
Neuroblastoma GD2 Expression and Computational Analysis of Aptamer-Based Bioaffinity Targeting



{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Epidemiology of NB
3. Genomics of Neuroblastoma-GD2 Synthase Gene
4. Minimal Residual Disease in NB
5. GD2 as a Diagnostic Biomarker
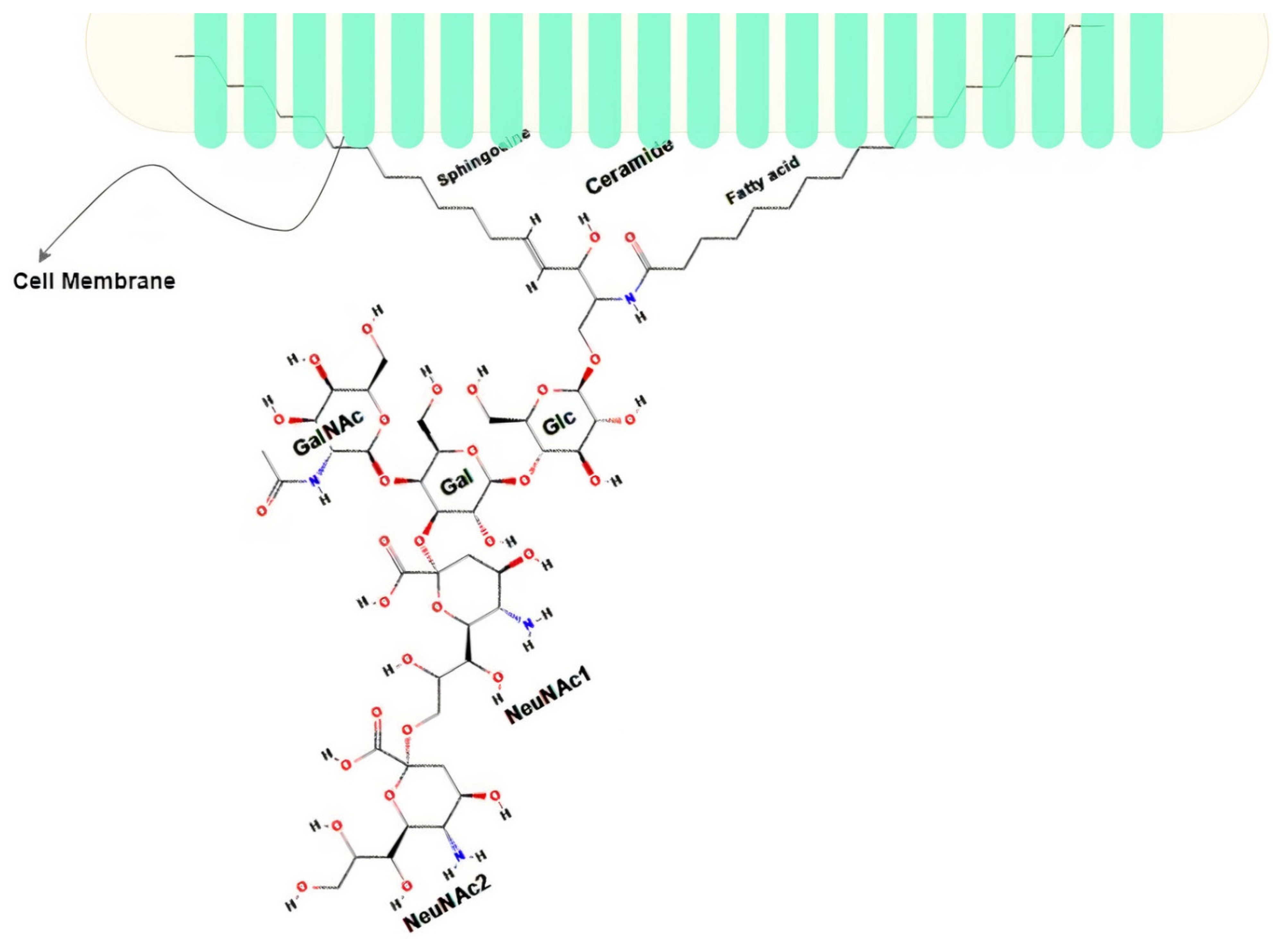
5.1. Disialoganglioside (GD2)
5.2. Neuroblastoma Detection: Focusing on GD2
5.3. NB Detection Techniques
5.4. GD2 Detection with Monoclonal Antibodies
5.5. GD2 Detection Using Aptamers
6. Computational Approaches for GD2 Characterization
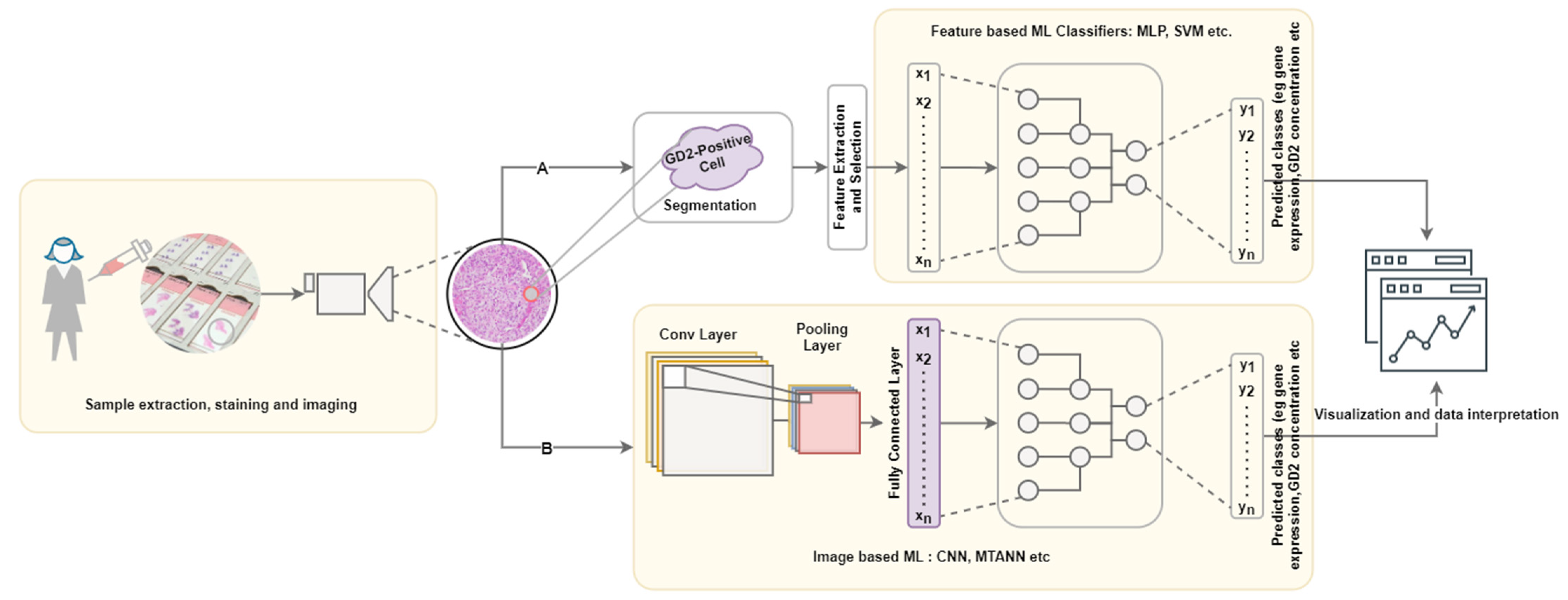
6.1. Immunohistolocal Image Processing Pipeline

6.2. Feature Extraction
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Castleberry, R.P. Biology and Treatment of Neuroblastoma. Pediatr. Clin. N. Am. 1997, 44, 919–937. [Google Scholar] [CrossRef]
- Park, J.R.; Bagatell, R. Chapter 24—Neuroblastoma. In Lanzkowsky’s Manual of Pediatric Hematology and Oncology, 6th ed.; Lanzkowsky, P., Lipton, J.M., Fish, J.D., Eds.; Academic Press: San Diego, CA, USA, 2016; pp. 473–490. [Google Scholar] [CrossRef]
- Cohn, S.L.; Pearson, A.D.J.; London, W.B.; Monclair, T.; Ambros, P.F.; Brodeur, G.M.; Faldum, A.; Hero, B.; Iehara, T.; Machin, D.; et al. The International Neuroblastoma Risk Group (Inrg) Classification System: An Inrg Task Force Report. J. Clin. Oncol. 2009, 27, 289–297. [Google Scholar] [CrossRef]
- Pastor, E.R.; Mousa, S.A. Current management of neuroblastoma and future direction. Crit. Rev. Oncol. Hematol. 2019, 138, 38–43. [Google Scholar] [CrossRef]
- Sait, S.; Modak, S.I. Anti-GD2 immunotherapy for neuroblastoma. Expert Rev. Anticancer Ther. 2017, 17, 889–904. [Google Scholar] [CrossRef]
- Yu, A.L.; Gilman, A.L.; Ozkaynak, M.F.; London, W.B.; Kreissman, S.G.; Chen, H.X.; Smith, M.; Anderson, B.; Villablanca, J.G.; Matthay, K.K.; et al. Anti-GD2 Antibody with GM-CSF, Interleukin-2, and Isotretinoin for Neuroblastoma. N. Engl. J. Med. 2010, 363, 1324–1334. [Google Scholar] [CrossRef] [Green Version]
- Quaglia, M.P.L. State of the art in oncology: High risk neuroblastoma, alveolar rhabdomyosarcoma, desmoplastic small round cell tumor, and post-text 3 and 4 hepatoblastoma. J. Pediatric Surg. 2014, 49, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Matthay, K.; O’Leary, M.; Ramsay, N.; Villablanca, J.; Reynolds, C.P.; Atkinson, J.; Haase, G.; Stram, D.; Seeger, R. Role of myeloablative therapy in improved outcome for high risk neuroblastoma: Review of recent children’s cancer group results. Eur. J. Cancer 1995, 31, 572–575. [Google Scholar] [CrossRef]
