
Prostaglandin E2 Receptor 4 (EP4) Affects Trophoblast Functions via Activating the cAMP-PKA-pCREB Signaling Pathway at the Maternal-Fetal Interface in Unexplained Recurrent Miscarriage

 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Results
2.1. Demographic and Clinical Presentation of the Patients
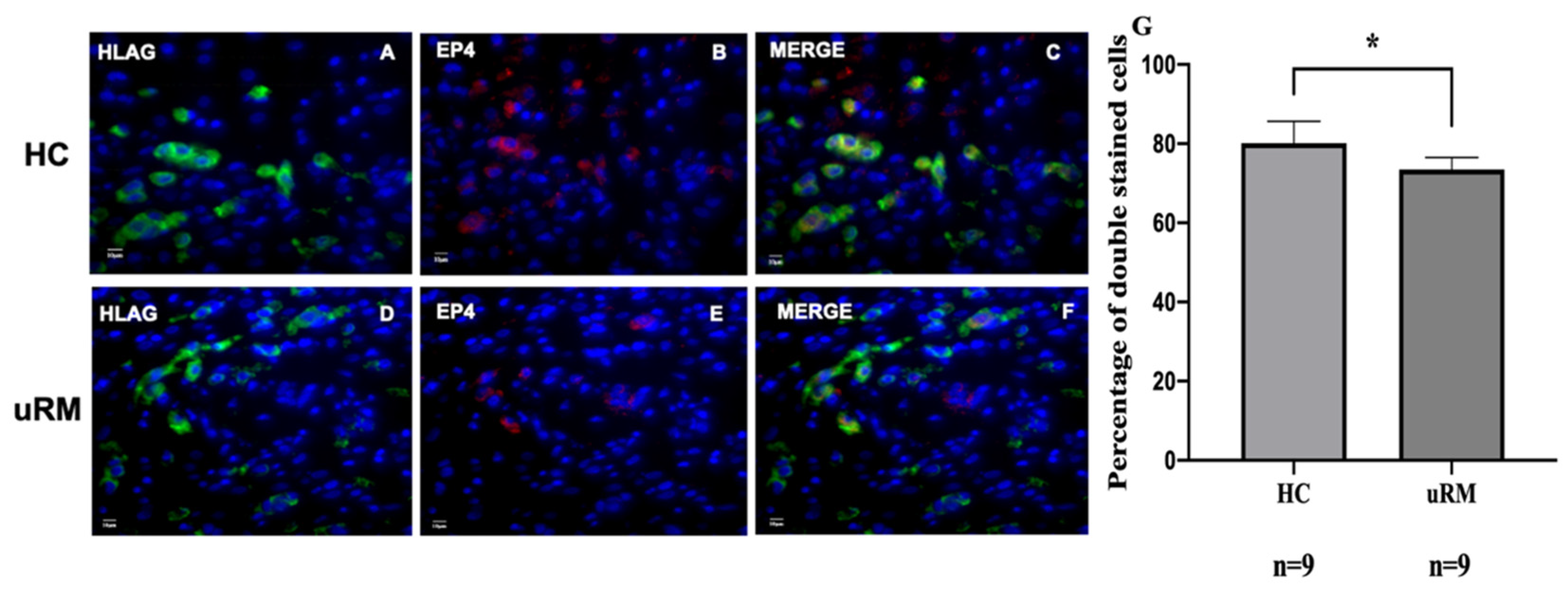
2.2. EP4 Is Downregulated in the Syncytium and EVTs in First Trimester Human Placentas with uRM
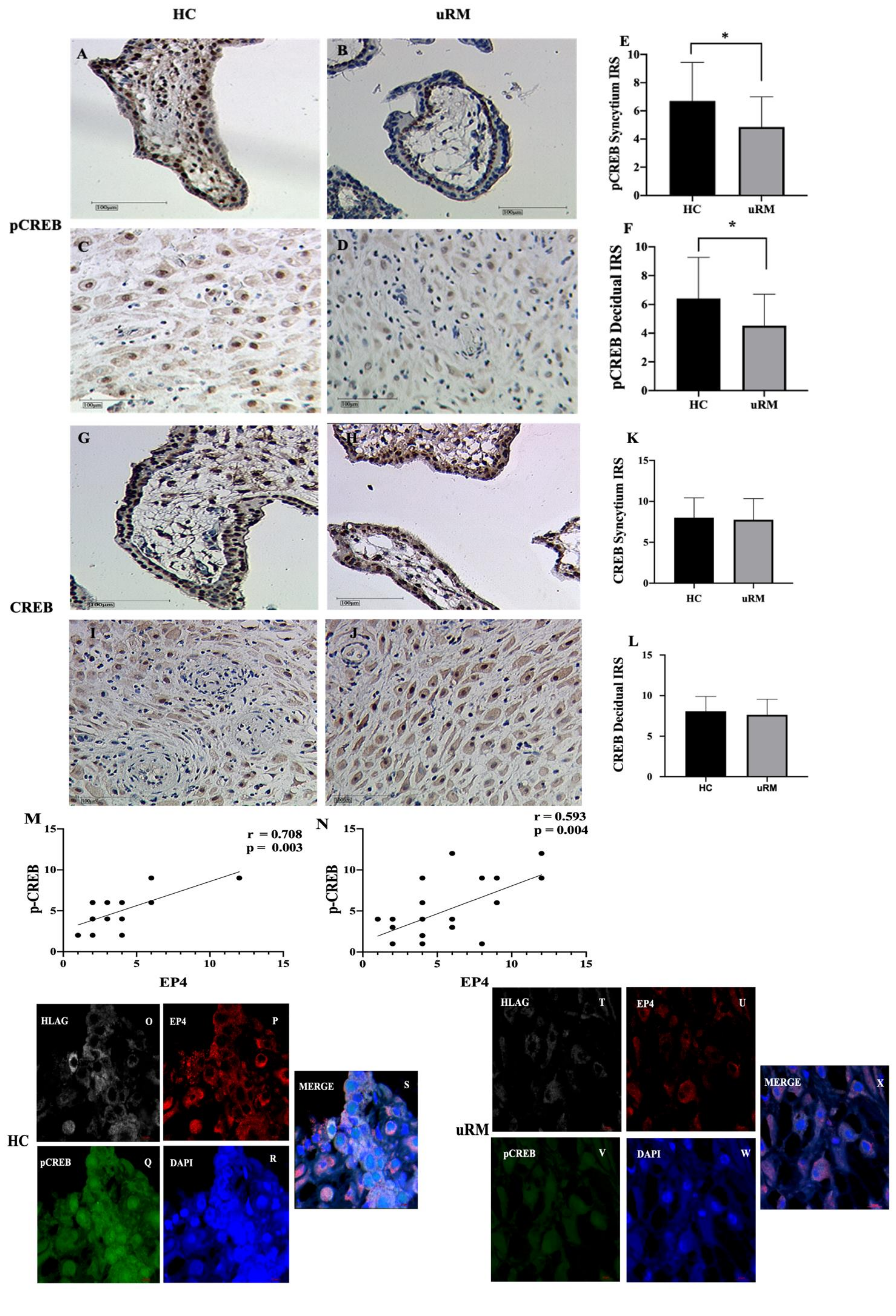
2.3. Phosphorylation CREB (pCREB) Is Involved in the Pathogenesis of uRM
