
Soluble Receptor for Advanced Glycation End Products (sRAGE) Is a Sensitive Biomarker in Human Pulmonary Arterial Hypertension

 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Demographic Characteristics of the Adult and Pediatric PAH Cohorts
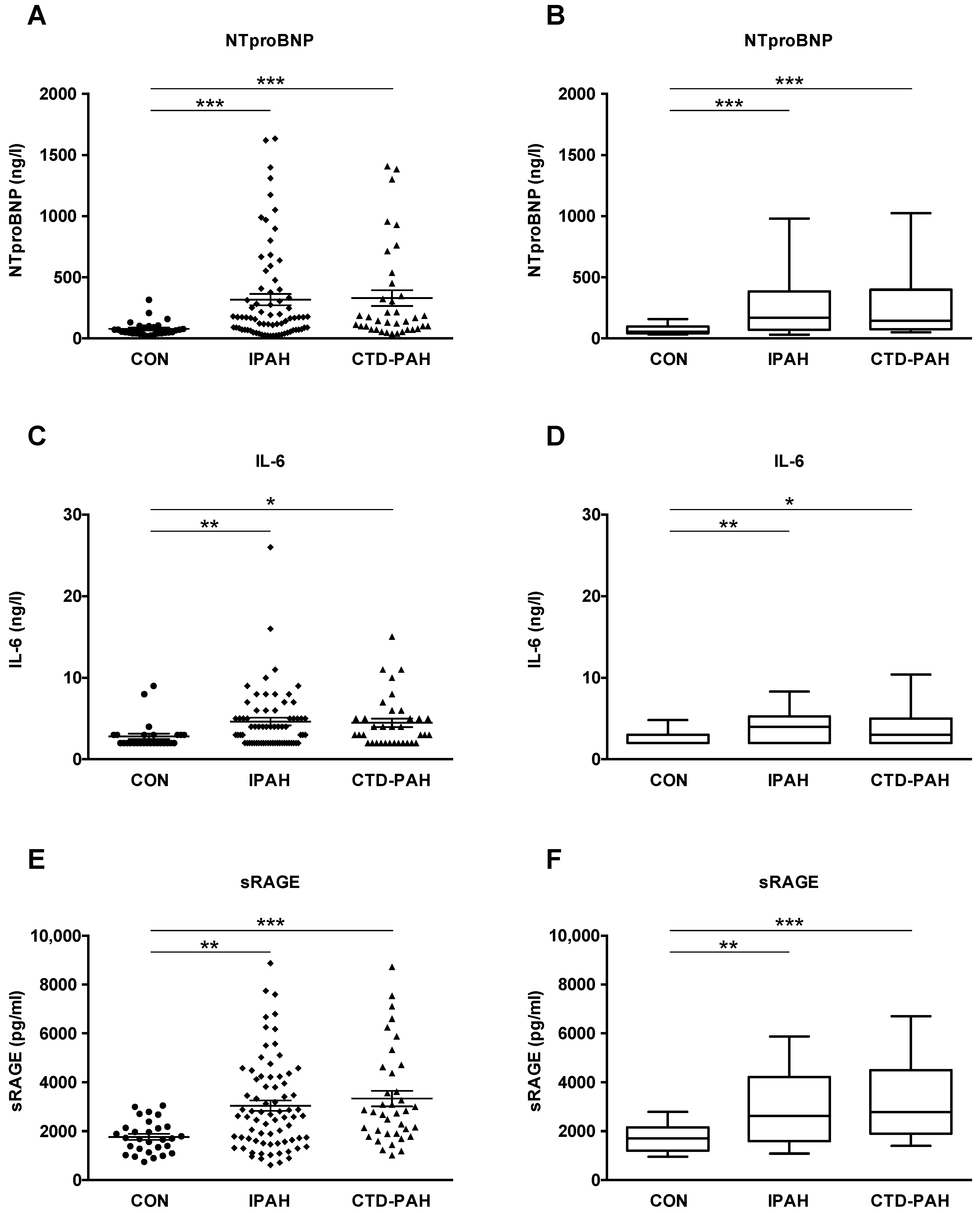
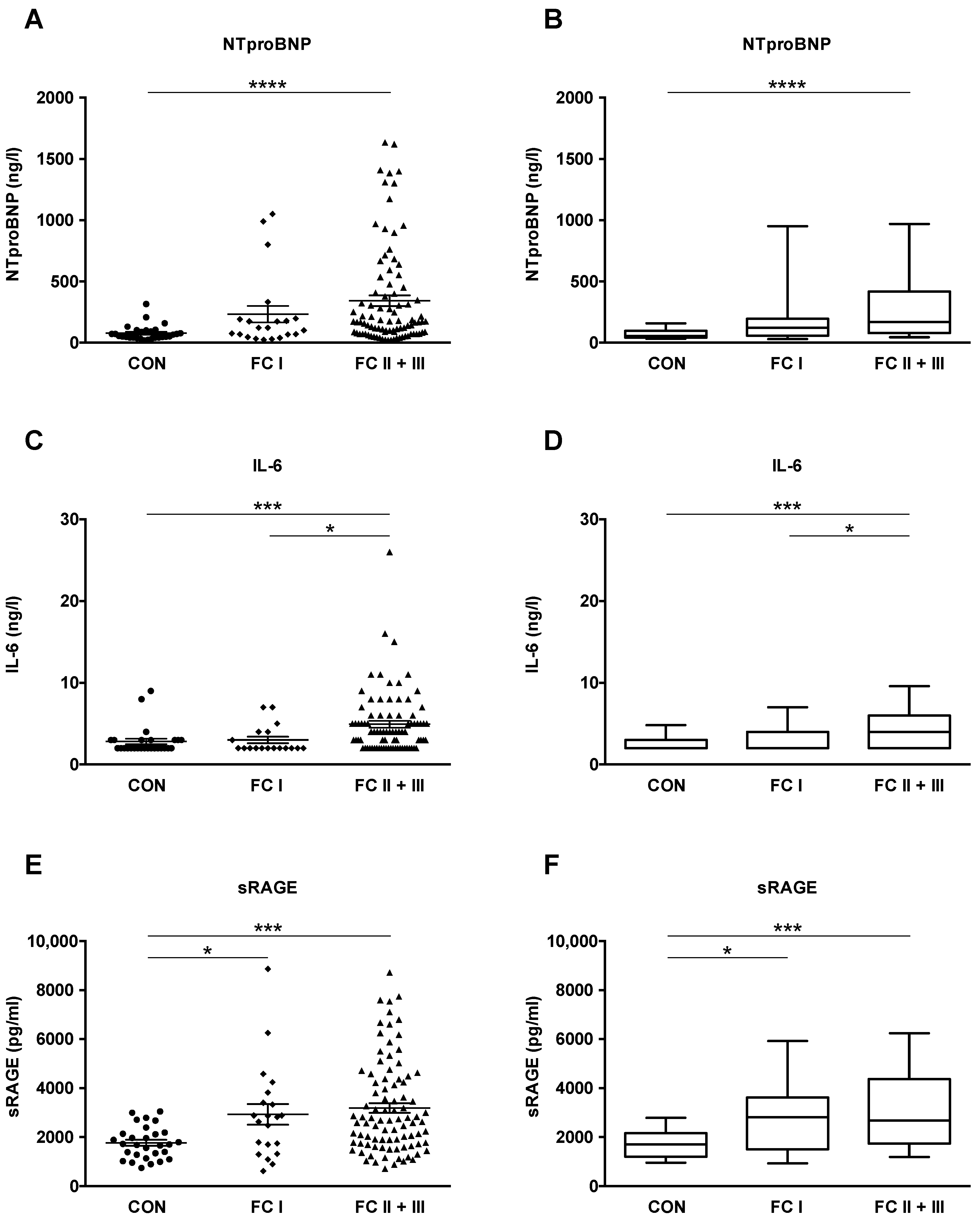
2.2. NTproBNP, IL-6 and sRAGE Plasma Levels Are Elevated in Adult IPAH and CTD-PAH Patients versus Healthy Control Subjects
2.3. Receiver-Operating Characteristic (ROC) Analysis: Plasma sRAGE Has Diagnostic Accuracy Comparable to the Established Biomarker NTproBNP
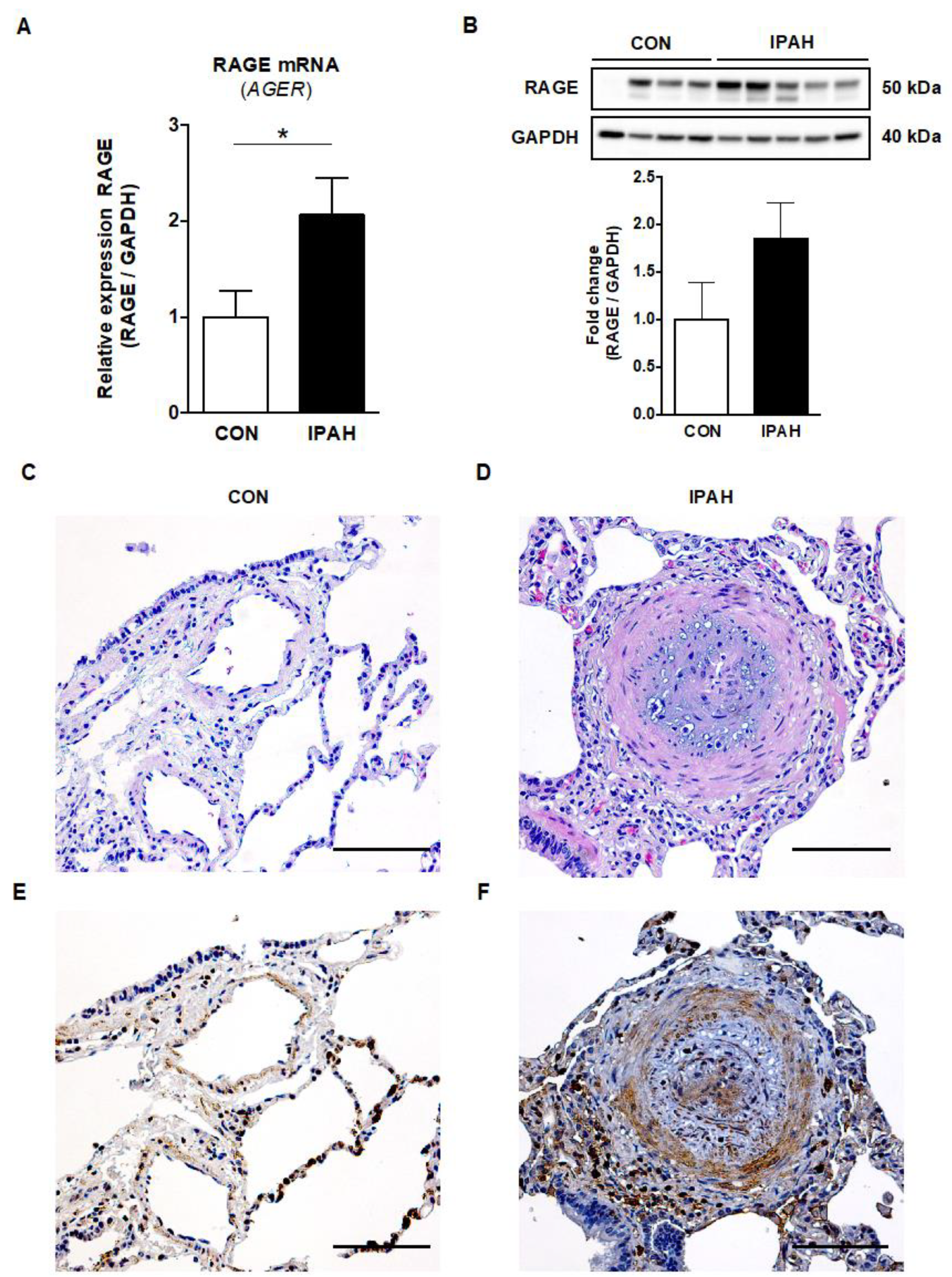
2.4. RAGE mRNA and Protein Expression Is Increased in Lung Tissue from End-Stage IPAH Patients versus Donor Controls
2.5. Immunohistochemistry RAGE Signal Is Augmented in the Pulmonary Vasculature of Adult IPAH Patients vs. Controls
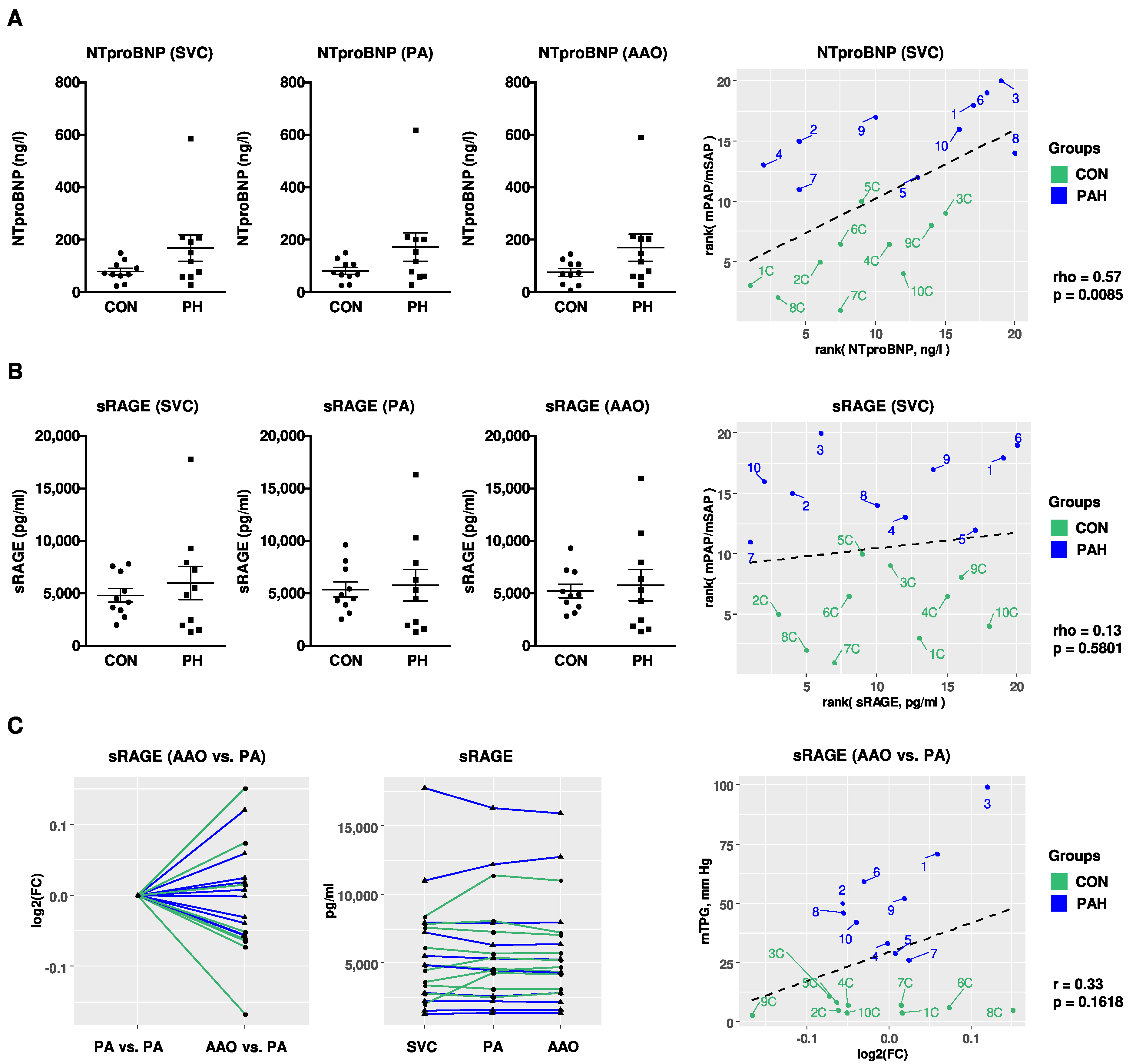
2.6. Compartment-Specific Plasma Concentrations of NTproBNP, IL-6, and sRAGE in Children with PH versus Non-PH Controls in the Systemic and Pulmonary Circulation
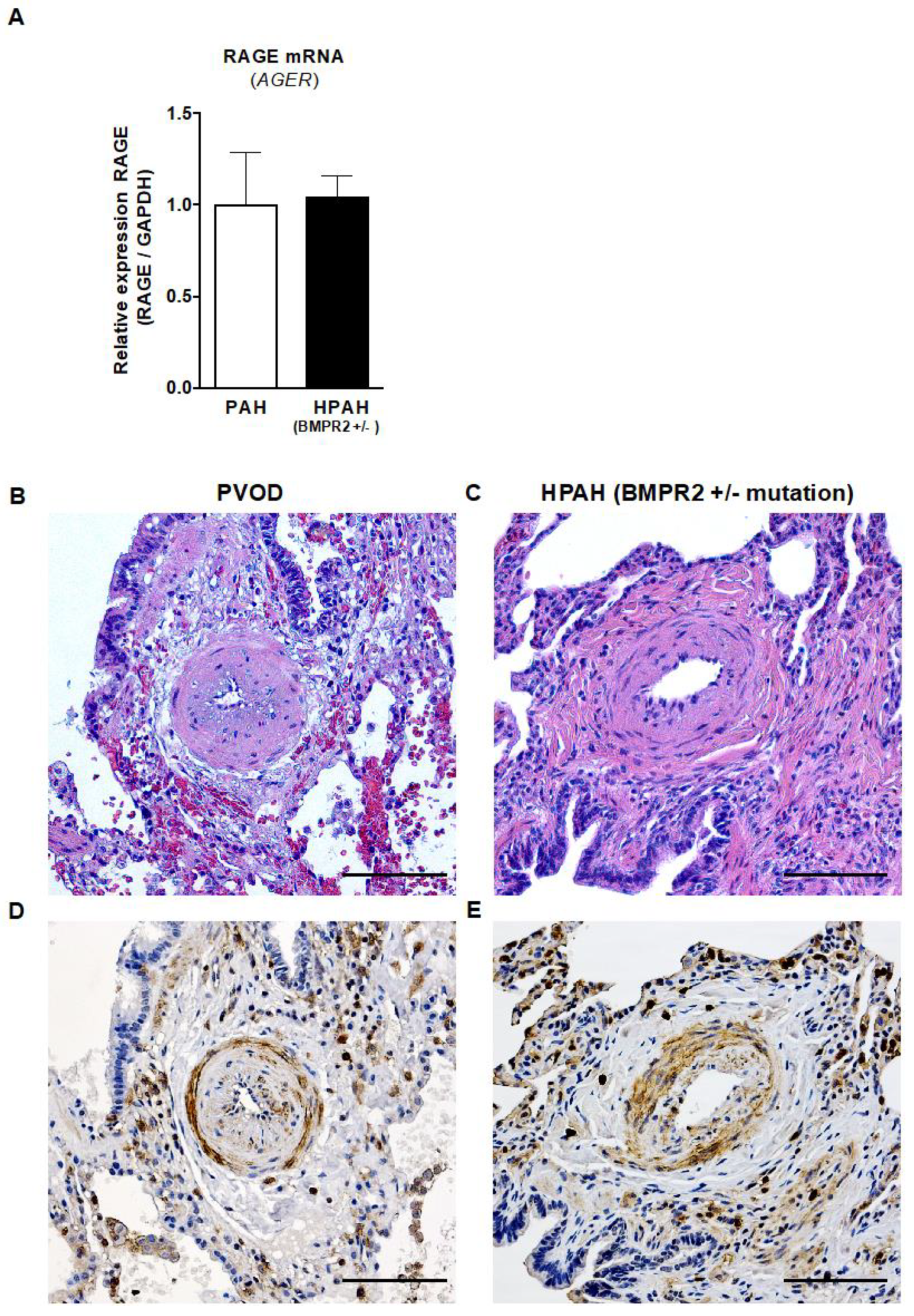
2.7. RAGE mRNA Expression in Explanted Lungs from Children with PAH and HPAH (Heterozygous BMPR2 Mutation)
2.8. Strong Immunohistochemistry Signal for RAGE in the Intima and Media of Pulmonary Vessels of Children with PAH/PVOD or Heritable PAH (BMPR2 +/− Mutation) Undergoing Lung Transplantation
3. Discussion
4. Materials and Methods
4.1. Clinical Study Design
4.2. Biomarker Assays
4.3. RNA Extraction and Quantitative Real-Time PCR
4.4. Protein Extraction and Western Blot
4.5. Immunohistochemistry
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 6MWD | six-minute walk distance |
| AAO | ascending aorta |
| AUC | area under the ROC curve |
| BMI | body mass index |
| BMPR2 | bone morphogenetic protein receptor 2 |
| CI | cardiac index |
| CON | control |
| CTD-PAH | connective tissue disease-associated pulmonary arterial hypertension |
| CTEPH | chronic thromboembolic pulmonary hypertension |
| DAMP | damage-associated molecular pattern |
| EPPVDN | European Pediatric Pulmonary Vascular Disease Network |
| FC | functional class |
| GAPDH | Glyceraldehyde 3-phosphate dehydrogenase |
| HMGB1 | high-mobility group box 1 |
| HPAH | heritable pulmonary arterial hypertension |
| IL-6 | interleukin-6 |
| IMPA1 | inositol monophosphatase 1 |
| IPAH | idiopathic pulmonary arterial hypertension |
| LV | left ventricle |
| LuTx | lung transplantation |