- Matthay, K.K.; Reynolds, C.P.; Seeger, R.C.; Shimada, H.; Adkins, E.S.; Haas-Kogan, D.; Gerbing, R.B.; London, W.B.; Villablanca, J.G. Long-Term Results for Children with High-Risk Neuroblastoma Treated on a Randomized Trial of Myeloablative Therapy Followed by 13-cis-Retinoic Acid: A Children’s Oncology Group Study. J. Clin. Oncol. 2009, 27, 1007–1013. [Google Scholar] [CrossRef] [Green Version]
- Morgenstern, D.A.; Irwin, M.S. Current and Future Strategies for Treatment of Relapsed Neuroblastoma. In Neuroblastoma; Ray, S.K., Ed.; Academic Press: San Diego, CA, USA, 2019; pp. 263–281. [Google Scholar] [CrossRef]
- Cheever, M.A.; Allison, J.P.; Ferris, A.S.; Finn, O.J.; Hastings, B.M.; Hecht, T.T.; Mellman, I.; Prindiville, S.A.; Viner, J.L.; Weiner, L.M.; et al. The prioritization of cancer antigens: A national cancer institute pilot project for the accel-eration of translational research. Clin. Cancer Res. 2009, 15, 5323–5337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Z.-L.; Ladisch, S.; Feig, S.; Ulsh, L.; Schwartz, E.; Floutsis, G.; Wiley, F.; Lenarsky, C.; Seeger, R. Shedding of GD2 ganglioside by human neuroblastoma. Int. J. Cancer 1987, 39, 73–76. [Google Scholar] [CrossRef] [PubMed]
- Colon, N.C.; Chung, D.H. Neuroblastoma. Adv. Pediatr. 2011, 58, 297–311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maris, J.M. Recent advances in neuroblastoma. N. Engl. J. Med. 2010, 362, 2202–2211. [Google Scholar] [CrossRef] [Green Version]
- Esiashvili, N.; Anderson, C.; Katzenstein, H.M. Neuroblastoma. Curr. Probl. Cancer 2009, 33, 333–360. [Google Scholar] [CrossRef]
- Trigg, R.M.; Turner, S.D. ALK in Neuroblastoma: Biological and Therapeutic Implications. Cancers 2018, 10, 113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Umapathy, G.; Mendoza-Garcia, P.; Hallberg, B.; Palmer, R.H. Targeting anaplastic lymphoma kinase in neuroblastoma. APMIS 2019, 127, 288–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simon, T.; Berthold, F.; Borkhardt, A.; Kremens, B.; Decarolis, B.; Hero, B. Treatment and outcomes of patients with relapsed, high-risk neuroblastoma: Results of German trials. Pediatr. Blood Cancer 2010, 56, 578–583. [Google Scholar] [CrossRef]
