
Spaceflight Modulates the Expression of Key Oxidative Stress and Cell Cycle Related Genes in Heart


Abstract
:1. Introduction
2. Results
2.1. Body Mass of Animals Pre- and Post-Flight
2.2. Spaceflight Altered Expression of Oxidative Stress Related Genes
2.3. Spaceflight Altered Expression of Cell Cycle/Proliferation Related Genes
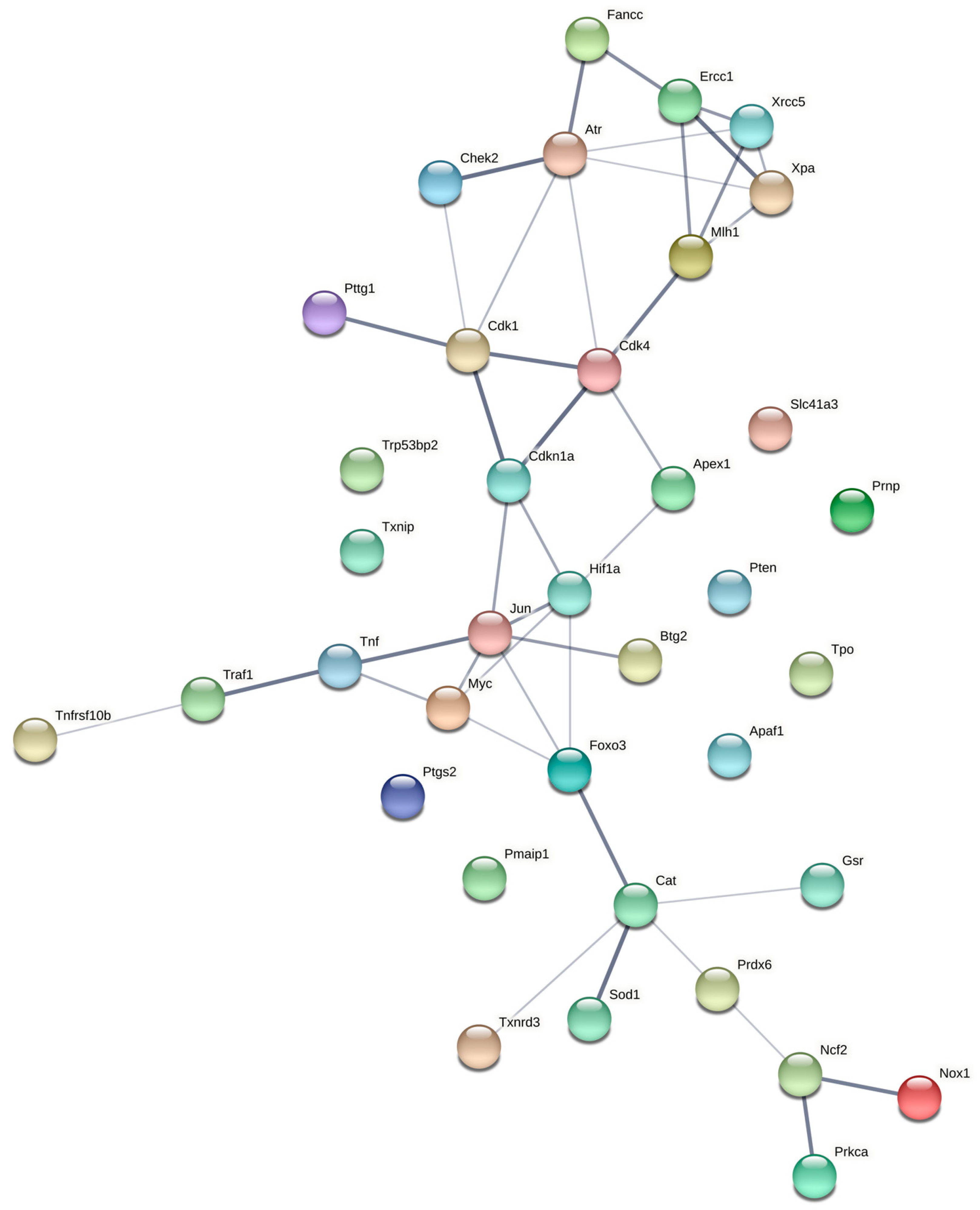
2.4. Gene Networks and Disease Predictions
3. Discussion
3.1. Oxidative Stress-Related Gene Expression in Spaceflight Heart
3.2. Cell Cycle and Senescence-Related Gene Expression in Spaceflight Heart
3.3. Apoptosis and DNA Damage Repair-Related Gene Expression in Spaceflight Hearts
3.4. Inflammatory Pathway Gene Expression in Spaceflight Heart
3.5. Gene Networks and Molecular Signatures of Disease
3.6. Limitations
4. Materials and Methods
4.1. Ethics Statement
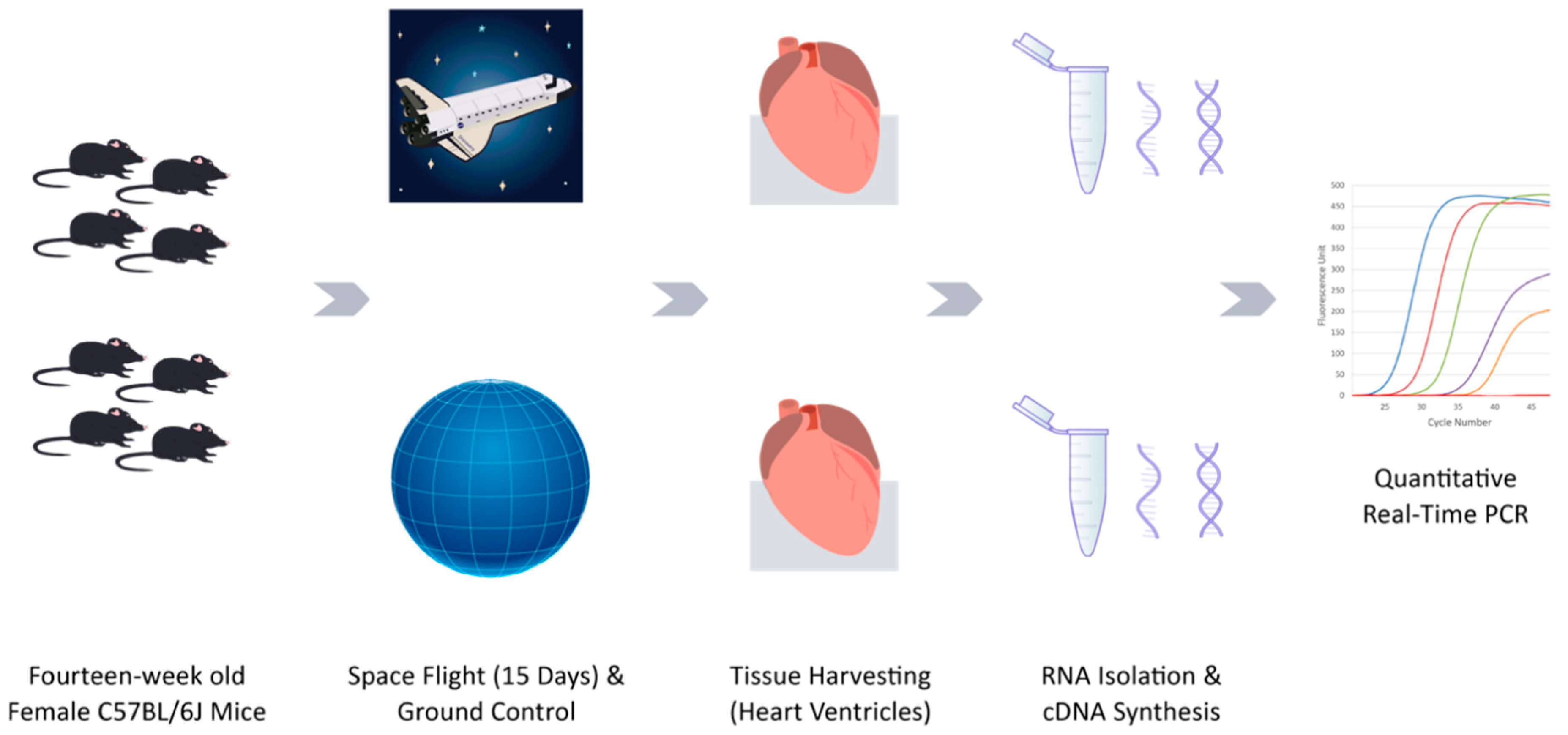
4.2. Animals and Spaceflight
4.3. RNA Isolation and cDNA Synthesis
4.4. Quantitative Real-Time PCR Array
4.5. Nfe2l2 Real-Time PCR
4.6. Data Analysis and Statistics
4.7. Gene Network Predictions and Functional Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hughson, R.L.; Shoemaker, J.K.; Blaber, A.P.; Arbeille, P.; Greaves, D.K.; Pereira-Junior, P.P.; Xu, D. Cardiovascular regulation during long-duration spaceflights to the International Space Station. J. Appl. Physiol. 2012, 112, 719–727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demontis, G.C.; Germani, M.M.; Caiani, E.G.; Barravecchia, I.; Passino, C.; Angeloni, D. Human Pathophysiological Adaptations to the Space Environment. Front. Physiol. 2017, 8, 547. [Google Scholar] [CrossRef] [PubMed]
- Marshall-Goebel, K.; Laurie, S.S.; Alferova, I.V.; Arbeille, P.; Aunon-Chancellor, S.M.; Ebert, D.J.; Lee, S.M.C.; Macias, B.R.; Martin, D.S.; Pattarini, J.M.; et al. Assessment of jugular venous blood flow stasis and thrombosis during spaceflight. JAMA Netw. Open 2019, 2, e1915011. [Google Scholar] [CrossRef] [PubMed]
- Buckey, J.C., Jr.; Gaffney, F.A.; Lane, L.D.; Levine, B.D.; Watenpaugh, D.E.; Wright, S.J.; Yancy, C.W., Jr.; Meyer, D.M.; Blomqvist, C.G. Central venous pressure in space. J. Appl. Physiol. 1996, 81, 19–25. [Google Scholar] [CrossRef]
- Moore, T.P.; Thornton, W.E. Space shuttle inflight and postflight fluid shifts measured by leg volume changes. Aviat. Space Environ. Med. 1987, 58, A91–A96. [Google Scholar]
- Vernice, N.A.; Meydan, C.; Afshinnekoo, E.; Mason, C.E. Long-term spaceflight and the cardiovascular system. Precis. Clin. Med. 2020, 3, 284–291. [Google Scholar] [CrossRef]
- Wieling, W.; Halliwill, J.R.; Karemaker, J.M. Orthostatic intolerance after space flight. J. Physiol. 2002, 538, 1. [Google Scholar] [CrossRef] [Green Version]
- Gallo, C.; Ridolfi, L.; Scarsoglio, S. Cardiovascular deconditioning during long-term spaceflight through multiscale modeling. NPJ Microgravity 2020, 6, 27. [Google Scholar] [CrossRef]
