
Effects of 4-Hexylresorcinol on Craniofacial Growth in Rats

 , , ,
, , ,  ,
,  and
and
Abstract
:1. Introduction
2. Results
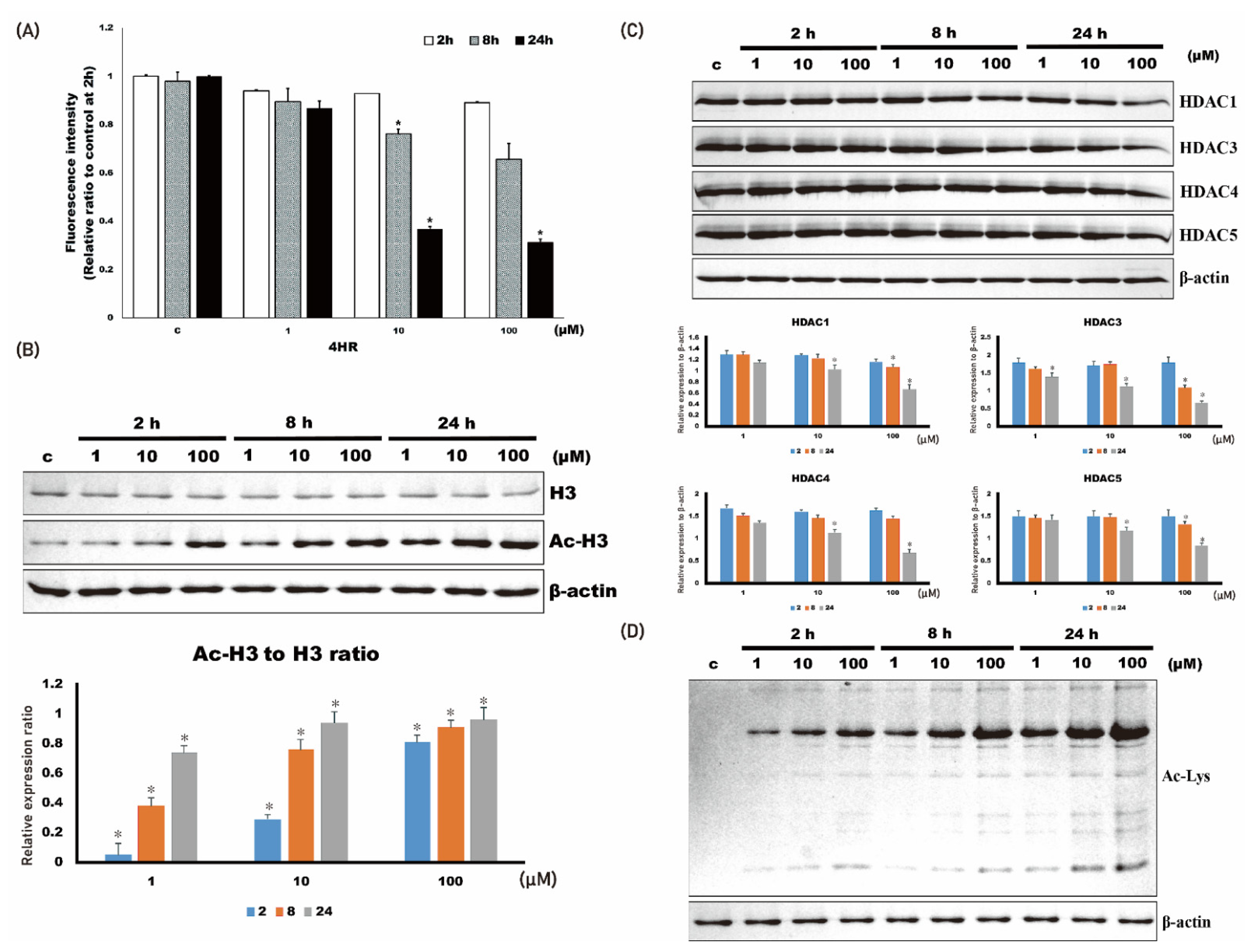
2.1. 4HR Administration Inhibits Class I HDAC Activity and Increases the Acetylation Level of Cellular Proteins
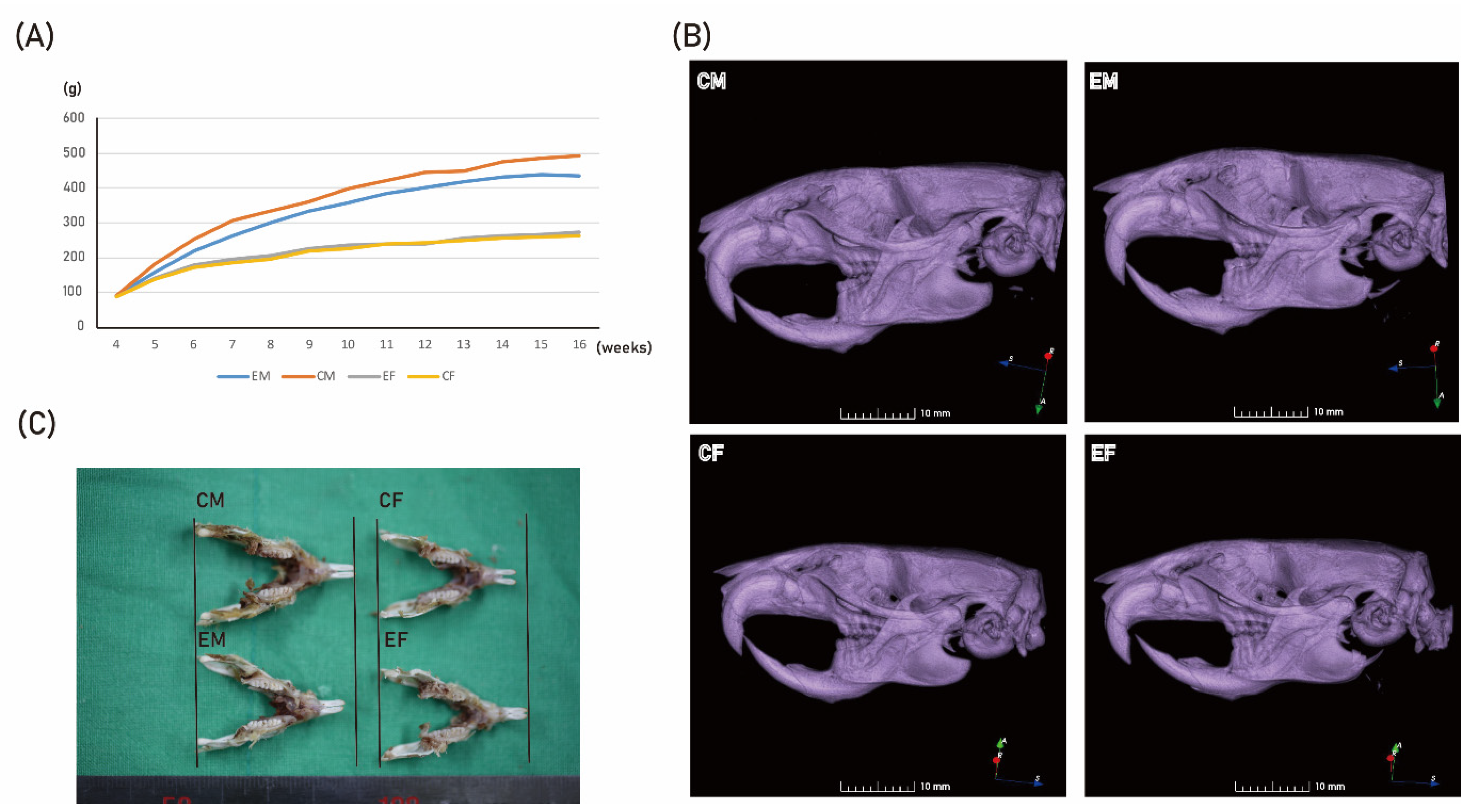
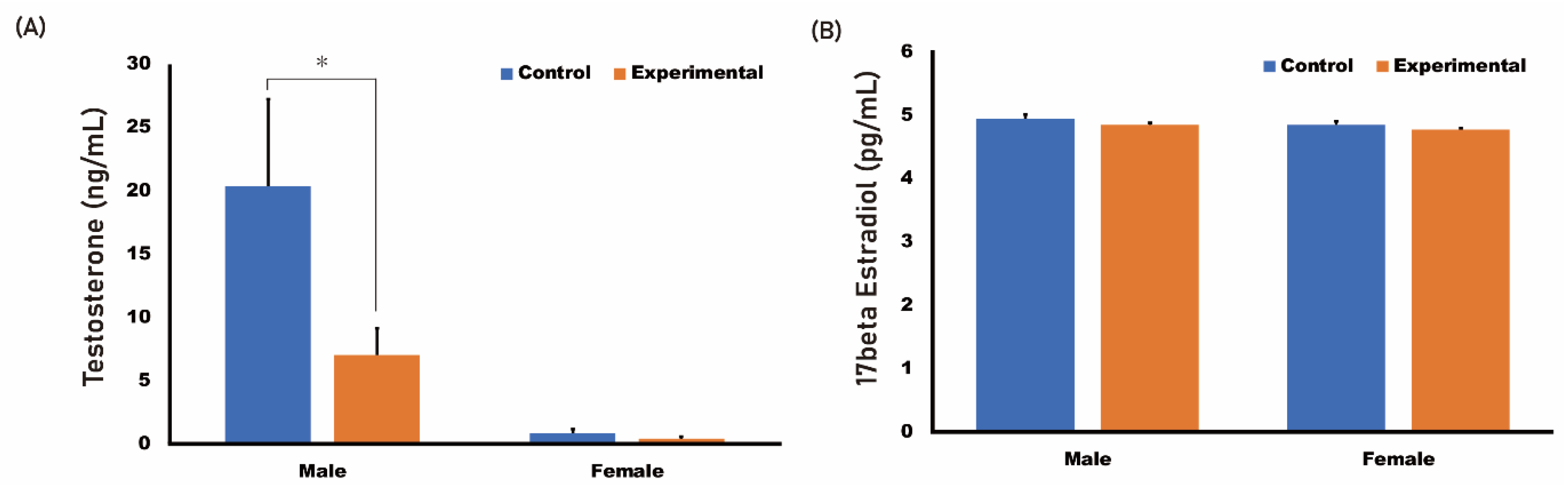
2.2. HR Administration Decreased Serum Testosterone Level and Mandibular Size in Male Rats
3. Discussion
4. Materials and Methods
4.1. Saos-2 Cell Culture
4.2. Western Blot and HDAC Inhibitory Assay
4.3. ATP, Mitochondrial Membrane Potential (MMP), and Oxygen-Consumption Assay
4.4. Immunoprecipitation High-Performance Liquid Chromatography (IP-HPLC)
4.5. Animal Experiments
4.6. Microcomputerized Tomography (mCT)
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fraga, M.; Ballestar, E.; Paz, M.F.; Ropero, S.; Setien, F.; Ballestar, M.L.; Suñer, D.H.; Cigudosa, J.C.; Urioste, M.; Benitez, J.; et al. From the cover: Epigenetic differences arise during the lifetime of monozygotic twins. Proc. Natl. Acad. Sci. USA 2005, 102, 10604–10609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavalli, G.; Heard, E. Advances in epigenetics link genetics to the environment and disease. Nat. Cell Biol. 2019, 571, 489–499. [Google Scholar] [CrossRef] [Green Version]
- Karagiannis, D.; Rampias, T. HDAC inhibitors: Dissecting mechanisms of action to counter tumor heterogeneity. Cancers 2021, 13, 3575. [Google Scholar] [CrossRef]
- Aka, J.A.; Kim, G.-W.; Yang, X.-J. K-acetylation and its enzymes: Overview and new developments. In Histone Deacetylases: The Biology and Clinical Implication; Yao, T.P., Seto, E., Eds.; Springer: New York, NY, USA, 2011; Volume 206, pp. 1–12. [Google Scholar]
- Yang, Y. Skeletal morphogenesis and embryonic development. Crit. Rev. Eukaryot. Gene Expr. 2009, 19, 1–10. [Google Scholar] [CrossRef]