- Speleman, F.; De Preter, K.; Vandesompele, J. Neuroblastoma genetics and phenotype: A tale of heterogeneity. Semin. Cancer Biol. 2011, 21, 238–244. [Google Scholar] [CrossRef]
- Kohl, N.E.; Kanda, N.; Schreck, R.R.; Bruns, G.; Latt, S.A.; Gilbert, F.; Alt, F.W. Transposition and amplification of onco-gene-related sequences in human neuroblastomas. Cell 1983, 35, 359–367. [Google Scholar] [CrossRef]
- Huang, M.; Weiss, W.A. Neuroblastoma and mycn. Cold Spring Harb. Perspect. Med. 2013, 3, a014415. [Google Scholar] [CrossRef]
- Lee, E.; Lee, J.W.; Lee, B.; Park, K.; Shim, J.; Yoo, K.H.; Koo, H.H.; Sung, K.W.; Park, W.-Y. Genomic profile of MYCN non-amplified neuroblastoma and potential for immunotherapeutic strategies in neuroblastoma. BMC Med. Genom. 2020, 13, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Attiyeh, E.F.; London, W.B.; Mossé, Y.P.; Wang, Q.; Winter, C.; Khazi, D.; McGrady, P.W.; Seeger, R.C.; Look, A.T.; Shimada, H.; et al. Chromosome 1p and 11q Deletions and Outcome in Neuroblastoma. N. Engl. J. Med. 2005, 353, 2243–2253. [Google Scholar] [CrossRef]
- Beheshti, B.; Braude, I.; Marrano, P.; Thorner, P.; Zielenska, M.; Squire, J.A. Chromosomal Localization of DNA Amplifications in Neuroblastoma Tumors Using cDNA Microarray Comparative Genomic Hybridization1. Neoplasia 2003, 5, 53–62. [Google Scholar] [CrossRef] [Green Version]
- Brinkschmidt, C.; Christiansen, H.; Terpe, H.J.; Simon, R.; Boecker, W.; Lampert, F.; Stoerkel, S. Comparative genomic hy-bridization (cgh) analysis of neuroblastomas—An important methodological approach in paediatric tumour pathology. J. Pathol. 1997, 181, 394–400. [Google Scholar] [CrossRef]
- Nazha, B.; Inal, C.; Owonikoko, T.K. Disialoganglioside GD2 Expression in Solid Tumors and Role as a Target for Cancer Therapy. Front. Oncol. 2020, 10, 1000. [Google Scholar] [CrossRef] [PubMed]
- Trinchera, M.; Parini, R.; Indellicato, R.; Domenighini, R.; dall’Olio, F. Diseases of ganglioside biosynthesis: An expanding group of congenital disorders of glycosylation. Mol. Genet. Metab. 2018, 124, 230–237. [Google Scholar] [CrossRef]
- Beta-1,4-N-acetyl-galactosaminyltransferase 1. Available online: https://www.ncbi.nlm.nih.gov/gene/2583 (accessed on 11 April 2021).
