
Development of a Rapid Fluorescent Diagnostic System to Detect Subtype H9 Influenza A Virus in Chicken Feces

 ,
,
Abstract
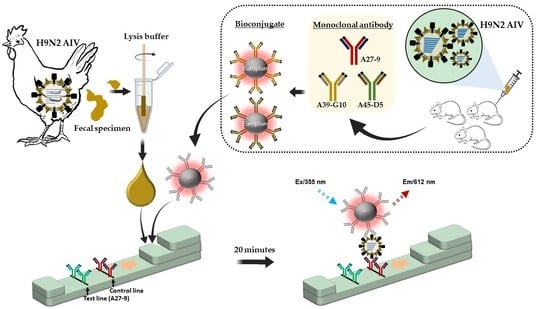
:
1. Introduction
2. Results
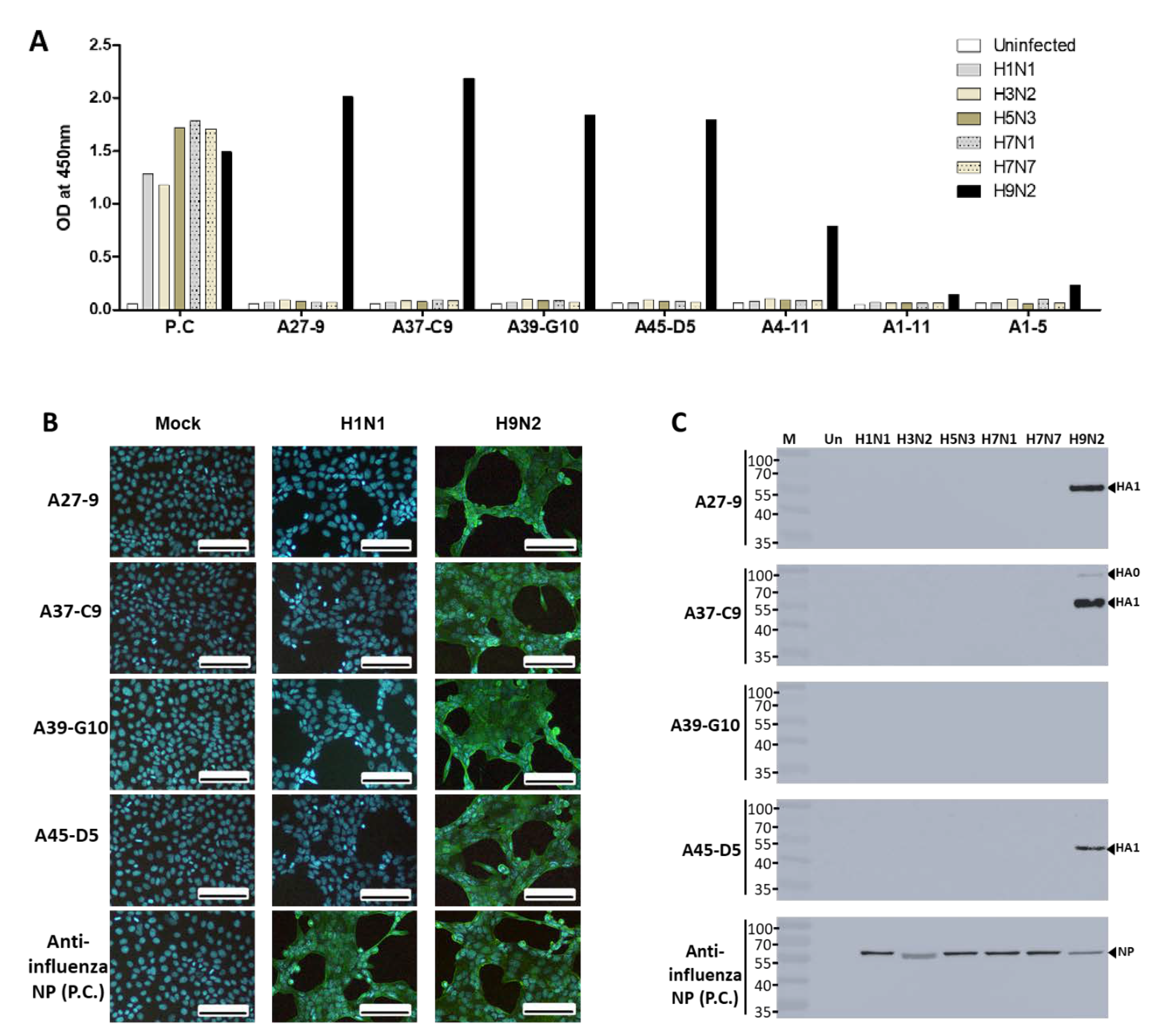
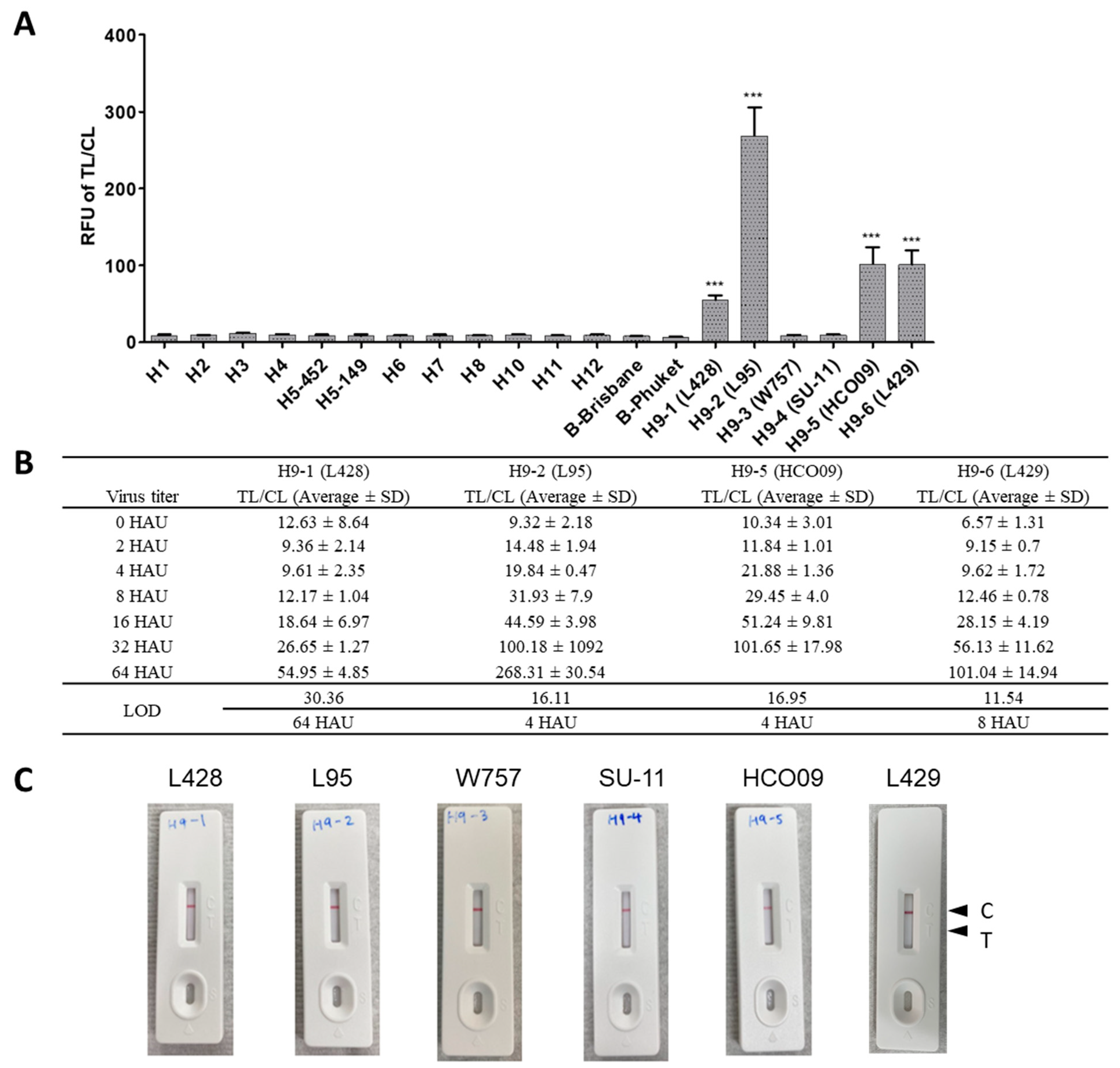
2.1. Development of McAbs
2.2. Selection of the Specific McAb Pair for FICT Assays
2.3. Optimization of Lysis Buffer
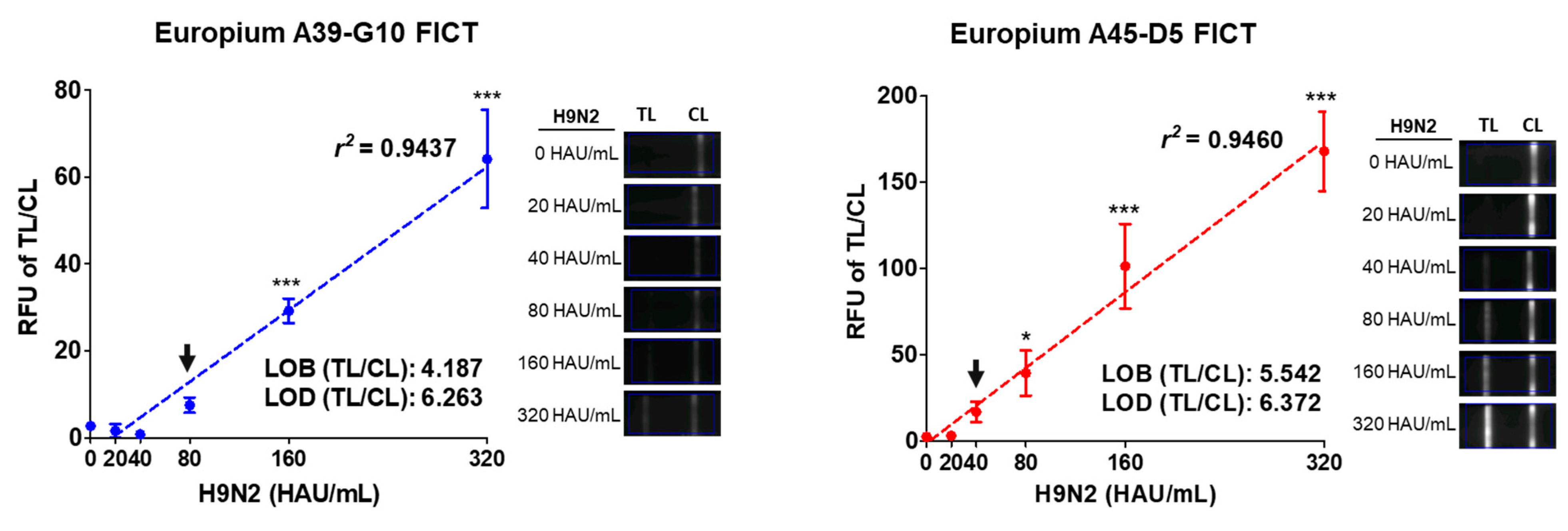
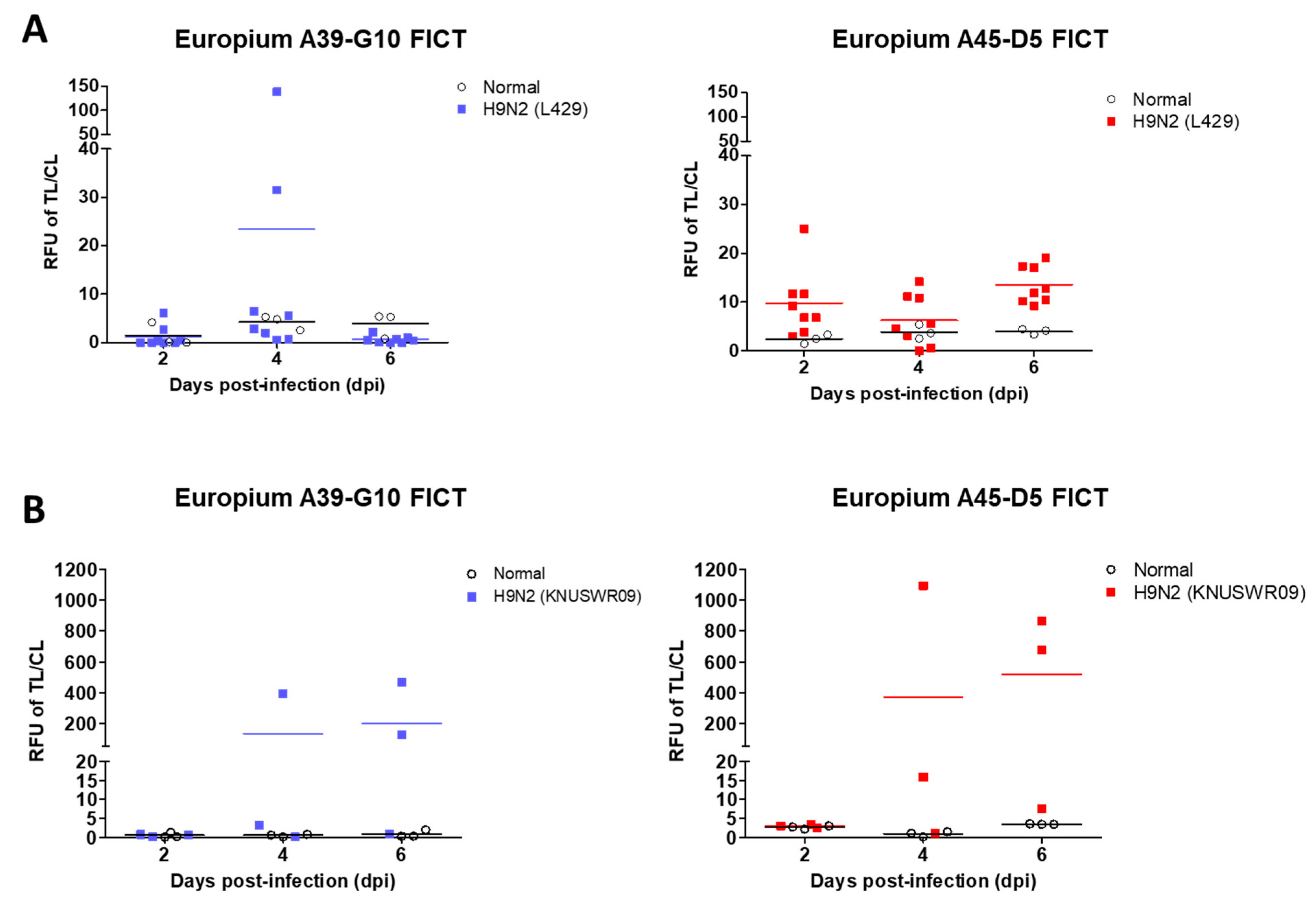
2.4. FICT Assay Performance
2.5. Clinical Study
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Cells and Viruses
4.3. Cell Fusion and Hybridoma Cell Cloning
4.4. Enzyme-Linked Immunosorbent Assay (ELISA)
4.5. Western Blot Analysis
4.6. Sandwich Fluorescent-Linked Immunosorbent Assay (FLISA)
4.7. Immunofluorescence Assay (IFA)
4.8. Conjugation of Europium Nanoparticles
4.9. Lateral Flow Test Strips for FICT
4.10. Real-Time Reverse Transcription-Polymerase Chain Reaction (rRT-PCR)
4.11. Rapid Detection Test Assay (RDT)
4.12. Clinical Study
4.13. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Homme, P.J.; Easterday, B.C. Avian Influenza Virus Infections. I. Characteristics of Influenza A/Turkey/Wisconsin/1966 Virus. Avian Dis. 1970, 14, 66. [Google Scholar] [CrossRef]
- Guo, Y.; Krauss, S.; Senne, D.; Mo, I.; Lo, K.; Xiong, X.; Norwood, M.; Shortridge, K.; Websterbh, R.G.; Guan, Y. Characterization of the Pathogenicity of Members of the Newly Established H9N2 Influenza Virus Lineages in Asia. Virology 2000, 267, 279–288. [Google Scholar] [CrossRef] [Green Version]
