
5-HT Receptors and the Development of New Antidepressants


Abstract
:1. Introduction
2. The Serotonergic System and Depression
3. The 5-HT Receptors
3.1. The 5-HT1A Receptors
- Animal studies show that both the stimulation and blockade of 5-HT1A receptors can cause or accelerate the antidepressant effect. It is difficult not to associate this with the functional differences of 5-HT1A auto- and hetero-receptors and the phenomenon of a biased 5-HT1A agonism;
- A single transcription factor, Freud-1, has been found to be crucial for the expression of the 5-HT1A autoreceptor. Targeting it may increase the response to antidepressant treatment;
- Observations on the 5-HT1A receptor population contributed to a fruitful search for potential multimodal antidepressants (vilazodone and vortioxetine) that incorporate 5-HT1A receptor activity into their mechanism of action.
3.2. The 5-HT1B Receptors
- Animal studies show that the involvement of 5-HT1B receptors in the pathophysiology of depression is partly related to their responsiveness to environmental stress as well as an exposure to antidepressants;
- The p11 protein improves 5-HT1B receptor function in various regions of the brain and contributes to an antidepressant-like effect in animal behavioral tests;
- The 5-HT1B heteroreceptors are involved in hippocampal neurogenesis, which may explain their importance for the antidepressant-like effect. The stimulation of postsynaptic receptors and the inhibition of presynaptic 5-HT1B receptors may be beneficial in the treatment of depression.
3.3. The 5-HT1D, 5-HT1E, and 5-HT1F Receptors
3.4. The 5-HT2A Receptors
- Many antidepressants and antipsychotic drugs have relatively high binding to 5-HT2A receptors;
- The high co-expression of 5-HT1A and 5-HT2A receptors in the neocortex may indicate that the blockade of 5-HT2A receptors enhances 5-HT1A receptor-mediated neurotransmission in the cortical and limbic regions, an activity associated with antidepressant efficacy;
- Increasing evidence shows that 5-HT2A receptor antagonists display antidepressant effects. A selective 5-HT2A receptor blockade may complement the behavioral effects of serotonin transporter inhibition.
3.5. The 5-HT2B Receptors
- 5-HT2B receptors directly and positively regulate the activity of serotonin neurons;
- There are reports of the antidepressant properties of selective 5-HT2B receptor agonists;
- Potential new antidepressants acting on 5-HT2B receptors may adversely affect the function of the respiratory and circulatory systems.
3.6. The 5-HT2C Receptors
- Several classes of antidepressants have an affinity for 5-HT2C receptors. Alterations in their functional status have been observed in depressive and anxiety states;
- Both 5-HT2C agonists and antagonists have been shown to be active in animal models of depression. Preclinical data show that the antagonism of 5-HT2C receptors increases the neurochemical and behavioral effects of SSRIs;
- There is still a need to further define the role of 5-HT2C receptor subtype in depression.
3.7. The 5-HT3 Receptors
- Activity towards other neurotransmission systems is the main mechanism of action for 5-HT3 receptor ligands;
- Rodent experiments show that the antidepressant-like effect should be attributed to postsynaptic rather than presynaptic 5-HT3 antagonism, since the presynaptic and somatodendritic 5-HT3 receptor blockade reduces serotonin levels;
- Some antidepressants with different mechanisms of action exhibit functional 5-HT3 receptor antagonism. The vortioxetine antagonism at the 5-HT3 receptor may underlie its faster onset of action.
3.8. The 5-HT4 Receptors
- There are reports linking the 5-HT4 receptor with depressive disorders. The 5-HT4 receptor interacts with the p11 protein, which determines the antidepressant activity mediated by 5-HT1B and 5-HT4 receptors;
- 5-HT4 receptor signaling may modulate the function of the dentate gyrus of the hippocampus by increasing the neurogenesis and expression of neurotrophic factors, which may contribute to the antidepressant effects of drugs that enhance serotonergic transmission.
- The activation of the 5-HT4 receptor may be a useful adjunct to antidepressant therapy, both to accelerate the onset of clinical antidepressant effects and to target cognitive symptoms that are not effectively treated with current therapies.
3.9. The 5-HT6 Receptors
- The distribution of the 5-HT6 receptor in the limbic and cortical regions of the brain may suggest that 5-HT6 receptors play an important role in the pathogenesis and/or treatment of depression;
- Several tricyclic and atypical antidepressants exhibit nanomolar 5-HT6 receptor binding. The inhibition of the 5-HT6 receptor potentiates the effects of clinically used antidepressants. This synergistic effect is interesting in the search for a multimodal antidepressant therapy with minimized side effects or a faster onset of action;
- It is not exactly clear what functional 5-HT6 receptor ligand profile (antagonism or agonism) will be more beneficial in the treatment of depression. Moreover, the exact mechanism by which 5-HT6 ligands induce antidepressant effects is unknown and may include effects on other neurotransmission systems.
3.10. The 5-HT7 Receptors
- Several antidepressants have been found to have a high affinity for the 5-HT7 receptor, leading to much further research into its antidepressant activity;
- The antagonists of the 5-HT7 receptor might represent a new class of antidepressants with a faster therapeutic effect.
- Preclinical evidence suggests that vortioxetine has a relatively low affinity for the 5-HT7 receptor compared to other 5-HT receptor subtypes but inhibits its action with a rapid 5-HT7 receptor downregulation as an inverse agonist. Both the intravenous and oral administration of vortioxetine resulted in a significant improvement in depression and anxiety after 3 days.
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- James, S.L.; Abate, D.; Abate, K.H.; Abay, S.M.; Abbafati, C.; Abbasi, N.; Abbastabar, H.; Abd-Allah, F.; Abdela, J.; Abdelalim, A.; et al. Global, regional, and national incidence, prevalence, and years lived with disability for 354 diseases and injuries for 195 countries and territories, 1990–2017: A systematic analysis for the Global Burden of Disease Study 2017. Lancet 2018, 392, 1789–1858. [Google Scholar] [CrossRef] [Green Version]
- Ögren, S.O.; Eriksson, T.M.; Elvander-Tottie, E.; D’Addario, C.; Ekström, J.C.; Svenningsson, P.; Meister, B.; Kehr, J.; Stiedl, O. The role of 5-HT1A receptors in learning and memory. Behav. Brain Res. 2008, 195, 54–77. [Google Scholar] [CrossRef]
- Day, J.J.; Sweatt, J.D. Epigenetic Treatments for Cognitive Impairments. Neuropsychopharmacology 2012, 37, 247–260. [Google Scholar] [CrossRef]
- Sheehan, D.V.; Nakagome, K.; Asami, Y.; Pappadopulos, E.A.; Boucher, M. Restoring function in major depressive disorder: A systematic review. J. Affect. Disord. 2017, 215, 299–313. [Google Scholar] [CrossRef]
- Sharp, T.; Cowen, P.J. 5-HT and depression: Is the glass half-full? Curr. Opin. Pharmacol. 2011, 11, 45–51. [Google Scholar] [CrossRef]
- Wiles, N.J.; Fischer, K.; Cowen, P.; Nutt, D.; Peters, T.J.; Lewis, G.; White, I.R. Allowing for non-adherence to treatment in a randomized controlled trial of two antidepressants (citalopram versus reboxetine): An example from the GENPOD trial. Psychol. Med. 2014, 44, 2855–2866. [Google Scholar] [CrossRef] [Green Version]
- Turcotte, J.E.; Debonnel, G.; De Montigny, C.; Hébert, C.; Blier, P. Assessment of the serotonin and norepinephrine reuptake blocking properties of duloxetine in healthy subjects. Neuropsychopharmacology 2001, 24, 511–521. [Google Scholar] [CrossRef] [Green Version]
- Cleare, A.; Pariante, C.; Young, A.; Anderson, I.; Christmas, D.; Cowen, P.; Dickens, C.; Ferrier, I.; Geddes, J.; Gilbody, S.; et al. Evidence-based guidelines for treating depressive disorders with antidepressants: A revision of the 2008 British Association for Psychopharmacology guidelines. J. Psychopharmacol. 2015, 29, 459–525. [Google Scholar] [CrossRef]
- Bauer, M.; Bschor, T.; Pfennig, A.; Whybrow, P.C.; Angst, J.; Versiani, M.; Möller, H.-J. World Federation of Societies of Biological Psychiatry (WFSBP) Guidelines for Biological Treatment of Unipolar Depressive Disorders in Primary Care. World J. Biol. Psychiatry 2007, 8, 67–104. [Google Scholar] [CrossRef]
- Harmer, C.J.; Duman, R.S.; Cowen, P.J. How do antidepressants work? New perspectives for refining future treatment approaches. Lancet Psychiatry 2017, 4, 409–418. [Google Scholar] [CrossRef] [Green Version]
- Rana, T.; Behl, T.; Sehgal, A.; Mehta, V.; Singh, S.; Kumar, R.; Bungau, S. Integrating Endocannabinoid Signalling In Depression. J. Mol. Neurosci. 2021. [Google Scholar] [CrossRef]
- Rana, T.; Behl, T.; Sehgal, A.; Mehta, V.; Singh, S.; Sharma, N.; Bungau, S. Elucidating the Possible Role of FoxO in Depression. Neurochem. Res. 2021. [Google Scholar] [CrossRef] [PubMed]
- Rana, T.; Behl, T.; Sehgal, A.; Sachdeva, M.; Mehta, V.; Sharma, N.; Singh, S.; Bungau, S. Exploring Sonic Hedgehog Cell Signaling in Neurogenesis: Its Potential Role in Depressive Behavior. Neurochem. Res. 2021, 46, 1589–1602. [Google Scholar] [CrossRef] [PubMed]
- Rana, T.; Behl, T.; Sehgal, A.; Srivastava, P.; Bungau, S. Unfolding the Role of BDNF as a Biomarker for Treatment of Depression. J. Mol. Neurosci. 2020. [Google Scholar] [CrossRef]
- Millan, M.J. On “polypharmacy” and multi-target agents, complementary strategies for improving the treatment of depression: A comparative appraisal. Int. J. Neuropsychopharmacol. 2014, 17, 1009–1037. [Google Scholar] [CrossRef]
- Duhr, F.; Déléris, P.; Raynaud, F.; Séveno, M.; Morisset-Lopez, S.; Mannoury la Cour, C.; Millan, M.J.; Bockaert, J.; Marin, P.; Chaumont-Dubel, S. Cdk5 induces constitutive activation of 5-HT6 receptors to promote neurite growth. Nat. Chem. Biol. 2014, 10, 590–597. [Google Scholar] [CrossRef] [Green Version]