- Cheung, N.-K.V.; Dyer, M.A. Neuroblastoma: Developmental biology, cancer genomics and immunotherapy. Nat. Rev. Cancer 2013, 13, 397–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mansoori, M.; Roudi, R.; Abbasi, A.; Abolhasani, M.; Rad, I.A.; Shariftabrizi, A.; Madjd, Z. High GD2 expression defines breast cancer cells with enhanced invasiveness. Exp. Mol. Pathol. 2019, 109, 25–35. [Google Scholar] [CrossRef]
- Furukawa, K.; Takamiya, K.; Furukawa, K. 1,4-n-acetylgalactosaminyltransferase—gm2/gd2 synthase: A key enzyme to control the synthesis of brain-enriched complex gangliosides. Biochim. Et Biophys. Acta Gen. Subj. 2002, 1573, 356–362. [Google Scholar] [CrossRef]
- Yoshida, H.; Koodie, L.; Jacobsen, K.; Hanzawa, K.; Miyamoto, Y.; Yamamoto, M. B4galnt1 induces angiogenesis, anchorage independence growth and motility, and promotes tumorigenesis in melanoma by induction of ganglioside gm2/gd2. Sci. Rep. 2020, 10, 1–12. [Google Scholar]
- Yamashiro, S.; Ruan, S.; Tai, T.; Lloyd, K.O.; Shiku, H.; Furukawa, K. Genetic and enzymatic basis for the differential expression of GM2 and GD2 gangliosides in human cancer cell lines. Cancer Res. 1993, 53, 5395–5400. [Google Scholar]
- Hoon, D.S.; Kuo, C.T.; Wen, S.; Wang, H.; Metelitsa, L.; Reynolds, C.P.; Seeger, R.C. Ganglioside GM2/GD2 Synthetase mRNA Is a Marker for Detection of Infrequent Neuroblastoma Cells in Bone Marrow. Am. J. Pathol. 2001, 159, 493–500. [Google Scholar] [CrossRef] [Green Version]
- Uemura, S.; Ishida, T.; Thwin, K.K.M.; Yamamoto, N.; Tamura, A.; Kishimoto, K.; Hasegawa, D.; Kosaka, Y.; Nino, N.; Lin, K.S.; et al. Dynamics of Minimal Residual Disease in Neuroblastoma Patients. Front. Oncol. 2019, 9, 455. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.K.; Sondel, P.M. Anti-GD2 Strategy in the Treatment of Neuroblastoma. Drugs Futur. 2010, 35, 665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Della Starza, I.; Chiaretti, S.; De Propris, M.S.; Elia, L.; Cavalli, M.; De Novi, L.A.; Soscia, R.; Messina, M.; Vitale, A.; Guarini, A.; et al. Minimal Residual Disease in Acute Lymphoblastic Leukemia: Technical and Clinical Advances. Front. Oncol. 2019, 9, 726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campana, D. Minimal residual disease. Leuk. Suppl. 2012, 1, S3–S4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kholodenko, I.V.; Kalinovsky, D.V.; Doronin, I.I.; Deyev, S.M.; Kholodenko, R.V. Neuroblastoma Origin and Therapeutic Targets for Immunotherapy. J. Immunol. Res. 2018, 2018, 1–25. [Google Scholar] [CrossRef]
- Yu, R.K.; Tsai, Y.-T.; Ariga, T.; Yanagisawa, M. Structures, biosynthesis, and functions of gangliosides—An overview. J. Oleo Sci. 2011, 60, 537–544. [Google Scholar] [CrossRef] [Green Version]
- Hung, J.-T.; Yu, A.L. Chapter 4—gd2-targeted immunotherapy of neuroblastoma. In Neuroblastoma; Ray, S.K., Ed.; Academic Press: San Diego, CA, USA, 2019; pp. 63–78. [Google Scholar]
- Suzuki, M.; Cheung, N.-K.V. Disialoganglioside GD2 as a therapeutic target for human diseases. Expert Opin. Ther. Targets 2014, 19, 349–362. [Google Scholar] [CrossRef] [PubMed]
- Hansen, J.N.; Li, X.; Zheng, Y.G.; Lotta, L.T.; Dedhe, A.; Schor, N.F. Using chemistry to target neuroblastoma. ACS Chem. Neurosci. 2017, 8, 2118–2123. [Google Scholar] [CrossRef]
- Cavdarli, S.; Groux-Degroote, S.; Delannoy, P. Gangliosides: The Double-Edge Sword of Neuro-Ectodermal Derived Tumors. Biomolecules 2019, 9, 311. [Google Scholar] [CrossRef] [Green Version]
- Seeger, R.C.; Reynolds, C.P.; Gallego, R.; Stram, D.O.; Gerbing, R.B.; Matthay, K.K. Quantitative Tumor Cell Content of Bone Marrow and Blood as a Predictor of Outcome in Stage IV Neuroblastoma: A Children’s Cancer Group Study. J. Clin. Oncol. 2000, 18, 4067–4076. [Google Scholar] [CrossRef]