- Xu, K.; Ferreri, L.; Rimondi, A.; Olivera, V.; Romano, M.; Ferreyra, H.; Rago, V.; Uhart, M.; Chen, H.; Sutton, T.; et al. Isolation and characterization of an H9N2 influenza virus isolated in Argentina. Virus Res. 2012, 168, 41–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, K.S.; Xu, K.M.; Peiris, J.S.M.; Poon, L.; Yu, K.Z.; Yuen, K.-Y.; Shortridge, K.F.; Webster, R.G.; Guan, Y. Characterization of H9 Subtype Influenza Viruses from the Ducks of Southern China: A Candidate for the Next Influenza Pandemic in Humans? J. Virol. 2003, 77, 6988–6994. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Liu, X.; Cheng, J.; Peng, D.; Jia, L.; Huang, Y. Phylogenetic Analysis of the Hemagglutinin Genes of Twenty-Six Avian Influenza Viruses of Subtype H9N2 Isolated from Chickens in China During 1996–2001. Avian Dis. 2003, 47, 116–127. [Google Scholar] [CrossRef]
- Xu, K.M.; Li, K.S.; Smith, G.J.; Li, J.W.; Tai, H.; Zhang, J.X.; Webster, R.G.; Peiris, J.S.M.; Chen, H.; Guan, Y. Evolution and Molecular Epidemiology of H9N2 Influenza A Viruses from Quail in Southern China, 2000 to 2005. J. Virol. 2007, 81, 2635–2645. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.-W.; Song, C.-S.; Lee, Y.-J.; Mo, I.-P.; Garcia, M.; Suarez, D.; Kim, S.-J. Sequence analysis of the hemagglutinin gene of H9N2 Korean avian influenza viruses and assessment of the pathogenic potential of isolate MS96. Avian Dis. 2000, 44, 527. [Google Scholar] [CrossRef]
- Kwon, H.-J.; Cho, S.-H.; Kim, M.-C.; Ahn, Y.-J.; Kim, S.-J. Molecular epizootiology of recurrent low pathogenic avian influenza by H9N2 subtype virus in Korea. Avian Pathol. 2006, 35, 309–315. [Google Scholar] [CrossRef]
