
Reconstruction of the Diaminopimelic Acid Pathway to Promote L-lysine Production in Corynebacterium glutamicum



Abstract
:
1. Introduction
2. Results and Discussion
2.1. The Effects of Different Ammonium (NH4+) Concentrations on L-lysine Production
2.2. Effect of The Upregulated Dehydrogenase Pathway on L-lysine Biosynthesis
2.3. Effect of Gene amtR Deletion on NH4+ Utilization and L-lysine Synthesis
2.4. Blocking the Succinylase Pathway to Upregulate the Dehydrogenase Pathway
2.5. Weakening the dapD Gene Makes the Metabolic Flux of the Two Pathways Reach the Best Balance
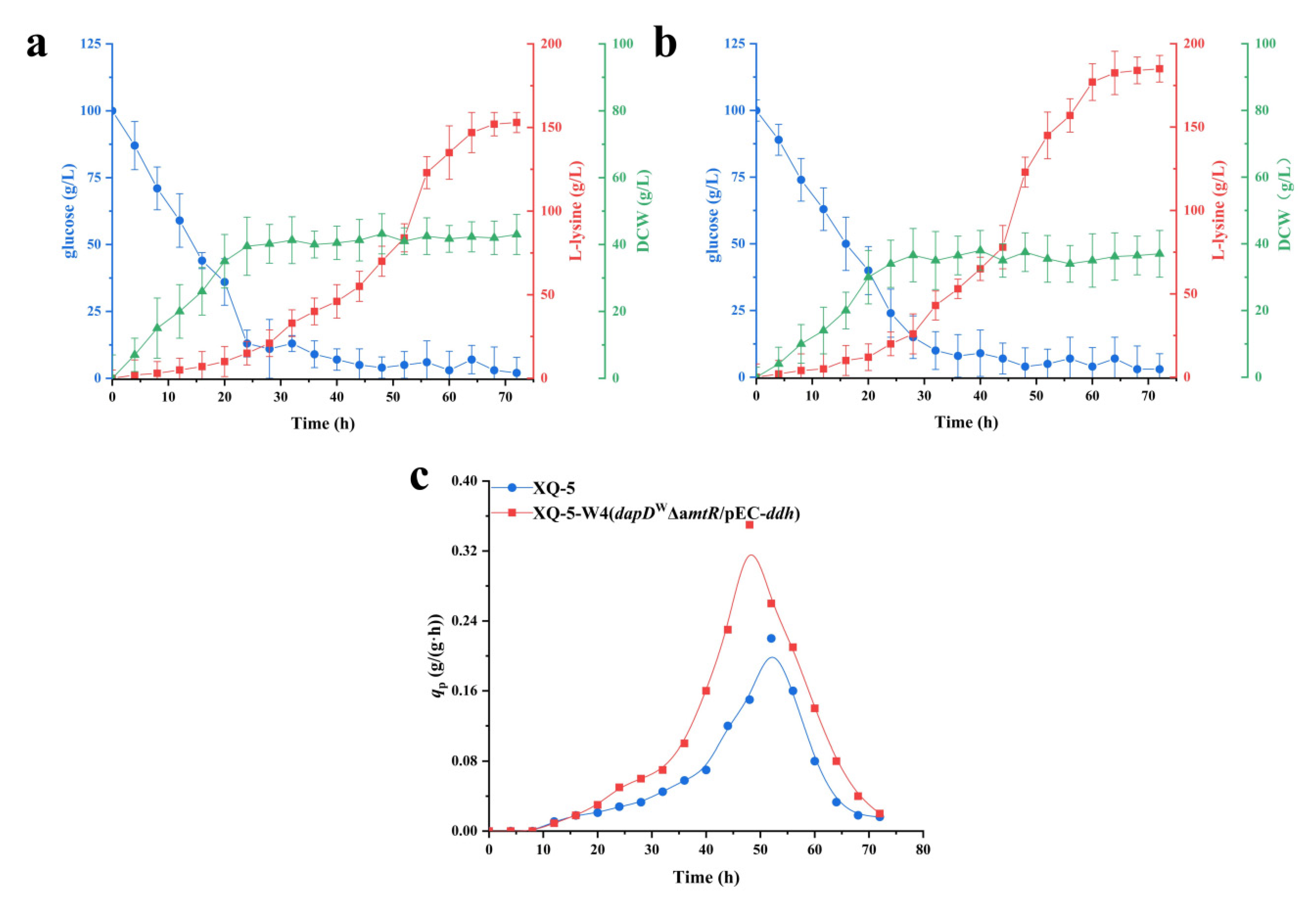
2.6. Fed-Batch Fermentation of C. glutamicum XQ-5-W4
3. Materials and Methods
3.1. Strains, Growth Medium, and Culture Conditions
3.2. Analytical Methods
3.3. Construction of C. glutamicum Recombinant Strains
3.4. Real-Time PCR
3.5. Preparation of Crude Extracts and Enzyme Assays
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Félix, F.; Letti, L.; Vinícius de Melo Pereira, G.; Bonfim, P.; Soccol, V.; Soccol, C. L-lysine production improvement: A review of the state of the art and patent landscape focusing on strain development and fermentation technologies. Crit. Rev. Biotechnol. 2019, 39, 1031–1055. [Google Scholar] [CrossRef]
- Leuchtenberger, W.; Huthmacher, K.; Drauz, K. Biotechnological production of amino acids and derivatives: Current status and prospects. Appl. Microbiol. Biotechnol. 2005, 69, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Nishida, H.; Nishiyama, M.; Kobashi, N.; Kosuge, T.; Hoshino, T.; Yamane, H. A prokaryotic gene cluster involved in synthesis of lysine through the amino adipate pathway: A key to the evolution of amino acid biosynthesis. Genome Res. 1999, 9, 1175–1183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Velasco, A.M.; Leguina, J.I.; Lazcano, A. Molecular Evolution of the Lysine Biosynthetic Pathways. J. Mol. Evol. 2002, 55, 445–449. [Google Scholar] [CrossRef] [PubMed]
- Jetten, M.; Sinskey, A. Recent advances in the physiology and genetics of amino acid-producing bacteria. Crit. Rev. Biotechnol. 1995, 15, 73–103. [Google Scholar] [CrossRef]
- Scapin, G.; Blanchard, J.S. Enzymology of Bacterial Lysine Biosynthesis. Adv. Enzymol. Relat. Areas Mol. Biol. 1998, 72, 279–324. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; White, R.H.; Whitman, W.B. Methanococci Use the Diaminopimelate Aminotransferase (DapL) Pathway for Lysine Biosynthesis. J. Bacteriol. 2010, 192, 3304–3310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Born, T.L.; Blanchard, J.S. Structure/function studies on enzymes in the diaminopimelate pathway of bacterial cell wall biosynthesis. Curr. Opin. Chem. Biol. 1999, 3, 607–613. [Google Scholar] [CrossRef]
