory neurons. Pain 2008, 138, 667–680. [Google Scholar] [CrossRef] [PubMed]
- Pistis, M.; Melis, M. From surface to nuclear receptors: The endocannabinoid family extends its assets. Curr. Med. Chem. 2010, 17, 1450–1467. [Google Scholar] [CrossRef] [PubMed]
- Rouzer, C.A.; Marnett, L.J. Endocannabinoid oxygenation by cyclooxygenases, lipoxygenases, and cytochromes P450: Cross-talk between the eicosanoid and endocannabinoid signaling pathways. Chem. Rev. 2011, 111, 5899–5921. [Google Scholar] [CrossRef] [PubMed]
- Zelasko, S.; Arnold, W.R.; Das, A. Endocannabinoid metabolism by cytochrome P450 monooxygenases. Prostaglandins Other Lipid Mediat. 2015, 116–117, 112–123. [Google Scholar] [CrossRef]
- El-Talatini, M.R.; Taylor, A.H.; Elson, J.C.; Brown, L.; Davidson, A.C.; Konje, J.C. Localisation and function of the endocannabinoid system in the human ovary. PLoS ONE 2009, 4, e4579. [Google Scholar] [CrossRef] [Green Version]
- Schuel, H.; Burkman, L.J.; Lippes, J.; Crickard, K.; Forester, E.; Piomelli, D.; Giuffrida, A. N-Acylethanolamines in human reproductive fluids. Chem. Phys. Lipids 2002, 121, 211–227. [Google Scholar] [CrossRef] [Green Version]
- Paria, B.C.; Das, S.K.; Dey, S.K. The preimplantation mouse embryo is a target for cannabinoid ligand-receptor signaling. Proc. Natl. Acad. Sci. USA 1995, 92, 9460–9464. [Google Scholar] [CrossRef] [Green Version]
- Das, S.K.; Paria, B.C.; Chakraborty, I.; Dey, S.K. Cannabinoid ligand-receptor signaling in the mouse uterus. Proc. Natl. Acad. Sci. USA 1995, 92, 4332–4336. [Google Scholar] [CrossRef] [Green Version]
- Scotchie, J.G.; Savaris, R.F.; Martin, C.E.; Young, S.L. Endocannabinoid regulation in human endometrium across the menstrual cycle. Reprod. Sci. 2015, 22, 113–123. [Google Scholar] [CrossRef] [Green Version]
- Park, B.; Gibbons, H.M.; Mitchell, M.D.; Glassa, M. Identification of the CB1 cannabinoid receptor and fatty acid amide hydrolase (FAAH) in the human placenta. Placenta 2003, 24, 473–478. [Google Scholar] [CrossRef]
- Correa, F.; Wolfson, M.L.; Valchi, P.; Aisemberg, J.; Franchi, A.M. Endocannabinoid system and pregnancy. Reproduction 2016, 152, R191–R200. [Google Scholar] [CrossRef]
- Schmid, P.C.; Paria, B.C.; Krebsbach, R.J.; Schmid, H.H.; Dey, S.K. Changes in anandamide levels in mouse uterus are associated with uterine receptivity for embryo implantation. Proc. Natl. Acad. Sci. USA 1997, 94, 4188–4192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Xie, H.; Guo, Y.; Zhang, H.; Takahashi, T.; Kingsley, P.J.; Marnett, L.J.; Das, S.K.; Cravatt, B.F.; Dey, S.K. Fatty acid amide hydrolase deficiency limits early pregnancy events. J. Clin. Investig. 2006, 116, 2122–2131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, H.; Sun, X.; Piao, Y.; Jegga, A.G.; Handwerger, S.; Ko, M.S.; Dey, S.K. Silencing or amplification of endocannabinoid signaling in blastocysts via CB1 compromises trophoblast cell migration. J. Biol. Chem. 2012, 287, 32288–32297. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.M.; Paria, B.C.; Dey, S.K. Activation of brain-type cannabinoid receptors interferes with preimplantation mouse embryo development. Biol. Reprod. 1996, 55, 756–761. [Google Scholar] [CrossRef]
- Burton, G.J.; Fowden, A.L. The placenta: A multifaceted, transient organ. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2015, 370, 20140066. [Google Scholar] [CrossRef] [Green Version]
- Knofler, M.; Haider, S.; Saleh, L.; Pollheimer, J.; Gamage, T.; James, J. Human placenta and trophoblast development: Key molecular mechanisms and model systems. Cell. Mol. Life Sci. 2019, 76, 3479–3496. [Google Scholar] [CrossRef] [Green Version]
- Costa, M.A. The endocannabinoid system: A novel player in human placentation. Reprod. Toxicol. 2016, 61, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Ezechukwu, H.C.; Diya, C.A.; Shrestha, N.; Hryciw, D.H. Role for endocannabinoids in early pregnancy: Recent advances and the effects of cannabis use. Am. J. Physiol. Endocrinol. Metab. 2020, 319, E557–E561. [Google Scholar] [CrossRef] [PubMed]
- Maia, J.; Fonseca, B.M.; Teixeira, N.; Correia-da-Silva, G. The fundamental role of the endocannabinoid system in endometrium and placenta: Implications in pathophysiological aspects of uterine and pregnancy disorders. Hum. Reprod. Update 2020, 26, 586–602. [Google Scholar] [CrossRef]
- Helliwell, R.J.; Chamley, L.W.; Blake-Palmer, K.; Mitchell, M.D.; Wu, J.; Kearn, C.S.; Glass, M. Characterization of the endocannabinoid system in early human pregnancy. J. Clin. Endocrinol. Metab. 2004, 89, 5168–5174. [Google Scholar] [CrossRef] [Green Version]