- Lee, Y.-J.; Shin, J.-Y.; Song, M.-S.; Lee, Y.-M.; Choi, J.-G.; Lee, E.-K.; Jeong, M.-O.; Sung, H.-W.; Kim, J.-H.; Kwon, Y.-K.; et al. Continuing evolution of H9 influenza viruses in Korean poultry. Virology 2007, 359, 313–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monne, I.; Cattoli, G.; Mazzacan, E.; Amarin, N.M.; Al Maaitah, H.M.; Al-Natour, M.Q.; Capua, I. Genetic Comparison of H9N2 AI Viruses Isolated in Jordan in 2003. Avian Dis. 2007, 51, 451–454. [Google Scholar] [CrossRef] [PubMed]
- Toroghi, R.; Momayez, R. Biological and molecular characterization of Avian influenza virus (H9N2) isolates from Iran. Acta Virol. 2006, 50, 163–168. [Google Scholar]
- Wu, R.; Sui, Z.W.; Zhang, H.B.; Chen, Q.J.; Liang, W.W.; Yang, K.L.; Xiong, Z.L.; Liu, Z.W.; Chen, Z.; Xu, D.P. Characterization of a pathogenic H9N2 influenza A virus isolated from central China in 2007. Arch. Virol. 2008, 153, 1549–1555. [Google Scholar] [CrossRef]
- Haghighat-Jahromi, M.; Asasi, K.; Nili, H.; Dadras, H.; Shooshtari, A.H. Coinfection of avian influenza virus (H9N2 subtype) with infectious bronchitis live vaccine. Arch. Virol. 2008, 153, 651–655. [Google Scholar] [CrossRef] [PubMed]
- Pan, Q.; Liu, A.; Zhang, F.; Ling, Y.; Ou, C.; Hou, N.; He, C. Co-infection of broilers with Ornithobacterium rhinotracheale and H9N2 avian influenza virus. BMC Vet.-Res. 2012, 8, 104. [Google Scholar] [CrossRef] [Green Version]
- Matrosovich, M.; Krauss, S.; Webster, R.G. H9N2 Influenza A Viruses from Poultry in Asia Have Human Virus-like Receptor Specificity. Virology 2001, 281, 156–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uyeki, T.M.; Nguyen, D.C.; Rowe, T.; Lu, X.; Hu-Primmer, J.; Huynh, L.P.; Hang, N.L.K.; Katz, J.M. Seroprevalence of Antibodies to Avian Influenza A (H5) and A (H9) Viruses among Market Poultry Workers, Hanoi, Vietnam, 2001. PLoS ONE 2012, 7, e43948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, R.; Wang, A.-R.; Liu, Z.-H.; Liang, W.; Li, X.-X.; Tang, Y.-J.; Miao, Z.-M.; Chai, T.-J. Seroprevalence of avian influenza H9N2 among poultry workers in Shandong Province, China. Eur. J. Clin. Microbiol. Infect. Dis. 2013, 32, 1347–1351. [Google Scholar] [CrossRef]