- Glozak, M.A.; Sengupta, N.; Zhang, X.; Seto, E. Acetylation and deacetylation of non-histone proteins. Gene 2005, 363, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Lagger, G.; O’Carroll, D.; Rembold, M.; Khier, H.; Tischler, J.; Weitzer, G.; Schuettengruber, B.; Hauser, C.; Brunmeir, R.; Jenuwein, T.; et al. Essential function of histone deacetylase 1 in proliferation control and CDK inhibitor repression. EMBO J. 2002, 21, 2672–2681. [Google Scholar] [CrossRef] [Green Version]
- Zimmermann, S.; Kiefer, F.; Prudenziati, M.; Spiller, C.; Hansen, J.; Floss, T.; Wurst, W.; Minucci, S.; Göttlicher, M. Reduced body size and decreased intestinal tumor rates in HDAC2-mutant mice. Cancer Res. 2007, 67, 9047–9054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vega, R.B.; Matsuda, K.; Oh, J.; Barbosa, A.C.; Yang, X.; Meadows, E.; McAnally, J.; Pomajzl, C.; Shelton, J.M.; Richardson, J.A.; et al. Histone deacetylase 4 controls chondrocyte hypertrophy during skeletogenesis. Cell 2004, 119, 555–566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haberland, M.; Mokalled, M.H.; Montgomery, R.L.; Olson, E.N. Epigenetic control of skull morphogenesis by histone deacetylase 8. Genes Dev. 2009, 23, 1625–1630. [Google Scholar] [CrossRef] [Green Version]
- Bondarev, A.D.; Attwood, M.M.; Jonsson, J.; Chubarev, V.N.; Tarasov, V.V.; Schiöth, H.B. Recent developments of HDAC inhibitors: Emerging indications and novel molecules. Br. J. Clin. Pharmacol. 2021, 15, 1–21. [Google Scholar]
- Schroeder, T.M.; Westendorf, J.J. Histone deacetylase inhibitors promote osteoblast maturation. J. Bone Miner. Res. 2005, 20, 2254–2263. [Google Scholar] [CrossRef]
- Cho, H.H.; Park, H.T.; Kim, Y.J.; Bae, Y.C.; Suh, K.T.; Jung, J.S. Induction of osteogenic differentiation of human mesenchymal stem cells by histone deacetylase inhibitors. J. Cell. Biochem. 2005, 96, 533–542. [Google Scholar] [CrossRef]