| LVOTO | left ventricular outflow tract obstruction |
| mPAP | mean pulmonary arterial pressure |
| mSAP | mean systemic arterial pressure |
| mTPG | mean transpulmonary pressure gradient |
| NTproBNP | N-terminal pro-brain natriuretic peptide |
| PA | pulmonary artery |
| PAH | pulmonary arterial hypertension |
| PASMC | pulmonary artery smooth muscle cells |
| PCH | pulmonary capillary hemangiomatosis |
| PH | pulmonary hypertension |
| PHA | Pulmonary Hypertension Association |
| PPARγ | peroxisome proliferator-activated receptor gamma |
| PRR | pattern recognition receptor |
| PVD | pulmonary vascular disease |
| PVOD | pulmonary veno-occlusive disease |
| PVR | pulmonary vascular resistance |
| ROC | receiver operating characteristic |
| RV | right ventricle |
| RVH | right ventricular hypertrophy |
| RVD | right ventricular dysfunction |
| SMC | smooth muscle cells |
| sRAGE | soluble receptor for advanced glycation end products |
| SVC | superior vena cava |
| WHO FC | World Health Organization functional class |
References
- Humbert, M.; Guignabert, C.; Bonnet, S.; Dorfmuller, P.; Klinger, J.R.; Nicolls, M.R.; Olschewski, A.J.; Pullamsetti, S.S.; Schermuly, R.T.; Stenmark, K.R.; et al. Pathology and pathobiology of pulmonary hypertension: State of the art and research perspectives. Eur. Respir. J. 2019, 53, 1801887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agrawal, V.; Lahm, T.; Hansmann, G.; Hemnes, A.R. Molecular mechanisms of right ventricular dysfunction in pulmonary arterial hypertension: Focus on the coronary vasculature, sex hormones, and glucose/lipid metabolism. Cardiovasc. Diagn. Ther. 2020, 10, 1522–1540. [Google Scholar] [CrossRef]
- Vonk Noordegraaf, A.; Chin, K.M.; Haddad, F.; Hassoun, P.M.; Hemnes, A.R.; Hopkins, S.R.; Kawut, S.M.; Langleben, D.; Lumens, J.; Naeije, R. Pathophysiology of the right ventricle and of the pulmonary circulation in pulmonary hypertension: An update. Eur. Respir. J. 2019, 53, 1801900. [Google Scholar] [CrossRef]
- Bernardo, R.J.; Haddad, F.; Couture, E.J.; Hansmann, G.; de Jesus Perez, V.A.; Denault, A.Y.; de Man, F.S.; Amsallem, M. Mechanics of right ventricular dysfunction in pulmonary arterial hypertension and heart failure with preserved ejection fraction. Cardiovasc. Diagn. Ther. 2020, 10, 1580–1603. [Google Scholar] [CrossRef] [PubMed]