- MacNamara, J.P.; Dias, K.A.; Sarma, S.; Lee, S.M.C.; Martin, D.; Romeijn, M.; Zaha, V.G.; Levine, B.D. Cardiac effects of repeated weightlessness during extreme duration swimming compared with spaceflight. Circulation 2021, 143, 1533–1535. [Google Scholar] [CrossRef]
- Perhonen, M.A.; Franco, F.; Lane, L.D.; Buckey, J.C.; Blomqvist, C.G.; Zerwekh, J.E.; Peshock, R.M.; Weatherall, P.T.; Levine, B.D. Cardiac atrophy after bed rest and spaceflight. J. Appl. Physiol. 2001, 91, 645–653. [Google Scholar] [CrossRef]
- Summers, R.L.; Martin, D.S.; Meck, J.V.; Coleman, T.G. Mechanism of spaceflight-induced changes in left ventricular mass. Am. J. Cardiol. 2005, 95, 1128–1130. [Google Scholar] [CrossRef]
- Goldstein, M.A.; Edwards, R.J.; Schroeter, J.P. Cardiac morphology after conditions of microgravity during COSMOS 2044. J. Appl. Physiol. 1992, 73, 94S–100S. [Google Scholar] [CrossRef] [PubMed]
- Ray, C.A.; Vasques, M.; Miller, T.A.; Wilkerson, M.K.; Delp, M.D. Effect of short-term microgravity and long-term hindlimb unloading on rat cardiac mass and function. J. Appl. Physiol. 2001, 91, 1207–1213. [Google Scholar] [CrossRef]
- Liang, L.; Li, H.; Cao, T.; Qu, L.; Zhang, L.; Fan, G.C.; Greer, P.A.; Li, J.; Jones, D.L.; Peng, T. Calpain activation mediates microgravity-induced myocardial abnormalities in mice via p38 and ERK1/2 MAPK pathways. J. Biol. Chem. 2020, 295, 16840–16851. [Google Scholar] [CrossRef]
- Wang, X.P.; Xing, C.Y.; Zhang, J.X.; Zhou, J.H.; Li, Y.C.; Yang, H.Y.; Zhang, P.F.; Zhang, W.; Huang, Y.; Long, J.G.; et al. Time-restricted feeding alleviates cardiac dysfunction induced by simulated microgravity via restoring cardiac FGF21 signaling. FASEB J. 2020, 34, 15180–15196. [Google Scholar] [CrossRef] [PubMed]
- Coleman, M.A.; Sasi, S.P.; Onufrak, J.; Natarajan, M.; Manickam, K.; Schwab, J.; Muralidharan, S.; Peterson, L.E.; Alekseyev, Y.O.; Yan, X.; et al. Low-dose radiation affects cardiac physiology: Gene networks and molecular signaling in cardiomyocytes. Am. J. Physiol. Heart Circ. Physiol. 2015, 309, H1947–H1963. [Google Scholar] [CrossRef] [PubMed]
- Seawright, J.W.; Samman, Y.; Sridharan, V.; Mao, X.W.; Cao, M.; Singh, P.; Melnyk, S.; Koturbash, I.; Nelson, G.A.; Hauer-Jensen, M.; et al. Effects of low-dose rate gamma-irradiation combined with simulated microgravity on markers of oxidative stress, DNA methylation potential, and remodeling in the mouse heart. PLoS ONE 2017, 12, e0180594. [Google Scholar] [CrossRef] [PubMed]
- Beheshti, A.; McDonald, J.T.; Miller, J.; Grabham, P.; Costes, S.V. GeneLab database analyses suggest long-term impact of space radiation on the cardiovascular system by the activation of FYN through reactive oxygen species. Int. J. Mol. Sci. 2019, 20, 661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baqai, F.P.; Gridley, D.S.; Slater, J.M.; Luo-Owen, X.; Stodieck, L.S.; Ferguson, V.; Chapes, S.K.; Pecaut, M.J. Effects of spaceflight on innate immune function and antioxidant gene expression. J. Appl. Physiol. 2009, 106, 1935–1942. [Google Scholar] [CrossRef] [Green Version]