- Żmudzka, E.; Sałaciak, K.; Sapa, J.; Pytka, K. Serotonin receptors in depression and anxiety: Insights from animal studies. Life Sci. 2018, 210, 106–124. [Google Scholar] [CrossRef]
- Miller, J.M.; Hesselgrave, N.; Ogden, R.T.; Zanderigo, F.; Oquendo, M.A.; Mann, J.J.; Parsey, R.V. Brain Serotonin 1A Receptor Binding as a Predictor of Treatment Outcome in Major Depressive Disorder. Biol. Psychiatry 2013, 74, 760–767. [Google Scholar] [CrossRef] [Green Version]
- Kaufman, J.; DeLorenzo, C.; Choudhury, S.; Parsey, R.V. The 5-HT1A receptor in Major Depressive Disorder. Eur. Neuropsychopharmacol. 2016, 26, 397–410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Artigas, F. Serotonin receptors involved in antidepressant effects. Pharmacol. Ther. 2013, 137, 119–131. [Google Scholar] [CrossRef] [Green Version]
- Artigas, F. Developments in the field of antidepressants, where do we go now? Eur. Neuropsychopharmacol. 2015, 25, 657–670. [Google Scholar] [CrossRef]
- Perez-Caballero, L.; Torres-Sanchez, S.; Romero-López-Alberca, C.; González-Saiz, F.; Mico, J.A.; Berrocoso, E. Monoaminergic system and depression. Cell Tissue Res. 2019, 377, 107–113. [Google Scholar] [CrossRef]
- Albert, P.R.; Vahid-Ansari, F. The 5-HT1A receptor: Signaling to behavior. Biochimie 2018, 161, 34–45. [Google Scholar] [CrossRef]
- Haleem, D.J. Targeting Serotonin1A Receptors for Treating Chronic Pain and Depression. Curr. Neuropharmacol. 2019, 17, 1098–1108. [Google Scholar] [CrossRef]
- Artigas, F.; Bortolozzi, A.; Celada, P. Can we increase speed and efficacy of antidepressant treatments? Part I: General aspects and monoamine-based strategies. Eur. Neuropsychopharmacol. 2018, 28, 445–456. [Google Scholar] [CrossRef]
- Kishi, T.; Meltzer, H.Y.; Matsuda, Y.; Iwata, N. Azapirone 5-HT1A receptor partial agonist treatment for major depressive disorder: Systematic review and meta-analysis. Psychol. Med. 2014, 44, 2255–2269. [Google Scholar] [CrossRef] [Green Version]
- Sahli, Z.T.; Banerjee, P.; Tarazi, F.I. The Preclinical and Clinical Effects of Vilazodone for the Treatment of Major Depressive Disorder. Expert Opin. Drug Discov. 2016, 11, 515–523. [Google Scholar] [CrossRef] [Green Version]
- James Deardorff, W.; Grossberg, G.T. A review of the clinical efficacy, safety and tolerability of the antidepressants vilazodone, levomilnacipran and vortioxetine. Expert Opin. Pharmacother. 2014, 15, 2525–2542. [Google Scholar] [CrossRef]
- Jonnakuty, C.; Gragnoli, C. What do we know about serotonin? J. Cell. Physiol. 2008, 217, 301–306. [Google Scholar] [CrossRef]
- Banerjee, P.; Mehta, M.; Kanjilal, B. The 5-HT1A Receptor: A Signaling Hub Linked to Emotional Balance; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2007; Chapter 7; ISBN 0849339774. [Google Scholar]
- Hasegawa, H.; Nakamura, K. Tryptophan Hydroxylase and Serotonin Synthesis Regulation. In Handbook of Behavioral Neuroscience; Elsevier B.V.: Amsterdam, The Netherlands, 2010; Volume 21, pp. 183–202. ISBN 9780123746344. [Google Scholar]
- Manocha, M.; Khan, W.I. Serotonin and GI Disorders: An Update on Clinical and Experimental Studies. Clin. Transl. Gastroenterol. 2012, 3, e13. [Google Scholar] [CrossRef]
- Savitz, J.; Lucki, I.; Drevets, W.C. 5-HT1A receptor function in major depressive disorder. Prog. Neurobiol. 2009, 88, 17–31. [Google Scholar] [CrossRef] [Green Version]
- Smythies, J. Section V. Serotonin System. Int. Rev. Neurobiol. 2005, 64, 217–268. [Google Scholar] [CrossRef]
- Diksic, M.; Young, S.N. Study of the brain serotonergic system with labeled α-methyl-l-tryptophan. J. Neurochem. 2001, 78, 1185–1200. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, B.L.; Azmitia, E.C. Structure and function of the brain serotonin system. Physiol. Rev. 1992, 72, 165–229. [Google Scholar] [CrossRef] [Green Version]
- Celada, P.; Puig, M.V.; Casanovas, J.M.; Guillazo, G.; Artigas, F. Control of dorsal raphe serotonergic neurons by the medial prefrontal cortex: Involvement of serotonin-1A, GABA(A), and glutamate receptors. J. Neurosci. 2001, 21, 9917–9929. [Google Scholar] [CrossRef] [Green Version]
- O’Leary, O.F.; Bechtholt, A.J.; Crowley, J.J.; Valentino, R.J.; Lucki, I. The role of noradrenergic tone in the dorsal raphe nucleus of the mouse in the acute behavioral effects of antidepressant drugs. Eur. Neuropsychopharmacol. 2007, 17, 215–226. [Google Scholar] [CrossRef]
- Varga, V.; Székely, A.D.; Csillag, A.; Sharp, T.; Hajós, M. Evidence for a role of GABA interneurones in the cortical modulation of midbrain 5-hydroxytryptamine neurones. Neuroscience 2001, 106, 783–792. [Google Scholar] [CrossRef]
- Martín-Ruiz, R.; Ugedo, L.; Honrubia, M.A.; Mengod, G.; Artigas, F. Control of serotonergic neurons in rat brain by dopaminergic receptors outside the dorsal raphe nucleus. J. Neurochem. 2001, 77, 762–775. [Google Scholar] [CrossRef] [Green Version]
- Ohno, Y. Serotonin Receptors as the Therapeutic Target for Central Nervous System Disorders; Elsevier Inc.: Amsterdam, The Netherlands, 2019; ISBN 9780128000502. [Google Scholar]
- Santana, N.; Bortolozzi, A.; Serrats, J.; Mengod, G.; Artigas, F. Expression of serotonin1A and serotonin2A receptors in pyramidal and GABAergic neurons of the rat prefrontal cortex. Cereb. Cortex 2004, 14, 1100–1109. [Google Scholar] [CrossRef] [Green Version]
- Riad, M.; Garcia, S.; Watkins, K.C.; Jodoin, N.; Doucet, É.; Langlois, X.; El Mestikawy, S.; Hamon, M.; Descarries, L. Somatodendritic localization of 5-HT1A and preterminal axonal localization of 5-HT1B serotonin receptors in adult rat brain. J. Comp. Neurol. 2000, 417, 181–194. [Google Scholar] [CrossRef]
- Liu, Y.F.; Ghahremani, M.H.; Rasenick, M.M.; Jakobs, K.H.; Albert, P.R. Stimulation of cAMP synthesis by G(i)-coupled receptors upon ablation of distinct Gα(i) protein expression. G(i) subtype specificity of the 5-HT1A receptor. J. Biol. Chem. 1999, 274, 16444–16450. [Google Scholar] [CrossRef] [Green Version]
- Loucif, A.J.C.; Bonnavion, P.; Macri, B.; Golmard, J.L.; Boni, C.; Melfort, M.; Leonard, G.; Lesch, K.P.; Adrien, J.; Jacquin, T.D. Gender-dependent regulation of G-protein-gated inwardly rectifying potassium current in dorsal raphe neurons in knock-out mice devoid of the 5-hydroxytryptamine transporter. J. Neurobiol. 2006, 66, 1475–1488. [Google Scholar] [CrossRef]
- Halasy, K.; Miettinen, R.; Szabat, E.; Freund, T.F. GABAergic Interneurons are the Major Postsynaptic Targets of Median Raphe Afferents in the Rat Dentate Gyrus. Eur. J. Neurosci. 1992, 4, 144–153. [Google Scholar] [CrossRef]
- Cassel, J.C.; Jeltsch, H. Serotonergic modulation of cholinergic function in the central nervous system: Cognitive implications. Neuroscience 1995, 69, 1–41. [Google Scholar] [CrossRef] [Green Version]
- Jeltsch-David, H.; Koenig, J.; Cassel, J.C. Modulation of cholinergic functions by serotonin and possible implications in memory: General data and focus on 5-HT1A receptors of the medial septum. Behav. Brain Res. 2008, 195, 86–97. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Gil, X.; Artigas, F.; Adell, A. Unraveling Monoamine Receptors Involved in the Action of Typical and Atypical Antipsychotics on Glutamatergic and Serotonergic Transmission in Prefrontal Cortex. Curr. Pharm. Des. 2010, 16, 502–515. [Google Scholar] [CrossRef] [Green Version]
- Di Matteo, V.; Di Giovanni, G.; Pierucci, M.; Esposito, E. Serotonin control of central dopaminergic function: Focus on in vivo microdialysis studies. Prog. Brain Res. 2008, 172, 7–44. [Google Scholar] [CrossRef]
- Andrade, R.; Nicoll, R.A. Pharmacologically distinct actions of serotonin on single pyramidal neurones of the rat hippocampus recorded in vitro. J. Physiol. 1987, 394, 99–124. [Google Scholar] [CrossRef]
- Newman-Tancredi, A. Biased agonism at serotonin 5-HT1A receptors: Preferential postsynaptic activity for improved therapy of CNS disorders. Neuropsychiatry 2011, 1, 149–164. [Google Scholar] [CrossRef]
- Mehta, M.; Ahmed, Z.; Fernando, S.S.; Cano-Sanchez, P.; Adayev, T.; Ziemnicka, D.; Wieraszko, A.; Banerjee, P. Plasticity of 5-HT1A receptor-mediated signaling during early postnatal brain development. J. Neurochem. 2007, 101, 918–928. [Google Scholar] [CrossRef]
- Kushwaha, N.; Albert, P.R. Coupling of 5-HT1A autoreceptors to inhibition of mitogen-activated protein kinase activation via Gβγ subunit signaling. Eur. J. Neurosci. 2005, 21, 721–732. [Google Scholar] [CrossRef]
- Heusler, P.; Pauwels, P.J.; Wurch, T.; Newman-Tancredi, A.; Tytgat, J.; Colpaert, F.C.; Cussac, D. Differential ion current activation by human 5-HT1A receptors in Xenopus oocytes: Evidence for agonist-directed trafficking of receptor signalling. Neuropharmacology 2005, 49, 963–976. [Google Scholar] [CrossRef]
- Valdizán, E.M.; Castro, E.; Pazos, A. Agonist-dependent modulation of G-protein coupling and transduction of 5-HT1A receptors in rat dorsal raphe nucleus. Int. J. Neuropsychopharmacol. 2010, 13, 835–843. [Google Scholar] [CrossRef] [Green Version]
- Heisler, L.K.; Chu, H.-M.; Brennan, T.J.; Danao, J.A.; Bajwa, P.; Parsons, L.H.; Tecott, L.H. Elevated anxiety and antidepressant-like responses in serotonin 5-HT1A receptor mutant mice. Proc. Natl. Acad. Sci. USA 1998, 95, 15049–15054. [Google Scholar] [CrossRef] [Green Version]
- Pytka, K.; Podkowa, K.; Rapacz, A.; Podkowa, A.; Zmudzka, E.; Olczyk, A.; Sapa, J.; Filipek, B. The role of serotonergic, adrenergic and dopaminergic receptors in antidepressant-like effect. Pharmacol. Rep. 2016, 68, 263–274. [Google Scholar] [CrossRef]
- Santarelli, L. Requirement of Hippocampal Neurogenesis for the Behavioral Effects of Antidepressants. Science 2003, 301, 805–809. [Google Scholar] [CrossRef] [Green Version]
- Assié, M.B.; Bardin, L.; Auclair, A.L.; Carilla-Durand, E.; Depoortère, R.; Koek, W.; Kleven, M.S.; Colpaert, F.; Vacher, B.; Newman-Tancredi, A. F15599, a highly selective post-synaptic 5-HT1A receptor agonist: In-vivo profile in behavioural models of antidepressant and serotonergic activity. Int. J. Neuropsychopharmacol. 2010, 13, 1285–1298. [Google Scholar] [CrossRef] [Green Version]