- Cirilli, M.; Zheng, R.J.; Scapin, G.; Blanchard, J.S. Structural symmetry: The three-dimensional structure of Haemophilus influenzae diaminopimelate epimerase. Biochemistry 1998, 37, 16452–16458. [Google Scholar] [CrossRef]
- White, P.J. The Essential Role of Diaminopimelate Dehydrogenase in the Biosynthesis of Lysine by Bacillus sphaericus. J. Gen. Microbiol. 1983, 129, 739–749. [Google Scholar] [CrossRef] [Green Version]
- Wenko, L.K.; Treick, R.W.; Wilson, K.G. Isolation and characterization of a gene encoding meso-diaminopimelate dehydrogenase from Glycine max. Plant Mol. Biol. 1985, 4, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Mccoy, A.J.; Adams, N.E.; Hudson, A.O.; Gilvarg, C.; Maurelli, A.T. L,L-diaminopimelate aminotransferase, a trans-kingdom enzyme shared by Chlamydia and plants for synthesis of diaminopimelate/lysine. Proc. Natl. Acad. Sci. USA 2018, 103, 17909–17914. [Google Scholar] [CrossRef] [Green Version]
- Dobson, R.C.J.; Girón, I.; Hudson, A.O. L,L-Diaminopimelate Aminotransferase from Chlamydomonas reinhardtii: A Target for Algaecide Development. PLoS ONE 2011, 6, e20439. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, N.; Cherney, M.M.; Belkum, M.J.V.; Marcus, S.L.; Flegel, M.D.; Clay, M.D.; Deyholos, M.K.; Vederas, J.C.; James, M.N.G. Crystal structure of L,L-diaminopimelate aminotransferase from Arabidopsis thaliana: A recently discovered enzyme in the biosynthesis of L-lysine by plants and Chlamydia. J. Mol. Biol. 2007, 371, 685–702. [Google Scholar] [CrossRef] [PubMed]
- Wehrmann, A.; Phillipp, B.; Sahm, H.; Eggeling, L. Different modes of diaminopimelate synthesis and their role in cell wall integrity: A study with Corynebacterium glutamicum. J. Bacteriol. 1998, 180, 3159–3165. [Google Scholar] [CrossRef] [Green Version]
- Sahm, H.; Eggeling, L.; de Graaf, A. Pathway analysis and metabolic engineering in Corynebacterium glutamicum. Biol. Chem. 2000, 381, 899–910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- UDAKA, S. Screening method for microorganisms accumulating metabolites and its use in the isolation of Micrococcus glutamicus. J. Bacteriol. 1960, 79, 754–755. [Google Scholar] [CrossRef] [Green Version]
- Burillo, S.; Luque, I.; Fuentes, I.; Contreras, A. Interactions between the nitrogen signal transduction protein PII and N-acetyl glutamate kinase in organisms that perform oxygenic photosynthesis. J. Bacteriol. 2004, 186, 3346–3354. [Google Scholar] [CrossRef] [Green Version]