- Kenney, S.P.; Kekuda, R.; Prasad, P.D.; Leibach, F.H.; Devoe, L.D.; Ganapathy, V. Cannabinoid receptors and their role in the regulation of the serotonin transporter in human placenta. Am. J. Obstet. Gynecol. 1999, 181, 491–497. [Google Scholar] [CrossRef]
- Aban, C.; Leguizamon, G.F.; Cella, M.; Damiano, A.; Franchi, A.M.; Farina, M.G. Differential expression of endocannabinoid system in normal and preeclamptic placentas: Effects on nitric oxide synthesis. Placenta 2013, 34, 67–74. [Google Scholar] [CrossRef]
- Costa, M.A.; Fonseca, B.M.; Keating, E.; Teixeira, N.A.; Correia-da-Silva, G. 2-arachidonoylglycerol effects in cytotrophoblasts: Metabolic enzymes expression and apoptosis in BeWo cells. Reproduction 2014, 147, 301–311. [Google Scholar] [CrossRef] [PubMed]
- Costa, M.A.; Keating, E.; Fonseca, B.M.; Teixeira, N.A.; Correia-da-Silva, G. 2-Arachidonoylglycerol impairs human cytotrophoblast cells syncytialization: Influence of endocannabinoid signalling in placental development. Mol. Cell Endocrinol. 2015, 399, 386–394. [Google Scholar] [CrossRef]
- Sun, X.; Xie, H.; Yang, J.; Wang, H.; Bradshaw, H.B.; Dey, S.K. Endocannabinoid signaling directs differentiation of trophoblast cell lineages and placentation. Proc. Natl. Acad. Sci. USA 2010, 107, 16887–16892. [Google Scholar] [CrossRef] [Green Version]
- Trabucco, E.; Acone, G.; Marenna, A.; Pierantoni, R.; Cacciola, G.; Chioccarelli, T.; Mackie, K.; Fasano, S.; Colacurci, N.; Meccariello, R.; et al. Endocannabinoid system in first trimester placenta: Low FAAH and high CB1 expression characterize spontaneous miscarriage. Placenta 2009, 30, 516–522. [Google Scholar] [CrossRef]
- Marczylo, T.H.; Lam, P.M.; Amoako, A.A.; Konje, J.C. Anandamide levels in human female reproductive tissues: Solid-phase extraction and measurement by ultraperformance liquid chromatography tandem mass spectrometry. Anal. Biochem. 2010, 400, 155–162. [Google Scholar] [CrossRef]
- Brocato, B.; Zoerner, A.A.; Janjetovic, Z.; Skobowiat, C.; Gupta, S.; Moore, B.M., 2nd; Slominski, A.; Zhang, J.; Schenone, M.; Phinehas, R.; et al. Endocannabinoid crosstalk between placenta and maternal fat in a baboon model (Papio spp.) of obesity. Placenta 2013, 34, 983–989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fonseca, B.M.; Correia-da-Silva, G.; Taylor, A.H.; Lam, P.M.; Marczylo, T.H.; Konje, J.C.; Teixeira, N.A. Characterisation of the endocannabinoid system in rat haemochorial placenta. Reprod. Toxicol. 2012, 34, 347–356. [Google Scholar] [CrossRef] [PubMed]
- Costa, M.A.; Fonseca, B.M.; Keating, E.; Teixeira, N.A.; Correia-da-Silva, G. Transient receptor potential vanilloid 1 is expressed in human cytotrophoblasts: Induction of cell apoptosis and impairment of syncytialization. Int. J. Biochem. Cell Biol. 2014, 57, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Kremshofer, J.; Siwetz, M.; Berghold, V.M.; Lang, I.; Huppertz, B.; Gauster, M. A role for GPR55 in human placental venous endothelial cells. Histochem. Cell Biol. 2015, 144, 49–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aban, C.E.; Accialini, P.L.; Etcheverry, T.; Leguizamon, G.F.; Martinez, N.A.; Farina, M.G. Crosstalk between nitric oxide and endocannabinoid signaling pathways in normal and pathological placentation. Front. Physiol. 2018, 9, 1699. [Google Scholar] [CrossRef]
- Costa, M.A.; Fonseca, B.M.; Teixeira, N.A.; Correia-da-Silva, G. The endocannabinoid anandamide induces apoptosis in cytotrophoblast cells: Involvement of both mitochondrial and death receptor pathways. Placenta 2015, 36, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Almada, M.; Costa, L.; Fonseca, B.; Alves, P.; Braga, J.; Goncalves, D.; Teixeira, N.; Correia-da-Silva, G. The endocannabinoid 2-arachidonoylglycerol promotes endoplasmic reticulum stress in placental cells. Reproduction 2020, 160, 171–180. [Google Scholar] [CrossRef]
- Etcheverry, T.; Accialini, P.; Palligas, M.; Loureiro, F.; Saraco, N.; Martinez, N.; Farina, M. Endocannabinoid signaling impairs syncytialization: Using flow cytometry to evaluate forskolin-induced cell fusion. Placenta 2021, 103, 152–155. [Google Scholar] [CrossRef]
- Szilagyi, J.T.; Composto-Wahler, G.M.; Joseph, L.B.; Wang, B.; Rosen, T.; Laskin, J.D.; Aleksunes, L.M. Anandamide down-regulates placental transporter expression through CB2 receptor-mediated inhibition of cAMP synthesis. Pharmacol. Res. 2019, 141, 331–342. [Google Scholar] [CrossRef] [PubMed]
- Habayeb, O.M.; Taylor, A.H.; Evans, M.D.; Cooke, M.S.; Taylor, D.J.; Bell, S.C.; Konje, J.C. Plasma levels of the endocannabinoid anandamide in women—A potential role in pregnancy maintenance and labor? J. Clin. Endocrinol. Metab. 2004, 89, 5482–5487. [Google Scholar] [CrossRef] [Green Version]