- Morrell, N.W.; Aldred, M.A.; Chung, W.K.; Elliott, C.G.; Nichols, W.C.; Soubrier, F.; Trembath, R.C.; Loyd, J.E. Genetics and genomics of pulmonary arterial hypertension. Eur. Respir. J. 2019, 53, 1801899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berger, R.M.; Beghetti, M.; Humpl, T.; Raskob, G.E.; Ivy, D.D.; Jing, Z.C.; Bonnet, D.; Schulze-Neick, I.; Barst, R.J. Clinical features of paediatric pulmonary hypertension: A registry study. Lancet 2012, 379, 537–546. [Google Scholar] [CrossRef] [Green Version]
- Barst, R.J.; Ertel, S.I.; Beghetti, M.; Ivy, D.D. Pulmonary arterial hypertension: A comparison between children and adults. Eur. Respir. J. 2011, 37, 665–677. [Google Scholar] [CrossRef] [Green Version]
- Pezzuto, B.; Badagliacca, R.; Poscia, R.; Ghio, S.; D’Alto, M.; Vitulo, P.; Mule, M.; Albera, C.; Volterrani, M.; Fedele, F.; et al. Circulating biomarkers in pulmonary arterial hypertension: Update and future direction. J. Heart Lung Transplant. 2015, 34, 282–305. [Google Scholar] [CrossRef]
- Sweatt, A.J.; Hedlin, H.K.; Balasubramanian, V.; Hsi, A.; Blum, L.K.; Robinson, W.H.; Haddad, F.; Hickey, P.M.; Condliffe, R.; Lawrie, A.; et al. Discovery of Distinct Immune Phenotypes Using Machine Learning in Pulmonary Arterial Hypertension. Circ. Res. 2019, 124, 904–919. [Google Scholar] [CrossRef] [Green Version]
- Calvier, L.; Legchenko, E.; Grimm, L.; Sallmon, H.; Hatch, A.; Plouffe, B.D.; Schroeder, C.; Bauersachs, J.; Murthy, S.K.; Hansmann, G. Galectin-3 and aldosterone as potential tandem biomarkers in pulmonary arterial hypertension. Heart 2016, 102, 390–396. [Google Scholar] [CrossRef] [Green Version]
- Rhodes, C.J.; Wharton, J.; Ghataorhe, P.; Watson, G.; Girerd, B.; Howard, L.S.; Gibbs, J.S.R.; Condliffe, R.; Elliot, C.A.; Kiely, D.G.; et al. Plasma proteome analysis in patients with pulmonary arterial hypertension: An observational cohort study. Lancet Respir. Med. 2017, 5, 717–726. [Google Scholar] [CrossRef] [Green Version]
- Galie, N.; Humbert, M.; Vachiery, J.L.; Gibbs, S.; Lang, I.; Torbicki, A.; Simonneau, G.; Peacock, A.; Vonk Noordegraaf, A.; Beghetti, M.; et al. 2015 ESC/ERS Guidelines for the diagnosis and treatment of pulmonary hypertension: The Joint Task Force for the Diagnosis and Treatment of Pulmonary Hypertension of the European Society of Cardiology (ESC) and the European Respiratory Society (ERS): Endorsed by: Association for European Paediatric and Congenital Cardiology (AEPC), International Society for Heart and Lung Transplantation (ISHLT). Eur. Heart J. 2016, 37, 67–119. [Google Scholar] [PubMed]