- Yi, T.; Baek, J.-H.; Kim, H.-J.; Choi, M.-H.; Seo, S.-B.; Ryoo, H.-M.; Kim, G.-S.; Woo, K.M. Trichostatin A-mediated upregulation of p21WAF1 contributes to osteoclast apoptosis. Exp. Mol. Med. 2007, 39, 213–221. [Google Scholar] [CrossRef] [Green Version]
- Jeon, E.-J.; Lee, K.-Y.; Choi, N.-S.; Lee, M.-H.; Kim, H.-N.; Jin, Y.-H.; Ryoo, H.-M.; Choi, J.-Y.; Yoshida, M.; Nishino, N.; et al. Bone morphogenetic protein-2 stimulates Runx2 acetylation. J. Biol. Chem. 2006, 281, 16502–16511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheth, R.D.; Wesolowski, C.; Jacob, J.; Penney, S.; Hobbs, G.R.; Riggs, J.E.; Bodensteiner, J.B. Effect of carbamazepine and valproate on bone mineral density. J. Pediatr. 1995, 127, 256–262. [Google Scholar] [CrossRef]
- Boluk, A.; Guzelipek, M.; Savli, H.; Temel, I.; Ozişik, H.I.; Kaygusuz, A. The effect of valproate on bone mineral density in adult epileptic patients. Pharmacol. Res. 2004, 50, 93–97. [Google Scholar] [CrossRef]
- Elliott, J.O.; Jacobson, M.P.; Haneef, Z. Homocysteine and bone loss in epilepsy. Seizure 2007, 16, 22–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vestergaard, P.; Rejnmark, L.; Mosekilde, L. Fracture risk associated with use of antiepileptic drugs. Epilepsia 2004, 45, 1330–1337. [Google Scholar] [CrossRef]
- Kozubek, A.; Tyman, J.H.P. Resorcinolic lipids, the natural non-isoprenoid phenolic amphiphiles and their biological activity. Chem. Rev. 1998, 99, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-H.; Kim, J.-M.; Lee, J.S.; Gang, S.-R.; Lim, H.-S.; Kim, M.; Lee, O.-H. Development and validation of an analytical method for the determination of 4-hexylresorcinol in food. Food Chem. 2016, 190, 1086–1092. [Google Scholar] [CrossRef]