- Afanas’ev, I. ROS and RNS signaling in heart disorders: Could antioxidant treatment be successful? Oxid. Med. Cell Longev. 2011, 2011, 293769. [Google Scholar] [CrossRef] [Green Version]
- Rizzo, A.M.; Corsetto, P.A.; Montorfano, G.; Milani, S.; Zava, S.; Tavella, S.; Cancedda, R.; Berra, B. Effects of long-term space flight on erythrocytes and oxidative stress of rodents. PLoS ONE 2012, 7, e32361. [Google Scholar] [CrossRef] [Green Version]
- Barrila, J.; Ott, C.M.; LeBlanc, C.; Mehta, S.K.; Crabbe, A.; Stafford, P.; Pierson, D.L.; Nickerson, C.A. Spaceflight modulates gene expression in the whole blood of astronauts. NPJ Microgravity 2016, 2, 16039. [Google Scholar] [CrossRef] [Green Version]
- Tauber, S.; Christoffel, S.; Thiel, C.S.; Ullrich, O. Transcriptional homeostasis of oxidative stress-related pathways in altered gravity. Int. J. Mol. Sci. 2018, 19, 2814. [Google Scholar] [CrossRef] [Green Version]
- Garrett-Bakelman, F.E.; Darshi, M.; Green, S.J.; Gur, R.C.; Lin, L.; Macias, B.R.; McKenna, M.J.; Meydan, C.; Mishra, T.; Nasrini, J.; et al. The NASA Twins Study: A multidimensional analysis of a year-long human spaceflight. Science 2019, 364. [Google Scholar] [CrossRef]
- Overbey, E.G.; da Silveira, W.A.; Stanbouly, S.; Nishiyama, N.C.; Roque-Torres, G.D.; Pecaut, M.J.; Zawieja, D.C.; Wang, C.; Willey, J.S.; Delp, M.D.; et al. Spaceflight influences gene expression, photoreceptor integrity, and oxidative stress-related damage in the murine retina. Sci. Rep. 2019, 9, 13304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- da Silveira, W.A.; Fazelinia, H.; Rosenthal, S.B.; Laiakis, E.C.; Kim, M.S.; Meydan, C.; Kidane, Y.; Rathi, K.S.; Smith, S.M.; Stear, B.; et al. Comprehensive multi-omics analysis reveals mitochondrial stress as a central biological hub for spaceflight impact. Cell 2020, 183, 1185–1201.e1120. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, N.; Waning, D.L.; Gautam, A.; Hoke, A.; Sowe, B.; Youssef, D.; Butler, S.; Savaglio, M.; Childress, P.J.; Kumar, R.; et al. Gene-Metabolite network linked to inhibited bioenergetics in association with spaceflight-induced loss of male mouse quadriceps muscle. J. Bone Miner. Res. 2020, 35, 2049–2057. [Google Scholar] [CrossRef] [PubMed]
- Ohnishi, T.; Inoue, N.; Matsumoto, H.; Omatsu, T.; Ohira, Y.; Nagaoka, S. Cellular content of p53 protein in rat skin after exposure to the space environment. J. Appl. Physiol. 1996, 81, 183–185. [Google Scholar] [CrossRef]
- Lu, T.; Zhang, Y.; Kidane, Y.; Feiveson, A.; Stodieck, L.; Karouia, F.; Ramesh, G.; Rohde, L.; Wu, H. Cellular responses and gene expression profile changes due to bleomycin-induced DNA damage in human fibroblasts in space. PLoS ONE 2017, 12, e0170358. [Google Scholar] [CrossRef] [PubMed]