- Bott, M. Offering surprises: TCA cycle regulation in Corynebacterium glutamicum. Trends Microbiol. 2007, 15, 417–425. [Google Scholar] [CrossRef]
- Misono, H.; Togawa, H.; Yamamoto, T.; Soda, K. Occurrence of meso-alpha, epsilon-diaminopimelate dehydrogenase in Bacillus sphaericus. Biochem. Biophys. Res. Commun. 1976, 72, 89–93. [Google Scholar] [CrossRef]
- Rehm, N.; Burkovski, A. Engineering of nitrogen metabolism and its regulation in Corynebacterium glutamicum: Influence on amino acid pools and production. Appl. Microbiol. Biotechnol. 2011, 89, 239–248. [Google Scholar] [CrossRef]
- Sonntag, K.; Eggeling, L.; Graaf, A.A.; Sahm, H. Flux partitioning in the split pathway of lysine synthesis in Corynebacterium glutamicum Quantification by 13C- and 1H-NMR spectroscopy. Eur. J. Biochem. 2010, 213, 1325–1331. [Google Scholar] [CrossRef] [Green Version]
- Xu, M.; Li, J.; Shu, Q.; Tang, M.; Rao, Z. Enhancement of l-arginine production by increasing ammonium uptake in an AmtR-deficient Corynebacterium crenatum mutant. J. Ind. Microbiol. Biotechnol. 2019, 46, 1155–1166. [Google Scholar] [CrossRef] [PubMed]
- Yeh, P.; Sicard, A.; Sinskey, A. General organization of the genes specifically involved in the diaminopimelate-lysine biosynthetic pathway of Corynebacterium glutamicum. Mol. Gen. Genet. 1988, 212, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Palanca, C.; Rubio, V. Structure of AmtR, the global nitrogen regulator of Corynebacterium glutamicum, in free and DNA-bound forms. FEBS J. 2016, 283, 1039–1059. [Google Scholar] [CrossRef] [PubMed]
- Rehm, N.; Georgi, T.; Hiery, E.; Degner, U.; Schmiedl, A.; Burkovski, A.; Bott, M. L-Glutamine as a nitrogen source for Corynebacterium glutamicum: Derepression of the AmtR regulon and implications for nitrogen sensing. Microbiology 2010, 156, 3180–3193. [Google Scholar] [CrossRef] [Green Version]
- Marc, J.; Reinhard, K.; Andreas, B. Nitrogen regulation in Corynebacterium glutamicum: Isolation of genes involved and biochemical characterization of corresponding proteins. FEMS Microbiol. Lett. 1999, 173, 303–310. [Google Scholar] [CrossRef] [Green Version]
- Buchinger, S.; StröSser, J.; Rehm, N.; Hänßler, E.; Hans, S.; Bathe, B.; Schomburg, D.; KräMer, R.; Burkovski, A. A combination of metabolome and transcriptome analyses reveals new targets of the Corynebacterium glutamicum nitrogen regulator AmtR. J. Biotechnol. 2009, 140, 68–74. [Google Scholar] [CrossRef]
- Scapin, G.; Cirilli, M.; Reddy, S.G.; Gao, Y.; Vederas, J.C.; Blanchard, J.S. Substrate and Inhibitor Binding Sites in Corynebacterium glutamicum Diaminopimelate Dehydrogenase. Biochemistry 1998, 37, 3278–3285. [Google Scholar] [CrossRef]