- Hansmann, G.; Koestenberger, M.; Alastalo, T.P.; Apitz, C.; Austin, E.D.; Bonnet, D.; Budts, W.; D’Alto, M.; Gatzoulis, M.A.; Hasan, B.S.; et al. 2019 updated consensus statement on the diagnosis and treatment of pediatric pulmonary hypertension: The European Pediatric Pulmonary Vascular Disease Network (EPPVDN), endorsed by AEPC, ESPR and ISHLT. J. Heart Lung Transplant. 2019, 38, 879–901. [Google Scholar] [CrossRef] [Green Version]
- Sims, G.P.; Rowe, D.C.; Rietdijk, S.T.; Herbst, R.; Coyle, A.J. HMGB1 and RAGE in inflammation and cancer. Annu. Rev. Immunol. 2010, 28, 367–388. [Google Scholar] [CrossRef]
- Lin, Q.; Fan, C.; Gomez-Arroyo, J.; Van Raemdonck, K.; Meuchel, L.W.; Skinner, J.T.; Everett, A.D.; Fang, X.; Macdonald, A.A.; Yamaji-Kegan, K.; et al. HIMF (Hypoxia-Induced Mitogenic Factor) Signaling Mediates the HMGB1 (High Mobility Group Box 1)-Dependent Endothelial and Smooth Muscle Cell Crosstalk in Pulmonary Hypertension. Arterioscler. Thromb. Vasc. Biol. 2019, 39, 2505–2519. [Google Scholar] [CrossRef]
- Steenbeke, M.; De Bruyne, S.; De Buyzere, M.; Lapauw, B.; Speeckaert, R.; Petrovic, M.; Delanghe, J.R.; Speeckaert, M.M. The role of soluble receptor for advanced glycation end-products (sRAGE) in the general population and patients with diabetes mellitus with a focus on renal function and overall outcome. Crit. Rev. Clin. Lab. Sci. 2020, 58, 113–130. [Google Scholar] [CrossRef]
- Raucci, A.; Cugusi, S.; Antonelli, A.; Barabino, S.M.; Monti, L.; Bierhaus, A.; Reiss, K.; Saftig, P.; Bianchi, M.E. A soluble form of the receptor for advanced glycation endproducts (RAGE) is produced by proteolytic cleavage of the membrane-bound form by the sheddase a disintegrin and metalloprotease 10 (ADAM10). FASEB J. 2008, 22, 3716–3727. [Google Scholar] [CrossRef] [PubMed]
- Meloche, J.; Courchesne, A.; Barrier, M.; Carter, S.; Bisserier, M.; Paulin, R.; Lauzon-Joset, J.F.; Breuils-Bonnet, S.; Tremblay, E.; Biardel, S.; et al. Critical role for the advanced glycation end-products receptor in pulmonary arterial hypertension etiology. J. Am. Heart Assoc. 2013, 2, e005157. [Google Scholar] [CrossRef] [Green Version]
- Jia, D.; He, Y.; Zhu, Q.; Liu, H.; Zuo, C.; Chen, G.; Yu, Y.; Lu, A. RAGE-mediated extracellular matrix proteins accumulation exacerbates HySu-induced pulmonary hypertension. Cardiovasc. Res. 2017, 113, 586–597. [Google Scholar] [CrossRef] [Green Version]
- Rafikov, R.; McBride, M.L.; Zemskova, M.; Kurdyukov, S.; McClain, N.; Niihori, M.; Langlais, P.R.; Rafikova, O. Inositol monophosphatase 1 as a novel interacting partner of RAGE in pulmonary hypertension. Am. J. Physiol. Lung Cell. Mol. Physiol. 2019, 316, L428–L444. [Google Scholar] [CrossRef]