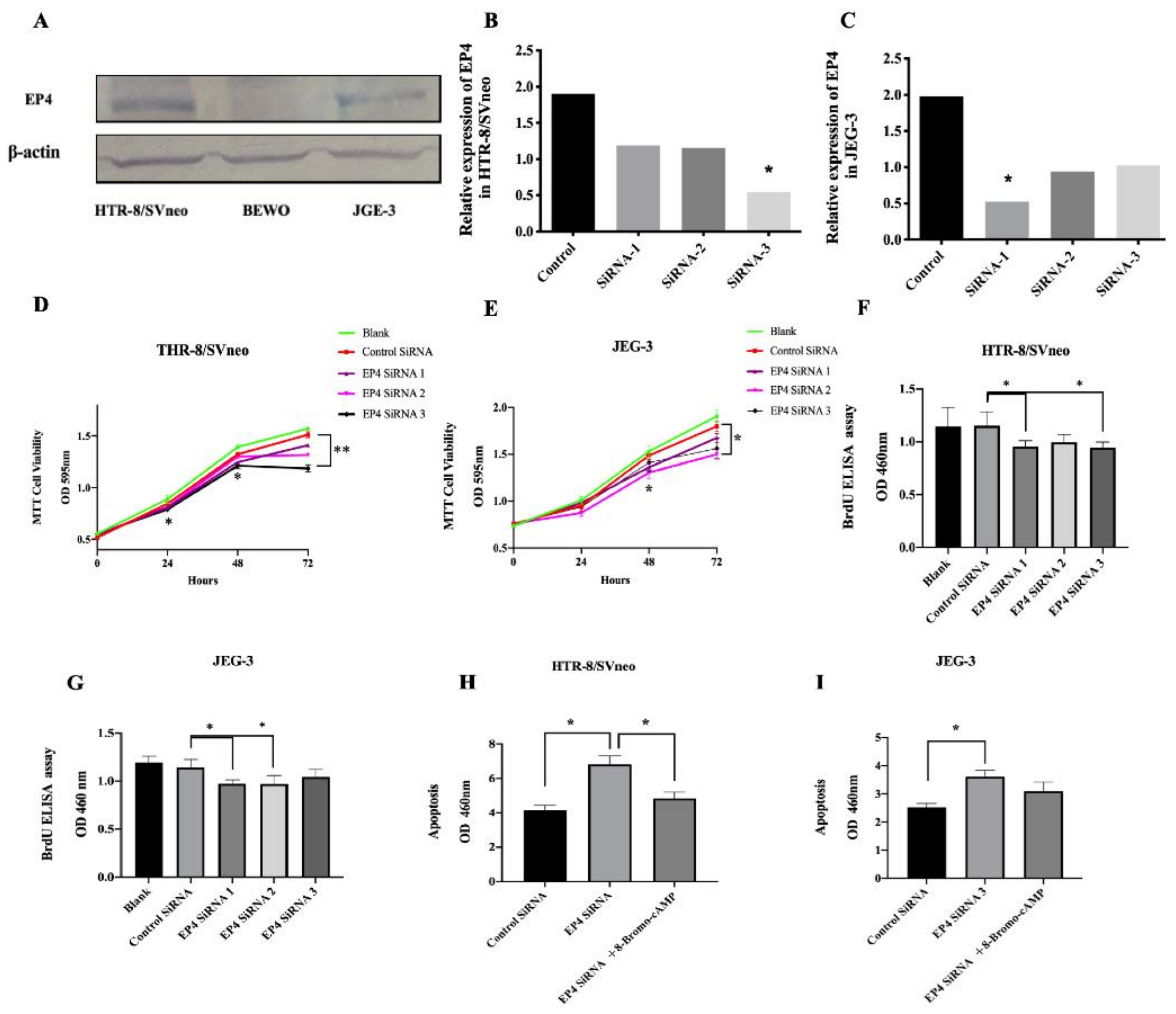
2.4. EP4 Downregulation Inhibits Proliferation and Induces Cell Apoptosis
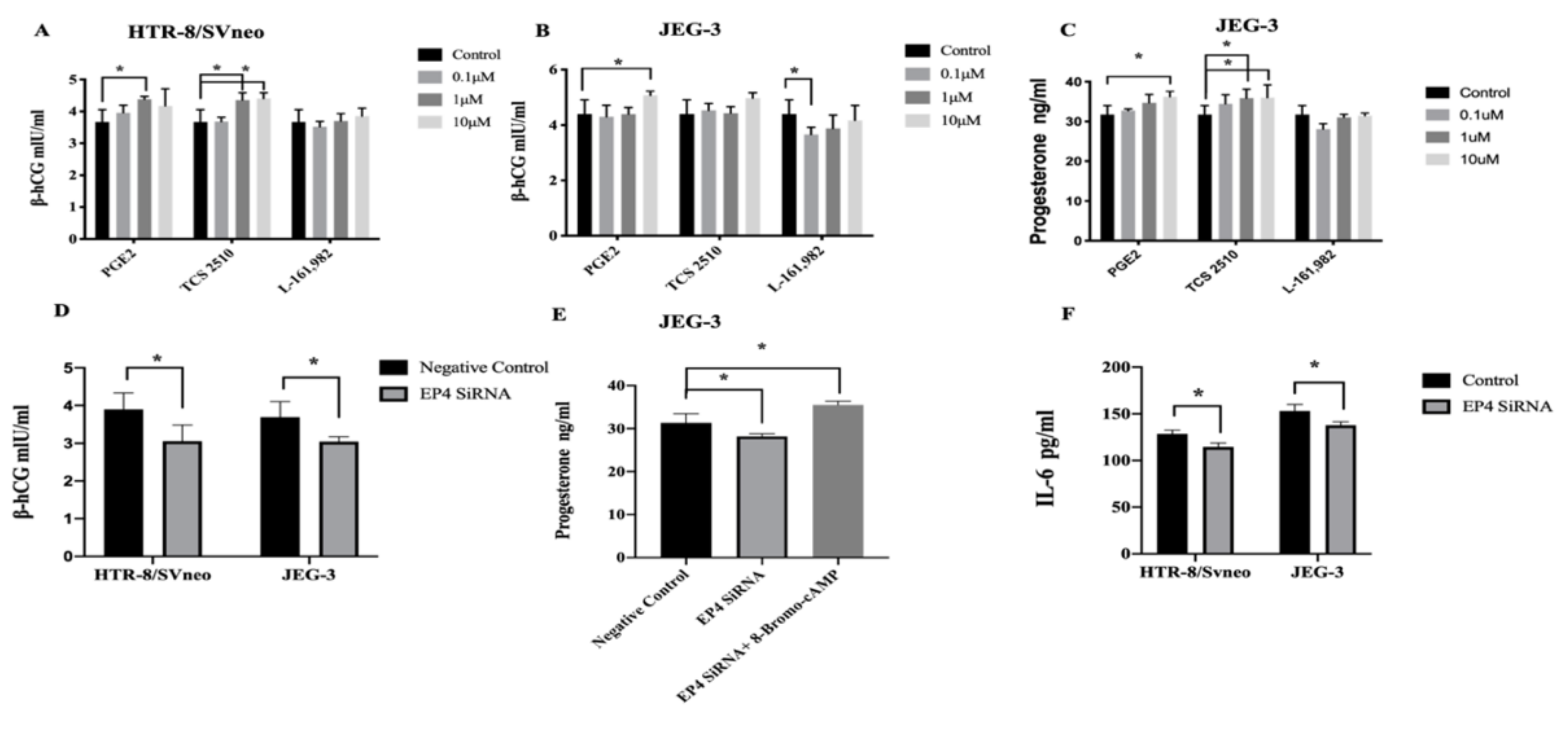
2.5. EP4 Effects Secretion of Hormones and Cytokines
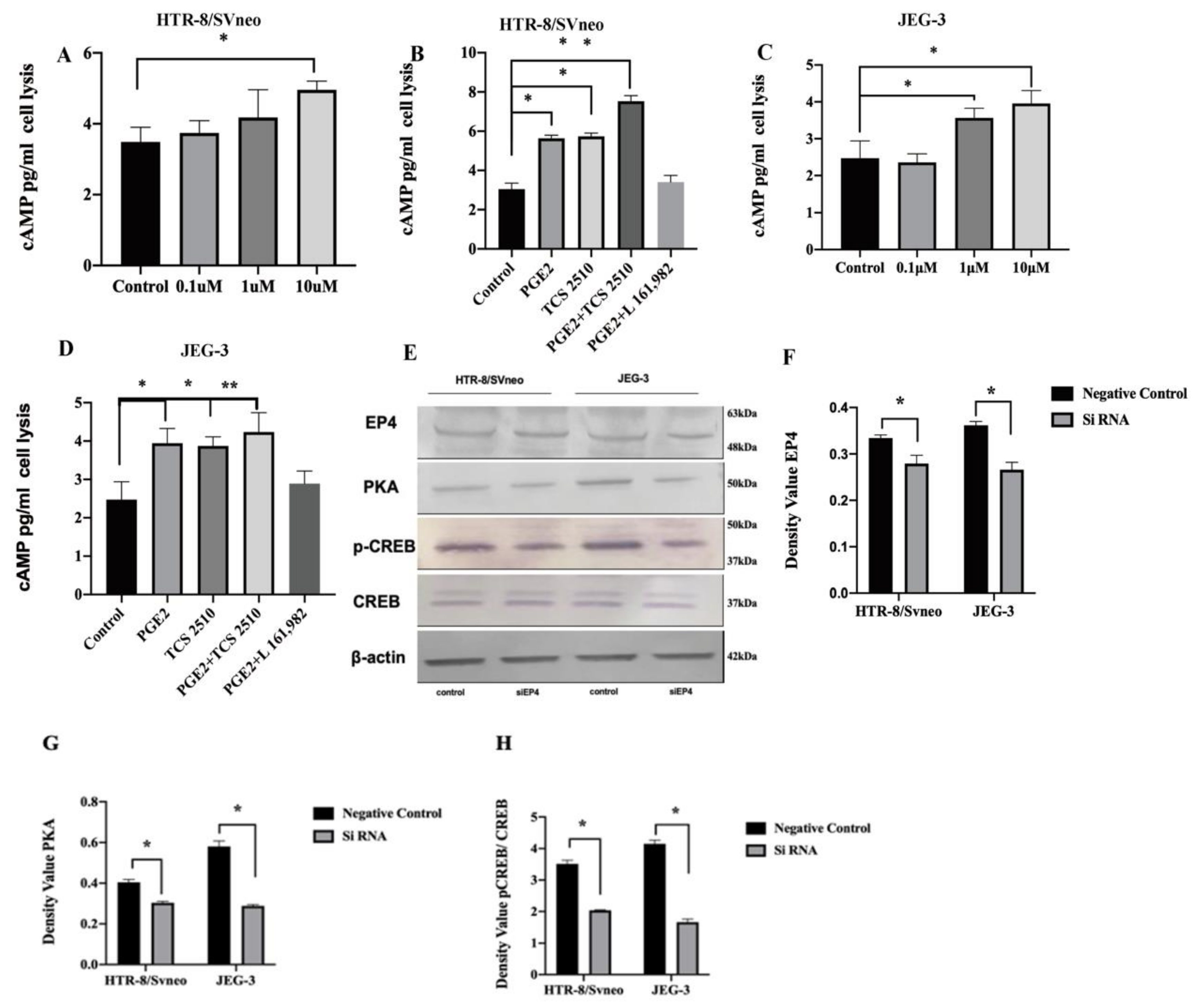
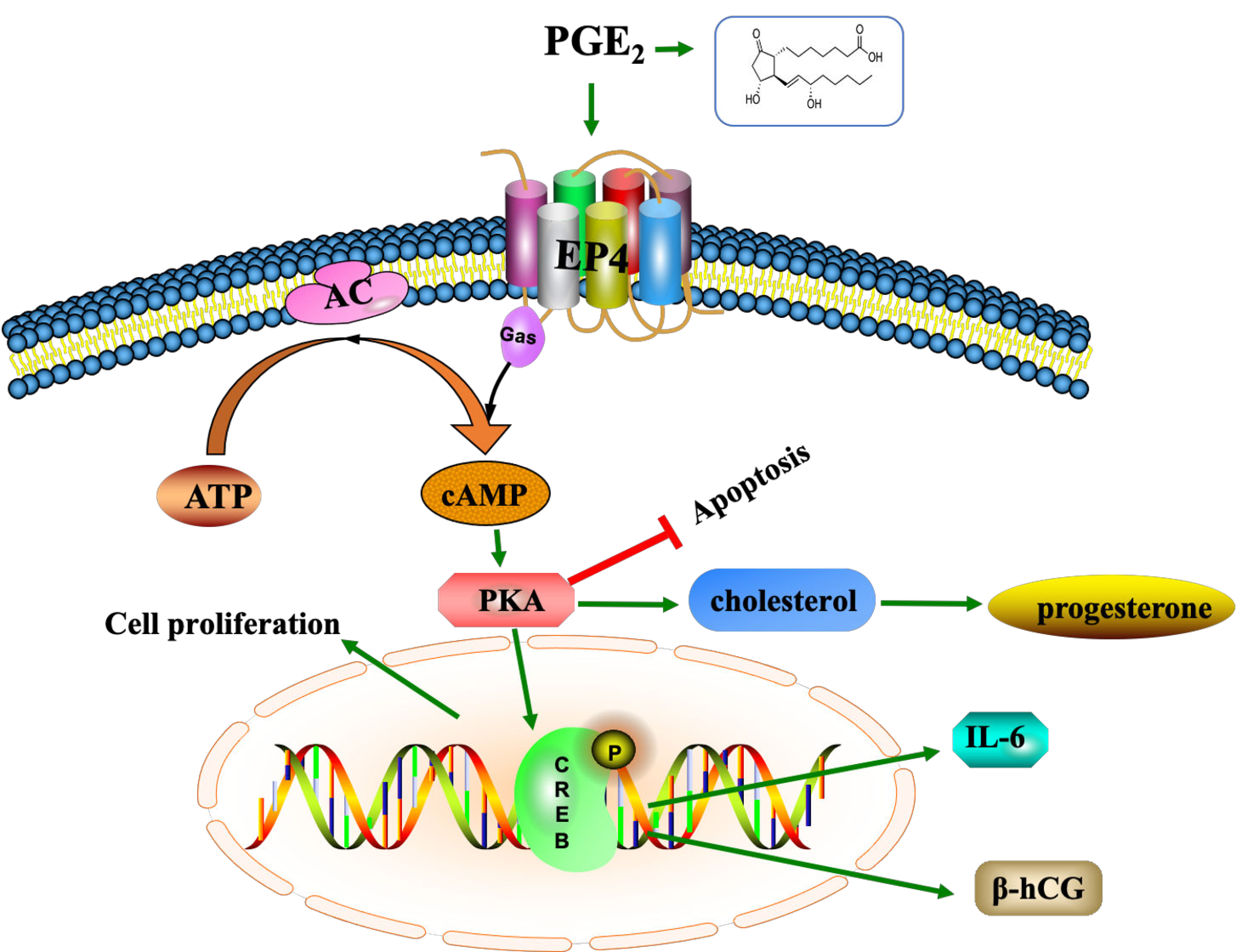
2.6. EP4 Modulates the Axis of cAMP-PKA-pCREB
3. Discussion
4. Materials and Methods
4.1. Ethics Approval and Sample Tissues
4.2. Immunohistochemistry
4.3. Immunofluorescence Staining
4.4. Cell Culture
4.5. Treatment with EP4 siRNAs