- Dutta, D.; Mishra, S. The structural and energetic aspects of substrate binding and the mechanism of action of the DapE-encoded N-succinyl-L,L-diaminopimelic acid desuccinylase (DapE) investigated using a hybrid QM/MM method. Phys. Chem. Chem. Phys. 2014, 16, 26348–26358. [Google Scholar] [CrossRef]
- Sagong, H.Y.; Kim, K.J. Structural basis for redox sensitivity in Corynebacterium glutamicum diaminopimelate epimerase: An enzyme involved in L-lysine biosynthesis. Sci. Rep. 2017, 7, 42318. [Google Scholar] [CrossRef] [Green Version]
- Sagong, H.; Kim, K. Crystal Structure and Biochemical Characterization of Tetrahydrodipicolinate N-Succinyltransferase from Corynebacterium glutamicum. J. Agric. Food. Chem. 2015, 63, 10641–10646. [Google Scholar] [CrossRef]
- Hartmann, M.; Tauch, A.; Eggeling, L.; Bathe, B.; Möckel, B.; Pühler, A.; Kalinowski, J. Identification and characterization of the last two unknown genes, dapC and dapF, in the succinylase branch of the L-lysine biosynthesis of Corynebacterium glutamicum. J. Biotechnol. 2003, 104, 199–211. [Google Scholar] [CrossRef]
- Shaw-Reid, C.; McCormick, M.; Sinskey, A.; Stephanopoulos, G. Flux through the tetrahydrodipicolinate succinylase pathway is dispensable for L-lysine production in Corynebacterium glutamicum. Appl. Microbiol. Biotechnol. 1999, 51, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Eggeling, L.; Bott, M. A giant market and a powerful metabolism: L-lysine provided by Corynebacterium glutamicum. Appl. Microbiol. Biotechnol. 2015, 99, 3387–3394. [Google Scholar] [CrossRef] [PubMed]
- Misono, H.; Soda, K. Purification and Properties of meso-α, ε-Diaminopimelate D-Dehydrogenase from Bacillus sphaericus. Agric. Biol. Chem. 1980, 44, 227–229. [Google Scholar] [CrossRef] [Green Version]
- He, Z.; Duan, Y.; Zhai, W.; Zhang, X.; Shi, J.; Zhang, X.; Xu, Z. Evaluating Terminator Strength Based on Differentiating Effects on Transcription and Translation. ChemBioChem 2020, 21, 2067–2072. [Google Scholar] [CrossRef]
- Man, Z.; Xu, M.; Rao, Z.; Guo, J.; Yang, T.; Zhang, X.; Xu, Z. Systems pathway engineering of Corynebacterium crenatum for improved L-arginine production. Sci. Rep. 2016, 6, 28629. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.J.; Liu, P.; Nielsen, A.A.K.; Brophy, J.A.N.; Clancy, K.; Peterson, T.; Voigt, C.A. Characterization of 582 natural and synthetic terminators and quantification of their design constraints. Nat. Methods 2013, 10, 659–664. [Google Scholar] [CrossRef]