- Pytka, K.; Głuch-Lutwin, M.; Kotańska, M.; Żmudzka, E.; Jakubczyk, M.; Waszkielewicz, A.; Janiszewska, P.; Walczak, M. HBK-15 protects mice from stress-induced behavioral disturbances and changes in corticosterone, BDNF, and NGF levels. Behav. Brain Res. 2017, 333, 54–66. [Google Scholar] [CrossRef]
- Depoortère, R.; Auclair, A.L.; Newman-Tancredi, A. NLX-101, a highly selective 5-HT1A receptor biased agonist, mediates antidepressant-like activity in rats via prefrontal cortex 5-HT1A receptors. Behav. Brain Res. 2021, 401, 113082. [Google Scholar] [CrossRef]
- Albert, P.R.; Le François, B.; Millar, A.M. Transcriptional dysregulation of 5-HT1A autoreceptors in mental illness. Mol. Brain 2011, 4, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terao, T.; Ishii, N.; Hirakawa, H.; Aoshima, E. Is the bell-shaped dose-response curve of the selective serotonin reuptake inhibitor due to 5-HT1A auto-receptors? Med. Hypotheses 2020, 140, 109681. [Google Scholar] [CrossRef]
- Hervás, I.; Vilaró, M.T.; Romero, L.; Scorza, M.C.; Mengod, G.; Artigas, F. Desensitization of 5-HT(1A) autoreceptors by a low chronic fluoxetine dose effect of the concurrent administration of WAY-100635. Neuropsychopharmacology 2001, 24, 11–20. [Google Scholar] [CrossRef] [Green Version]
- Artigas, F.; Celada, P.; Laruelle, M.; Adell, A. How does pindolol improve antidepressant action? Trends Pharmacol. Sci. 2001, 22, 224–228. [Google Scholar] [CrossRef]
- Celada, P.; Artigas, F. Monoamine oxidase inhibitors increase preferentially extracellular 5-hydroxytryptamine in the midbrain raphe nuclei. A brain microdialysis study in the awake rat. Naunyn-Schmiedebergs Arch. Pharmacol. 1993, 347, 583–590. [Google Scholar] [CrossRef] [Green Version]
- Blier, P.; De Montigny, C. Current advances and trends in the treatment of depression. Trends Pharmacol. Sci. 1994, 15, 220–226. [Google Scholar] [CrossRef]
- Romero, L.; Bel, N.; Artigas, F.; De Montigny, C.; Blier, P. Effect of pindolol on the function of pre- and postsynaptic 5-HT(1A) receptors: In vivo microdialysis and electrophysiological studies in the rat brain. Neuropsychopharmacology 1996, 15, 349–360. [Google Scholar] [CrossRef]
- El Mansari, M.; Sánchez, C.; Chouvet, G.; Renaud, B.; Haddjeri, N. Effects of acute and long-term administration of escitalopram and citalopram on serotonin neurotransmission: An in vivo electrophysiological study in rat brain. Neuropsychopharmacology 2005, 30, 1269–1277. [Google Scholar] [CrossRef]
- Ohno, Y. New Insight into the Therapeutic Role of 5-HT1A Receptors in Central Nervous System Disorders. Cent. Nerv. Syst. Agents Med. Chem. 2010, 10, 148–157. [Google Scholar] [CrossRef]
- Richardson-Jones, J.W.; Craige, C.P.; Guiard, B.P.; Stephen, A.; Metzger, K.L.; Kung, H.F.; Gardier, A.M.; Dranovsky, A.; David, D.J.; Beck, S.G.; et al. 5-HT1A Autoreceptor Levels Determine Vulnerability to Stress and Response to Antidepressants. Neuron 2010, 65, 40–52. [Google Scholar] [CrossRef] [Green Version]
- Whale, R.; Terao, T.; Cowen, P.; Freemantle, N.; Geddes, J. Pindolol augmentation of serotonin reuptake inhibitors for the treatment of depressive disorder: A systematic review. J. Psychopharmacol. 2010, 24, 513–520. [Google Scholar] [CrossRef]
- Arborelius, L.; Linner, L.; Wallsten, C.; Ahlenius, S.; Svensson, T.H. Partial 5-HT(1A) receptor agonist properties of (-)pindolol in combination with citalopram on serotonergic dorsal raphe cell firing in vivo. Psychopharmacology 2000, 151, 77–84. [Google Scholar] [CrossRef]
- Trivedi, M.H.; Fava, M.; Wisniewski, S.R.; Thase, M.E.; Quitkin, F.; Warden, D.; Ritz, L.; Nierenberg, A.A.; Lebowitz, B.D.; Biggs, M.M.; et al. Medication Augmentation after the Failure of SSRIs for Depression. N. Engl. J. Med. 2006, 354, 1243–1252. [Google Scholar] [CrossRef]
- Shelton, R.C.; Tollefson, G.D.; Tohen, M.; Stahl, S.; Gannon, K.S.; Jacobs, T.G.; Buras, W.R.; Bymaster, F.P.; Zhang, W.; Spencer, K.A.; et al. A novel augmentation strategy for treating resistant major depression. Am. J. Psychiatry 2001, 158, 131–134. [Google Scholar] [CrossRef]
- Vieta, E.; Florea, I.; Schmidt, S.N.; Areberg, J.; Ettrup, A. Intravenous vortioxetine to accelerate onset of effect in major depressive disorder: A 2-week, randomized, double-blind, placebo-controlled study. Int. Clin. Psychopharmacol. 2019, 34, 153–160. [Google Scholar] [CrossRef]
- MacQueen, G.M.; Yucel, K.; Taylor, V.H.; Macdonald, K.; Joffe, R. Posterior Hippocampal Volumes Are Associated with Remission Rates in Patients with Major Depressive Disorder. Biol. Psychiatry 2008, 64, 880–883. [Google Scholar] [CrossRef]
- Bremner, J.D.; Narayan, M.; Anderson, E.R.; Staib, L.H.; Miller, H.L.; Charney, D.S. Hippocampal volume reduction in major depression. Am. J. Psychiatry 2000, 157, 115–117. [Google Scholar] [CrossRef] [PubMed]
- Hong, W.; Fan, J.; Yuan, C.; Zhang, C.; Hu, Y.; Peng, D.; Wang, Y.; Huang, J.; Li, Z.; Yu, S.; et al. Significantly decreased mRNA levels of BDNF and MEK1 genes in treatment-resistant depression. Neuroreport 2014, 25, 753–755. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, R.; Ghosh, A.K.; Ghosh, B.; Bhattacharyya, S.; Mondal, A.C. Decreased mRNA and protein expression of BDNF, NGF, and their receptors in the hippocampus from suicide: An analysis in human postmortem brain. Clin. Med. Insights Pathol. 2013, 1–11. [Google Scholar] [CrossRef]
- Filho, C.B.; Jesse, C.R.; Donato, F.; Giacomeli, R.; Del Fabbro, L.; Da Silva Antunes, M.; De Gomes, M.G.; Goes, A.T.R.; Boeira, S.P.; Prigol, M.; et al. Chronic unpredictable mild stress decreases BDNF and NGF levels and Na+,K+-ATPase activity in the hippocampus and prefrontal cortex of mice: Antidepressant effect of chrysin. Neuroscience 2015, 289, 367–380. [Google Scholar] [CrossRef]
- Mao, Q.Q.; Xian, Y.F.; Ip, S.P.; Tsai, S.H.; Che, C.T. Long-term treatment with peony glycosides reverses chronic unpredictable mild stress-induced depressive-like behavior via increasing expression of neurotrophins in rat brain. Behav. Brain Res. 2010, 210, 171–177. [Google Scholar] [CrossRef]
- Su, G.Y.; Yang, J.Y.; Wang, F.; Ma, J.; Zhang, K.; Dong, Y.X.; Song, S.J.; Lu, X.M.; Wu, C.F. Antidepressant-like effects of Xiaochaihutang in a rat model of chronic unpredictable mild stress. J. Ethnopharmacol. 2014, 152, 217–226. [Google Scholar] [CrossRef]
- Antunes, M.S.; Jesse, C.R.; Ruff, J.R.; De Oliveira Espinosa, D.; Gomes, N.S.; Altvater, E.E.T.; Donato, F.; Giacomeli, R.; Boeira, S.P. Hesperidin reverses cognitive and depressive disturbances induced by olfactory bulbectomy in mice by modulating hippocampal neurotrophins and cytokine levels and acetylcholinesterase activity. Eur. J. Pharmacol. 2016, 789, 411–420. [Google Scholar] [CrossRef]
- Noorafshan, A.; Abdollahifar, M.A.; Asadi-Golshan, R.; Rashidian-Rashidabadi, A.; Karbalay-Doust, S. Curcumin and sertraline prevent the reduction of the number of neurons and glial cells and the volume of rats’ medial prefrontal cortex induced by stress. Acta Neurobiol. Exp. 2014, 74, 44–53. [Google Scholar]
- Luscombe, G.P.; Martin, K.F.; Hutchins, L.J.; Gosden, J.; Heal, D.J. Mediation of the antidepressant-like effect of 8-OH-DPAT in mice by postsynaptic 5-HT1A receptors. Br. J. Pharmacol. 1993, 108, 669–677. [Google Scholar] [CrossRef] [Green Version]
- Samuels, B.A.; Anacker, C.; Hu, A.; Levinstein, M.R.; Pickenhagen, A.; Tsetsenis, T.; Madroñal, N.; Donaldson, Z.R.; Drew, L.J.; Dranovsky, A.; et al. 5-HT1A receptors on mature dentate gyrus granule cells are critical for the antidepressant response. Nat. Neurosci. 2015, 18, 1606–1616. [Google Scholar] [CrossRef] [Green Version]
- Richardson-Jones, J.W.; Craige, C.P.; Nguyen, T.H.; Kung, H.F.; Gardier, A.M.; Dranovsky, A.; David, D.J.; Guiard, B.P.; Beck, S.G.; Hen, R.; et al. Serotonin-1A autoreceptors are necessary and sufficient for the normal formation of circuits underlying innate anxiety. J. Neurosci. 2011, 31, 6008–6018. [Google Scholar] [CrossRef] [Green Version]
- Vahid-Ansari, F.; Daigle, M.; Manzini, M.C.; Tanaka, K.F.; Hen, R.; Geddes, S.D.; Béïque, J.-C.; James, J.; Merali, Z.; Albert, P.R. Abrogated Freud-1/Cc2d1a Repression of 5-HT1A Autoreceptors Induces Fluoxetine-Resistant Anxiety/Depression-Like Behavior. J. Neurosci. 2017, 37, 11967–11978. [Google Scholar] [CrossRef] [Green Version]
- Stockmeier, C.A.; Shapiro, L.A.; Dilley, G.E.; Kolli, T.N.; Friedman, L.; Rajkowska, G. Increase in serotonin-1A autoreceptors in the midbrain of suicide victims with major depression—Postmortem evidence for decreased serotonin activity. J. Neurosci. 1998, 18, 7394–7401. [Google Scholar] [CrossRef] [Green Version]
- Parsey, R.V.; Oquendo, M.A.; Ogden, R.T.; Olvet, D.M.; Simpson, N.; Huang, Y.Y.; Van Heertum, R.L.; Arango, V.; Mann, J.J. Altered serotonin 1A binding in major depression: A [carbonyl-C-11] WAY100635 positron emission tomography study. Biol. Psychiatry 2006, 59, 106–113. [Google Scholar] [CrossRef]
- Boldrini, M.; Underwood, M.D.; Mann, J.J.; Arango, V. Serotonin-1A autoreceptor binding in the dorsal raphe nucleus of depressed suicides. J. Psychiatr. Res. 2008, 42, 433–442. [Google Scholar] [CrossRef] [Green Version]
- Lemonde, S.; Turecki, G.; Bakish, D.; Du, L.; Hrdina, P.D.; Bown, C.D.; Sequeira, A.; Kushwaha, N.; Morris, S.J.; Basak, A.; et al. Impaired repression at a 5-hydroxytryptamine 1A receptor gene polymorphism associated with major depression and suicide. J. Neurosci. 2003, 23, 8788–8799. [Google Scholar] [CrossRef] [Green Version]
- Neff, C.D.; Abkevich, V.; Packer, J.C.L.; Chen, Y.; Potter, J.; Riley, R.; Davenport, C.; DeGrado Warren, J.; Jammulapati, S.; Bhathena, A.; et al. Evidence for HTR1A and LHPP as interacting genetic risk factors in major depression. Mol. Psychiatry 2009, 14, 621–630. [Google Scholar] [CrossRef] [Green Version]
- Bhagwagar, Z.; Rabiner, E.A.; Sargent, P.A.; Grasby, P.M.; Cowen, P.J. Persistent reduction in brain serotonin1A receptor binding in recovered depressed men measured by positron emission tomography with [ 11C]WAY-100635. Mol. Psychiatry 2004, 9, 386–392. [Google Scholar] [CrossRef] [Green Version]