- Cheung, I.Y.; Cheung, N.-K.V. Detection of microscopic disease: Comparing histology, immunocytology, and RT-PCR of tyrosine hydroxylase, GAGE, andMAGE. Med. Pediatr. Oncol. 2001, 36, 210–212. [Google Scholar] [CrossRef]
- Méhes, G.; Luegmayr, A.; Kornmüller, R.; Ambros, I.M.; Ladenstein, R.; Gadner, H.; Ambros, P.F. Detection of Disseminated Tumor Cells in Neuroblastoma: 3 Log Improvement in Sensitivity by Automatic Immunofluorescence plus FISH (AIPF) Analysis Compared with Classical Bone Marrow Cytology. Am. J. Pathol. 2003, 163, 393–399. [Google Scholar] [CrossRef]
- Moss, T.J.; Reynolds, C.P.; Sather, H.N.; Romansky, S.G.; Hammond, G.D.; Seeger, R.C. Prognostic value of immuno-cytologic detection of bone marrow metastases in neuroblastoma. N. Engl. J. Med. 1991, 324, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Merugu, S.; Chen, L.; Gavens, E.; Gabra, H.; Brougham, M.; Makin, G.; Ng, A.; Murphy, D.; Gabriel, A.S.; Robinson, M.L.; et al. Detection of Circulating and Disseminated Neuroblastoma Cells Using the ImageStream Flow Cytometer for Use as Predictive and Pharmacodynamic Biomarkers. Clin. Cancer Res. 2019, 26, 122–134. [Google Scholar] [CrossRef]
- Reynolds, C.P. Detection and treatment of minimal residual disease in high-risk neuroblastoma. Pediatr. Transplant. 2004, 8, 56–66. [Google Scholar] [CrossRef]
- Jeevanandam, J.; Tan, K.X.; Danquah, M.K.; Guo, H.-B.; Turgeson, A. Advancing Aptamers as Molecular Probes for Cancer Theranostic Applications—The Role of Molecular Dynamics Simulation. Biotechnol. J. 2019, 15. [Google Scholar] [CrossRef]
- Acquah, C.; Chan, Y.W.; Pan, S.; Yon, L.S.; Ongkudon, C.M.; Guo, H.-B.; Danquah, M.K. Characterisation of aptamer-anchored poly(EDMA-co-GMA) monolith for high throughput affinity binding. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Maris, J.M.; Hogarty, M.D.; Bagatell, R.; Cohn, S.L. Neuroblastoma. Lancet 2007, 369, 2106–2120. [Google Scholar] [CrossRef]
- Kuci, Z.; Seitz, G.; CuCI, S.; Kreyenberg, H.; Schumm, M.; Lang, P.; Niethammer, D.; Handgretinger, R.; Bruchelt, G. Pitfalls in detection of contaminating neuroblastoma cells by tyrosine hydroxylase rt-pcr due to catecholamine-producing hema-topoietic cells. Anticancer. Res. 2006, 26, 2075–2080. [Google Scholar]
- Berthold, F.; Schneider, A.; Schumacher, R.; Bosslet, K. Detection of Minimal Disease in Bone Marrow of Neuroblastoma Patients by Immunofluorescence. Pediatr. Hematol. Oncol. 1989, 6, 73–83. [Google Scholar] [CrossRef]
- Cheung, N.K.; Von Hoff, D.D.; Strandjord, S.E.; Coccia, P.F. Detection of neuroblastoma cells in bone marrow using GD2 specific monoclonal antibodies. J. Clin. Oncol. 1986, 4, 363–369. [Google Scholar] [CrossRef]
- Swerts, K.; Ambros, P.F.; Brouzes, C.; Navarro, J.M.F.; Gross, N.; Rampling, D.; Schumacher-Kuckelkorn, R.; Sementa, A.R.; Ladenstein, R.; Beiske, K. Standardization of the immunocytochemical detection of neuroblastoma cells in bone marrow. J. Histochem. Cytochem. Off. J. Histochem. Soc. 2005, 53, 1433–1440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Treleaven, J.; Ugelstad, J.; Philip, T.; Gibson, F.; Rembaum, A.; Caine, G.; Kemshead, J. Removal of neuroblastoma cells from bone marrow with monoclonal antibodies conjugated to magnetic microspheres. Lancet 1984, 323, 70–73. [Google Scholar] [CrossRef]
- Cai, W.; Hong, H. Applications of Aptamers in Targeted Imaging: State of the Art. Curr. Top. Med. Chem. 2015, 15, 1138–1152. [Google Scholar] [CrossRef]
- Hays, E.M.; Duan, W.; Shigdar, S. Aptamers and glioblastoma: Their potential use for imaging and therapeutic applications. Int. J. Mol. Sci. 2017, 18, 2576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruscito, A.; DeRosa, M.C. Small-Molecule binding aptamers: Selection strategies, characterization, and applications. Front. Chem. 2016, 4, 14. [Google Scholar] [CrossRef]