- Lin, Q.; Fan, C.; Skinner, J.T.; Hunter, E.N.; Macdonald, A.A.; Illei, P.B.; Yamaji-Kegan, K.; Johns, R.A. RELMalpha Licenses Macrophages for Damage-Associated Molecular Pattern Activation to Instigate Pulmonary Vascular Remodeling. J. Immunol. 2019, 203, 2862–2871. [Google Scholar] [CrossRef]
- Suzuki, S.; Nakazato, K.; Sugimoto, K.; Yoshihisa, A.; Yamaki, T.; Kunii, H.; Suzuki, H.; Saitoh, S.; Takeishi, Y. Plasma Levels of Receptor for Advanced Glycation End-Products and High-Mobility Group Box 1 in Patients with Pulmonary Hypertension. Int. Heart J. 2016, 57, 234–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moser, B.; Megerle, A.; Bekos, C.; Janik, S.; Szerafin, T.; Birner, P.; Schiefer, A.I.; Mildner, M.; Lang, I.; Skoro-Sajer, N.; et al. Local and systemic RAGE axis changes in pulmonary hypertension: CTEPH and iPAH. PLoS ONE 2014, 9, e106440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cracowski, J.-L.; Leuchte, H.H. The potential of biomarkers in pulmonary arterial hypertension. Am. J. Cardiol. 2012, 110 (Suppl. S6), S32–S38. [Google Scholar] [CrossRef]
- Badesch, D.B.; Raskob, G.E.; Elliott, C.G.; Krichman, A.M.; Farber, H.W.; Frost, A.E.; Barst, R.J.; Benza, R.L.; Liou, T.G.; Turner, M.; et al. Pulmonary arterial hypertension: Baseline characteristics from the REVEAL Registry. Chest 2010, 137, 376–387. [Google Scholar] [CrossRef] [Green Version]
- Humbert, M.; Sitbon, O.; Chaouat, A.; Bertocchi, M.; Habib, G.; Gressin, V.; Yaici, A.; Weitzenblum, E.; Cordier, J.F.; Chabot, F.; et al. Pulmonary arterial hypertension in France: Results from a national registry. Am. J. Respir. Crit. Care Med. 2006, 173, 1023–1030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chouvarine, P.; Legchenko, E.; Geldner, J.; Riehle, C.; Hansmann, G. Hypoxia drives cardiac miRNAs and inflammation in the right and left ventricle. J. Mol. Med. 2019, 97, 1427–1438. [Google Scholar] [CrossRef] [PubMed]
- Abdul-Salam, V.B.; Wharton, J.; Cupitt, J.; Berryman, M.; Edwards, R.J.; Wilkins, M.R. Proteomic analysis of lung tissues from patients with pulmonary arterial hypertension. Circulation 2010, 122, 2058–2067. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, K.; Sakaguchi, M.; Matsubara, H.; Akagi, S.; Sarashina, T.; Ejiri, K.; Akazawa, K.; Kondo, M.; Nakagawa, K.; Yoshida, M.; et al. Crucial role of RAGE in inappropriate increase of smooth muscle cells from patients with pulmonary arterial hypertension. PLoS ONE 2018, 13, e0203046. [Google Scholar] [CrossRef]