- Haddjeri, N.; Blier, P.; De Montigny, C. Long-term antidepressant treatments result in a tonic activation of forebrain 5-HT(1A) receptors. J. Neurosci. 1998, 18, 10150–10156. [Google Scholar] [CrossRef] [Green Version]
- Jacobs, B.; Van Praag, H.; Gage, F. Adult brain neurogenesis and psychiatry a no. Mol. Psychiatry 2000, 5, 262–269. [Google Scholar] [CrossRef]
- Blier, P.; Ward, N.M. Is there a role for 5-HT1A agonists in the treatment of depression? Biol. Psychiatry 2003, 53, 193–203. [Google Scholar] [CrossRef]
- Rabiner, E.A.; Wilkins, M.R.; Turkheimer, F.; Gunn, R.N.; De Haes, J.U.; De Vries, M.; Grasby, P.M. 5-Hydroxytryptamine1A receptor occupancy by novel full antagonist 2-[4-[4-(7-Chloro-2,3-dihydro-1,4-benzdioxyn-5-yl)-1-piperazinyl]butyl] -1,2-benzisothiazol-3-(2H)-one-1,1-dioxide: A [11C][O-methyl-3H]-N-(2-(4-(2-methoxyphenyl)-1-piperazinyl) ethyl)-N-(2-pyridinyl) cyclohexanecarboxamide trihydrochloride (WAY-100635) positron emission tomography study in humans. J. Pharmacol. Exp. Ther. 2002, 301, 1144–1150. [Google Scholar] [CrossRef] [Green Version]
- Scorza, M.; Lladó-Pelfort, L.; Oller, S.; Cortés, R.; Puigdemont, D.; Portella, M.J.; Pérez-Egea, R.; Alvarez, E.; Celada, P.; Pérez, V.; et al. Preclinical and clinical characterization of the selective 5-HT1A receptor antagonist DU-125530 for antidepressant treatment. Br. J. Pharmacol. 2012, 167, 1021–1034. [Google Scholar] [CrossRef] [Green Version]
- Mørk, A.; Pehrson, A.; Brennum, L.T.; Møller Nielsen, S.; Zhong, H.; Lassen, A.B.; Miller, S.; Westrich, L.; Boyle, N.J.; Sánchez, C.; et al. Pharmacological effects of Lu AA21004: A novel multimodal compound for the treatment of major depressive disorder. J. Pharmacol. Exp. Ther. 2012, 340, 666–675. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.-M.; Han, C.; Lee, S.-J.; Patkar, A.A.; Masand, P.S.; Pae, C.-U. Vilazodone for the Treatment of Depression: An Update. Chonnam Med. J. 2016, 52, 91. [Google Scholar] [CrossRef] [Green Version]
- Sowa-Kućma, M.; Pańczyszyn-Trzewik, P.; Misztak, P.; Jaeschke, R.R.; Sendek, K.; Styczeń, K.; Datka, W.; Koperny, M. Vortioxetine: A review of the pharmacology and clinical profile of the novel antidepressant. Pharmacol. Rep. 2017, 69, 595–601. [Google Scholar] [CrossRef]
- Sanchez, C.; Asin, K.E.; Artigas, F. Vortioxetine, a novel antidepressant with multimodal activity: Review of preclinical and clinical data. Pharmacol. Ther. 2015, 145, 43–57. [Google Scholar] [CrossRef]
- Schreiber, R.; De Vry, J. 5-HT1A receptor ligands in animal models of anxiety, impulsivity and depression: Multiple mechanisms of action? Prog. Neuropsychopharmacol. Biol. Psychiatry 1993, 17, 87–104. [Google Scholar] [CrossRef]
- Depoortère, R.; Bardin, L.; Varney, M.A.; Newman-Tancredi, A. Serotonin 5-HT1A Receptor Biased Agonists Display Differential Anxiolytic Activity in a Rat Social Interaction Model. ACS Chem. Neurosci. 2019, 10, 3101–3107. [Google Scholar] [CrossRef]
- Robinson, D.S.; Kajdasz, D.K.; Gallipoli, S.; Whalen, H.; Wamil, A.; Reed, C.R. A 1-year, open-label study assessing the safety and tolerability of vilazodone in patients with major depressive disorder. J. Clin. Psychopharmacol. 2011, 31, 643–646. [Google Scholar] [CrossRef] [PubMed]
- De Boer, S.F.; Koolhaas, J.M. 5-HT1A and 5-HT1B receptor agonists and aggression: A pharmacological challenge of the serotonin deficiency hypothesis. Eur. J. Pharmacol. 2005, 526, 125–139. [Google Scholar] [CrossRef] [PubMed]
- Naranjo, C.A.; Bremner, K.E. Serotonin-altering medications and desire, consumption and effects of alcohol-treatment implications. EXS 1994, 71, 209–219. [Google Scholar] [CrossRef]
- Prinssen, E.P.M.; Colpaert, F.C.; Koek, W. 5-HT1A receptor activation and anti-cataleptic effects: High-efficacy agonists maximally inhibit haloperidol-induced catalepsy. Eur. J. Pharmacol. 2002, 453, 217–221. [Google Scholar] [CrossRef]
- Sari, Y. Serotonin 1B receptors: From protein to physiological function and behavior. Neurosci. Biobehav. Rev. 2004, 28, 565–582. [Google Scholar] [CrossRef]
- Morelli, N.; Gori, S.; Choub, A.; Maluccio, M.R.; Orlandi, G.; Guazzelli, M.; Murri, L. Do 5HT1B/1D Receptor Agonists have an Effect on Mood and Anxiety Disorders? Cephalalgia 2007, 27, 471–472. [Google Scholar] [CrossRef]
- Li, X.; Sun, X.; Sun, J.; Zu, Y.; Zhao, S.; Sun, X.; Li, L.; Zhang, X.; Wang, W.; Liang, Y.; et al. Depressive-like state sensitizes 5-HT1A and 5-HT1B auto-receptors in the dorsal raphe nucleus sub-system. Behav. Brain Res. 2020, 389. [Google Scholar] [CrossRef]
- Carr, G.V.; Lucki, I. The role of serotonin receptor subtypes in treating depression: A review of animal studies. Psychopharmacology 2011, 213, 265–287. [Google Scholar] [CrossRef] [Green Version]
- Banasr, M.; Hery, M.; Printemps, R.; Daszuta, A. Serotonin-induced increases in adult cell proliferation and neurogenesis are mediated through different and common 5-HT receptor subtypes in the dentate gyrus and the subventricular zone. Neuropsychopharmacology 2004, 29, 450–460. [Google Scholar] [CrossRef] [Green Version]
- Nautiyal, K.M.; Tritschler, L.; Ahmari, S.E.; David, D.J.; Gardier, A.M.; Hen, R. A lack of serotonin 1B autoreceptors results in decreased anxiety and depression-related behaviors. Neuropsychopharmacology 2016, 41, 2941–2950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.Y.; Oquendo, M.A.; Harkavy Friedman, J.M.; Greenhill, L.L.; Brodsky, B.; Malone, K.M.; Khait, V.; Mann, J.J. Substance abuse disorder and major depression are associated with the human 5-HTIB receptor gene(HTRIB) G86IC polymorphism. Neuropsychopharmacology 2003, 28, 163–169. [Google Scholar] [CrossRef] [Green Version]
- Ruf, B.; Bhagwagar, Z. The 5-HT1B Receptor: A Novel Target for the Pathophysiology of Depression (Supplementary Tables). Curr. Drug Targets 2009, 10, 1118–1138. [Google Scholar] [CrossRef]
- Nishiguchi, N.; Shirakawa, O.; Ono, H.; Nishimura, A.; Nushida, H.; Ueno, Y.; Maeda, K. No evidence of an association between 5HT1B receptor gene polymorphism and suicide victims in a Japanese population. Am. J. Med. Genet. Neuropsychiatr. Genet. 2001, 105, 343–345. [Google Scholar] [CrossRef] [PubMed]
- Egeland, M.; Warner-Schmidt, J.; Greengard, P.; Svenningsson, P. Co-expression of serotonin 5-HT1B and 5-HT4 receptors in p11 containing cells in cerebral cortex, hippocampus, caudate-putamen and cerebellum. Neuropharmacology 2011, 61, 442–450. [Google Scholar] [CrossRef] [PubMed]
- Svenningsson, P.; Tzavara, E.T.; Qi, H.; Carruthers, R.; Witkin, J.M.; Nomikos, G.G.; Greengard, P. Biochemical and behavioral evidence for antidepressant-like effects of 5-HT6 receptor stimulation. J. Neurosci. 2007, 27, 4201–4209. [Google Scholar] [CrossRef] [Green Version]
- Svenningsson, P. Alterations in 5-HT1B Receptor Function by p11 in Depression-Like States. Science 2006, 311, 77–80. [Google Scholar] [CrossRef] [Green Version]
- Piñeyro, G.; Blier, P. Autoregulation of serotonin neurons: Role in antidepressant drug action. Pharmacol. Rev. 1999, 51, 533–591. [Google Scholar]
- Blier, P.; Chaput, Y.; De Montigny, C. Long-term 5-HT reuptake blockade, but not monoamine oxidase inhibition, decreases the function of terminal 5-HT autoreceptors: An electrophysiological study in the rat brain. Naunyn-Schmiedebergs Arch. Pharmacol. 1988, 337, 246–254. [Google Scholar] [CrossRef]
- Davidson, C.; Stamford, J.A. The effect of paroxetine on 5-HT efflux in the rat dorsal raphe nucleus is potentiated by both 5-HT1A and 5-HT1B D receptor antagonists. Neurosci. Lett. 1995, 188, 41–44. [Google Scholar] [CrossRef]
- Tatarczyńska, E.; Kłodzińska, A.; Stachowicz, K.; Chojnacka-Wójcik, E. Effects of a selective 5-HT1B receptor agonist and antagonists in animal models of anxiety and depression. Behav. Pharmacol. 2004, 15, 523–534. [Google Scholar] [CrossRef]
- Hervás, I.; Queiroz, C.M.T.; Adell, A.; Artigas, F. Role of uptake inhibition and autoreceptor activation in the control of 5-HT release in the frontal cortex and dorsal hippocampus of the rat. Br. J. Pharmacol. 2000, 130, 160–166. [Google Scholar] [CrossRef]
- Knobelman, D.A.; Hen, R.; Lucki, I. Genetic regulation of extracellular serotonin by 5-hydroxytryptamine(1A) and 5-hydroxytryptamine(1B) autoreceptors in different brain regions of the mouse. J. Pharmacol. Exp. Ther. 2001, 298, 1083–1091. [Google Scholar]
- Dawson, L.A.; Hughes, Z.A.; Starr, K.R.; Storey, J.D.; Bettelini, L.; Bacchi, F.; Arban, R.; Poffe, A.; Melotto, S.; Hagan, J.J.; et al. Characterisation of the selective 5-HT1B receptor antagonist SB-616234-A (1-[6-(cis-3,5-dimethylpiperazin-1-yl)-2,3-dihydro-5-methoxyindol-1-yl]- 1-[2′-methyl-4′-(5-methyl-1,2,4-oxadiazol-3-yl)biphenyl-4-yl]methanone hydrochloride): In vivo neurochemical. Neuropharmacology 2006, 50, 975–983. [Google Scholar] [CrossRef]
- Roberts, C.; Watson, J.; Price, G.W.; Middlemiss, D.N. SB-236057-A: A selective 5-HT1B receptor inverse agonist. CNS Drug Rev. 2001, 7, 433–444. [Google Scholar] [CrossRef]
- Chenu, F.; David, D.J.P.; Leroux-Nicollet, I.; Le Maêtre, E.; Gardier, A.M.; Bourin, M. Serotonin1B heteroreceptor activation induces an antidepressant-like effect in mice with an alteration of the serotonergic system. J. Psychiatry Neurosci. 2008, 33, 541–550. [Google Scholar] [PubMed]
- Tatarczyńska, E.; Antkiewicz-Michaluk, L.; Kłodzińska, A.; Stachowicz, K.; Chojnacka-Wójcik, E. Antidepressant-like effect of the selective 5-HT1B receptor agonist CP 94253: A possible mechanism of action. Eur. J. Pharmacol. 2005, 516, 46–50. [Google Scholar] [CrossRef] [PubMed]
- Tiger, M.; Varnäs, K.; Okubo, Y.; Lundberg, J. The 5-HT1B receptor—A potential target for antidepressant treatment. Psychopharmacology 2018, 235, 1317–1334. [Google Scholar] [CrossRef] [Green Version]
- Neumaier, J.F.; Root, D.C.; Hamblin, M.W. Chronic fluoxetine reduces serotonin transporter mRNA and 5-HT(1B) mRNA in a sequential manner in the rat dorsal raphe nucleus. Neuropsychopharmacology 1996, 15, 515–522. [Google Scholar] [CrossRef]