- Kim, M.-K.; Kim, S.-G.; Lee, S.K. 4-hexylresorcinol induced angiogenesis potential in human endothelial cells. Maxillofac. Plast. Reconstr. Surg. 2020, 42, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-Y.; Kweon, H.-Y.; Kim, D.-W.; Choi, J.-Y.; Kim, S.-G. 4-hexylresorcinol inhibits class I histone deacetylases in human umbilical cord endothelial cells. Appl. Sci. 2021, 11, 3486. [Google Scholar] [CrossRef]
- Amadasi, A.; Mozzarelli, A.; Meda, C.; Maggi, A.; Cozzini, P. Identification of xenoestrogens in food additives by an integrated in silico and in vitro approach. Chem. Res. Toxicol. 2008, 22, 52–63. [Google Scholar] [CrossRef] [Green Version]
- Kang, Y.-J.; Oh, J.-H.; Seok, H.; Jo, Y.-Y.; Kim, D.-W.; Garagiola, U.; Choi, J.-Y.; Kim, S.-G. 4-hexylresorcinol exhibits different characteristics to estrogen. Appl. Sci. 2020, 10, 1737. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.-G.; Jeong, J.-H.; Choi, J.-Y.; Kweon, H. 4-hexylresorcinol stimulates the differentiation of SCC-9 cells through the suppression of E2F2, E2F3 and Sp3 expression and the promotion of Sp1 expression. Oncol. Rep. 2012, 28, 677–681. [Google Scholar] [CrossRef] [Green Version]
- Ahn, J.; Kim, S.-G.; Kim, M.-K.; Kim, D.-W.; Lee, J.-H.; Seok, H.; Choi, J.-Y. Topical delivery of 4-hexylresorcinol promotes wound healing via tumor necrosis factor-α suppression. Burns 2016, 42, 1534–1541. [Google Scholar] [CrossRef]
- Heywood, L.H. Testosterone levels in the male laboratory rat: Variation under experimental conditions. Int. J. Androl. 1980, 3, 519–529. [Google Scholar] [CrossRef]
- Fujita, T.; Ohtani, J.; Shigekawa, M.; Kawata, T.; Kaku, M.; Kohno, S.; Tsutsui, K.; Tenjo, K.; Motokawa, M.; Tohma, Y.; et al. Effects of sex hormone disturbances on craniofacial growth in newborn mice. J. Dent. Res. 2004, 83, 250–254. [Google Scholar] [CrossRef]