- Hansmann, G.; Wagner, R.A.; Schellong, S.; Perez, V.A.; Urashima, T.; Wang, L.; Sheikh, A.Y.; Suen, R.S.; Stewart, D.J.; Rabinovitch, M. Pulmonary arterial hypertension is linked to insulin resistance and reversed by peroxisome proliferator-activated receptor-gamma activation. Circulation 2007, 115, 1275–1284. [Google Scholar] [CrossRef]
- Hansmann, G.; de Jesus Perez, V.A.; Alastalo, T.P.; Alvira, C.M.; Guignabert, C.; Bekker, J.M.; Schellong, S.; Urashima, T.; Wang, L.; Morrell, N.W.; et al. An antiproliferative BMP-2/PPARgamma/apoE axis in human and murine SMCs and its role in pulmonary hypertension. J. Clin. Investig. 2008, 118, 1846–1857. [Google Scholar] [CrossRef] [Green Version]
- Calvier, L.; Chouvarine, P.; Legchenko, E.; Hoffmann, N.; Geldner, J.; Borchert, P.; Jonigk, D.; Mozes, M.M.; Hansmann, G. PPARgamma links BMP2 and TGFbeta1 pathways in vascular smooth muscle cells, regulating cell proliferation and glucose metabolism. Cell Metab. 2017, 25, 1118–1134.e7. [Google Scholar] [CrossRef] [Green Version]
- Calvier, L.; Chouvarine, P.; Legchenko, E.; Kokeny, G.; Mozes, M.M.; Hansmann, G. Chronic TGF-beta1 Signaling in Pulmonary Arterial Hypertension Induces Sustained Canonical Smad3 Pathways in Vascular Smooth Muscle Cells. Am. J. Respir. Cell Mol. Biol. 2019, 61, 121–123. [Google Scholar] [CrossRef]
- Legchenko, E.; Chouvarine, P.; Borchert, P.; Fernandez-Gonzalez, A.; Snay, E.; Meier, M.; Maegel, L.; Mitsialis, S.A.; Rog-Zielinska, E.A.; Kourembanas, S.; et al. The PPARgamma agonist pioglitazone reverses pulmonary hypertension and prevents right heart failure via fatty acid oxidation. Sci. Transl. Med. 2018, 10, eaao0303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meloche, J.; Paulin, R.; Courboulin, A.; Lambert, C.; Barrier, M.; Bonnet, P.; Bisserier, M.; Roy, M.; Sussman, M.A.; Agharazii, M.; et al. RAGE-dependent activation of the oncoprotein Pim1 plays a critical role in systemic vascular remodeling processes. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 2114–2124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spiekerkoetter, E.; Guignabert, C.; de Jesus Perez, V.; Alastalo, T.P.; Powers, J.M.; Wang, L.; Lawrie, A.; Ambartsumian, N.; Schmidt, A.M.; Berryman, M.; et al. S100A4 and bone morphogenetic protein-2 codependently induce vascular smooth muscle cell migration via phospho-extracellular signal-regulated kinase and chloride intracellular channel 4. Circ. Res. 2009, 105, 639–647. [Google Scholar] [CrossRef] [Green Version]