- Blaber, E.A.; Dvorochkin, N.; Lee, C.; Alwood, J.S.; Yousuf, R.; Pianetta, P.; Globus, R.K.; Burns, B.P.; Almeida, E.A. Microgravity induces pelvic bone loss through osteoclastic activity, osteocytic osteolysis, and osteoblastic cell cycle inhibition by CDKN1a/p21. PLoS ONE 2013, 8, e61372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammond, T.G.; Allen, P.L.; Birdsall, H.H. Effects of space flight on mouse liver versus kidney: Gene pathway analyses. Int. J. Mol. Sci. 2018, 19, 4106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Behnke, B.J.; Stabley, J.N.; McCullough, D.J.; Davis, R.T., 3rd; Dominguez, J.M., 2nd; Muller-Delp, J.M.; Delp, M.D. Effects of spaceflight and ground recovery on mesenteric artery and vein constrictor properties in mice. FASEB J. 2013, 27, 399–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailey, J.F.; Hargens, A.R.; Cheng, K.K.; Lotz, J.C. Effect of microgravity on the biomechanical properties of lumbar and caudal intervertebral discs in mice. J. Biomech. 2014, 47, 2983–2988. [Google Scholar] [CrossRef] [PubMed]
- Connor, M.K.; Hood, D.A. Effect of microgravity on the expression of mitochondrial enzymes in rat cardiac and skeletal muscles. J. Appl. Physiol. 1998, 84, 593–598. [Google Scholar] [CrossRef] [PubMed]
- Ogneva, I.V.; Loktev, S.S.; Sychev, V.N. Cytoskeleton structure and total methylation of mouse cardiac and lung tissue during space flight. PLoS ONE 2018, 13, e0192643. [Google Scholar] [CrossRef]
- Walls, S.; Diop, S.; Birse, R.; Elmen, L.; Gan, Z.; Kalvakuri, S.; Pineda, S.; Reddy, C.; Taylor, E.; Trinh, B.; et al. Prolonged exposure to microgravity reduces cardiac contractility and initiates remodeling in Drosophila. Cell Rep. 2020, 33, 108445. [Google Scholar] [CrossRef]
- Camberos, V.; Baio, J.; Mandujano, A.; Martinez, A.F.; Bailey, L.; Hasaniya, N.; Kearns-Jonker, M. The impact of spaceflight and microgravity on the human islet-1+ cardiovascular progenitor cell transcriptome. Int. J. Mol. Sci. 2021, 22, 3577. [Google Scholar] [CrossRef]
- Blaber, E.A.; Pecaut, M.J.; Jonscher, K.R. Spaceflight activates autophagy programs and the proteasome in mouse liver. Int. J. Mol. Sci. 2017, 18, 2062. [Google Scholar] [CrossRef]
- Suzuki, T.; Uruno, A.; Yumoto, A.; Taguchi, K.; Suzuki, M.; Harada, N.; Ryoke, R.; Naganuma, E.; Osanai, N.; Goto, A.; et al. Nrf2 contributes to the weight gain of mice during space travel. Commun. Biol. 2020, 3, 496. [Google Scholar] [CrossRef]
- Hayashi, T.; Kudo, T.; Fujita, R.; Fujita, S.I.; Tsubouchi, H.; Fuseya, S.; Suzuki, R.; Hamada, M.; Okada, R.; Muratani, M.; et al. Nuclear factor E2-related factor 2 (NRF2) deficiency accelerates fast fibre type transition in soleus muscle during space flight. Commun. Biol. 2021, 4, 787. [Google Scholar] [CrossRef]
- Lambeth, J.D.; Neish, A.S. Nox enzymes and new thinking on reactive oxygen: A double-edged sword revisited. Annu. Rev. Pathol. 2014, 9, 119–145. [Google Scholar] [CrossRef]
- Bedard, K.; Krause, K.H. The NOX family of ROS-generating NADPH oxidases: Physiology and pathophysiology. Physiol. Rev. 2007, 87, 245–313. [Google Scholar] [CrossRef] [PubMed]
- Brandes, R.P.; Weissmann, N.; Schroder, K. NADPH oxidases in cardiovascular disease. Free Radic. Biol. Med. 2010, 49, 687–706. [Google Scholar] [CrossRef] [PubMed]