- Armada, L.; Nogueira, C.; Neves, U.; Souza, P.S.; Detogne, J.; Armada-Dias, L.; Moreira, R.; Nascimento-Saba, C. Mandible analysis in sex steroid-deficient rats. Oral Dis. 2006, 12, 181–186. [Google Scholar] [CrossRef]
- Armada, L.; Brasil, S.D.C.; Dias, L.; Bezerra, J.; Pereira, R.M.R.; Takayama, L.; Dos Santos, R.M.M.; Gonçalves, L.S.; Nascimento-Saba, C.C.A. Effects of aging, gender, and hypogonadism on mandibular bone density. J. Investig. Clin. Dent. 2018, 9, e12310. [Google Scholar] [CrossRef]
- He, S.; Hartsfield, J.; Guo, Y.; Cao, Y.; Wang, S.; Chen, S. Association between CYP19A1 genotype and pubertal sagittal jaw growth. Am. J. Orthod. Dentofac. Orthop. 2012, 142, 662–670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noda, K.; Chang, H.-P.; Takahashi, I.; Kinoshita, Z.; Kawamoto, T. Effects of the anabolic steroid nandrolone phenylpropionate on craniofacial growth in rats. J. Morphol. 1994, 220, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.-H.; Chai, J.H.; Chang, S.-Y.; Kim, S.A. Acute valproate exposure induces sex-specific changes in steroid hormone metabolism in the cerebral cortex of juvenile mice. Neurochem. Res. 2020, 45, 2044–2051. [Google Scholar] [CrossRef] [PubMed]
- Taubøll, E.; Gregoraszczuk, E.L.; Kołodziej, A.; Kajta, M.; Ropstad, E. Valproate inhibits the conversion of testosterone to estradiol and acts as an apoptotic agent in growing porcine ovarian follicular cells. Epilepsia 2003, 44, 1014–1021. [Google Scholar] [CrossRef]
- Murray, E.K.; Hien, A.; de Vries, G.; Forger, N.G. Epigenetic control of sexual differentiation of the bed nucleus of the stria terminalis. Endocrinology 2009, 150, 4241–4247. [Google Scholar] [CrossRef]
- Gamez-Del-Estal, M.M.; Contreras, I.; Prieto-Pérez, R.; Ruiz-Rubio, M. Epigenetic effect of testosterone in the behavior of C. elegans. A clue to explain androgen-dependent autistic traits? Front. Cell. Neurosci. 2014, 8, 1–12. [Google Scholar]