- Gomaa, M.R.; Kayed, A.S.; Elabd, M.A.; Abu Zeid, D.; Zaki, S.A.; El Rifay, A.S.; Sherif, L.S.; McKenzie, P.P.; Webster, R.G.; Webby, R.J.; et al. Avian Influenza A(H5N1) and A(H9N2) Seroprevalence and Risk Factors for Infection Among Egyptians: A Prospective, Controlled Seroepidemiological Study. J. Infect. Dis. 2015, 211, 1399–1407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heidari, A.; Mancin, M.; Nili, H.; Pourghanbari, G.H.; Lankarani, K.B.; Leardini, S.; Cattoli, G.; Monne, I.; Piccirillo, A. Serological evidence of H9N2 avian influenza virus exposure among poultry workers from Fars province of Iran. Virol. J. 2016, 13, 16. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Tian, B.; Jianfang, Z.; Yongkun, C.; Xiaodan, L.; Wenfei, Z.; Yan, L.; Jing, T.; Junfeng, G.; Tao, C.; et al. A comprehensive retrospective study of the seroprevalence of H9N2 avian influenza viruses in occupationally exposed populations in China. PLoS ONE 2017, 12, e0178328. [Google Scholar] [CrossRef]
- CDC. Rapid Diagnostic Testing for Influenza: Information for Clinical Laboratory Directors. 2019. Available online: https://www.cdc.gov/flu/professionals/diagnosis/rapidlab.htm (accessed on 28 January 2021).
- Toriyama, K.; Suzuki, T.; Inoue, T.; Eguchi, H.; Hoshi, S.; Inoue, Y.; Aizawa, H.; Miyoshi, K.; Ohkubo, M.; Hiwatashi, E.; et al. Development of an Immunochromatographic Assay Kit Using Fluorescent Silica Nanoparticles for Rapid Diagnosis of Acanthamoeba Keratitis. J. Clin. Microbiol. 2014, 53, 273–277. [Google Scholar] [CrossRef] [Green Version]