- Liang, L.; Yuan, W.; Qu, L.; Li, H.; Zhang, L.; Fan, G.C.; Peng, T. Administration of losartan preserves cardiomyocyte size and prevents myocardial dysfunction in tail-suspended mice by inhibiting p47(phox) phosphorylation, NADPH oxidase activation and MuRF1 expression. J. Transl. Med. 2019, 17, 279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D.; Patel, V.V.; Ricciotti, E.; Zhou, R.; Levin, M.D.; Gao, E.; Yu, Z.; Ferrari, V.A.; Lu, M.M.; Xu, J.; et al. Cardiomyocyte cyclooxygenase-2 influences cardiac rhythm and function. Proc. Natl. Acad. Sci. USA 2009, 106, 7548–7552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sellers, R.S.; Radi, Z.A.; Khan, N.K. Pathophysiology of cyclooxygenases in cardiovascular homeostasis. Vet. Pathol. 2010, 47, 601–613. [Google Scholar] [CrossRef]
- Hughes-Fulford, M.; Rodenacker, K.; Jutting, U. Reduction of anabolic signals and alteration of osteoblast nuclear morphology in microgravity. J. Cell. Biochem. 2006, 99, 435–449. [Google Scholar] [CrossRef]
- Bondesen, B.A.; Mills, S.T.; Pavlath, G.K. The COX-2 pathway regulates growth of atrophied muscle via multiple mechanisms. Am. J. Physiol. Cell Physiol. 2006, 290, C1651–C1659. [Google Scholar] [CrossRef] [Green Version]
- Nakai, K.; Tanaka, S.; Sakai, A.; Nagashima, M.; Tanaka, M.; Otomo, H.; Nakamura, T. Cyclooxygenase-2 selective inhibition suppresses restoration of tibial trabecular bone formation in association with restriction of osteoblast maturation in skeletal reloading after hindlimb elevation of mice. Bone 2006, 39, 83–92. [Google Scholar] [CrossRef]
- Nishiyama, A.; Matsui, M.; Iwata, S.; Hirota, K.; Masutani, H.; Nakamura, H.; Takagi, Y.; Sono, H.; Gon, Y.; Yodoi, J. Identification of thioredoxin-binding protein-2/vitamin D(3) up-regulated protein 1 as a negative regulator of thioredoxin function and expression. J. Biol. Chem. 1999, 274, 21645–21650. [Google Scholar] [CrossRef] [Green Version]
- Versari, S.; Longinotti, G.; Barenghi, L.; Maier, J.A.; Bradamante, S. The challenging environment on board the International Space Station affects endothelial cell function by triggering oxidative stress through thioredoxin interacting protein overexpression: The ESA-SPHINX experiment. FASEB J. 2013, 27, 4466–4475. [Google Scholar] [CrossRef]
- Saadatzadeh, M.R.; Bijangi-Vishehsaraei, K.; Hong, P.; Bergmann, H.; Haneline, L.S. Oxidant hypersensitivity of Fanconi anemia type C-deficient cells is dependent on a redox-regulated apoptotic pathway. J. Biol. Chem. 2004, 279, 16805–16812. [Google Scholar] [CrossRef] [Green Version]
- Galow, A.M.; Wolfien, M.; Muller, P.; Bartsch, M.; Brunner, R.M.; Hoeflich, A.; Wolkenhauer, O.; David, R.; Goldammer, T. Integrative cluster analysis of whole hearts reveals proliferative cardiomyocytes in adult mice. Cells 2020, 9, 1144. [Google Scholar] [CrossRef]
- Tong, Y.F.; Wang, Y.; Ding, Y.Y.; Li, J.M.; Pan, X.C.; Lu, X.L.; Chen, X.H.; Liu, Y.; Zhang, H.G. Cyclin-Dependent Kinase Inhibitor p21WAF1/CIP1 facilitates the development of cardiac hypertrophy. Cell Physiol. Biochem. 2017, 42, 1645–1656. [Google Scholar] [CrossRef]