- Song, J.S.; Kang, C.M.; Park, C.K.; Yoon, H.K.; Lee, S.Y.; Ahn, J.H.; Moon, H.S. Inhibitory effect of receptor for advanced glycation end products (RAGE) on the TGF-beta-induced alveolar epithelial to mesenchymal transition. Exp. Mol. Med. 2011, 43, 517–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yerkovich, S.T.; Chang, A.B.; Carroll, M.L.; Petsky, H.L.; Scrivener, G.; Upham, J.W. Soluble receptor for advanced glycation end products (sRAGE) is present at high concentrations in the lungs of children and varies with age and the pattern of lung inflammation. Respirology 2012, 17, 841–846. [Google Scholar] [CrossRef]
- Chouvarine, P.; Giera, M.; Kastenmuller, G.; Artati, A.; Adamski, J.; Bertram, H.; Hansmann, G. Trans-right ventricle and transpulmonary metabolite gradients in human pulmonary arterial hypertension. Heart 2020, 106, 1332–1341. [Google Scholar] [CrossRef] [Green Version]
- Chouvarine, P.; Geldner, J.; Giagnorio, R.; Legchenko, E.; Bertram, H.; Hansmann, G. Trans-Right-Ventricle and Transpulmonary MicroRNA Gradients in Human Pulmonary Arterial Hypertension. Pediatr. Crit. Care Med. 2020, 21, 340–349. [Google Scholar] [CrossRef]
- Simonneau, G.; Montani, D.; Celermajer, D.S.; Denton, C.P.; Gatzoulis, M.A.; Krowka, M.; Williams, P.G.; Souza, R. Haemodynamic definitions and updated clinical classification of pulmonary hypertension. Eur. Respir. J. 2019, 53, 1801913. [Google Scholar] [CrossRef] [PubMed]
- Rosenzweig, E.B.; Abman, S.H.; Adatia, I.; Beghetti, M.; Bonnet, D.; Haworth, S.; Ivy, D.D.; Berger, R.M.F. Paediatric pulmonary arterial hypertension: Updates on definition, classification, diagnostics and management. Eur. Respir. J. 2019, 53, 1801916. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CON (n = 29) | PAH Total (n = 111) | IPAH (n = 74) | CTD-PAH (n = 37) | |
|---|---|---|---|---|
| Demographics | ||||
| Age—years | 44.8 (21–76) | 48.7 (20–80) | 47.6 (20–79) | 50.0 (26–80) |
| Male sex—n | 0 | 0 | 0 | 0 |
| Height—m | 1.64 ± 0.01 | 1.63 ± 0.01 | 1.63 ± 0.01 | 1.63 ± 0.01 |
| Weight—kg | 74.7 ± 3.6 | 71.9 ± 1.5 | 71.0 ± 2.0 | 73.8 ± 2.2 |
| BMI—kg/m2 | 27.8 ± 1.4 | 27.0 ± 0.5 | 26.7 ± 0.7 | 27.7 ± 0.8 |
| Functional Status | ||||
| WHO FC I—n (%) | - | 21 (19%) | 20 (27%) | 1 (3%) |
| WHO FC II—n (%) | - | 67 (60%) | 40 (54%) | 27 (73%) |
| WHO FC III—n (%) | - | 23 (21%) | 14 (19%) | 9 (24%) |
| Biomarker | ||||