- Itzhak, Y.; Liddie, S.; Anderson, K.L. Sodium butyrate-induced histone acetylation strengthens the expression of cocaine-associated contextual memory. Neurobiol. Learn. Mem. 2013, 102, 34–42. [Google Scholar] [CrossRef] [Green Version]
- Mendonça, M.L.; Pereira, F.A.; Nogueira-Barbosa, M.H.; Monsignore, L.M.; Teixeira, S.R.; Watanabe, P.C.; Maciel, L.M.; De Paula, F.J. Increased vertebral morphometric fracture in patients with postsurgical hypoparathyroidism despite normal bone mineral density. BMC Endocr. Disord. 2013, 13, 1. [Google Scholar] [CrossRef]
- Hassona, Y.; Rajab, L.; Taimeh, D.; Scully, C. Sanjad-Sakati Syndrome: Oral health care. Med. Princ. Pr. 2018, 27, 293–296. [Google Scholar] [CrossRef] [Green Version]
- Shimizu, E.; Selvamurugan, N.; Westendorf, J.J.; Olson, E.N.; Partridge, N. HDAC4 represses matrix metalloproteinase-13 transcription in osteoblastic cells, and parathyroid hormone controls this repression. J. Biol. Chem. 2010, 285, 9616–9626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutiérrez, S.J.; Gomez, M.; Rey, J.A.; Ochoa, M.; Prieto, J.C. Polymorphisms of the noggin gene and mandibular micrognathia: A first approximation. AOL Acta Odontol. Latinoam. 2010, 23, 13–19. [Google Scholar] [PubMed]
- Wójcik, D.; Beń-Skowronek, I. Craniofacial morphology in children with growth hormone deficiency and turner syndrome. Diagnostics 2020, 10, 88. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Zhu, W.; Chen, L.; Liang, L. Early growth hormone intervention improves glucose metabolism in adult rats born small for gestational age. Exp. Clin. Endocrinol. Diabetes 2018, 128, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Villalba, P.; Jiménez-Lara, A.M.; Castillo, A.I.; Aranda, A. Histone acetylation influences thyroid hormone and retinoic acid-mediated gene expression. DNA Cell Biol. 1997, 16, 421–431. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Males (EM) | Control Males (CM) | Experimental Females (EF) | Control Females (CF) | |||||