- Maccarrone, M.; Valensise, H.; Bari, M.; Lazzarin, N.; Romanini, C.; Finazzi-Agro, A. Relation between decreased anandamide hydrolase concentrations in human lymphocytes and miscarriage. Lancet 2000, 355, 1326–1329. [Google Scholar] [CrossRef]
- Maccarrone, M.; Bisogno, T.; Valensise, H.; Lazzarin, N.; Fezza, F.; Manna, C.; Di Marzo, V.; Finazzi-Agro, A. Low fatty acid amide hydrolase and high anandamide levels are associated with failure to achieve an ongoing pregnancy after IVF and embryo transfer. Mol. Hum. Reprod. 2002, 8, 188–195. [Google Scholar] [CrossRef]
- Molvarec, A.; Fugedi, G.; Szabo, E.; Stenczer, B.; Walentin, S.; Rigo, J., Jr. Decreased circulating anandamide levels in preeclampsia. Hypertens. Res. 2015, 38, 413–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fugedi, G.; Molnar, M.; Rigo, J., Jr.; Schonleber, J.; Kovalszky, I.; Molvarec, A. Increased placental expression of cannabinoid receptor 1 in preeclampsia: An observational study. BMC Pregnancy Childbirth 2014, 14, 395. [Google Scholar] [CrossRef] [Green Version]
- Thompson, R.; DeJong, K.; Lo, J. Marijuana use in pregnancy: A review. Obstet. Gynecol. Surv. 2019, 74, 415–428. [Google Scholar] [CrossRef]
- Bailey, J.R.; Cunny, H.C.; Paule, M.G.; Slikker, W., Jr. Fetal disposition of delta 9-tetrahydrocannabinol (THC) during late pregnancy in the rhesus monkey. Toxicol. Appl. Pharmacol. 1987, 90, 315–321. [Google Scholar] [CrossRef]
- Blackard, C.; Tennes, K. Human placental transfer of cannabinoids. N. Engl. J. Med. 1984, 311, 797. [Google Scholar] [CrossRef]
- Hutchings, D.E.; Martin, B.R.; Gamagaris, Z.; Miller, N.; Fico, T. Plasma concentrations of delta-9-tetrahydrocannabinol in dams and fetuses following acute or multiple prenatal dosing in rats. Life Sci. 1989, 44, 697–701. [Google Scholar] [CrossRef]
- Khare, M.; Taylor, A.H.; Konje, J.C.; Bell, S.C. Delta9-tetrahydrocannabinol inhibits cytotrophoblast cell proliferation and modulates gene transcription. Mol. Hum. Reprod. 2006, 12, 321–333. [Google Scholar] [CrossRef] [Green Version]
- Walker, O.S.; Ragos, R.; Gurm, H.; Lapierre, M.; May, L.L.; Raha, S. Delta-9-tetrahydrocannabinol disrupts mitochondrial function and attenuates syncytialization in human placental BeWo cells. Physiol. Rep. 2020, 8, e14476. [Google Scholar] [CrossRef]
- Costa, M.A.; Fonseca, B.M.; Marques, F.; Teixeira, N.A.; Correia-da-Silva, G. The psychoactive compound of Cannabis sativa, Delta(9)-tetrahydrocannabinol (THC) inhibits the human trophoblast cell turnover. Toxicology 2015, 334, 94–103. [Google Scholar] [CrossRef]
- Almada, M.; Alves, P.; Fonseca, B.M.; Carvalho, F.; Queiros, C.R.; Gaspar, H.; Amaral, C.; Teixeira, N.A.; Correia-da-Silva, G. Synthetic cannabinoids JWH-018, JWH-122, UR-144 and the phytocannabinoid THC activate apoptosis in placental cells. Toxicol. Lett. 2020, 319, 129–137. [Google Scholar] [CrossRef]
- Almada, M.; Costa, L.; Fonseca, B.M.; Amaral, C.; Teixeira, N.; Correia-da-Silva, G. The synthetic cannabinoid WIN-55,212 induced-apoptosis in cytotrophoblasts cells by a mechanism dependent on CB1 receptor. Toxicology 2017, 385, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Lojpur, T.; Easton, Z.; Raez-Villanueva, S.; Laviolette, S.; Holloway, A.C.; Hardy, D.B. Delta9-Tetrahydrocannabinol leads to endoplasmic reticulum stress and mitochondrial dysfunction in human BeWo trophoblasts. Reprod. Toxicol. 2019, 87, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Walker, O.S.; Gurm, H.; Sharma, R.; Verma, N.; May, L.L.; Raha, S. Delta-9-tetrahydrocannabinol inhibits invasion of HTR8/SVneo human extravillous trophoblast cells and negatively impacts mitochondrial function. Sci. Rep. 2021, 11, 4029. [Google Scholar] [CrossRef]
- Araujo, J.R.; Goncalves, P.; Martel, F. Effect of cannabinoids upon the uptake of folic acid by BeWo cells. Pharmacology 2009, 83, 170–176. [Google Scholar] [CrossRef]
- Keating, E.; Goncalves, P.; Campos, I.; Costa, F.; Martel, F. Folic acid uptake by the human syncytiotrophoblast: Interference by pharmacotherapy, drugs of abuse and pathological conditions. Reprod. Toxicol. 2009, 28, 511–520. [Google Scholar] [CrossRef]
- Almada, M.; Amaral, C.; Oliveira, A.; Fernandes, P.A.; Ramos, M.J.; Fonseca, B.M.; Correia-da-Silva, G.; Teixeira, N. Cannabidiol (CBD) but not tetrahydrocannabinol (THC) dysregulate in vitro decidualization of human endometrial stromal cells by disruption of estrogen signaling. Reprod. Toxicol. 2020, 93, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Feinshtein, V.; Erez, O.; Ben-Zvi, Z.; Eshkoli, T.; Sheizaf, B.; Sheiner, E.; Holcberg, G. Cannabidiol enhances xenobiotic permeability through the human placental barrier by direct inhibition of breast cancer resistance protein: An ex vivo study. Am. J. Obstet. Gynecol. 2013, 209, 573.e1–573.e15. [Google Scholar] [CrossRef]