- Yeo, S.-J.; Choi, K.; Cuc, B.T.; Hong, N.N.; Bao, D.T.; Ngoc, N.M.; Le, M.Q.; Hang, N.L.K.; Thach, N.C.; Mallik, S.K.; et al. Smartphone-Based Fluorescent Diagnostic System for Highly Pathogenic H5N1 Viruses. Theranostics 2016, 6, 231–242. [Google Scholar] [CrossRef]
- Yeo, S.-J.; Cuc, B.T.; Sung, H.W.; Park, H. Evaluation of a smartphone-based rapid fluorescent diagnostic system for H9N2 virus in specific-pathogen-free chickens. Arch. Virol. 2016, 161, 2249–2256. [Google Scholar] [CrossRef] [Green Version]
- Yeo, S.-J.; Bao, D.T.; Seo, G.-E.; Bui, C.T.; Kim, D.T.H.; Anh, N.T.V.; Tien, T.T.T.; Linh, N.T.P.; Sohn, H.-J.; Chong, C.-K.; et al. Improvement of a rapid diagnostic application of monoclonal antibodies against avian influenza H7 subtype virus using Europium nanoparticles. Sci. Rep. 2017, 7, 7933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ming, F.; Cheng, Y.; Ren, C.; Suolang, S.; Zhou, H. Development of a DAS-ELISA for detection of H9N2 avian influenza virus. J. Virol. Methods 2019, 263, 38–43. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Yuan, H.; Zhou, C.; Mao, M.; Liu, Q.; Shen, H.; Cen, Y.; Qin, Z.; Ma, L.; Li, L.S. Multiplexed detection of influenza A virus subtype H5 and H9 via quantum dot-based immunoassay. Biosens. Bioelectron. 2016, 77, 464–470. [Google Scholar] [CrossRef] [PubMed]
- Yao, M.; Lv, J.; Huang, R.; Yang, Y.; Chai, T. Determination of Infective Dose of H9N2 Avian Influenza Virus in Different Routes: Aerosol, Intranasal, and Gastrointestinal. Intervirology 2014, 57, 369–374. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Ashraf, S.; Gao, S.; Lu, J.; Liu, X. Evaluation of Transmission Route and Replication Efficiency of H9N2 Avian Influenza Virus. Avian Dis. 2010, 54, 22–27. [Google Scholar] [CrossRef]