|---|---|---|---|---|---|---|---|---|
| Mean | SE | Mean | SE | Mean | SE | Mean | SE | |
| Total mandibular length I | 27.39 | 0.16 | 27.86 | 0.18 | 25.70 * | 0.18 | 24.94 | 0.09 |
| Total mandibular length II | 25.69 * | 0.19 | 26.35 | 0.17 | 24.18 * | 0.17 | 23.61 | 0.11 |
| Total mandibular length III | 23.63 * | 0.17 | 24.19 | 0.13 | 21.92 * | 0.15 | 21.41 | 0.10 |
| Corpus length I | 27.81 * | 0.16 | 28.54 | 0.20 | 26.47 * | 0.21 | 25.73 | 0.16 |
| Corpus length II | 24.42 * | 0.19 | 25.43 | 0.22 | 23.37 | 0.19 | 22.91 | 0.18 |
| Corpus length III | 21.26 * | 0.15 | 22.20 | 0.16 | 20.09 | 0.17 | 19.88 | 0.24 |
| Corpus length IV | 20.06 * | 0.14 | 20.80 | 0.17 | 18.92 | 0.17 | 18.52 | 0.14 |
| Ramus height I | 9.48 | 0.06 | 9.39 | 0.08 | 8.59 * | 0.07 | 8.19 | 0.09 |
| Ramus height II | 14.75 | 0.08 | 15.13 | 0.13 | 13.27 | 0.10 | 12.98 | 0.09 |
| Ramus height III | 12.41 | 0.11 | 12.74 | 0.09 | 11.56 | 0.07 | 11.40 | 0.08 |
| Bicondylior mandibular width | 19.70 | 0.16 | 19.80 | 0.10 | 18.00 | 0.33 | 18.04 | 0.09 |
| Bigonial mandibular width | 21.75 * | 0.21 | 22.34 | 0.14 | 18.84 | 0.41 | 18.51 | 0.37 |
| Maxillary molar width | 9.58 | 0.05 | 9.61 | 0.06 | 9.28 | 0.16 | 9.30 | 0.08 |
| Zygomatic width | 25.27 | 0.12 | 25.53 | 0.25 | 23.66 | 0.21 | 23.10 | 0.19 |
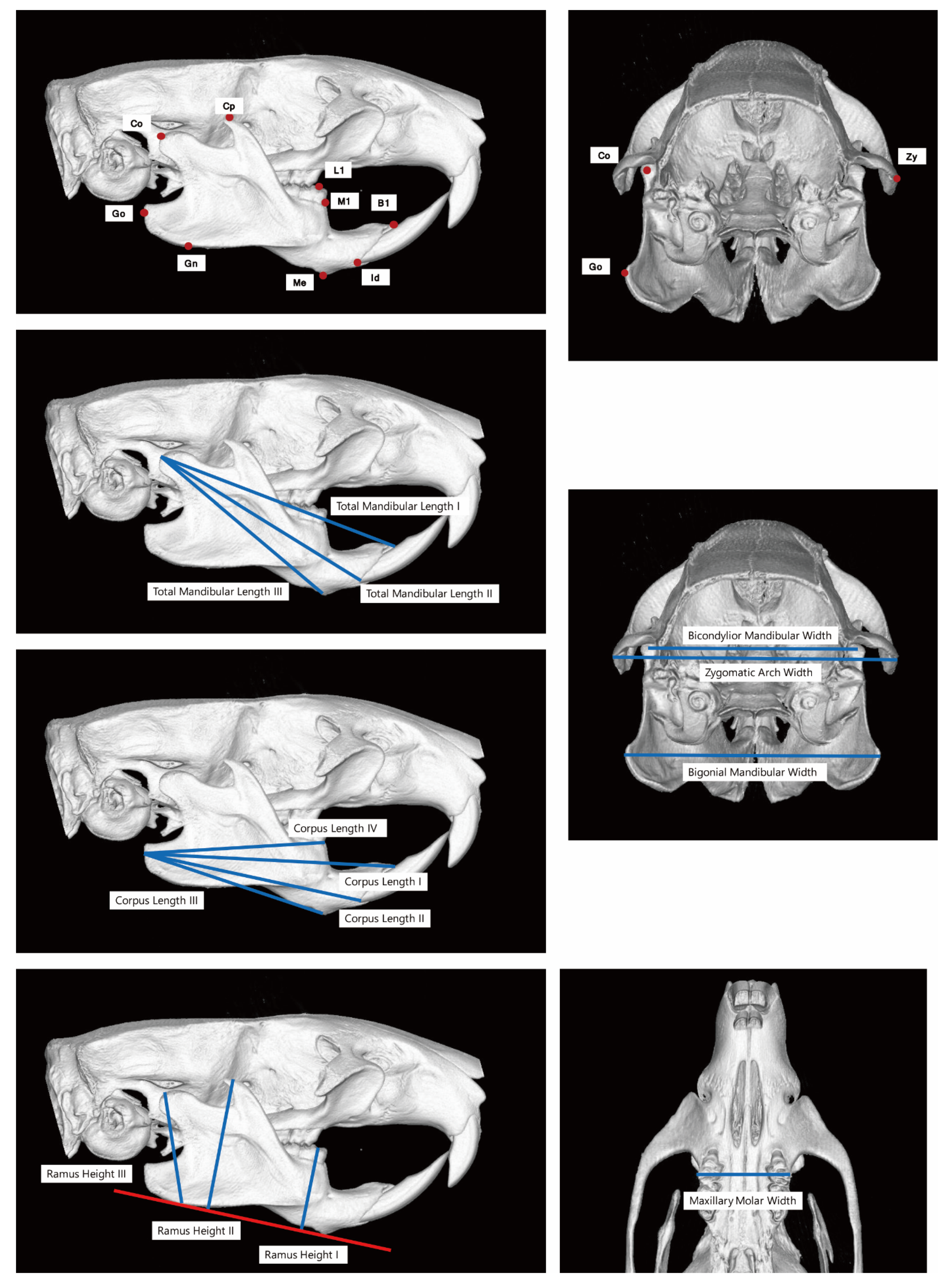
| Condylion | Co | Most posterosuperior point of condylar process |
| Gonion | Go | Most posterior point of angular process of mandible |
| Gnathion | Gn | Point on most inferior contour of angular process of mandible |