- Costa, M.A. The endocrine function of human placenta: An overview. Reprod. Biomed. Online 2016, 32, 14–43. [Google Scholar] [CrossRef] [Green Version]
- Berkane, N.; Liere, P.; Oudinet, J.P.; Hertig, A.; Lefevre, G.; Pluchino, N.; Schumacher, M.; Chabbert-Buffet, N. From pregnancy to preeclampsia: A key role for estrogens. Endocr. Rev. 2017, 38, 123–144. [Google Scholar] [CrossRef]
- Maia, J.; Almada, M.; Midao, L.; Fonseca, B.M.; Braga, J.; Goncalves, D.; Teixeira, N.; Correia-da-Silva, G. The cannabinoid delta-9-tetrahydrocannabinol disrupts estrogen signaling in human placenta. Toxicol. Sci. 2020, 177, 420–430. [Google Scholar] [CrossRef]
- Maia, J.; Midao, L.; Cunha, S.C.; Almada, M.; Fonseca, B.M.; Braga, J.; Goncalves, D.; Teixeira, N.; Correia-da-Silva, G. Effects of cannabis tetrahydrocannabinol on endocannabinoid homeostasis in human placenta. Arch. Toxicol. 2019, 93, 649–658. [Google Scholar] [CrossRef] [Green Version]
- Maia, J.; Fonseca, B.M.; Cunha, S.C.; Braga, J.; Goncalves, D.; Teixeira, N.; Correia-da-Silva, G. Impact of tetrahydrocannabinol on the endocannabinoid 2-arachidonoylglycerol metabolism: ABHD6 and ABHD12 as novel players in human placenta. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2020, 1865, 158807. [Google Scholar] [CrossRef]
- Maccarrone, M.; Guzman, M.; Mackie, K.; Doherty, P.; Harkany, T. Programming of neural cells by (endo)cannabinoids: From physiological rules to emerging therapies. Nat. Rev. Neurosci. 2014, 15, 786–801. [Google Scholar] [CrossRef] [Green Version]
- Malenczyk, K.; Keimpema, E.; Piscitelli, F.; Calvigioni, D.; Bjorklund, P.; Mackie, K.; Di Marzo, V.; Hokfelt, T.G.; Dobrzyn, A.; Harkany, T. Fetal endocannabinoids orchestrate the organization of pancreatic islet microarchitecture. Proc. Natl. Acad. Sci. USA 2015, 112, E6185–E6194. [Google Scholar] [CrossRef] [Green Version]
- Oltrabella, F.; Melgoza, A.; Nguyen, B.; Guo, S. Role of the endocannabinoid system in vertebrates: Emphasis on the zebrafish model. Dev. Growth Differ. 2017, 59, 194–210. [Google Scholar] [CrossRef]
- Paria, B.C.; Deutsch, D.D.; Dey, S.K. The uterus is a potential site for anandamide synthesis and hydrolysis: Differential profiles of anandamide synthase and hydrolase activities in the mouse uterus during the periimplantation period. Mol. Reprod. Dev. 1996, 45, 183–192. [Google Scholar] [CrossRef]
- Paria, B.C.; Song, H.; Wang, X.; Schmid, P.C.; Krebsbach, R.J.; Schmid, H.H.; Bonner, T.I.; Zimmer, A.; Dey, S.K. Dysregulated cannabinoid signaling disrupts uterine receptivity for embryo implantation. J. Biol. Chem. 2001, 276, 20523–20528. [Google Scholar] [CrossRef] [Green Version]
- Berrendero, F.; Sepe, N.; Ramos, J.A.; Di Marzo, V.; Fernandez-Ruiz, J.J. Analysis of cannabinoid receptor binding and mRNA expression and endogenous cannabinoid contents in the developing rat brain during late gestation and early postnatal period. Synapse 1999, 33, 181–191. [Google Scholar] [CrossRef]
- Maccarrone, M.; Finazzi-Agro, A. Anandamide hydrolase: A guardian angel of human reproduction? Trends Pharmacol. Sci. 2004, 25, 353–357. [Google Scholar] [CrossRef]
- Wang, J.; Paria, B.C.; Dey, S.K.; Armant, D.R. Stage-specific excitation of cannabinoid receptor exhibits differential effects on mouse embryonic development. Biol. Reprod. 1999, 60, 839–844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barker, D.J. In utero programming of chronic disease. Clin. Sci. (Lond.) 1998, 95, 115–128. [Google Scholar] [CrossRef] [PubMed]
- Bernard, C.; Milh, M.; Morozov, Y.M.; Ben-Ari, Y.; Freund, T.F.; Gozlan, H. Altering cannabinoid signaling during development disrupts neuronal activity. Proc. Natl. Acad. Sci. USA 2005, 102, 9388–9393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calvigioni, D.; Hurd, Y.L.; Harkany, T.; Keimpema, E. Neuronal substrates and functional consequences of prenatal cannabis exposure. Eur. Child Adolesc. Psychiatry 2014, 23, 931–941. [Google Scholar] [CrossRef] [Green Version]
- Fried, P.A.; Smith, A.M. A literature review of the consequences of prenatal marihuana exposure. An emerging theme of a deficiency in aspects of executive function. Neurotoxicol. Teratol. 2001, 23, 1–11. [Google Scholar] [CrossRef]
- Navarro, M.; Rubio, P.; de Fonseca, F.R. Behavioural consequences of maternal exposure to natural cannabinoids in rats. Psychopharmacology 1995, 122, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Schneider, M. Cannabis use in pregnancy and early life and its consequences: Animal models. Eur. Arch. Psychiatry Clin. Neurosci. 2009, 259, 383–393. [Google Scholar] [CrossRef]