- Dilantika, C.; Sedyaningsih, E.R.; Kasper, M.R.; Agtini, M.; Listiyaningsih, E.; Uyeki, T.M.; Burgess, T.H.; Blair, P.J.; Putnam, S.D. Influenza virus infection among pediatric patients reporting diarrhea and influenza-like illness. BMC Infect. Dis. 2010, 10, 3. [Google Scholar] [CrossRef] [Green Version]
- Guan, Y.; Shortridge, K.F.; Krauss, S.; Webster, R.G. Molecular characterization of H9N2 influenza viruses: Were they the donors of the “internal” genes of H5N1 viruses in Hong Kong? Proc. Natl. Acad. Sci. USA 1999, 96, 9363–9367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, D.; Shi, W.; Shi, Y.; Wang, D.; Xiao, H.; Li, W.; Bi, Y.; Wu, Y.; Li, X.; Yan, J.; et al. Origin and diversity of novel avian influenza A H7N9 viruses causing human infection: Phylogenetic, structural, and coalescent analyses. Lancet 2013, 381, 1926–1932. [Google Scholar] [CrossRef]
- Chen, H.; Yuan, H.; Gao, R.; Zhang, J.; Wang, D.; Xiong, Y.; Fan, G.; Yang, F.; Li, X.; Zhou, J.; et al. Clinical and epidemiological characteristics of a fatal case of avian influenza A H10N8 virus infection: A descriptive study. Lancet 2014, 383, 714–721. [Google Scholar] [CrossRef]
- Yang, J.; Dai, X.; Chen, H.; Teng, Q.; Li, X.; Rong, G.; Yan, L.; Liu, Q.; Li, Z. Development of blocking ELISA for detection of antibodies against H9N2 avian influenza viruses. J. Virol. Methods 2016, 229, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Forsström, B.; Axnäs, B.B.; Rockberg, J.; Danielsson, H.; Bohlin, A.; Uhlen, M. Dissecting Antibodies with Regards to Linear and Conformational Epitopes. PLoS ONE 2015, 10, e0121673. [Google Scholar] [CrossRef]