- Xu, M.; Wan, C.X.; Huang, S.H.; Wang, H.B.; Fan, D.; Wu, H.M.; Wu, Q.Q.; Ma, Z.G.; Deng, W.; Tang, Q.Z. Oridonin protects against cardiac hypertrophy by promoting P21-related autophagy. Cell Death Dis. 2019, 10, 403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, S.; Xu, M.; Liu, L.; Yang, J.; Wang, H.; Wan, C.; Deng, W.; Tang, Q. Autophagy is involved in the protective effect of p21 on LPS-induced cardiac dysfunction. Cell Death Dis. 2020, 11, 554. [Google Scholar] [CrossRef]
- Hauck, L.; Grothe, D.; Billia, F. p21(CIP1/WAF1)-dependent inhibition of cardiac hypertrophy in response to Angiotensin II involves Akt/Myc and pRb signaling. Peptides 2016, 83, 38–48. [Google Scholar] [CrossRef]
- Burton, P.B.; Yacoub, M.H.; Barton, P.J. Cyclin-dependent kinase inhibitor expression in human heart failure. A comparison with fetal development. Eur. Heart J. 1999, 20, 604–611. [Google Scholar] [CrossRef] [Green Version]
- Pei, L.; Melmed, S. Isolation and characterization of a pituitary tumor-transforming gene (PTTG). Mol. Endocrinol. 1997, 11, 433–441. [Google Scholar] [CrossRef]
- Hoglund, A.; Stromvall, K.; Li, Y.; Forshell, L.P.; Nilsson, J.A. Chk2 deficiency in Myc overexpressing lymphoma cells elicits a synergistic lethal response in combination with PARP inhibition. Cell Cycle 2011, 10, 3598–3607. [Google Scholar] [CrossRef] [Green Version]
- Seawright, J.W.; Sridharan, V.; Landes, R.D.; Cao, M.; Singh, P.; Koturbash, I.; Mao, X.W.; Miousse, I.R.; Singh, S.P.; Nelson, G.A.; et al. Effects of low-dose oxygen ions and protons on cardiac function and structure in male C57BL/6J mice. Life Sci. Space Res. 2019, 20, 72–84. [Google Scholar] [CrossRef] [PubMed]
- Prasad, B.; Grimm, D.; Strauch, S.M.; Erzinger, G.S.; Corydon, T.J.; Lebert, M.; Magnusson, N.E.; Infanger, M.; Richter, P.; Kruger, M. Influence of microgravity on apoptosis in cells, tissues, and other systems in vivo and in vitro. Int. J. Mol. Sci. 2020, 21, 9373. [Google Scholar] [CrossRef] [PubMed]
- Congdon, C.C.; Allebban, Z.; Gibson, L.A.; Kaplansky, A.; Strickland, K.M.; Jago, T.L.; Johnson, D.L.; Lange, R.D.; Ichiki, A.T. Lymphatic tissue changes in rats flown on Spacelab Life Sciences-2. J. Appl. Physiol. 1996, 81, 172–177. [Google Scholar] [CrossRef] [PubMed]
- Gridley, D.S.; Mao, X.W.; Stodieck, L.S.; Ferguson, V.L.; Bateman, T.A.; Moldovan, M.; Cunningham, C.E.; Jones, T.A.; Slater, J.M.; Pecaut, M.J. Changes in mouse thymus and spleen after return from the STS-135 mission in space. PLoS ONE 2013, 8, e75097. [Google Scholar] [CrossRef] [PubMed]
- Locksley, R.M.; Killeen, N.; Lenardo, M.J. The TNF and TNF receptor superfamilies: Integrating mammalian biology. Cell 2001, 104, 487–501. [Google Scholar] [CrossRef] [Green Version]
- Gridley, D.S.; Slater, J.M.; Luo-Owen, X.; Rizvi, A.; Chapes, S.K.; Stodieck, L.S.; Ferguson, V.L.; Pecaut, M.J. Spaceflight effects on T lymphocyte distribution, function and gene expression. J. Appl. Physiol. 2009, 106, 194–202. [Google Scholar] [CrossRef]