- Zhang, B.; Yu, M.; Zhou, Y.; Ye, B.C. Improvement of L-ornithine production by attenuation of argF in engineered Corynebacterium glutamicum S9114. Amb Express 2018, 8, 26. [Google Scholar] [CrossRef]
- Wang, L.; Yu, H.; Xu, J.; Ruan, H.; Zhang, W. Deciphering the crucial roles of AraC-type transcriptional regulator Cgl2680 on NADPH metabolism and L-lysine production in Corynebacterium glutamicum. World J. Microbiol. Biotechnol. 2020, 36, 82. [Google Scholar] [CrossRef]
- Xu, J.; Han, M.; Ren, X.; Zhang, W. Modification of aspartokinase III and dihydrodipicolinate synthetase increases the production of L-lysine in Escherichia coli. Biochem. Eng. J. 2016, 114, 79–86. [Google Scholar] [CrossRef]
- Rest, M.E.V.D.; Lange, C.; Molenaar, D. A heat shock following electroporation induces highly efficient transformation of Corynebacterium glutamicum with xenogeneic plasmid DNA. Appl. Microbiol. Biotechnol. 1999, 52, 541–545. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Wu, Z.; Gao, S.; Zhang, W. Rational modification of tricarboxylic acid cycle for improving L-lysine production in Corynebacterium glutamicum. Microb. Cell Factories 2018, 17, 105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.Y.; Zhang, F.; Xu, J.Z.; Zhang, W.G.; Chen, X.L.; Liu, L.M. Improvement of L-Leucine Production in Corynebacterium glutamicum by Altering the Redox Flux. Int. J. Mol. Sci. 2019, 20, 2020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cremer, J.; Treptow, C.; Eggeling, L.; Sahm, H. Regulation of Enzymes of Lysine Biosynthesis in Corynebacterium glutamicum. J. Gen. Microbiol. 1988, 134, 3221–3229. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| XQ-5-1 (XQ-5/pEC-ddh) | XQ-5-3 (XQ-5-ΔamtR/pEC-ddh) | XQ-5-8 (XQ-5-ΔamtRΔdapD/pEC-ddh) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| by-Products | 200 mM | 300 mM | 400 mM | 200 mM | 300 mM | 400 mM | 200 mM | 300 mM | 400 mM |
| glutamate | 0.26 ± 0.05 | 0.35 ± 0.11 | 0.54 ± 0.02 | 0.49 ± 0.06 | 0.97 ± 0.11 | 1.23 ± 0.02 | 1.07 ± 0.04 | 1.75 ± 0.15 | 2.01 ± 0.02 |
| pyruvate | 1.94 ± 0.04 | 2.08 ± 0.14 | 2.36 ± 0.02 | 2.21 ± 0.04 | 2.65 ± 0.03 | 3.32 ± 0.04 | 3.01 ± 0.12 | 3.43 ± 0.09 | 3.91 ± 0.12 |
| isoleucine | 1.06 ± 0.16 | 1.33 ± 0.11 | 1.35 ± 0.12 | 1.57 ± 0.02 | 2.03 ± 0.03 | 2.37 ± 0.02 | 2.31 ± 0.04 | 2.93 ± 0.11 | 3.51 ± 0.02 |
| aspartate | 2.06 ± 0.06 | 2.87 ± 0.05 | 2.92 ± 0.07 | 2.98 ± 0.08 | 3.52 ± 0.12 | 3.91 ± 0.12 | 3.88 ± 0.16 | 4.01 ± 0.11 | 4.22 ± 0.12 |
| methionine | 1.12 ± 0.11 | 1.33 ± 0.07 | 1.41 ± 0.15 | 1.22 ± 0.02 | 1.54 ± 0.11 | 1.87 ± 0.08 | 1.46 ± 0.02 | 1.98 ± 0.05 | 2.31 ± 0.06 |
| threonine | 0.98 ± 0.13 | 1.15 ± 0.14 | 1.23 ± 0.02 | 1.06 ± 0.16 | 2.01 ± 0.11 | 3.91 ± 0.12 | 2.17 ± 0.13 | 3.23 ± 0.11 | 3.91 ± 0.12 |