- Jeevanandam, J.; Sabbih, G.; Tan, K.X.; Danquah, M.K. Oncological Ligand-Target Binding Systems and Developmental Approaches for Cancer Theranostics. Mol. Biotechnol. 2021, 63, 167–183. [Google Scholar] [CrossRef]
- Hong, K.L.; Sooter, L.J. Single-Stranded DNA Aptamers against Pathogens and Toxins: Identification and Biosensing Applications. BioMed Res. Int. 2015, 2015, 1–31. [Google Scholar] [CrossRef]
- Tang, J.; Yu, T.; Guo, L.; Xie, J.; Shao, N.; He, Z. In Vitro selection of DNA aptamer against abrin toxin and aptamer-based abrin direct detection. Biosens. Bioelectron. 2007, 22, 2456–2463. [Google Scholar] [CrossRef]
- Delač, M.; Motaln, H.; Ulrich, H.; Lah, T.T. Aptamer for imaging and therapeutic targeting of brain tumor glioblastoma. Cytom. Part. A 2015, 87, 806–816. [Google Scholar] [CrossRef] [Green Version]
- Borghei, Y.-S.; Hosseini, M.; Dadmehr, M.; Hosseinkhani, S.; Ganjali, M.R.; Sheikhnejad, R. Visual detection of cancer cells by colorimetric aptasensor based on aggregation of gold nanoparticles induced by DNA hybridization. Anal. Chim. Acta 2015, 904, 92–97. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, M. A novel ph-sensitive multifunctional dna nanomedicine: An enhanced and harmless gd2 ap-tamer-mediated strategy for guiding neuroblastoma antitumor therapy. Front. Chem. 2021, 4, 14. [Google Scholar]
- di Pietrantonio, F.; Cannatà, D.; Benetti, M. Chapter 8—Biosensor technologies based on nanomaterials. In Functional Nanostructured Interfaces for Environmental and Biomedical Applications, Micro and Nano Technologies; Dinca, V., Suchea, M.P., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 181–242. [Google Scholar]
- Murray, J.L.; Kleinerman, E.S.; Jia, S.-F.; Rosenblum, M.G.; Eton, O.; Buzaid, A.; Legha, S.; Ross, M.I.; Thompson, L.; Mujoo, K.; et al. Phase ia/ib trial of anti-gd2 chimeric monoclonal antibody 14.18 (ch l4.18) and recombinant human granulocyte-macrophage colony-stimulating factor (rhgm-csf) in metastatic melanoma. J. Immunother. 1996, 19, 206–217. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Tang, Z.; Kim, Y.; Nie, H.; Huang, Y.F.; He, X.; Deng, K.; Wang, K.; Tan, W. In Vivo Fluorescence Imaging of Tumors using Molecular Aptamers Generated by Cell-SELEX. Chem. Asian J. 2010, 5, 2209–2213. [Google Scholar] [CrossRef] [PubMed]
- Hong, H.; Goel, S.; Zhang, Y.; Cai, W. Molecular imaging with nucleic acid aptamers. Curr. Med. Chem. 2011, 18, 4195–4205. [Google Scholar] [CrossRef] [Green Version]
- Shi, H.; He, X.; Wang, K.; Wu, X.; Ye, X.; Guo, Q.; Tan, W.; Qing, Z.; Yang, X.; Zhou, B. Activatable aptamer probe for con-trast-enhanced in vivo cancer imaging based on cell membrane protein-triggered conformation alteration. Proc. Natl. Acad. Sci. USA 2011, 108, 3900–3905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, E.-K.; Kim, B.; Choi, Y.; Ro, Y.; Cho, E.-J.; Lee, J.H.; Ryu, S.-H.; Suh, J.-S.; Haam, S.; Huh, Y.-M. Aptamer-Conjugated magnetic nanoparticles enable efficient targeted detection of integrin v3 via magnetic resonance imaging. J. Biomed. Mater. Res. Part A 2014, 102, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Heo, D.; Ku, M.; Kim, J.-H.; Yang, J.; Suh, J.-S. Aptamer-modified Magnetic Nanosensitizer for in vivo MR imaging of HER2-expressing Cancer. Nanoscale Res. Lett. 2018, 13, 288. [Google Scholar] [CrossRef] [Green Version]
- Dai, Y.; Zhang, A.; You, J.; Li, J.; Xu, H.; Xu, K. Fabrication of AS1411 aptamer functionalized Gd2O3-based molecular magnetic resonance imaging (mMRI) nanoprobe for renal carcinoma cell imaging. RSC Adv. 2015, 5, 77204–77210. [Google Scholar] [CrossRef]