- Dong, C.; Chen, J.; Harrington, A.; Vinod, K.Y.; Hegde, M.L.; Hegde, V.L. Cannabinoid exposure during pregnancy and its impact on immune function. Cell. Mol. Life Sci. 2019, 76, 729–743. [Google Scholar] [CrossRef]
- Zumbrun, E.E.; Sido, J.M.; Nagarkatti, P.S.; Nagarkatti, M. Epigenetic regulation of immunological alterations following prenatal exposure to marijuana cannabinoids and its long term consequences in offspring. J. Neuroimmune Pharmacol. 2015, 10, 245–254. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.; Laviolette, S.R.; Hardy, D.B. Exposure to Delta9-tetrahydrocannabinol during rat pregnancy leads to impaired cardiac dysfunction in postnatal life. Pediatr. Res. 2021. [Google Scholar] [CrossRef] [PubMed]
- Fride, E.; Gobshtis, N.; Dahan, H.; Weller, A.; Giuffrida, A.; Ben-Shabat, S. Chapter 6 The endocannabinoid system during development: Emphasis on perinatal events and delayed effects. Vitam. Horm. 2009, 81, 139–158. [Google Scholar] [CrossRef]
- Harkany, T.; Keimpema, E.; Barabas, K.; Mulder, J. Endocannabinoid functions controlling neuronal specification during brain development. Mol. Cell Endocrinol. 2008, 286, S84–S90. [Google Scholar] [CrossRef] [Green Version]
- Jutras-Aswad, D.; DiNieri, J.A.; Harkany, T.; Hurd, Y.L. Neurobiological consequences of maternal cannabis on human fetal development and its neuropsychiatric outcome. Eur. Arch. Psychiatry Clin. Neurosci. 2009, 259, 395–412. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Ruiz, J.; Gomez, M.; Hernandez, M.; de Miguel, R.; Ramos, J.A. Cannabinoids and gene expression during brain development. Neurotox. Res. 2004, 6, 389–401. [Google Scholar] [CrossRef]
- Mulder, J.; Aguado, T.; Keimpema, E.; Barabas, K.; Ballester Rosado, C.J.; Nguyen, L.; Monory, K.; Marsicano, G.; Di Marzo, V.; Hurd, Y.L.; et al. Endocannabinoid signaling controls pyramidal cell specification and long-range axon patterning. Proc. Natl. Acad. Sci. USA 2008, 105, 8760–8765. [Google Scholar] [CrossRef] [Green Version]
- Galve-Roperh, I.; Chiurchiu, V.; Diaz-Alonso, J.; Bari, M.; Guzman, M.; Maccarrone, M. Cannabinoid receptor signaling in progenitor/stem cell proliferation and differentiation. Prog. Lipid Res. 2013, 52, 633–650. [Google Scholar] [CrossRef]
- Berghuis, P.; Rajnicek, A.M.; Morozov, Y.M.; Ross, R.A.; Mulder, J.; Urban, G.M.; Monory, K.; Marsicano, G.; Matteoli, M.; Canty, A.; et al. Hardwiring the brain: Endocannabinoids shape neuronal connectivity. Science 2007, 316, 1212–1216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diaz-Alonso, J.; Guzman, M.; Galve-Roperh, I. Endocannabinoids via CB(1) receptors act as neurogenic niche cues during cortical development. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2012, 367, 3229–3241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaffuri, A.L.; Ladarre, D.; Lenkei, Z. Type-1 cannabinoid receptor signaling in neuronal development. Pharmacology 2012, 90, 19–39. [Google Scholar] [CrossRef]
- Goldschmidt, L.; Richardson, G.A.; Cornelius, M.D.; Day, N.L. Prenatal marijuana and alcohol exposure and academic achievement at age 10. Neurotoxicol. Teratol. 2004, 26, 521–532. [Google Scholar] [CrossRef]
- Smith, A.M.; Fried, P.A.; Hogan, M.J.; Cameron, I. Effects of prenatal marijuana on visuospatial working memory: An fMRI study in young adults. Neurotoxicol. Teratol. 2006, 28, 286–295. [Google Scholar] [CrossRef]
- Campolongo, P.; Trezza, V.; Cassano, T.; Gaetani, S.; Morgese, M.G.; Ubaldi, M.; Soverchia, L.; Antonelli, T.; Ferraro, L.; Massi, M.; et al. Perinatal exposure to delta-9-tetrahydrocannabinol causes enduring cognitive deficits associated with alteration of cortical gene expression and neurotransmission in rats. Addict. Biol. 2007, 12, 485–495. [Google Scholar] [CrossRef]
- Mereu, G.; Fa, M.; Ferraro, L.; Cagiano, R.; Antonelli, T.; Tattoli, M.; Ghiglieri, V.; Tanganelli, S.; Gessa, G.L.; Cuomo, V. Prenatal exposure to a cannabinoid agonist produces memory deficits linked to dysfunction in hippocampal long-term potentiation and glutamate release. Proc. Natl. Acad. Sci. USA 2003, 100, 4915–4920. [Google Scholar] [CrossRef] [Green Version]
- Silva, L.; Zhao, N.; Popp, S.; Dow-Edwards, D. Prenatal tetrahydrocannabinol (THC) alters cognitive function and amphetamine response from weaning to adulthood in the rat. Neurotoxicol. Teratol. 2012, 34, 63–71. [Google Scholar] [CrossRef] [Green Version]
- O’Shea, M.; McGregor, I.S.; Mallet, P.E. Repeated cannabinoid exposure during perinatal, adolescent or early adult ages produces similar longlasting deficits in object recognition and reduced social interaction in rats. J. Psychopharmacol. 2006, 20, 611–621. [Google Scholar] [CrossRef]