- Gobert, A.; Dekeyne, A.; Millan, M.J. The ability of WAY100,635 to potentiate the neurochemical and functional actions of fluoxetine is enhanced by co-administration of SB224,289, but not BRL15572. Neuropharmacology 2000, 39, 1608–1616. [Google Scholar] [CrossRef]
- Whale, R.; Clifford, E.M.; Bhagwagar, Z.; Cowen, P.J. Decreased sensitivity of 5-HTID receptors in melancholic depression. Br. J. Psychiatry 2001, 178, 454–457. [Google Scholar] [CrossRef] [Green Version]
- Lowther, S.; Katona, C.L.E.; Crompton, M.R.; Horton, R.W. 5-HT(1D) and 5-HT(1E/1F) binding sites in depressed suicides: Increased 5-HT(1D) binding in globus pallidus but not cortex. Mol. Psychiatry 1997, 2, 314–321. [Google Scholar] [CrossRef] [Green Version]
- Klein, M.T.; Dukat, M.; Glennon, R.A.; Teitler, M. Toward Selective Drug Development for the Human 5-Hydroxytryptamine 1E Receptor: A Comparison of 5-Hydroxytryptamine 1E and 1F Receptor Structure-Affinity Relationships. J. Pharmacol. Exp. Ther. 2011, 337, 860–867. [Google Scholar] [CrossRef]
- Burnet, P.W.J.; Eastwood, S.L.; Lacey, K.; Harrison, P.J. The distribution of 5-HT1A and 5-HT2A receptor mRNA in human brain. Brain Res. 1995, 676, 157–168. [Google Scholar] [CrossRef]
- López-Giménez, J.F.; Vilaró, M.T.; Palacios, J.M.; Mengod, G. Mapping of 5-HT2A receptors and their mRNA in monkey brain: [3H]MDL100,907 autoradiography and in situ hybridization studies. J. Comp. Neurol. 2001, 429, 571–589. [Google Scholar] [CrossRef]
- Hannon, J.; Hoyer, D. Molecular biology of 5-HT receptors. Behav. Brain Res. 2008, 195, 198–213. [Google Scholar] [CrossRef]
- Bécamel, C.; Berthoux, C.; Barre, A.; Marin, P. Growing Evidence for Heterogeneous Synaptic Localization of 5-HT2A Receptors. ACS Chem. Neurosci. 2017, 8, 897–899. [Google Scholar] [CrossRef] [Green Version]
- De Almeida, J.; Mengod, G. Quantitative analysis of glutamatergic and GABAergic neurons expressing 5-HT2A receptors in human and monkey prefrontal cortex. J. Neurochem. 2007, 103, 475–486. [Google Scholar] [CrossRef]
- Mestre, T.A.; Zurowski, M.; Fox, S.H. 5-Hydroxytryptamine 2A receptor antagonists as potential treatment for psychiatric disorders. Expert Opin. Investig. Drugs 2013, 22, 411–421. [Google Scholar] [CrossRef]
- Gawliński, D.; Smaga, I.; Zaniewska, M.; Gawlińska, K.; Faron-Górecka, A.; Filip, M. Adaptive mechanisms following antidepressant drugs: Focus on serotonin 5-HT2A receptors. Pharmacol. Rep. 2019, 71, 994–1000. [Google Scholar] [CrossRef] [PubMed]
- Howland, R.H. Vilazodone: Another novel atypical antidepressant drug. J. Psychosoc. Nurs. Ment. Health Serv. 2011, 49, 19–22. [Google Scholar] [CrossRef]
- Richelson, E. Pharmacology of antidepressants. Mayo Clin. Proc. 2001, 76, 511–527. [Google Scholar] [CrossRef] [Green Version]
- Butler, M.O.; Morinobu, S.; Duman, R.S. Chronic Electroconvulsive Seizures Increase the Expression of Serotonin2 Receptor mRNA in Rat Frontal Cortex. J. Neurochem. 1993, 61, 1270–1276. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, A.F.; Machado, J.R.; Cavalcante, J.L. Augmentation strategies for treatment-resistant depression. Curr. Opin. Psychiatry 2009, 22, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, L.L.; Leon, Z.; Yasmin, S.; Price, L.H. Clinical Experience with Mirtazapine in the Treatment of Panic Disorder. Ann. Clin. Psychiatry 1999, 11, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Marek, G.J.; Carpenter, L.L.; McDougle, C.J.; Price, L.H. Synergistic action of 5-HT2A antagonists and selective serotonin reuptake inhibitors in neuropsychiatric disorders. Neuropsychopharmacology 2003, 28, 402–412. [Google Scholar] [CrossRef]
- Amargós-Bosch, M.; Bortolozzi, A.; Puig, M.V.; Serrats, J.; Adell, A.; Celada, P.; Toth, M.; Mengod, G.; Artigas, F. Co-expression and In Vivo Interaction of Serotonin1A and Serotonin2A Receptors in Pyramidal Neurons of Pre-frontal Cortex. Cereb. Cortex 2004, 14, 281–299. [Google Scholar] [CrossRef] [Green Version]
- Gray, J.A.; Roth, B.L. Paradoxical trafficking and regulation of 5-HT2A receptors by agonists and antagonists. Brain Res. Bull. 2001, 56, 441–451. [Google Scholar] [CrossRef] [Green Version]
- Van Oekelen, D.; Luyten, W.H.M.L.; Leysen, J.E. 5-HT2A and 5-HT2C receptors and their atypical regulation properties. Life Sci. 2003, 72, 2429–2449. [Google Scholar] [CrossRef]
- Bressa, G.M.; Marini, S.; Gregori, S. Serotonin S2 receptors blockage and generalized anxiety disorders. A double-blind study on ritanserin and lorazepam. Int. J. Clin. Pharmacol. Res. 1987, 7, 111–119. [Google Scholar]
- Blier, P.; El-Mansari, M. Serotonin and beyond: Therapeutics for major depression. Philos. Trans. R. Soc. B Biol. Sci. 2013, 368. [Google Scholar] [CrossRef]
- Szabo, S.T.; Blier, P. Serotonin 1A receptor ligands act on norepinephrine neuron firing through excitatory amino acid and GABAA receptors: A microiontophoretic study in the rat locus coeruleus. Synapse 2001, 42, 203–212. [Google Scholar] [CrossRef]
- Szabo, S.T.; Blier, P. Response of the norepinephrine system to antidepressant drugs. CNS Spectr. 2001, 6, 679–684. [Google Scholar] [CrossRef] [PubMed]
- Dremencov, E.; El Mansari, M.; Blier, P. Noradrenergic Augmentation of Escitalopram Response by Risperidone: Electrophysiologic Studies in the Rat Brain. Biol. Psychiatry 2007, 61, 671–678. [Google Scholar] [CrossRef] [PubMed]
- Kawahara, Y.; Kawahara, H.; Kaneko, F.; Tanaka, M. Long-term administration of citalopram reduces basal and stress-induced extracellular noradrenaline levels in rat brain. Psychopharmacology 2007, 194, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Seager, M.A.; Barth, V.N.; Phebus, L.A.; Rasmussen, K. Chronic coadministration of olanzapine and fluoxetine activates locus coeruleus neurons in rats: Implications for bipolar disorder. Psychopharmacology 2005, 181, 126–133. [Google Scholar] [CrossRef]
- Chernoloz, O.; El Mansari, M.; Blier, P. Electrophysiological studies in the rat brain on the basis for aripiprazole augmentation of antidepressants in major depressive disorder. Psychopharmacology 2009, 206, 335–344. [Google Scholar] [CrossRef]
- Chernoloz, O.; El Mansari, M.; Blier, P. Effects of sustained administration of quetiapine alone and in combination with a serotonin reuptake inhibitor on norepinephrine and serotonin transmission. Neuropsychopharmacology 2012, 37, 1717–1728. [Google Scholar] [CrossRef] [Green Version]
- Patel, J.G.; Bartoszyk, G.D.; Edwards, E.; Ashby, C.R. The Highly Selective 5-Hydroxytryptamine (5-HT)2A Receptor Antagonist, EMD 281014, Significantly Increases Swimming and Decreases Immobility in Male Congenital Learned Helpless Rats in the Forced Swim Test. Synapse 2004, 52, 73–75. [Google Scholar] [CrossRef]
- Albinsson, A.; Björk, A.; Svartengren, J.; Klint, T.; Andersson, G. Preclinical pharmacology of FG5893: A potential anxiolytic drug with high affinity for both 5-HT1A and 5-HT2A receptors. Eur. J. Pharmacol. 1994, 261, 285–294. [Google Scholar] [CrossRef]
- Marek, G.J.; Martin-Ruiz, R.; Abo, A.; Artigas, F. The selective 5-HT2A receptor antagonist M100907 enhances antidepressant-like behavioral effects of the SSRI fluoxetine. Neuropsychopharmacology 2005, 30, 2205–2215. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Wu, R.; Davis, C.; Li, M. Activation of 5-HT2A receptor disrupts rat maternal behavior. Neuropharmacology 2018, 128, 96–105. [Google Scholar] [CrossRef]
- Berthoux, C.; Barre, A.; Bockaert, J.; Marin, P.; Bécamel, C. Sustained Activation of Postsynaptic 5-HT 2A Receptors Gates Plasticity at Prefrontal Cortex Synapses. Cereb. Cortex 2019, 29, 1659–1669. [Google Scholar] [CrossRef]
- Jaggar, M.; Weisstaub, N.; Gingrich, J.A.; Vaidya, V.A. 5-HT2A receptor deficiency alters the metabolic and transcriptional, but not the behavioral, consequences of chronic unpredictable stress. Neurobiol. Stress 2017, 7, 89–102. [Google Scholar] [CrossRef]
- Duxon, M.S.; Flanigan, T.P.; Reavley, T.A.C.; Baxter, T.G.S.; Blackburn, T.P.; Fone, K.C.F. 5-Hydroxytryptamine-2B receptor protein in the rat central nervous system. Neuroscience 1997, 76, 323–329. [Google Scholar] [CrossRef]
- McCorvy, J.D.; Roth, B.L. Structure and function of serotonin G protein-coupled receptors. Pharmacol. Ther. 2015, 150, 129–142. [Google Scholar] [CrossRef] [Green Version]
- Diaz, S.L.; Doly, S.; Narboux-Nme, N.; Fernández, S.; Mazot, P.; Banas, S.M.; Boutourlinsky, K.; Moutkine, I.; Belmer, A.; Roumier, A.; et al. 5-HT 2B receptors are required for serotonin-selective antidepressant actions. Mol. Psychiatry 2012, 17, 154–163. [Google Scholar] [CrossRef] [Green Version]
- Belmer, A.; Quentin, E.; Diaz, S.L.; Guiard, B.P.; Fernandez, S.P.; Doly, S.; Banas, S.M.; Pitychoutis, P.M.; Moutkine, I.; Muzerelle, A.; et al. Positive regulation of raphe serotonin neurons by serotonin 2B receptors. Neuropsychopharmacology 2018, 43, 1623–1632. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Zhang, S.; Zhang, H.; Nu, W.; Cai, L.; Hertz, L.; Peng, L. Fluoxetine-mediated 5-HT2B receptor stimulation in astrocytes causes EGF receptor transactivation and ERK phosphorylation. Psychopharmacology 2008, 201, 443–458. [Google Scholar] [CrossRef] [PubMed]
- Diaz, S.L.; Narboux-Nême, N.; Boutourlinsky, K.; Doly, S.; Maroteaux, L. Mice lacking the serotonin 5-HT2B receptor as an animal model of resistance to selective serotonin reuptake inhibitors antidepressants. Eur. Neuropsychopharmacol. 2016, 26, 265–279. [Google Scholar] [CrossRef] [Green Version]
- D’Andrea, I.; Béchade, C.; Maroteaux, L. Serotonin and 5-HT2B receptors in microglia control of behavior. Handb. Behav. Neurosci. 2020, 31, 589–599. [Google Scholar] [CrossRef]
- Millan, M.J. Serotonin 5-HT2C receptors as a target for the treatment of depressive and anxious states: Focus on novel therapeutic strategies. Therapie 2005, 60, 441–460. [Google Scholar] [CrossRef]