- Zhu, H.; Zhang, L.; Liu, Y.; Zhou, Y.; Wang, K.; Xie, X.; Song, L.; Wang, D.; Han, C.; Chen, Q. Aptamer-PEG-Modified Fe3O4@Mn as a novel T1- and T2- dual-model MRI contrast agent targeting hypoxia-induced cancer stem cells. Sci. Rep. 2016, 6, 39245. [Google Scholar] [CrossRef] [Green Version]
- Lu, F.-M.; Yuan, Z. Pet/spect molecular imaging in clinical neuroscience: Recent advances in the investigation of cns diseases. Quant. Imaging Med. Surg. 2015, 5, 433–447. [Google Scholar] [PubMed]
- Li, J.; Zheng, H.; Bates, P.J.; Malik, T.; Li, X.-F.; Trent, J.O.; Ng, C.K. Aptamer imaging with Cu-64 labeled AS1411: Preliminary assessment in lung cancer. Nucl. Med. Biol. 2013, 41, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, O.; Yan, X.; Niu, G.; Weiss, I.D.; Ma, Y.; Szajek, L.P.; Shen, B.; Kiesewetter, D.O.; Chen, X. PET Imaging of Tenascin-C with a Radiolabeled Single-Stranded DNA Aptamer. J. Nucl. Med. 2015, 56, 616–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.; Jeong, Y.Y.; Jon, S. A Drug-Loaded Aptamer−Gold Nanoparticle Bioconjugate for Combined CT Imaging and Therapy of Prostate Cancer. ACS Nano 2010, 4, 3689–3696. [Google Scholar] [CrossRef] [PubMed]
- Li, C.-H.; Kuo, T.-R.; Su, H.-J.; Lai, W.-Y.; Yang, P.-C.; Chen, J.-S.; Wang, D.-Y.; Wu, Y.-C.; Chen, C.-C. Fluorescence-Guided Probes of Aptamer-Targeted Gold Nanoparticles with Computed Tomography Imaging Accesses for in Vivo Tumor Resection. Sci. Rep. 2015, 5, 15675. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Wang, Y. Paclitaxel-Loaded and a10-3.2 aptamer-targeted poly(lactide-co-glycolic acid) nanobubbles for ultrasound imaging and therapy of prostate cancer. Int. J. Nanomed. 2017, 12, 5313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmauch, B.; Romagnoni, A.; Pronier, E.; Saillard, C.; Maillé, P.; Calderaro, J.; Kamoun, A.; Sefta, M.; Toldo, S.; Zaslavskiy, M.; et al. A deep learning model to predict RNA-Seq expression of tumours from whole slide images. Nat. Commun. 2020, 11, 1–15. [Google Scholar] [CrossRef]
- Zarella, M.; Bowman, D.; Aeffner, F.; Farahani, N.; Xthona, A.; Absar, S.F.; Parwani, A.; Bui, M.; Hartman, D.J. A Practical Guide to Whole Slide Imaging: A White Paper from the Digital Pathology Association. Arch. Pathol. Lab. Med. 2018, 143, 222–234. [Google Scholar] [CrossRef] [Green Version]
- Srinidhi, C.L.; Ciga, O.; Martel, A.L. Deep neural network models for computational histopathology: A survey. Med. Image Anal. 2020, 67, 101813. [Google Scholar] [CrossRef]
- Simonyan, K.; Zisserman, A. Very deep convolutional networks for large-scale image recognition. arXiv 2015, arXiv:1409.1556. [Google Scholar]
- He, K.; Zhang, X.; Ren, S.; Sun, J. Deep residual learning for image recognition. In Proceedings of the IEEE Conference on Computer Vision and Pattern Recognition (CVPR), Las Vegas, NV, USA, 27–30 June 2016. [Google Scholar]
- Szegedy, C.; Liu, W.; Jia, Y.; Sermanet, P.; Reed, S.; Anguelov, D.; Erhan, D.; Vanhoucke, V.; Rabinovich, A. Going deeper with convolutions. In Proceedings of the IEEE Conference on Computer Vision and Pattern Recognition (CVPR), Boston, MA, USA, 7–12 June 2015. [Google Scholar]
- Ronneberger, O.; Fischer, P.; Brox, T. U-Net: Convolutional Networks for Biomedical Image Segmentation. In International Conference on Medical Image Computing and Computer-Assisted Intervention; Springer: Berlin/Heidelberg, Germany, 2015; pp. 234–241. [Google Scholar] [CrossRef] [Green Version]
- Long, J.; Shelhamer, E.; Darrell, T. Fully convolutional networks for semantic segmentation. In Proceedings of the IEEE Conference on Computer Vision and Pattern Recognition (CVPR), Boston, MA, USA, 7–12 June 2015. [Google Scholar]
- Girshick, R. Fast r-cnn. arXiv 2015, arXiv:1504.08083. [Google Scholar]