- Spano, M.S.; Ellgren, M.; Wang, X.; Hurd, Y.L. Prenatal cannabis exposure increases heroin seeking with allostatic changes in limbic enkephalin systems in adulthood. Biol. Psychiatry 2007, 61, 554–563. [Google Scholar] [CrossRef] [Green Version]
- Vela, G.; Martin, S.; Garcia-Gil, L.; Crespo, J.A.; Ruiz-Gayo, M.; Fernandez-Ruiz, J.J.; Garcia-Lecumberri, C.; Pelaprat, D.; Fuentes, J.A.; Ramos, J.A.; et al. Maternal exposure to delta9-tetrahydrocannabinol facilitates morphine self-administration behavior and changes regional binding to central mu opioid receptors in adult offspring female rats. Brain Res. 1998, 807, 101–109. [Google Scholar] [CrossRef]
- Molina-Holgado, F.; Alvarez, F.J.; Gonzalez, I.; Antonio, M.T.; Leret, M.L. Maternal exposure to delta 9-tetrahydrocannabinol (delta 9-THC) alters indolamine levels and turnover in adult male and female rat brain regions. Brain Res. Bull. 1997, 43, 173–178. [Google Scholar] [CrossRef]
- DiNieri, J.A.; Wang, X.; Szutorisz, H.; Spano, S.M.; Kaur, J.; Casaccia, P.; Dow-Edwards, D.; Hurd, Y.L. Maternal cannabis use alters ventral striatal dopamine D2 gene regulation in the offspring. Biol. Psychiatry 2011, 70, 763–769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Dow-Edwards, D.; Anderson, V.; Minkoff, H.; Hurd, Y.L. In utero marijuana exposure associated with abnormal amygdala dopamine D2 gene expression in the human fetus. Biol. Psychiatry 2004, 56, 909–915. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Gil, L.; de Miguel, R.; Romero, J.; Perez, A.; Ramos, J.A.; Fernandez-Ruiz, J.J. Perinatal delta9-tetrahydrocannabinol exposure augmented the magnitude of motor inhibition caused by GABA(B), but not GABA(A), receptor agonists in adult rats. Neurotoxicol. Teratol. 1999, 21, 277–283. [Google Scholar] [CrossRef]
- Saez, T.M.; Aronne, M.P.; Caltana, L.; Brusco, A.H. Prenatal exposure to the CB1 and CB2 cannabinoid receptor agonist WIN 55,212-2 alters migration of early-born glutamatergic neurons and GABAergic interneurons in the rat cerebral cortex. J. Neurochem. 2014, 129, 637–648. [Google Scholar] [CrossRef]
- Antonelli, T.; Tanganelli, S.; Tomasini, M.C.; Finetti, S.; Trabace, L.; Steardo, L.; Sabino, V.; Carratu, M.R.; Cuomo, V.; Ferraro, L. Long-term effects on cortical glutamate release induced by prenatal exposure to the cannabinoid receptor agonist (R)-(+)-[2,3-dihydro-5-methyl-3-(4-morpholinyl-methyl)pyrrolo[1,2,3-de]-1,4-benzo xazin-6-yl]-1-naphthalenylmethanone: An in vivo microdialysis study in the awake rat. Neuroscience 2004, 124, 367–375. [Google Scholar] [CrossRef] [PubMed]
- Suarez, I.; Bodega, G.; Rubio, M.; Fernandez-Ruiz, J.J.; Ramos, J.A.; Fernandez, B. Prenatal cannabinoid exposure down- regulates glutamate transporter expressions (GLAST and EAAC1) in the rat cerebellum. Dev. Neurosci. 2004, 26, 45–53. [Google Scholar] [CrossRef]
- Kumar, A.M.; Haney, M.; Becker, T.; Thompson, M.L.; Kream, R.M.; Miczek, K. Effect of early exposure to delta-9-tetrahydrocannabinol on the levels of opioid peptides, gonadotropin-releasing hormone and substance P in the adult male rat brain. Brain Res. 1990, 525, 78–83. [Google Scholar] [CrossRef]
- Miranda, C.C.; Barata, T.; Vaz, S.H.; Ferreira, C.; Quintas, A.; Bekman, E.P. hiPSC-based model of prenatal exposure to cannabinoids: Effect on neuronal differentiation. Front. Mol. Neurosci. 2020, 13, 119. [Google Scholar] [CrossRef]
- Wanner, N.M.; Colwell, M.; Drown, C.; Faulk, C. Developmental cannabidiol exposure increases anxiety and modifies genome-wide brain DNA methylation in adult female mice. Clin. Epigenetics 2021, 13, 4. [Google Scholar] [CrossRef]
- Schlosburg, J.E.; Blankman, J.L.; Long, J.Z.; Nomura, D.K.; Pan, B.; Kinsey, S.G.; Nguyen, P.T.; Ramesh, D.; Booker, L.; Burston, J.J.; et al. Chronic monoacylglycerol lipase blockade causes functional antagonism of the endocannabinoid system. Nat. Neurosci. 2010, 13, 1113–1119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, C.S.; Morgan, D.; Jew, C.P.; Haskins, C.; Andrews, M.J.; Leishman, E.; Spencer, C.M.; Czyzyk, T.; Bradshaw, H.; Mackie, K.; et al. Long-term consequences of perinatal fatty acid amino hydrolase inhibition. Br. J. Pharmacol. 2014, 171, 1420–1434. [Google Scholar] [CrossRef] [Green Version]
- Fried, P.A.; Makin, J.E. Neonatal behavioural correlates of prenatal exposure to marihuana, cigarettes and alcohol in a low risk population. Neurotoxicol. Teratol. 1987, 9, 1–7. [Google Scholar] [CrossRef]
- El Marroun, H.; Hudziak, J.J.; Tiemeier, H.; Creemers, H.; Steegers, E.A.; Jaddoe, V.W.; Hofman, A.; Verhulst, F.C.; van den Brink, W.; Huizink, A.C. Intrauterine cannabis exposure leads to more aggressive behavior and attention problems in 18-month-old girls. Drug Alcohol Depend. 2011, 118, 470–474. [Google Scholar] [CrossRef] [Green Version]