| Strains | Specific Activity (mU/mg of Protein) | ||
|---|---|---|---|
| DapD | DapDH(F-Reaction) | DapDH (R-Reaction) | |
| XQ-5-W1 | 5.8 ± 0.13 | 201 ± 13.6 | 130 ± 13.8 |
| XQ-5-W2 | 5.3 ± 0.21 | 227 ± 22.1 | 138 ± 18.1 |
| XQ-5-W3 | 4.5 ± 0.39 | 241 ± 26.8 | 145 ± 17.4 |
| XQ-5-W4 | 3.3 ± 0.26 | 260 ± 18.4 | 151 ± 21.2 |
| XQ-5-W5 | 2.1 ± 0.22 | 268 ± 19.3 | 177 ± 19.3 |
| XQ-5-W6 | 1.3 ± 0.42 | 275 ± 20.2 | 194 ± 18.9 |
| Strains | by-Products (g/L) | |||||
|---|---|---|---|---|---|---|
| Glutamate | Pyruvate | Isoleucine | Aspartate | Methionine | Threonine | |
| XQ-5-W1 | 0.4 ± 0.03 | 2.11 ± 0.14 | 1.5 ± 0.09 | 2.93 ± 0.18 | 1.54 ± 0.04 | 1.26 ± 0.08 |
| XQ-5-W2 | 0.54 ± 0.04 | 2.23 ± 0.13 | 1.64 ± 0.07 | 2.85 ± 0.12 | 1.25 ± 0.02 | 1.75 ± 0.11 |
| XQ-5-W3 | 0.64 ± 0.12 | 2.41 ± 0.10 | 1.32 ± 0.05 | 2.42 ± 0.12 | 1.05 ± 0.06 | 1.35 ± 0.02 |
| XQ-5-W4 | 0.78 ± 0.04 | 2.32 ± 0.13 | 1.24 ± 0.09 | 2.15 ± 0.11 | 0.85 ± 0.12 | 1.65 ± 0.08 |
| XQ-5-W5 | 1.32 ± 0.07 | 2.87 ± 0.11 | 2.01 ± 0.12 | 2.78 ± 0.05 | 1.05 ± 0.10 | 2.75 ± 0.02 |
| XQ-5-W6 | 1.64 ± 0.12 | 3.12 ± 0.03 | 2.64 ± 0.14 | 3.65 ± 0.02 | 1.75 ± 0.04 | 3.01 ± 0.12 |
| Strains and Plasmids | Characters | Reference |
|---|---|---|
| C. glutamicum strains | ||
| XQ-5 | C. glutamicum AECr 2-TAr MFr Mets, L-lysine-producing bacteria derived from strain C. glutamicum ATCC13032 | [41] |
| XQ-5-1 | strain XQ-5 harboring plasmid pEC-XK99E-ddh | this study |
| XQ-5-2 | derivative of strain XQ-5 with deletion of amtR | this study |
| XQ-5-3 | strain XQ-5-2 harboring plasmid pEC-XK99E-ddh | this study |
| XQ-5-4 | derivative of strain XQ-5 with deletion of dapD | this study |
| XQ-5-5 | derivative of strain XQ-5 with deletion of ddh | this study |
| XQ-5-6 | derivative of strain XQ-5-2 with deletion of dapD | this study |
| XQ-5-7 | strain XQ-5-4 harboring plasmid pEC-XK99E-ddh | this study |
| XQ-5-8 | strain XQ-5-6 harboring plasmid pEC-XK99E-ddh | this study |
| XQ-5-W1 | derivative of strain XQ-5-3 with weaking of dapD(T1-Terminator) | this study |
| XQ-5-W2 | derivative of strain XQ-5-3 with weaking of dapD(T2-Terminator) | this study |
| XQ-5-W3 | derivative of strain XQ-5-3 with weaking of dapD(T3-Terminator) | this study |
| XQ-5-W4 | derivative of strain XQ-5-3 with weaking of dapD(T4-Terminator) | this study |
| XQ-5-W5 | derivative of strain XQ-5-3 with weaking of dapD(T5-Terminator) | this study |
| XQ-5-W6 | derivative of strain XQ-5-3 with weaking of dapD(T6-Terminator) | this study |
| plasmid | ||
| pEC-XK99E | Kanr, Expression vector with pMB1 replicon | stratagene |