- Crucian, B.; Stowe, R.P.; Mehta, S.; Quiriarte, H.; Pierson, D.; Sams, C. Alterations in adaptive immunity persist during long-duration spaceflight. NPJ Microgravity 2015, 1, 15013. [Google Scholar] [CrossRef]
- Chang, T.T.; Walther, I.; Li, C.F.; Boonyaratanakornkit, J.; Galleri, G.; Meloni, M.A.; Pippia, P.; Cogoli, A.; Hughes-Fulford, M. The Rel/NF-kappaB pathway and transcription of immediate early genes in T cell activation are inhibited by microgravity. J. Leukoc. Biol. 2012, 92, 1133–1145. [Google Scholar] [CrossRef] [Green Version]
- Gertz, M.L.; Chin, C.R.; Tomoiaga, D.; MacKay, M.; Chang, C.; Butler, D.; Afshinnekoo, E.; Bezdan, D.; Schmidt, M.A.; Mozsary, C.; et al. Multi-omic, single-cell, and biochemical profiles of astronauts guide Pharmacological strategies for returning to gravity. Cell Rep. 2020, 33, 108429. [Google Scholar] [CrossRef]
- Sasi, S.P.; Yan, X.; Zuriaga-Herrero, M.; Gee, H.; Lee, J.; Mehrzad, R.; Song, J.; Onufrak, J.; Morgan, J.; Enderling, H.; et al. Different sequences of fractionated low-dose proton and single iron-radiation-induced divergent biological responses in the heart. Radiat. Res. 2017, 188, 191–203. [Google Scholar] [CrossRef]
- Garikipati, V.N.S.; Arakelyan, A.; Blakely, E.A.; Chang, P.Y.; Truongcao, M.M.; Cimini, M.; Malaredy, V.; Bajpai, A.; Addya, S.; Bisserier, M.; et al. Long-term effects of very low dose particle radiation on gene expression in the heart: Degenerative disease risks. Cells 2021, 10, 387. [Google Scholar] [CrossRef]
- Hughson, R.L.; Robertson, A.D.; Arbeille, P.; Shoemaker, J.K.; Rush, J.W.; Fraser, K.S.; Greaves, D.K. Increased postflight carotid artery stiffness and inflight insulin resistance resulting from 6-mo spaceflight in male and female astronauts. Am. J. Physiol. Heart Circ. Physiol. 2016, 310, H628–H638. [Google Scholar] [CrossRef] [Green Version]
- Mehta, S.K.; Laudenslager, M.L.; Stowe, R.P.; Crucian, B.E.; Feiveson, A.H.; Sams, C.F.; Pierson, D.L. Latent virus reactivation in astronauts on the international space station. NPJ Microgravity 2017, 3, 11. [Google Scholar] [CrossRef] [Green Version]
- Roberts, D.R.; Inglesby, D.C.; Brown, T.R.; Collins, H.R.; Eckert, M.A.; Asemani, D. Longitudinal change in ventricular volume is accelerated in astronauts undergoing long-duration spaceflight. Aging Brain 2021, 1, 100017. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Nastou, K.C.; Lyon, D.; Kirsch, R.; Pyysalo, S.; Doncheva, N.T.; Legeay, M.; Fang, T.; Bork, P.; et al. The STRING database in 2021: Customizable protein-protein networks, and functional characterization of user-uploaded gene/measurement sets. Nucleic Acids Res. 2021, 49, D605–D612. [Google Scholar] [CrossRef]
- Chen, J.; Bardes, E.E.; Aronow, B.J.; Jegga, A.G. ToppGene Suite for gene list enrichment analysis and candidate gene prioritization. Nucleic Acids Res. 2009, 37, W305–W311. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Gene Name | Fold Regulation |
|---|---|---|
| Nox1 | NADPH oxidase 1 | 6.62 |
| Tpo | Thyroid peroxidase | 2.35 |