- López-Giménez, J.F.; Mengod, G.; Palacios, J.M.; Vilaró, M.T. Regional distribution and cellular localization of 5-HT2C receptor mRNA in monkey brain: Comparison with [3H]mesulergine binding sites and choline acetyltransferase mRNA. Synapse 2001, 42, 12–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serrats, J.; Mengod, G.; Cortés, R. Expression of serotonin 5-HT2C receptors in GABAergic cells of the anterior raphe nuclei. J. Chem. Neuroanat. 2005, 29, 83–91. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Bubar, M.J.; Lanfranco, M.F.; Hillman, G.R.; Cunningham, K.A. Serotonin2C receptor localization in GABA neurons of the rat medial prefrontal cortex: Implications for understanding the neurobiology of addiction. Neuroscience 2007, 146, 1677–1688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bubar, M.J.; Stutz, S.J.; Cunningham, K.A. 5-HT2C Receptors localize to dopamine and gaba neurons in the rat mesoaccumbens pathway. PLoS ONE 2011, 6, e20508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dekeyne, A.; Mannoury La Cour, C.; Gobert, A.; Brocco, M.; Lejeune, F.; Serres, F.; Sharp, T.; Daszuta, A.; Soumier, A.; Papp, M.; et al. S32006, a novel 5-HT2C receptor antagonist displaying broad-based antidepressant and anxiolytic properties in rodent models. Psychopharmacology 2008, 199, 549–568. [Google Scholar] [CrossRef] [PubMed]
- Dekeyne, A.; Brocco, M.; Loiseau, F.; Gobert, A.; Rivet, J.M.; Di Cara, B.; Cremers, T.I.; Flik, G.; Fone, K.C.F.; Watson, D.J.G.; et al. S32212, a novel serotonin type 2C receptor inverse agonist/α 2- adrenoceptor antagonist and potential antidepressant: II. A behavioral, neurochemical, and electrophysiological characterization. J. Pharmacol. Exp. Ther. 2012, 340, 765–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cryan, J.F.; Lucki, I. Antidepressant-like behavioral effects mediated by 5-Hydroxytryptamine(2C) receptors. J. Pharmacol. Exp. Ther. 2000, 295, 1120–1126. [Google Scholar]
- Rosenzweig-Lipson, S.; Sabb, A.; Stack, G.; Mitchell, P.; Lucki, I.; Malberg, J.E.; Grauer, S.; Brennan, J.; Cryan, J.F.; Sukoff Rizzo, S.J.; et al. Antidepressant-like effects of the novel, selective, 5-HT2C receptor agonist WAY-163909 in rodents. Psychopharmacology 2007, 192, 159–170. [Google Scholar] [CrossRef] [PubMed]
- Dunlop, J.; Marquis, K.L.; Lim, H.K.; Leung, L.; Kao, J.; Cheesman, C.; Rosenzweig-Lipson, S. Pharmacological profile of the 5-HT2C receptor agonist WAY-163909; therapeutic potential in multiple indications. CNS Drug Rev. 2006, 12, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Esposito, E. Serotonin-Dopamine Interaction as a Focus of Novel Antidepressant Drugs. Curr. Drug Targets 2006, 7, 177–185. [Google Scholar] [CrossRef]
- Jenck, F.; Moreau, J.; Berendsen, H.H.; Boes, M.; Broekkamp, C.L.; Martin, J.R.; Wichmann, J.; Van Delft, A.M. Antiaversive effects of 5HT2C receptor agonists and fluoxetine in a model of panic-like anxiety in rats. Eur. Neuropsychopharmacol. 1998, 8, 161–168. [Google Scholar] [CrossRef]
- Cremers, T.I.F.H.; Giorgetti, M.; Bosker, F.J.; Hogg, S.; Arnt, J.; Mørk, A.; Honig, G.; Bøgesø, K.P.; Westerink, B.H.C.; Den Boer, H.; et al. Inactivation of 5-HT2C receptors potentiates consequences of serotonin reuptake blockade. Neuropsychopharmacology 2004, 29, 1782–1789. [Google Scholar] [CrossRef] [Green Version]
- Cremers, T.I.F.H.; Rea, K.; Bosker, F.J.; Wikström, H.V.; Hogg, S.; Mørk, A.; Westerink, B.H.C. Augmentation of SSRI effects on serotonin by 5-HT2C antagonists: Mechanistic studies. Neuropsychopharmacology 2007, 32, 1550–1557. [Google Scholar] [CrossRef]
- Clenet, F.; De Vos, A.; Bourin, M. Involvement of 5-HT2C receptors in the anti-immobility effects of antidepressants in the forced swimming test in mice. Eur. Neuropsychopharmacol. 2001, 11, 145–152. [Google Scholar] [CrossRef]
- Gurevich, I.; Tamir, H.; Arango, V.; Dwork, A.J.; Mann, J.J.; Schmauss, C. Altered editing of serotonin 2C receptor pre-mRNA in the prefrontal cortex of depressed suicide victims. Neuron 2002, 34, 349–356. [Google Scholar] [CrossRef] [Green Version]
- Yamauchi, M.; Tatebayashi, T.; Nagase, K.; Kojima, M.; Imanishi, T. Chronic treatment with fluvoxamine desensitizes 5-HT2C receptor-mediated hypolocomotion in rats. Pharmacol. Biochem. Behav. 2004, 78, 683–689. [Google Scholar] [CrossRef] [PubMed]
- Invernizzi, R.W.; Pierucci, M.; Calcagno, E.; Di Giovanni, G.; Di Matteo, V.; Benigno, A.; Esposito, E. Selective activation of 5-HT2C receptors stimulates GABA-ergic function in the rat substantia nigra pars reticulata: A combined in vivo electrophysiological and neurochemical study. Neuroscience 2007, 144, 1523–1535. [Google Scholar] [CrossRef]
- Meltzer, H.Y.; Li, Z.; Kaneda, Y.; Ichikawa, J. Serotonin receptors: Their key role in drugs to treat schizophrenia. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2003, 27, 1159–1172. [Google Scholar] [CrossRef] [PubMed]
- Demireva, E.Y.; Suri, D.; Morelli, E.; Mahadevia, D.; Chuhma, N.; Teixeira, C.M.; Ziolkowski, A.; Hersh, M.; Fifer, J.; Bagchi, S.; et al. 5-HT2C receptor blockade reverses SSRI-associated basal ganglia dysfunction and potentiates therapeutic efficacy. Mol. Psychiatry 2020, 25, 3304–3321. [Google Scholar] [CrossRef]
- Angel, I.; Schoemaker, H.; Prouteau, M.; Garreau, M.; Langer, S.Z. Litoxetine: A selective 5-HT uptake inhibitor with concomitant 5-HT3 receptor antagonist and antiemetic properties. Eur. J. Pharmacol. 1993, 232, 139–145. [Google Scholar] [CrossRef]
- Thompson, A.; Lummis, S.R. 5-HT3 Receptors. Curr. Pharm. Des. 2006, 12, 3615–3630. [Google Scholar] [CrossRef]
- Morales, M.; Bloom, F.E. The 5-HT3 receptor is present in different subpopulations of GABAergic neurons in the rat telencephalon. J. Neurosci. 1997, 17, 3157–3167. [Google Scholar] [CrossRef] [Green Version]
- Puig, M.V.; Santana, N.; Celada, P.; Mengod, G.; Artigas, F. In vivo excitation of GABA interneurons in the medial prefrontal cortex through 5-HT3 receptors. Cereb. Cortex 2004, 14, 1365–1375. [Google Scholar] [CrossRef] [Green Version]
- Gupta, D.; Prabhakar, V.; Radhakrishnan, M. 5HT3 receptors: Target for new antidepressant drugs. Neurosci. Biobehav. Rev. 2016, 64, 311–325. [Google Scholar] [CrossRef]
- Ramamoorthy, R.; Radhakrishnan, M.; Borah, M. Antidepressant-like effects of serotonin type-3 antagonist, ondansetron: An investigation in behaviour-based rodent models. Behav. Pharmacol. 2008, 19, 29–40. [Google Scholar] [CrossRef]
- Bravo, G.; Maswood, S. Acute treatment with 5-HT3 receptor antagonist, tropisetron, reduces immobility in intact female rats exposed to the forced swim test. Pharmacol. Biochem. Behav. 2006, 85, 362–368. [Google Scholar] [CrossRef]
- Eisensamer, B.; Rammes, G.; Gimpl, G.; Shapa, M.; Ferrari, U.; Hapfelmeier, G.; Bondy, B.; Parsons, C.; Gilling, K.; Zieglgänsberger, W.; et al. Antidepressants are functional antagonists at the serotonin type 3 (5-HT3) receptor. Mol. Psychiatry 2003, 8, 994–1007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, P. Facilitation of 5-hydroxytryptamine3 receptor desensitization by fluoxetine. Neuroscience 1994, 62, 515–522. [Google Scholar] [CrossRef]
- Mössner, R.; Schmitt, A.; Hennig, T.; Benninghoff, J.; Gerlach, M.; Riederer, P.; Deckert, J.; Lesch, K.P. Quantitation of 5HT3 receptors in forebrain of serotonin transporter deficient mice. J. Neural Transm. 2004, 111, 27–35. [Google Scholar] [CrossRef]
- Redrobe, J.P.; Bourin, M. Partial role of 5-HT2 and 5-HT3 receptors in the activity of antidepressants in the mouse forced swimming test. Eur. J. Pharmacol. 1997, 325, 129–135. [Google Scholar] [CrossRef]
- Alvarez, E.; Perez, V.; Dragheim, M.; Loft, H.; Artigas, F. A double-blind, randomized, placebo-controlled, active reference study of Lu AA21004 in patients with major depressive disorder. Int. J. Neuropsychopharmacol. 2012, 15, 589–600. [Google Scholar] [CrossRef] [Green Version]
- Baldwin, D.S.; Loft, H.; Dragheim, M. A randomised, double-blind, placebo controlled, duloxetine-referenced, fixed-dose study of three dosages of Lu AA21004 in acute treatment of major depressive disorder (MDD). Eur. Neuropsychopharmacol. 2012, 22, 482–491. [Google Scholar] [CrossRef]
- Kelliny, M.; Croarkin, P.E.; Moore, K.M.; Bobo, W.V. Profile of vortioxetine in the treatment of major depressive disorder: An overview of the primary and secondary literature. Ther. Clin. Risk Manag. 2015, 11, 1193–1212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bétry, C.; Pehrson, A.L.; Etiévant, A.; Ebert, B.; Sánchez, C.; Haddjeri, N. The rapid recovery of 5-HT cell firing induced by the antidepressant vortioxetine involves 5-HT3 receptor antagonism. Int. J. Neuropsychopharmacol. 2013, 16, 1115–1127. [Google Scholar] [CrossRef] [Green Version]
- Gupta, D.; Radhakrishnan, M.; Thangaraj, D.; Kurhe, Y. Antidepressant and anti-anxiety like effects of 4i (N-(3-chloro-2- methylphenyl) quinoxalin-2-carboxamide), a novel 5-HT3 receptor antagonist in acute and chronic neurobehavioral rodent models. Eur. J. Pharmacol. 2014, 735, 59–67. [Google Scholar] [CrossRef]
- Gupta, D.; Radhakrishnan, M.; Kurhe, Y. Effect of a novel 5-HT3 receptor antagonist 4i, in corticosterone-induced depression-like behavior and oxidative stress in mice. Steroids 2015, 96, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Gupta, D.; Thangaraj, D.; Radhakrishnan, M. A novel 5HT3 antagonist 4i (N-(3-chloro-2-methylphenyl)quinoxalin-2-carboxamide) prevents diabetes-induced depressive phenotypes in mice: Modulation of serotonergic system. Behav. Brain Res. 2016, 297, 41–50. [Google Scholar] [CrossRef]
- Dhar, A.K.; Mahesh, R.; Jindal, A.; Bhatt, S. Piperazine analogs of naphthyridine-3-carboxamides and indole-2-carboxamides: Novel 5-HT3 receptor antagonists with antidepressant-like activity. Arch. Pharm. 2015, 348, 34–45. [Google Scholar] [CrossRef]
- Kurhe, Y.; Radhakrishnan, M.; Gupta, D.; Devadoss, T. QCM-4 a novel 5-HT3 antagonist attenuates the behavioral and biochemical alterations on chronic unpredictable mild stress model of depression in Swiss albino mice. J. Pharm. Pharmacol. 2014, 66, 122–132. [Google Scholar] [CrossRef] [PubMed]