- Sela, M.; Schechter, B.; Schechter, I.; Borek, F. Antibodies to Sequential and Conformational Determinants. Cold Spring Harb. Symp. Quant. Biol. 1967, 32, 537–545. [Google Scholar] [CrossRef]
- Chan, M.C.; Lee, N.; Chan, P.; Leung, T.; Sung, J.J.Y. Fecal detection of influenza A virus in patients with concurrent respiratory and gastrointestinal symptoms. J. Clin. Virol. 2009, 45, 208–211. [Google Scholar] [CrossRef] [PubMed]
- Pawar, S.D.; Pande, S.A.; Tare, D.S.; Keng, S.S.; Kode, S.S.; Singh, D.K.; Mullick, J. Morphological and Biochemical Characteristics of Avian Faecal Droppings and Their Impact on Survival of Avian Influenza Virus. Food Environ. Virol. 2017, 10, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Linke, D. Detergents: An overview. Methods Enzymol. 2009, 463, 603–617. [Google Scholar] [PubMed]
- Yeo, S.-J.; Huong, D.T.; Hong, N.N.; Li, C.-Y.; Choi, K.; Yu, K.; Choi, D.-Y.; Chong, C.-K.; Choi, H.S.; Mallik, S.K.; et al. Rapid and Quantitative Detection of Zoonotic Influenza A Virus Infection Utilizing Coumarin-derived dendrimer-based Fluorescent Immunochromatographic Strip Test (FICT). Theranostics 2014, 4, 1239–1249. [Google Scholar] [CrossRef] [Green Version]
- Tien, T.T.T.; Park, H.; Tuong, H.T.; Yu, S.-T.; Choi, D.-Y.; Yeo, S.-J. Development of a Rapid Fluorescent Immunochromatographic Test to Detect Respiratory Syncytial Virus. Int. J. Mol. Sci. 2018, 19, 3013. [Google Scholar] [CrossRef] [Green Version]
- Yeo, J.-S.; Liu, D.-X.; Park, H. Potential interaction of Plasmodium falciparum Hsp60 and Calpain. Korean J. Parasitol. 2015, 53, 665. [Google Scholar] [CrossRef] [PubMed]
- GraphPad Prism; Version 5.0; Software for Scientific Analysis and Graphing; GraphPad Research: La Jolla, CA, USA, 2007.