| Slc41a3 | Solute carrier family 41, member 3 | 1.97 |
| Txnip | Thioredoxin interacting protein | 1.94 |
| Prnp | Prion protein | 1.44 |
| Txnrd3 | Thioredoxin reductase 3 | 1.43 |
| Gsr | Glutathione reductase | 1.31 |
| Prdx6 | Peroxiredoxin 6 | 1.23 |
| Cat | Catalase | 1.20 |
| Xpa | Xeroderma pigmentosum, complementation group A | 1.18 |
| Sod1 | Superoxide dismutase 1, soluble | 1.18 |
| Ncf2 | Neutrophil cytosolic factor 2 | −1.49 |
| Fancc | Fanconi anemia, complementation group C | −1.62 |
| Ptgs2 | Prostaglandin-endoperoxide synthase 2 | −3.44 |
| Gene Symbol | Gene Name | Fold Regulation |
|---|---|---|
| Cdkn1a | Cyclin-dependent kinase inhibitor 1A (P21) | 6.80 |
| Myc | Myelocytomatosis oncogene | 3.91 |
| Hif1a | Hypoxia inducible factor 1, alpha subunit | 1.58 |
| Pmaip1 | Phorbol-12-myristate-13-acetate-induced protein 1 | 1.54 |
| Prkca | Protein kinase C, alpha | 1.49 |
| Trp53bp2 | Transformation related protein 53 binding protein 2 | 1.41 |
| Apex1 | Apurinic/apyrimidinic endonuclease 1 | 1.28 |
| Ercc1 | Excision repair cross-complementing rodent repair deficiency, complementation group 1 | 1.27 |
| Foxo3 | Forkhead box O3 | 1.27 |
| Mlh1 | MutL homolog 1 (E. coli) | 1.19 |
| Pten | Phosphatase and tensin homolog | 1.18 |
| Xrcc5 | X-ray repair complementing defective repair in Chinese hamster cells 5 | 1.13 |
| Atr | Ataxia telangiectasia and rad3 related | 1.10 |
| Cdk4 | Cyclin-dependent kinase 4 | −1.16 |
| Apaf1 | Apoptotic peptidase activating factor 1 | −1.41 |
| Tnfrsf10b | Tumor necrosis factor receptor superfamily, member 10b | −1.92 |
| Chek2 | CHK2 checkpoint homolog (S. pombe) | −2.15 |
| Traf1 | Tnf receptor-associated factor 1 | −2.25 |
| Jun | Jun oncogene | −2.31 |
| Pttg1 | Pituitary tumor-transforming gene 1 | −2.42 |
| Btg2 | B-cell translocation gene 2, anti-proliferative | −3.50 |
| Cdk1 | Cyclin-dependent kinase 1 | −4.11 |
| Tnf | Tumor necrosis factor | −9.86 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kumar, A.; Tahimic, C.G.T.; Almeida, E.A.C.; Globus, R.K. Spaceflight Modulates the Expression of Key Oxidative Stress and Cell Cycle Related Genes in Heart. Int. J. Mol. Sci. 2021, 22, 9088. https://doi.org/10.3390/ijms22169088
Kumar A, Tahimic CGT, Almeida EAC, Globus RK. Spaceflight Modulates the Expression of Key Oxidative Stress and Cell Cycle Related Genes in Heart. International Journal of Molecular Sciences. 2021; 22(16):9088. https://doi.org/10.3390/ijms22169088
Chicago/Turabian StyleKumar, Akhilesh, Candice G. T. Tahimic, Eduardo A. C. Almeida, and Ruth K. Globus. 2021. "Spaceflight Modulates the Expression of Key Oxidative Stress and Cell Cycle Related Genes in Heart" International Journal of Molecular Sciences 22, no. 16: 9088. https://doi.org/10.3390/ijms22169088
APA StyleKumar, A., Tahimic, C. G. T., Almeida, E. A. C., & Globus, R. K. (2021). Spaceflight Modulates the Expression of Key Oxidative Stress and Cell Cycle Related Genes in Heart. International Journal of Molecular Sciences, 22(16), 9088. https://doi.org/10.3390/ijms22169088