- Chen, L.; Papandreou, G.; Kokkinos, I.; Murphy, K.; Yuille, A.L. Deeplab: Semantic image segmentation with deep convolutional nets, atrous convolution, and fully connected crfs. IEEE Trans. Pattern Anal. Mach. Intell. 2018, 40, 834–848. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K. Overview of deep learning in medical imaging. Radiol. Phys. Technol. 2017, 10, 257–273. [Google Scholar] [CrossRef]
- Jiang, D.; Liao, J.; Duan, H.; Wu, Q.; Owen, G.; Shu, C.; Chen, L.; He, Y.; Wu, Z.; He, D.; et al. A machine learning-based prognostic predictor for stage iii colon cancer. Sci. Rep. 2020, 10, 10333. [Google Scholar] [CrossRef] [PubMed]
- Park, A.; Nam, S. Deep learning for stage prediction in neuroblastoma using gene expression data. Genom. Inform. 2019, 17, e30. [Google Scholar] [CrossRef]
- Maggio, V.; Chierici, M.; Jurman, G.; Furlanello, C. A multiobjective deep learning approach for predictive classification in neuroblastoma. arXiv 2018, arXiv:1711.08198. [Google Scholar]
- Tranchevent, L.-C.; Azuaje, F.; Rajapakse, J.C. A deep neural network approach to predicting clinical outcomes of neuroblastoma patients. BMC Med. Genom. 2019, 12, 178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, H.; Momeni, A.; Cedoz, P.-L.; Vogel, H.; Gevaert, O. Whole slide images reflect DNA methylation patterns of human tumors. NPJ Genom. Med. 2020, 5, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, F.; Oh, T.W.; Vergara-Niedermayr, C.; Kurc, T.; Saltz, J. Managing and querying whole slide images. Proc. SPIE Int. Soc. Opt. Eng. 2012, 8319, 83190. [Google Scholar]
- Kurc, T.; Qi, X.; Wang, D.; Wang, F.; Teodoro, G.; Cooper, L.; Nalisnik, M.; Yang, L.; Saltz, J.; Foran, D.J. Scalable analysis of Big pathology image data cohorts using efficient methods and high-performance computing strategies. BMC Bioinform. 2015, 16, 399. [Google Scholar] [CrossRef] [Green Version]
- Kong, J.; Cooper, L.A.D.; Wang, F.; Gao, J.; Teodoro, G.; Scarpace, L.; Mikkelsen, T.; Schniederjan, M.J.; Moreno, C.S.; Saltz, J.H.; et al. Machine-Based Morphologic Analysis of Glioblastoma Using Whole-Slide Pathology Images Uncovers Clinically Relevant Molecular Correlates. PLoS ONE 2013, 8, e81049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; Huang, Y.; Han, L.; Ozolek, J.A.; Rohde, G.K. Hierarchical feature extraction for nuclear morphometry-based cancer diagnosis. In Deep Learning and Data Labeling for Medical Applications; Carneiro, G., Mateus, D., Peter, L., Bradley, A., Tavares, J.M.R.S., Belagiannis, V., Papa, J.P., Nascimento, J.C., Loog, M., Lu, Z., et al., Eds.; Springer International Publishing: Berlin/Heidelberg, Germany, 2016; pp. 219–227. [Google Scholar]
- Chen, C.; Wang, W.; Ozolek, J.A.; Rohde, G.K. A flexible and robust approach for segmenting cell nuclei from 2d microscopy images using supervised learning and template matching. Cytom. Part A 2013, 83, 495–507. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sabbih, G.O.; Danquah, M.K. Neuroblastoma GD2 Expression and Computational Analysis of Aptamer-Based Bioaffinity Targeting. Int. J. Mol. Sci. 2021, 22, 9101. https://doi.org/10.3390/ijms22169101
Sabbih GO, Danquah MK. Neuroblastoma GD2 Expression and Computational Analysis of Aptamer-Based Bioaffinity Targeting. International Journal of Molecular Sciences. 2021; 22(16):9101. https://doi.org/10.3390/ijms22169101
Chicago/Turabian StyleSabbih, Godfred O., and Michael K. Danquah. 2021. "Neuroblastoma GD2 Expression and Computational Analysis of Aptamer-Based Bioaffinity Targeting" International Journal of Molecular Sciences 22, no. 16: 9101. https://doi.org/10.3390/ijms22169101
APA StyleSabbih, G. O., & Danquah, M. K. (2021). Neuroblastoma GD2 Expression and Computational Analysis of Aptamer-Based Bioaffinity Targeting. International Journal of Molecular Sciences, 22(16), 9101. https://doi.org/10.3390/ijms22169101