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Specimen a | Sensitivity (%) | ||||
|---|---|---|---|---|---|
| 2 dpi | |||||
| RT-PCR | FICT | RDT | |||
| D5 | G10 | D5 | G10 | ||
| Cloacal | 0 (0/6) | 0 (0/6) | 0 (0/6) | 0 (0/6) | 0 (0/6) |
| Feces | 37.50 (3/8) | 75.00 (6/8) | 12.50 (1/8) | 12.50 (1/8) | 12.50 (1/8) |
| 4 dpi | |||||
| RT-PCR | FICT | RDT | |||
| D5 | G10 | D5 | G10 | ||
| Cloacal | 50.00 (3/6) | 0 (0/6) | 0 (0/6) | 0 (0/6) | 0 (0/6) |
| Feces | 75.00 (6/8) | 37.50 (3/8) | 37.50 (3/8) | 25.00 (2/8) | 12.50 (1/8) |
| 6 dpi | |||||
| RT-PCR | FICT | RDT | |||
| D5 | G10 | D5 | G10 | ||
| Cloacal | 50.00 (3/6) | 0 (0/6) | 0 (0/6) | 0 (0/6) | 0 (0/6) |
| Feces | 100 (8/8) | 100 (8/8) | 0 (0/8) | 0 (0/8) | 0 (0/8) |
| Specimen b | Sensitivity (%) | ||||
| 2 dpi | |||||
| RT-PCR | FICT | RDT | |||
| D5 | G10 | D5 | G10 | ||
| Cloacal | 33.33 (1/3) | 0 (0/3) | 0 (0/3) | 0 (0/3) | 0 (0/3) |
| Feces | 33.33 (1/3) | 0 (0/3) | 0 (0/3) | 0 (0/3) | 0 (0/3) |
| 4 dpi | |||||
| RT-PCR | FICT | RDT | |||
| D5 | G10 | D5 | G10 | ||
| Cloacal | 66.67 (2/3) | 33.33 (1/3) | 0 (0/3) | 0 (0/3) | 0 (0/3) |
| Feces | 100 (3/3) | 66.67 (2/3) | 33.33 (1/3) | 33.33 (1/3) | 33.33 (1/3) |
| 6 dpi | |||||
| RT-PCR | FICT | RDT | |||
| D5 | G10 | D5 | G10 | ||
| Cloacal | 66.67 (2/3) | 66.67 (2/3) | 0 (0/3) | 33.33 (1/3) | 0 (0/3) |
| Feces | 100 (3/3) | 100 (3/3) | 66.67 (2/3) | 66.67 (2/3) | 66.67 (2/3) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tuong, H.T.; Jeong, J.H.; Choi, Y.K.; Park, H.; Baek, Y.H.; Yeo, S.-J. Development of a Rapid Fluorescent Diagnostic System to Detect Subtype H9 Influenza A Virus in Chicken Feces. Int. J. Mol. Sci. 2021, 22, 8823. https://doi.org/10.3390/ijms22168823
Tuong HT, Jeong JH, Choi YK, Park H, Baek YH, Yeo S-J. Development of a Rapid Fluorescent Diagnostic System to Detect Subtype H9 Influenza A Virus in Chicken Feces. International Journal of Molecular Sciences. 2021; 22(16):8823. https://doi.org/10.3390/ijms22168823
Chicago/Turabian StyleTuong, Hien Thi, Ju Hwan Jeong, Young Ki Choi, Hyun Park, Yun Hee Baek, and Seon-Ju Yeo. 2021. "Development of a Rapid Fluorescent Diagnostic System to Detect Subtype H9 Influenza A Virus in Chicken Feces" International Journal of Molecular Sciences 22, no. 16: 8823. https://doi.org/10.3390/ijms22168823
APA StyleTuong, H. T., Jeong, J. H., Choi, Y. K., Park, H., Baek, Y. H., & Yeo, S. -J. (2021). Development of a Rapid Fluorescent Diagnostic System to Detect Subtype H9 Influenza A Virus in Chicken Feces. International Journal of Molecular Sciences, 22(16), 8823. https://doi.org/10.3390/ijms22168823