| Menton | Me | Point on most inferior contour of lower border of mandible, adjacent to incisors |
| Infradentale | Id | Most inferior point of the marginal alveolar bone of the lower central incisor |
| M1 | Point on intersection between the mandibular alveolar bone and mesial surface of first molar | |
| B1 | Point on intersection between lingual surface of lower incisor and anteriormost part of lingual alveolar bone | |
| L1 | Point on mesial occlusal fossa of lower first molar | |
| L1’ | Crossing point on Me–Gn perpendicular to Me–Gn from L1 | |
| L2 | Most external point on buccal surface of lower first molar | |
| Coronoid Process | Cp | Point on most superior contour of coronoid process |
| Cp’ | Crossing point on Me–Gn perpendicular to Me–Gn from Cp | |
| Zygion | Zy | Most external point of the zygomatic arch |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, I.-S.; Kim, D.-W.; Oh, J.-H.; Lee, S.K.; Choi, J.-Y.; Kim, S.-G.; Kim, T.-W. Effects of 4-Hexylresorcinol on Craniofacial Growth in Rats. Int. J. Mol. Sci. 2021, 22, 8935. https://doi.org/10.3390/ijms22168935
Lee I-S, Kim D-W, Oh J-H, Lee SK, Choi J-Y, Kim S-G, Kim T-W. Effects of 4-Hexylresorcinol on Craniofacial Growth in Rats. International Journal of Molecular Sciences. 2021; 22(16):8935. https://doi.org/10.3390/ijms22168935
Chicago/Turabian StyleLee, In-Song, Dae-Won Kim, Ji-Hyeon Oh, Suk Keun Lee, Je-Yong Choi, Seong-Gon Kim, and Tae-Woo Kim. 2021. "Effects of 4-Hexylresorcinol on Craniofacial Growth in Rats" International Journal of Molecular Sciences 22, no. 16: 8935. https://doi.org/10.3390/ijms22168935
APA StyleLee, I. -S., Kim, D. -W., Oh, J. -H., Lee, S. K., Choi, J. -Y., Kim, S. -G., & Kim, T. -W. (2021). Effects of 4-Hexylresorcinol on Craniofacial Growth in Rats. International Journal of Molecular Sciences, 22(16), 8935. https://doi.org/10.3390/ijms22168935