| pK18mobSacB | Kanr, Integration vector | stratagene |
| pEC-XK99E-ddh | pEC-XK99E carrying gene ddh from C.glutamicum | this study |
| pK18mobsacB-ΔamtR | pK18mobsacB carrying amtR-L and amtR-R fragments | this study |
| pK18mobsacB-ΔdapD pK18mobsacB-Δddh | pK18mobsacB carrying dapD-L and dapD-R fragments | this study |
| pK18mobsacB carrying ddh-L and ddh-R fragments | this study | |
| pK18mobsacB-T1 | a derivative of pK18mobsacB, harboring the fragment of inserting T1 terminator in front of dapD | this study |
| pK18mobsacB-T2 | a derivative of pK18mobsacB, harboring the fragment of inserting T2 terminator in front of dapD | this study |
| pK18mobsacB-T3 | a derivative of pK18mobsacB, harboring the fragment of inserting T3 terminator in front of dapD | this study |
| pK18mobsacB-T4 | a derivative of pK18mobsacB, harboring the fragment of inserting T4 terminator in front of dapD | this study |
| pK18mobsacB-T5 | a derivative of pK18mobsacB, harboring the fragment of inserting T5 terminator in front of dapD | this study |
| pK18mobsacB-T6 | a derivative of pK18mobsacB, harboring the fragment of inserting T6 terminator in front of dapD | this study |
| XQ-5-W4 | derivative of strain XQ-5-3 with weaking of dapD(T4-Terminator) | this study |
| XQ-5-W5 | derivative of strain XQ-5-3 with weaking of dapD(T5-Terminator) | this study |
| XQ-5-W6 | derivative of strain XQ-5-3 with weaking of dapD(T6-Terminator) | this study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, N.; Zhang, T.-T.; Rao, Z.-M.; Zhang, W.-G.; Xu, J.-Z. Reconstruction of the Diaminopimelic Acid Pathway to Promote L-lysine Production in Corynebacterium glutamicum. Int. J. Mol. Sci. 2021, 22, 9065. https://doi.org/10.3390/ijms22169065
Liu N, Zhang T-T, Rao Z-M, Zhang W-G, Xu J-Z. Reconstruction of the Diaminopimelic Acid Pathway to Promote L-lysine Production in Corynebacterium glutamicum. International Journal of Molecular Sciences. 2021; 22(16):9065. https://doi.org/10.3390/ijms22169065
Chicago/Turabian StyleLiu, Ning, Ting-Ting Zhang, Zhi-Ming Rao, Wei-Guo Zhang, and Jian-Zhong Xu. 2021. "Reconstruction of the Diaminopimelic Acid Pathway to Promote L-lysine Production in Corynebacterium glutamicum" International Journal of Molecular Sciences 22, no. 16: 9065. https://doi.org/10.3390/ijms22169065
APA StyleLiu, N., Zhang, T. -T., Rao, Z. -M., Zhang, W. -G., & Xu, J. -Z. (2021). Reconstruction of the Diaminopimelic Acid Pathway to Promote L-lysine Production in Corynebacterium glutamicum. International Journal of Molecular Sciences, 22(16), 9065. https://doi.org/10.3390/ijms22169065