- Fried, P.A.; Watkinson, B. 36- and 48-month neurobehavioral follow-up of children prenatally exposed to marijuana, cigarettes, and alcohol. J. Dev. Behav. Pediatr. 1990, 11, 49–58. [Google Scholar] [CrossRef]
- Day, N.L.; Richardson, G.A.; Goldschmidt, L.; Robles, N.; Taylor, P.M.; Stoffer, D.S.; Cornelius, M.D.; Geva, D. Effect of prenatal marijuana exposure on the cognitive development of offspring at age three. Neurotoxicol. Teratol. 1994, 16, 169–175. [Google Scholar] [CrossRef]
- Goldschmidt, L.; Richardson, G.A.; Willford, J.; Day, N.L. Prenatal marijuana exposure and intelligence test performance at age 6. J. Am. Acad. Child Adolesc. Psychiatry 2008, 47, 254–263. [Google Scholar] [CrossRef] [Green Version]
- Richardson, G.A.; Ryan, C.; Willford, J.; Day, N.L.; Goldschmidt, L. Prenatal alcohol and marijuana exposure: Effects on neuropsychological outcomes at 10 years. Neurotoxicol. Teratol. 2002, 24, 309–320. [Google Scholar] [CrossRef]
- Smith, A.M.; Fried, P.A.; Hogan, M.J.; Cameron, I. Effects of prenatal marijuana on response inhibition: An fMRI study of young adults. Neurotoxicol. Teratol. 2004, 26, 533–542. [Google Scholar] [CrossRef] [PubMed]
- Tortoriello, G.; Morris, C.V.; Alpar, A.; Fuzik, J.; Shirran, S.L.; Calvigioni, D.; Keimpema, E.; Botting, C.H.; Reinecke, K.; Herdegen, T.; et al. Miswiring the brain: Delta9-tetrahydrocannabinol disrupts cortical development by inducing an SCG10/stathmin-2 degradation pathway. EMBO J. 2014, 33, 668–685. [Google Scholar] [CrossRef] [PubMed]
- de Salas-Quiroga, A.; Diaz-Alonso, J.; Garcia-Rincon, D.; Remmers, F.; Vega, D.; Gomez-Canas, M.; Lutz, B.; Guzman, M.; Galve-Roperh, I. Prenatal exposure to cannabinoids evokes long-lasting functional alterations by targeting CB1 receptors on developing cortical neurons. Proc. Natl. Acad. Sci. USA 2015, 112, 13693–13698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fride, E. Multiple roles for the endocannabinoid system during the earliest stages of life: Pre- and postnatal development. J. Neuroendocrinol. 2008, 20 (Suppl. 1), 75–81. [Google Scholar] [CrossRef]
- Harkany, T.; Guzman, M.; Galve-Roperh, I.; Berghuis, P.; Devi, L.A.; Mackie, K. The emerging functions of endocannabinoid signaling during CNS development. Trends Pharmacol. Sci. 2007, 28, 83–92. [Google Scholar] [CrossRef]
- Higuera-Matas, A.; Ucha, M.; Ambrosio, E. Long-term consequences of perinatal and adolescent cannabinoid exposure on neural and psychological processes. Neurosci. Biobehav. Rev. 2015, 55, 119–146. [Google Scholar] [CrossRef]
- Scheyer, A.F.; Melis, M.; Trezza, V.; Manzoni, O.J.J. Consequences of perinatal cannabis exposure. Trends Neurosci. 2019, 42, 871–884. [Google Scholar] [CrossRef]
- Dall’Aglio, C.; Polisca, A.; Cappai, M.G.; Mercati, F.; Troisi, A.; Pirino, C.; Scocco, P.; Maranesi, M. Immunohistochemistry detected and localized cannabinoid receptor type 2 in bovine fetal pancreas at late gestation. Eur. J. Histochem. 2017, 61, 2761. [Google Scholar] [CrossRef] [Green Version]
- Hales, C.N.; Barker, D.J. Type 2 (non-insulin-dependent) diabetes mellitus: The thrifty phenotype hypothesis. Diabetologia 1992, 35, 595–601. [Google Scholar] [CrossRef]
- Bazwinsky-Wutschke, I.; Zipprich, A.; Dehghani, F. Endocannabinoid system in hepatic glucose metabolism, fatty liver disease, and cirrhosis. Int. J. Mol. Sci. 2019, 20, 2516. [Google Scholar] [CrossRef] [Green Version]
- Kim, W.; Doyle, M.E.; Liu, Z.; Lao, Q.; Shin, Y.K.; Carlson, O.D.; Kim, H.S.; Thomas, S.; Napora, J.K.; Lee, E.K.; et al. Cannabinoids inhibit insulin receptor signaling in pancreatic beta-cells. Diabetes 2011, 60, 1198–1209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Athanasiou, A.; Clarke, A.B.; Turner, A.E.; Kumaran, N.M.; Vakilpour, S.; Smith, P.A.; Bagiokou, D.; Bradshaw, T.D.; Westwell, A.D.; Fang, L.; et al. Cannabinoid receptor agonists are mitochondrial inhibitors: A unified hypothesis of how cannabinoids modulate mitochondrial function and induce cell death. Biochem. Biophys. Res. Commun. 2007, 364, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Mendizabal-Zubiaga, J.; Melser, S.; Benard, G.; Ramos, A.; Reguero, L.; Arrabal, S.; Elezgarai, I.; Gerrikagoitia, I.; Suarez, J.; Rodriguez De Fonseca, F.; et al. Cannabinoid CB1 receptors are localized in striated muscle mitochondria and regulate mitochondrial respiration. Front. Physiol. 2016, 7, 476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bansal, A.; Rashid, C.; Simmons, R.A. Chapter 14—Impact of fetal programming on mitochondrial function and susceptibility to obesity and type 2 diabetes. In Mitochondria in Obesity and Type 2 Diabetes; Morio, B., Pénicaud, L., Rigoulet, M., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 325–345. [Google Scholar]