- Dukat, M.; Alix, K.; Worsham, J.; Khatri, S.; Schulte, M.K. 2-Amino-6-chloro-3,4-dihydroquinazoline: A novel 5-HT3 receptor antagonist with antidepressant character. Bioorg. Med. Chem. Lett. 2013, 23, 5945–5948. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, Y.; Ishima, T.; Takashima, T. The 5-HT3 receptor agonist attenuates the action of antidepressants in the forced swim test in rats. Brain Res. 1998, 786, 189–193. [Google Scholar] [CrossRef]
- Kos, T.; Popik, P.; Pietraszek, M.; Schäfer, D.; Danysz, W.; Dravolina, O.; Blokhina, E.; Galankin, T.; Bespalov, A.Y. Effect of 5-HT3 receptor antagonist MDL 72222 on behaviors induced by ketamine in rats and mice. Eur. Neuropsychopharmacol. 2006, 16, 297–310. [Google Scholar] [CrossRef] [PubMed]
- Gupta, D.; Radhakrishnan, M.; Kurhe, Y. 5HT3 receptor antagonist (ondansetron) reverses depressive behavior evoked by chronic unpredictable stress in mice: Modulation of hypothalamic-pituitary-adrenocortical and brain serotonergic system. Pharmacol. Biochem. Behav. 2014, 124, 129–136. [Google Scholar] [CrossRef]
- Gupta, D.; Radhakrishnan, M.; Kurhe, Y. Ondansetron, a 5HT3 receptor antagonist reverses depression and anxiety-like behavior in streptozotocin-induced diabetic mice: Possible implication of serotonergic system. Eur. J. Pharmacol. 2015, 744, 59–66. [Google Scholar] [CrossRef]
- Kordjazy, N.; Haj-Mirzaian, A.; Amiri, S.; Ostadhadi, S.; Amini-Khoei, H.; Dehpour, A.R. Involvement of N-methyl-d-aspartate receptors in the antidepressant-like effect of 5-hydroxytryptamine 3 antagonists in mouse forced swimming test and tail suspension test. Pharmacol. Biochem. Behav. 2016, 141, 1–9. [Google Scholar] [CrossRef]
- Haj-Mirzaian, A.; Kordjazy, N.; Amiri, S.; Haj-Mirzaian, A.; Amini-Khoei, H.; Ostadhadi, S.; Dehpour, A.R. Involvement of nitric oxide-cyclic guanosine monophosphate pathway in the antidepressant-like effect of tropisetron and ondansetron in mice forced swimming test and tail suspension test. Eur. J. Pharmacol. 2016, 780, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Martin, V.; Riffaud, A.; Marday, T.; Brouillard, C.; Franc, B.; Tassin, J.P.; Sevoz-Couche, C.; Mongeau, R.; Lanfumey, L. Response of Htr3a knockout mice to antidepressant treatment and chronic stress. Br. J. Pharmacol. 2017, 174, 2471–2483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez-Palomar, B.; Mollinedo-Gajate, I.; Berrocoso, E.; Meana, J.J.; Ortega, J.E. Serotonin 5-HT3 receptor antagonism potentiates the antidepressant activity of citalopram. Neuropharmacology 2018, 133, 491–502. [Google Scholar] [CrossRef]
- Ostacher, M.J. Comorbid Alcohol and Substance Abuse Dependence in Depression: Impact on the Outcome of Antidepressant Treatment. Psychiatr. Clin. N. Am. 2007, 30, 69–76. [Google Scholar] [CrossRef]
- Engleman, E.A.; Rodd, Z.A.; Bell, R.L.; Murphy, J.M. The role of 5-HT3 receptors in drug abuse and as a target for pharmacotherapy. CNS Neurol. Disord. Drug Targets 2008, 7, 454–467. [Google Scholar] [CrossRef] [Green Version]
- Rammes, G.; Eisensamer, B.; Ferrari, U.; Shapa, M.; Gimpl, G.; Gilling, K.; Parsons, C.; Riering, K.; Hapfelmeier, G.; Bondy, B.; et al. Antipsychotic drugs antagonize human serotonin type 3 receptor currents in a noncompetitive manner. Mol. Psychiatry 2004, 9, 846–858. [Google Scholar] [CrossRef] [Green Version]
- Kelley, S.P.; Bratt, A.M.; Hodge, C.W. Targeted gene deletion of the 5-HT3A receptor subunit produces an anxiolytic phenotype in mice. Eur. J. Pharmacol. 2003, 461, 19–25. [Google Scholar] [CrossRef]
- Martin, P.; Gozlan, H.; Puech, A.J. 5-HT3 receptor antagonists reverse helpless behaviour in rats. Eur. J. Pharmacol. 1992, 212, 73–78. [Google Scholar] [CrossRef]
- Harmer, C.J.; Reid, C.B.; Ray, M.K.; Goodwin, G.M.; Cowen, P.J. 5HT3 antagonism abolishes the emotion potentiated startle effect in humans. Psychopharmacology 2006, 186, 18–24. [Google Scholar] [CrossRef]
- Varnäs, K.; Halldin, C.; Pike, V.W.; Hall, H. Distribution of 5-HT4 receptors in the postmortem human brain—An autoradiographic study using [125I]SB 207710. Eur. Neuropsychopharmacol. 2003, 13, 228–234. [Google Scholar] [CrossRef]
- Marner, L.; Gillings, N.; Madsen, K.; Erritzoe, D.; Baaré, W.F.C.; Svarer, C.; Hasselbalch, S.G.; Knudsen, G.M. Brain imaging of serotonin 4 receptors in humans with [11C]SB207145-PET. Neuroimage 2010, 50, 855–861. [Google Scholar] [CrossRef]
- Samuels, B.A.; Mendez-David, I.; Faye, C.; David, S.A.; Pierz, K.A.; Gardier, A.M.; Hen, R.; David, D.J. Serotonin 1A and Serotonin 4 Receptors: Essential Mediators of the Neurogenic and Behavioral Actions of Antidepressants. Neuroscientist 2016, 22, 26–45. [Google Scholar] [CrossRef]
- Vidal, R.; Castro, E.; Pilar-Cuellar, F.; Pascual-Brazo, J.; Diaz, A.; Rojo, M.; Linge, R.; Martin, A.; Valdizan, E.; Pazos, A. Serotonin 5-HT4 Receptors: A New Strategy for Developing Fast Acting Antidepressants? Curr. Pharm. Des. 2014, 20, 3751–3762. [Google Scholar] [CrossRef] [Green Version]
- Licht, C.L.; Kirkegaard, L.; Zueger, M.; Chourbaji, S.; Gass, P.; Aznar, S.; Knudsen, G.M. Changes in 5-HT4 receptor and 5-HT transporter binding in olfactory bulbectomized and glucocorticoid receptor heterozygous mice. Neurochem. Int. 2010, 56, 603–610. [Google Scholar] [CrossRef]
- Licht, C.L.; Marcussen, A.B.; Wegener, G.; Overstreet, D.H.; Aznar, S.; Knudsen, G.M. The brain 5-HT4 receptor binding is down-regulated in the Flinders Sensitive Line depression model and in response to paroxetine administration. J. Neurochem. 2009, 109, 1363–1374. [Google Scholar] [CrossRef]
- Vidal, R.; Pilar-Cuellar, F.; Dos Anjos, S.; Linge, R.; Treceno, B.; Ines Vargas, V.; Rodriguez-Gaztelumendi, A.; Mostany, R.; Castro, E.; Diaz, A.; et al. New Strategies in the Development of Antidepressants: Towards the Modulation of Neuroplasticity Pathways. Curr. Pharm. Des. 2011, 17, 521–533. [Google Scholar] [CrossRef] [Green Version]
- Pittenger, C.; Duman, R.S. Stress, depression, and neuroplasticity: A convergence of mechanisms. Neuropsychopharmacology 2008, 33, 88–109. [Google Scholar] [CrossRef]
- Segi-Nishida, E. The effect of serotonin-targeting antidepressants on neurogenesis and neuronal maturation of the hippocampus mediated via 5-HT1A and 5-HT4 receptors. Front. Cell. Neurosci. 2017, 11, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Warner-Schmidt, J.L.; Flajolet, M.; Maller, A.; Chen, E.Y.; Qi, H.; Svenningsson, P.; Greengard, P. Role of p11 in cellular and behavioral effects of 5-HT4 receptor stimulation. J. Neurosci. 2009, 29, 1937–1946. [Google Scholar] [CrossRef] [Green Version]
- Conductier, G.; Dusticier, N.; Lucas, G.; Côté, F.; Debonnel, G.; Daszuta, A.; Dumuis, A.; Nieoullon, A.; Hen, R.; Bockaert, J.; et al. Adaptive changes in serotonin neurons of the raphe nuclei in 5-HT 4 receptor knock-out mouse. Eur. J. Neurosci. 2006, 24, 1053–1062. [Google Scholar] [CrossRef]
- Lucas, G.; Compan, V.; Charnay, Y.; Neve, R.L.; Nestler, E.J.; Bockaert, J.; Barrot, M.; Debonnel, G. Frontocortical 5-HT4 receptors exert positive feedback on serotonergic activity: Viral transfections, subacute and chronic treatments with 5-HT4 agonists. Biol. Psychiatry 2005, 57, 918–925. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, K.; Ikeda, Y.; Haneda, E.; Suzuki, H. Chronic fluoxetine bidirectionally modulates potentiating effects of serotonin on the hippocampal mossy fiber synaptic transmission. J. Neurosci. 2008, 28, 6272–6280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vidal, R.; Valdizan, E.M.; Vilaró, M.T.; Pazos, A.; Castro, E. Reduced signal transduction by 5-HT 4 receptors after long-term venlafaxine treatment in rats. Br. J. Pharmacol. 2010, 161, 695–706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cryan, J.F.; Lucki, I. 5-HT4 receptors do not mediate the antidepressant-like behavioral effects of fluoxetine in a modified forced swim test. Eur. J. Pharmacol. 2000, 409, 295–299. [Google Scholar] [CrossRef]
- Mendez-David, I.; David, D.J.; Darcet, F.; Wu, M.V.; Kerdine-Römer, S.; Gardier, A.M.; Hen, R. Rapid anxiolytic effects of a 5-HT4 receptor agonist are mediated by a neurogenesis-independent mechanism. Neuropsychopharmacology 2014, 39, 1366–1378. [Google Scholar] [CrossRef] [Green Version]
- Amigó, J.; Díaz, A.; Pilar-Cuéllar, F.; Vidal, R.; Martín, A.; Compan, V.; Pazos, A.; Castro, E. The absence of 5-HT4 receptors modulates depression- and anxiety-like responses and influences the response of fluoxetine in olfactory bulbectomised mice: Adaptive changes in hippocampal neuroplasticity markers and 5-HT1A autoreceptor. Neuropharmacology 2016, 111, 47–58. [Google Scholar] [CrossRef] [Green Version]
- Amigo, J.; Garro-Martinez, E.; Vidal Casado, R.; Compan, V.; Pilar-Cuéllar, F.; Pazos, A.; Díaz, A.; Castro, E. 5-HT4 Receptors Are Not Involved in the Effects of Fluoxetine in the Corticosterone Model of Depression. ACS Chem. Neurosci. 2021, 12, 2036–2044. [Google Scholar] [CrossRef]
- Lucas, G.; Rymar, V.V.; Du, J.; Mnie-Filali, O.; Bisgaard, C.; Manta, S.; Lambas-Senas, L.; Wiborg, O.; Haddjeri, N.; Piñeyro, G.; et al. Serotonin4 (5-HT4) Receptor Agonists Are Putative Antidepressants with a Rapid Onset of Action. Neuron 2007, 55, 712–725. [Google Scholar] [CrossRef] [Green Version]
- Darcet, F.; Gardier, A.M.; David, D.J.; Guilloux, J.P. Chronic 5-HT4 receptor agonist treatment restores learning and memory deficits in a neuroendocrine mouse model of anxiety/depression. Neurosci. Lett. 2016, 616, 197–203. [Google Scholar] [CrossRef]
- Duman, R.S. A Silver Bullet for the Treatment of Depression? Neuron 2007, 55, 679–681. [Google Scholar] [CrossRef] [Green Version]
- Murphy, S.E.; De Cates, A.N.; Gillespie, A.L.; Godlewska, B.R.; Scaife, J.C.; Wright, L.C.; Cowen, P.J.; Harmer, C.J. Translating the promise of 5HT4 receptor agonists for the treatment of depression. Psychol. Med. 2020, 51, 1111–1120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Branchek, T.A.; Blackburn, T.P. 5-ht 6 Receptors as Emerging Targets for Drug Discovery. Annu. Rev. Pharmacol. Toxicol. 2000, 40, 319–334. [Google Scholar] [CrossRef] [PubMed]