| NTproBNP—ng/L | 78.0 ± 11.5 | 321.0 ± 37.2 | 316.8 ± 45.7 | 329.3 ± 65.0 |
| Race/ethnicity | ||||
| White | 21 | 80 | 56 | 24 |
| Black | 3 | 7 | 2 | 5 |
| Asian | 1 | 6 | 3 | 3 |
| Hispanic | 3 | 8 | 6 | 2 |
| other | 1 | 10 | 7 | 3 |
| Group | Gender | Age (Years) | Diagnosis |
|---|---|---|---|
| IPAH | female | 41 | IPAH |
| IPAH | female | 31 | IPAH |
| IPAH | female | 29 | IPAH |
| IPAH | female | 42 | IPAH |
| IPAH | female | 36 | IPAH |
| IPAH | female | 53 | IPAH |
| IPAH | female | 25 | IPAH |
| Control | female | N/A | Downsizing lung |
| Control | male | N/A | Unused donor lung |
| Control | female | N/A | Unused donor lung |
| Control | male | N/A | Unused donor lung |
| Control | male | N/A | Downsizing lung |
| Control | N/A | N/A | Unused donor lung |
| Control | N/A | N/A | Unused donor lung |
| Control | male | N/A | Unused donor lung |
| Control | N/A | N/A | Downsizing Lung |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Diekmann, F.; Chouvarine, P.; Sallmon, H.; Meyer-Kobbe, L.; Kieslich, M.; Plouffe, B.D.; Murthy, S.K.; Lichtinghagen, R.; Legchenko, E.; Hansmann, G. Soluble Receptor for Advanced Glycation End Products (sRAGE) Is a Sensitive Biomarker in Human Pulmonary Arterial Hypertension. Int. J. Mol. Sci. 2021, 22, 8591. https://doi.org/10.3390/ijms22168591
Diekmann F, Chouvarine P, Sallmon H, Meyer-Kobbe L, Kieslich M, Plouffe BD, Murthy SK, Lichtinghagen R, Legchenko E, Hansmann G. Soluble Receptor for Advanced Glycation End Products (sRAGE) Is a Sensitive Biomarker in Human Pulmonary Arterial Hypertension. International Journal of Molecular Sciences. 2021; 22(16):8591. https://doi.org/10.3390/ijms22168591
Chicago/Turabian StyleDiekmann, Franziska, Philippe Chouvarine, Hannes Sallmon, Louisa Meyer-Kobbe, Moritz Kieslich, Brian D. Plouffe, Shashi K. Murthy, Ralf Lichtinghagen, Ekaterina Legchenko, and Georg Hansmann. 2021. "Soluble Receptor for Advanced Glycation End Products (sRAGE) Is a Sensitive Biomarker in Human Pulmonary Arterial Hypertension" International Journal of Molecular Sciences 22, no. 16: 8591. https://doi.org/10.3390/ijms22168591
APA StyleDiekmann, F., Chouvarine, P., Sallmon, H., Meyer-Kobbe, L., Kieslich, M., Plouffe, B. D., Murthy, S. K., Lichtinghagen, R., Legchenko, E., & Hansmann, G. (2021). Soluble Receptor for Advanced Glycation End Products (sRAGE) Is a Sensitive Biomarker in Human Pulmonary Arterial Hypertension. International Journal of Molecular Sciences, 22(16), 8591. https://doi.org/10.3390/ijms22168591