- Dias-Rocha, C.P.; Almeida, M.M.; Santana, E.M.; Costa, J.C.B.; Franco, J.G.; Pazos-Moura, C.C.; Trevenzoli, I.H. Maternal high-fat diet induces sex-specific endocannabinoid system changes in newborn rats and programs adiposity, energy expenditure and food preference in adulthood. J. Nutr. Biochem. 2018, 51, 56–68. [Google Scholar] [CrossRef]
- Brents, L.K. Marijuana, the endocannabinoid system and the female reproductive system. Yale J. Biol. Med. 2016, 89, 175–191. [Google Scholar] [PubMed]
- Gundersen, T.D.; Jorgensen, N.; Andersson, A.M.; Bang, A.K.; Nordkap, L.; Skakkebaek, N.E.; Priskorn, L.; Juul, A.; Jensen, T.K. Association between use of marijuana and male reproductive hormones and semen quality: A study among 1,215 healthy young men. Am. J. Epidemiol. 2015, 182, 473–481. [Google Scholar] [CrossRef]
- Mendelson, J.H.; Mello, N.K.; Ellingboe, J.; Skupny, A.S.; Lex, B.W.; Griffin, M. Marihuana smoking suppresses luteinizing hormone in women. J. Pharmacol. Exp. Ther. 1986, 237, 862–866. [Google Scholar]
- Reich, R.; Laufer, N.; Lewysohn, O.; Cordova, T.; Ayalon, D.; Tsafriri, A. In vitro effects of cannabinoids on follicular function in the rat. Biol. Reprod. 1982, 27, 223–231. [Google Scholar] [CrossRef] [Green Version]
- Rosenkrantz, H.; Grant, R.J.; Fleischman, R.W.; Baker, J.R. Marihuana-induced embryotoxicity in the rabbit. Fundam. Appl. Toxicol. 1986, 7, 236–243. [Google Scholar] [CrossRef]
- Treinen, K.A.; Sneeden, J.L.; Heindel, J.J. Specific inhibition of FSH-stimulated cAMP accumulation by delta 9-tetrahydrocannabinol in cultured rat granulosa cells. Toxicol. Appl. Pharmacol. 1993, 118, 53–57. [Google Scholar] [CrossRef]
- Smith, C.G.; Almirez, R.G.; Berenberg, J.; Asch, R.H. Tolerance develops to the disruptive effects of delta 9-tetrahydrocannabinol on primate menstrual cycle. Science 1983, 219, 1453–1455. [Google Scholar] [CrossRef]
- De Domenico, E.; Todaro, F.; Rossi, G.; Dolci, S.; Geremia, R.; Rossi, P.; Grimaldi, P. Overactive type 2 cannabinoid receptor induces meiosis in fetal gonads and impairs ovarian reserve. Cell Death Dis. 2017, 8, e3085. [Google Scholar] [CrossRef] [PubMed]
- Dalterio, S.L.; deRooij, D.G. Maternal cannabinoid exposure. Effects on spermatogenesis in male offspring. Int. J. Androl. 1986, 9, 250–258. [Google Scholar] [CrossRef] [PubMed]
- GW Pharmaceuticals Plc Reports Fiscal Fourth Quarter and Year-End 2018 Financial Results and Operational Progress. Available online: https://ir.gwpharm.com/news-releases/news-release-details/gw-pharmaceuticals-plc-reports-fiscal-fourth-quarter-and-year-0 (accessed on 24 June 2021).
- Martinez-Pena, A.A.; Lee, K.; Petrik, J.J.; Hardy, D.B.; Holloway, A.C. Gestational exposure to Delta(9)-THC impacts ovarian follicular dynamics and angiogenesis in adulthood in Wistar rats. J. Dev. Orig. Health Dis. 2021, 1–5. [Google Scholar] [CrossRef]
- Castel, P.; Barbier, M.; Poumerol, E.; Mandon-Pepin, B.; Tassistro, V.; Lepidi, H.; Pelissier-Alicot, A.L.; Manzoni, O.J.; Courbiere, B. Prenatal cannabinoid exposure alters the ovarian reserve in adult offspring of rats. Arch. Toxicol. 2020, 94, 4131–4141. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martínez-Peña, A.A.; Perono, G.A.; Gritis, S.A.; Sharma, R.; Selvakumar, S.; Walker, O.S.; Gurm, H.; Holloway, A.C.; Raha, S. The Impact of Early Life Exposure to Cannabis: The Role of the Endocannabinoid System. Int. J. Mol. Sci. 2021, 22, 8576. https://doi.org/10.3390/ijms22168576
Martínez-Peña AA, Perono GA, Gritis SA, Sharma R, Selvakumar S, Walker OS, Gurm H, Holloway AC, Raha S. The Impact of Early Life Exposure to Cannabis: The Role of the Endocannabinoid System. International Journal of Molecular Sciences. 2021; 22(16):8576. https://doi.org/10.3390/ijms22168576
Chicago/Turabian StyleMartínez-Peña, Annia A., Genevieve A. Perono, Sarah Alexis Gritis, Reeti Sharma, Shamini Selvakumar, O’Llenecia S. Walker, Harmeet Gurm, Alison C. Holloway, and Sandeep Raha. 2021. "The Impact of Early Life Exposure to Cannabis: The Role of the Endocannabinoid System" International Journal of Molecular Sciences 22, no. 16: 8576. https://doi.org/10.3390/ijms22168576
APA StyleMartínez-Peña, A. A., Perono, G. A., Gritis, S. A., Sharma, R., Selvakumar, S., Walker, O. S., Gurm, H., Holloway, A. C., & Raha, S. (2021). The Impact of Early Life Exposure to Cannabis: The Role of the Endocannabinoid System. International Journal of Molecular Sciences, 22(16), 8576. https://doi.org/10.3390/ijms22168576