- Rosse, G.; Schaffhauser, H. 5-HT6 Receptor Antagonists as Potential Therapeutics for Cognitive Impairment. Curr. Top. Med. Chem. 2010, 10, 207–221. [Google Scholar] [CrossRef]
- Heal, D.J.; Smith, S.L.; Fisas, A.; Codony, X.; Buschmann, H. Selective 5-HT6 receptor ligands: Progress in the development of a novel pharmacological approach to the treatment of obesity and related metabolic disorders. Pharmacol. Ther. 2008, 117, 207–231. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, E.S.; Hoplight, B.J.; Lear, S.P.; Neumaier, J.F. BGC20-761, a novel tryptamine analog, enhances memory consolidation and reverses scopolamine-induced memory deficit in social and visuospatial memory tasks through a 5-HT6 receptor-mediated mechanism. Neuropharmacology 2006, 50, 412–420. [Google Scholar] [CrossRef] [PubMed]
- Monsma, F.J.; Shen, Y.; Ward, R.P.; Hamblin, M.W.; Sibley, D.R. Cloning and expression of a novel serotonin receptor with high affinity for tricyclic psychotropic drugs. Mol. Pharmacol. 1993, 43, 320–327. [Google Scholar]
- Wesołowska, A.; Nikiforuk, A. Effects of the brain-penetrant and selective 5-HT6 receptor antagonist SB-399885 in animal models of anxiety and depression. Neuropharmacology 2007, 52, 1274–1283. [Google Scholar] [CrossRef]
- Wesołowska, A.; Nikiforuk, A. The selective 5-HT6 receptor antagonist SB-399885 enhances anti-immobility action of antidepressants in rats. Eur. J. Pharmacol. 2008, 582, 88–93. [Google Scholar] [CrossRef]
- Pereira, M.; Martynhak, B.J.; Andreatini, R.; Svenningsson, P. 5-HT6 receptor agonism facilitates emotional learning. Front. Pharmacol. 2015, 6, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Nikiforuk, A.; Kos, T.; Wesołowska, A. The 5-HT6 receptor agonist EMD 386088 produces antidepressant and anxiolytic effects in rats after intrahippocampal administration. Psychopharmacology 2011, 217, 411–418. [Google Scholar] [CrossRef] [PubMed]
- Jastrzębska-Więsek, M.; Siwek, A.; Partyka, A.; Szewczyk, B.; Sowa-Kućma, M.; Wasik, A.; Kołaczkowski, M.; Wesołowska, A. Antidepressant-like activity of EMD 386088, a 5-HT6 receptor partial agonist, following systemic acute and chronic administration to rats. Naunyn. Schmiedebergs. Arch. Pharmacol. 2015, 388, 1079–1088. [Google Scholar] [CrossRef]
- De Foubert, G.; O’Neill, M.J.; Zetterström, T.S.C. Acute onset by 5-HT6-receptor activation on rat brain brain-derived neurotrophic factor and activity-regulated cytoskeletal-associated protein mRNA expression. Neuroscience 2007, 147, 778–785. [Google Scholar] [CrossRef] [PubMed]
- Dawson, L.A.; Li, P. Effects of 5-HT6 receptor blockade on the neurochemical outcome of antidepressant treatment in the frontal cortex of the rat. J. Neural Transm. 2003, 110, 577–590. [Google Scholar] [CrossRef] [PubMed]
- Hedlund, P.B.; Sutcliffe, J.G. Functional, molecular and pharmacological advances in 5-HT 7 receptor research. Trends Pharmacol. Sci. 2004, 25, 481–486. [Google Scholar] [CrossRef]
- Neumaier, J.F.; Sexton, T.J.; Yracheta, J.; Diaz, A.M.; Brownfield, M. Localization of 5-HT7 receptors in rat brain by immunocytochemistry, in situ hybridization, and agonist stimulated cFos expression. J. Chem. Neuroanat. 2001, 21, 63–73. [Google Scholar] [CrossRef]
- Varnäs, K.; Thomas, D.R.; Tupala, E.; Tiihonen, J.; Hall, H. Distribution of 5-HT7 receptors in the human brain: A preliminary autoradiographic study using [3H]SB-269970. Neurosci. Lett. 2004, 367, 313–316. [Google Scholar] [CrossRef]
- Hedlund, P.B. The 5-HT7 receptor and disorders of the nervous system: An overview. Psychopharmacology 2009, 206, 345–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mullins, U.L.; Gianutsos, G.; Eison, A.S. Effects of antidepressants on 5-HT7 receptor regulation in the rat hypothalamus. Neuropsychopharmacology 1999, 21, 352–367. [Google Scholar] [CrossRef] [Green Version]
- Roth, B.L.; Craigo, S.C.; Choudhary, M.S.; Uluer, A.; Monsma, F.J.J.; Shen, Y.; Meltzer, H.Y.; Sibley, D.R. Binding of typical and atypical antipsychotic agents to 5-hydroxytryptamine-6 and 5-hydroxytryptamine-7 receptors. J. Pharmacol. Exp. Ther. 1994, 268, 1403–1410. [Google Scholar]
- Wesołowska, A.; Nikiforuk, A.; Stachowicz, K.; Tatarczyńska, E. Effect of the selective 5-HT7 receptor antagonist SB 269970 in animal models of anxiety and depression. Neuropharmacology 2006, 51, 578–586. [Google Scholar] [CrossRef]
- Bonaventure, P.; Kelly, L.; Aluisio, L.; Shelton, J.; Lord, B.; Galici, R.; Miller, K.; Atack, J.; Lovenberg, T.W.; Dugovic, C. Selective blockade of 5-hydroxytryptamine (5-HT)7 receptors enhances 5-HT transmission, antidepressant-like behavior, and rapid eye movement sleep suppression induced by citalopram in rodents. J. Pharmacol. Exp. Ther. 2007, 321, 690–698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wesołowska, A.; Tatarczyńska, E.; Nikiforuk, A.; Chojnacka-Wójcik, E. Enhancement of the anti-immobility action of antidepressants by a selective 5-HT7 receptor antagonist in the forced swimming test in mice. Eur. J. Pharmacol. 2007, 555, 43–47. [Google Scholar] [CrossRef]
- Wesołowska, A.; Nikiforuk, A.; Stachowicz, K. Potential anxiolytic and antidepressant effects of the selective 5-HT7 receptor antagonist SB 269970 after intrahippocampal administration to rats. Eur. J. Pharmacol. 2006, 553, 185–190. [Google Scholar] [CrossRef] [PubMed]
- Bonaventure, P.; Dugovic, C.; Kramer, M.; De Boer, P.; Singh, J.; Wilson, S.; Bertelsen, K.; Di, J.; Shelton, J.; Aluisio, L.; et al. Translational evaluation of JNJ-18038683, a 5-hydroxytryptamine type 7 receptor antagonist, on rapid eye movement sleep and in major depressive disorder. J. Pharmacol. Exp. Ther. 2012, 342, 429–440. [Google Scholar] [CrossRef] [Green Version]
- Semenova, S.; Geyer, M.A.; Sutcliffe, J.G.; Markou, A.; Hedlund, P.B. Inactivation of the 5-HT7 Receptor Partially Blocks Phencyclidine-Induced Disruption of Prepulse Inhibition. Biol. Psychiatry 2008, 63, 98–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berman, R.M.; Fava, M.; Thase, M.E.; Trivedi, M.H.; Swanink, R.; McQuade, R.D.; Carson, W.H.; Adson, D.; Taylor, L.; Hazel, J.; et al. Aripiprazole augmentation in major depressive disorder: A double-blind, placebo-controlled study in patients with inadequate response to antidepressants. CNS Spectr. 2009, 14, 197–206. [Google Scholar] [CrossRef] [PubMed]
- Abbas, A.I.; Hedlund, P.B.; Huang, X.P.; Tran, T.B.; Meltzer, H.Y.; Roth, B.L. Amisulpride is a potent 5-HT7 antagonist: Relevance for antidepressant actions in vivo. Psychopharmacology 2009, 205, 119–128. [Google Scholar] [CrossRef] [Green Version]
- Okubo, R.; Hasegawa, T.; Fukuyama, K.; Shiroyama, T.; Okada, M. Current Limitations and Candidate Potential of 5-HT7 Receptor Antagonism in Psychiatric Pharmacotherapy. Front. Psychiatry 2021, 12, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Mørk, A.; Montezinho, L.P.; Miller, S.; Trippodi-Murphy, C.; Plath, N.; Li, Y.; Gulinello, M.; Sanchez, C. Vortioxetine (Lu AA21004), a novel multimodal antidepressant, enhances memory in rats. Pharmacol. Biochem. Behav. 2013, 105, 41–50. [Google Scholar] [CrossRef] [Green Version]
- Okada, M.; Matsumoto, R.; Yamamoto, Y.; Fukuyama, K. Effects of subchronic administrations of vortioxetine, lurasidone, and escitalopram on thalamocortical glutamatergic transmission associated with serotonin 5-ht7 receptor. Int. J. Mol. Sci. 2021, 22, 1351. [Google Scholar] [CrossRef]
- Andressen, K.W.; Manfra, O.; Brevik, C.H.; Ulsund, A.H.; Vanhoenacker, P.; Levy, F.O.; Krobert, K.A. The atypical antipsychotics clozapine and olanzapine promote down-regulation and display functional selectivity at human 5-HT7 receptors. Br. J. Pharmacol. 2015, 172, 3846–3860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stahl, S.M. The serotonin-7 receptor as a novel therapeutic target. J. Clin. Psychiatry 2010, 71, 1414–1415. [Google Scholar] [CrossRef] [PubMed]











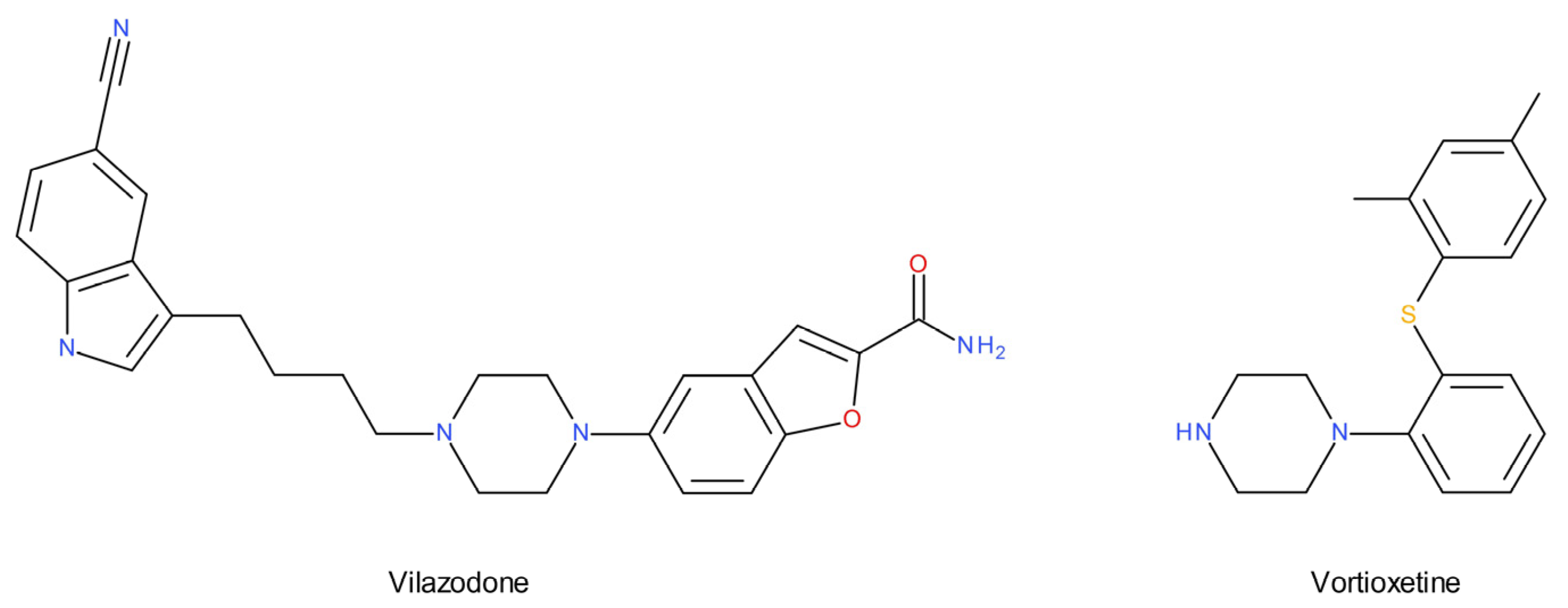{kind=link}
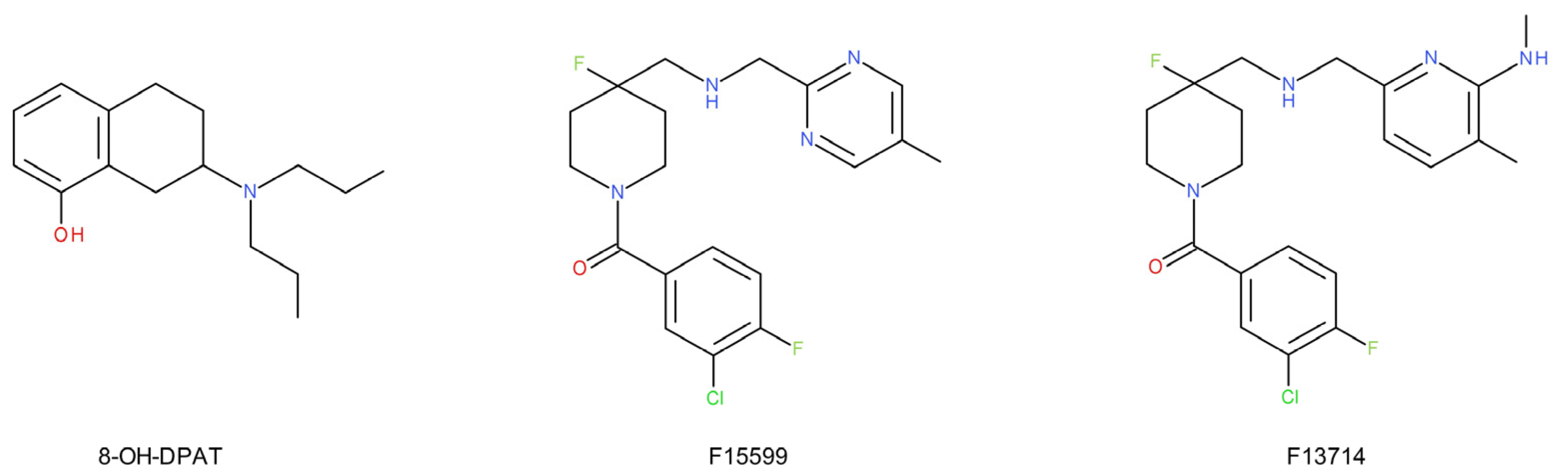{kind=link}
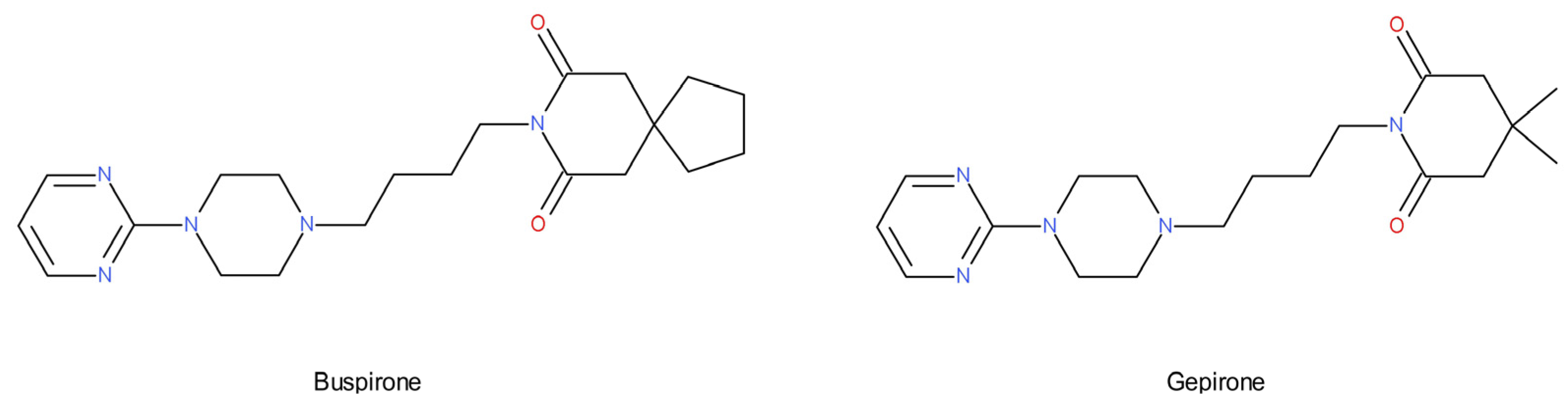{kind=link}
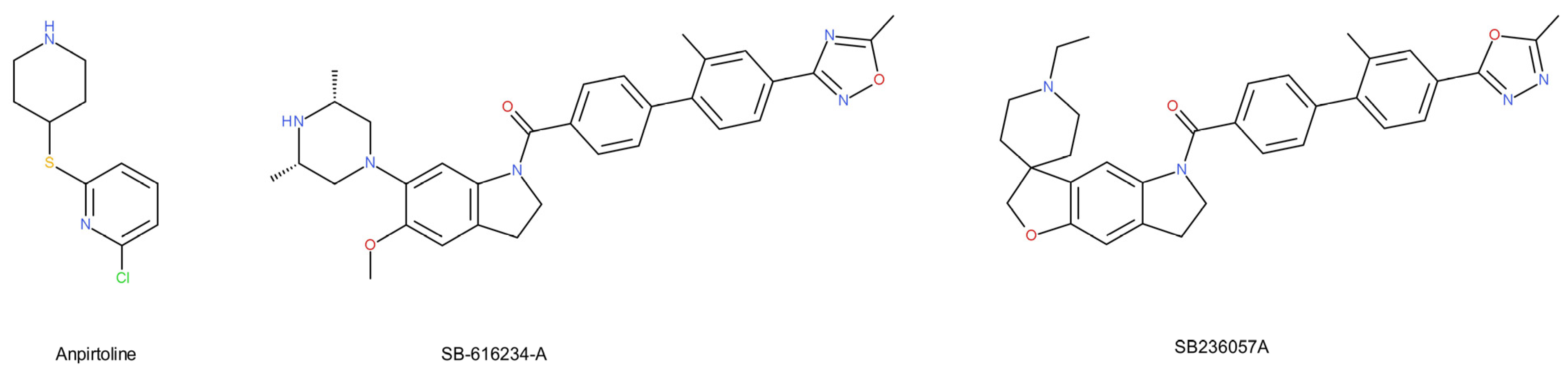{kind=link}
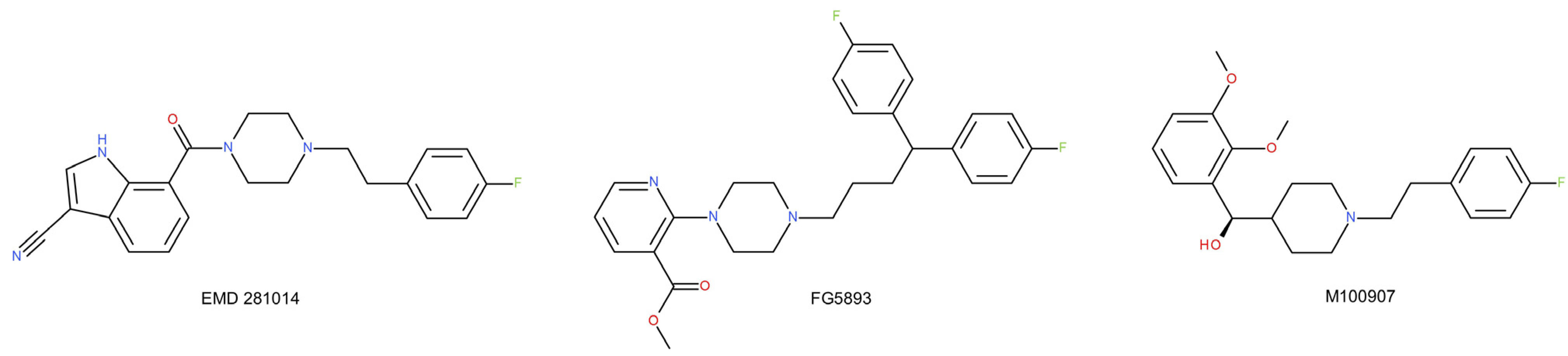{kind=link}
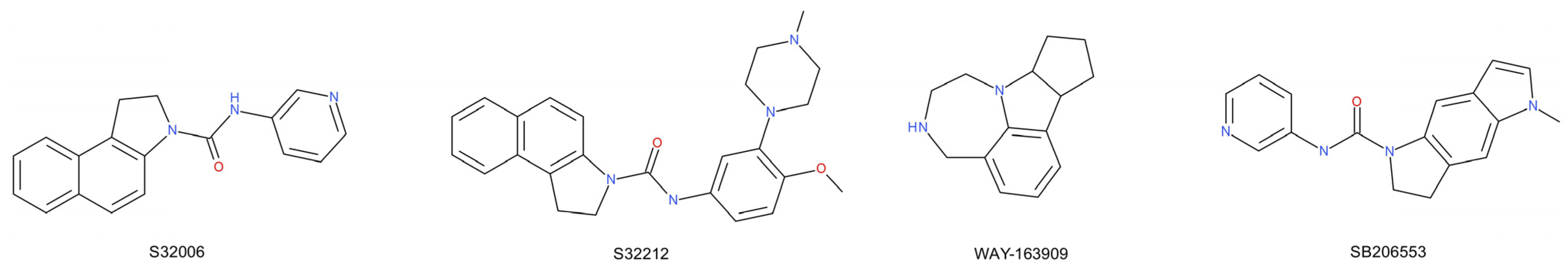{kind=link}
{kind=link}
{kind=link}
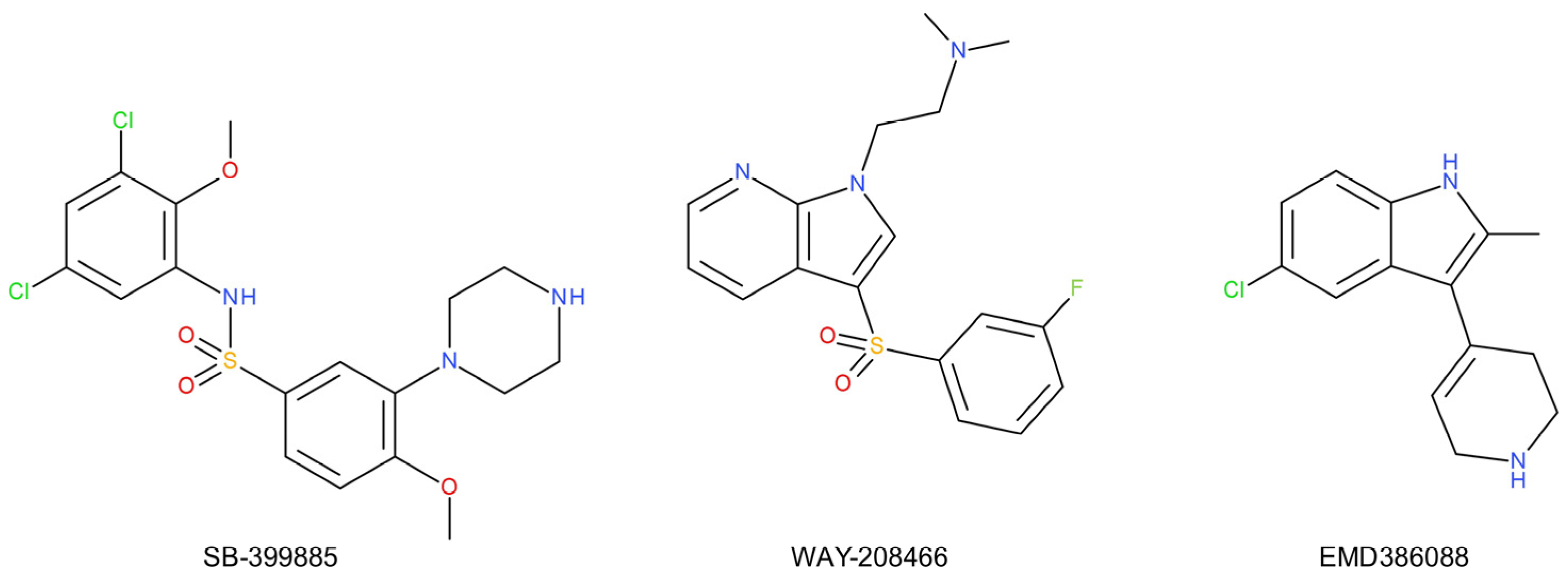{kind=link}
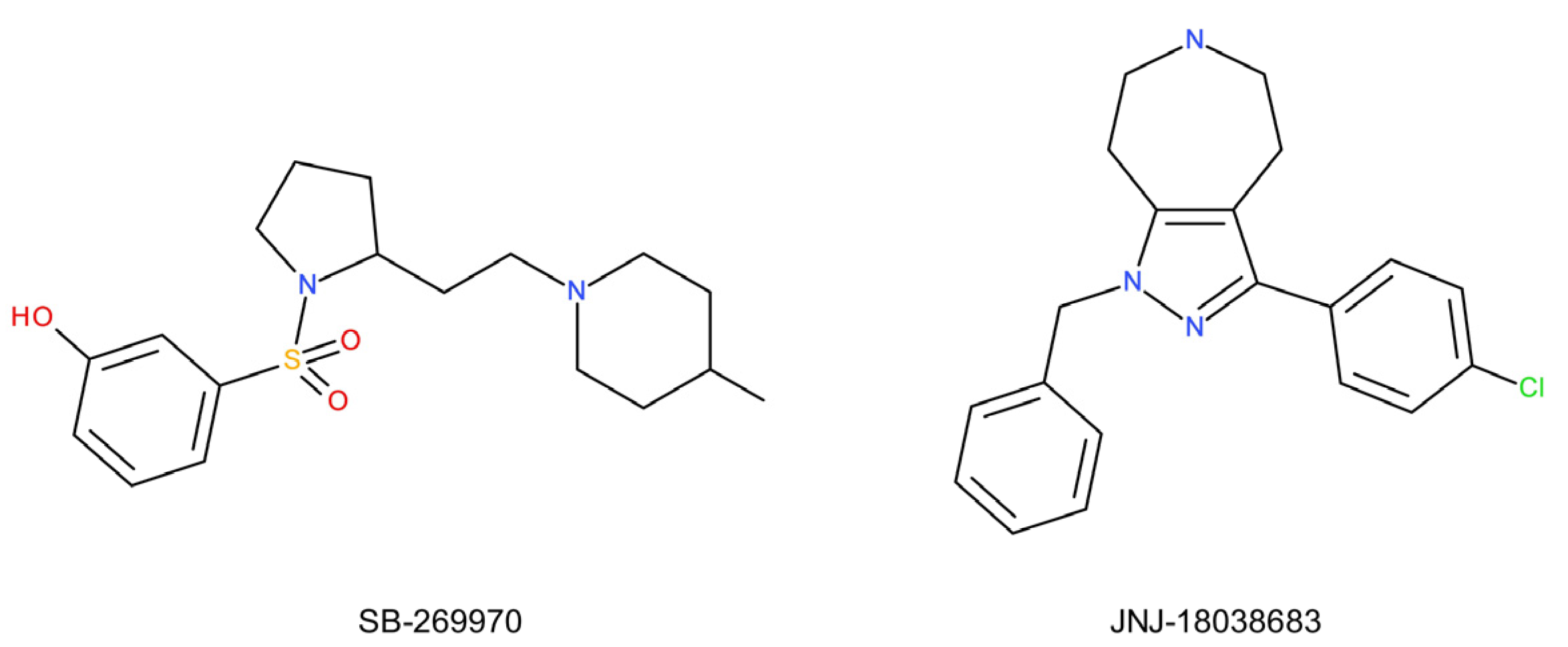{kind=link}
| Clinical Intervention | Mechanism of Action | Effect | References |
|---|---|---|---|
| SSRI + pindolol | SERT inhibition + 5-HT1A agonism | Reduced latency of the antidepressant response and improved the clinical response in previously untreated MDD patients | [21] |
| SSRI + buspirone | SERT inhibition + 5-HT1A partial agonism | Symptom remission in patients unsuccessfully treated with SSRIs | [75] |
| SSRI + mirtazapine | SERT inhibition + 5-HT2A antagonism | Augmentation of the clinical response to SSRIs in treatment-resistant patients | [76] |
| Vilazodone | SERT inhibition + 5-HT1A partial agonism | In contrast to prototypical SSRIs, vilazodone has not been associated with treatment-emergent sexual difficulties or dysfunction | [27] |
| Vortioxetine | SERT, 5-HT3 and 5-HT7 receptors inhibition, 5-HT1A agonism | Potential rapid onset of action | [77] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ślifirski, G.; Król, M.; Turło, J. 5-HT Receptors and the Development of New Antidepressants. Int. J. Mol. Sci. 2021, 22, 9015. https://doi.org/10.3390/ijms22169015
Ślifirski G, Król M, Turło J. 5-HT Receptors and the Development of New Antidepressants. International Journal of Molecular Sciences. 2021; 22(16):9015. https://doi.org/10.3390/ijms22169015
Chicago/Turabian StyleŚlifirski, Grzegorz, Marek Król, and Jadwiga Turło. 2021. "5-HT Receptors and the Development of New Antidepressants" International Journal of Molecular Sciences 22, no. 16: 9015. https://doi.org/10.3390/ijms22169015
APA StyleŚlifirski, G., Król, M., & Turło, J. (2021). 5-HT Receptors and the Development of New Antidepressants. International Journal of Molecular Sciences, 22(16), 9015. https://doi.org/10.3390/ijms22169015