4.6. Real-Time Quantitative Extraction
4.7. Western Blot Analysis
4.8. Cell Viability Assays and Cell Proliferation
4.9. ELISA of cAMP Assay
4.10. Cell Death Detection ELISA
4.11. β-hCG and Progesterone Measurement
4.12. Detection of Cytokines by ELISA
4.13. Statistic Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| EP4 | prostaglandin E2 receptor 4 |
| EVTs | extravillous trophoblast cells |
| uRM | unexplained recurrent marriage |
| cAMP | cyclic adenosine monophosphate |
| PKA | cAMP-dependent protein kinase A |
| HCG | human chorionic gonadotropin |
| IL | interleukin |
| pCREB | phosphorylating CREB |
| ESHRE | European Society of Human Reproduction and Embryology |
| APS | antiphospholipid syndrome |
| PGE2 | Prostaglandin E2 |
| EP | G-protein-coupled EP receptors |
| HLA-G | human leukocyte antigen G |
| OD | optical density |
| PPAR | peroxisome proliferator-activated receptors |
| IHC | immunohistochemical |
| ABC | Avidin-Biotin Complex |
| IRS | immunoreactive score |
| RT-PCR | real-time polymerase chain reaction |
| iEVT | invasive extravillous trophoblast |
| EPAC | exchange protein directly activated by cAMP |
| PKA | cAMP-dependent protein kinase A |
| CREB | cAMP response element–binding protein |
| AMPK | AMP-activated protein kinase |
| LH | luteinizing hormone |
| GCM-1 | chorion-specific transcription factor-1 |
References
- Bender Atik, R.; Christiansen, O.B.; Elson, J.; Kolte, A.M.; Lewis, S.; Middeldorp, S.; Nelen, W.; Peramo, B.; Quenby, S.; Vermeulen, N.; et al. ESHRE guideline: Recurrent pregnancy loss. Hum. Reprod. Open 2018, 2018, hoy004. [Google Scholar]
- Red-Horse, K.; Zhou, Y.; Genbacev, O.; Prakobphol, A.; Foulk, R.; McMaster, M.; Fisher, S.J. Trophoblast differentiation during embryo implantation and formation of the maternal-fetal interface. J. Clin. Investig. 2004, 114, 744–754. [Google Scholar] [CrossRef]
- Burton, G.J.; Jauniaux, E.; Charnock-Jones, D.S. The influence of the intrauterine environment on human placental development. Int. J. Dev. Biol. 2010, 54, 303–312. [Google Scholar] [CrossRef] [Green Version]
- Okae, H.; Toh, H.; Sato, T.; Hiura, H.; Takahashi, S.; Shirane, K.; Kabayama, Y.; Suyama, M.; Sasaki, H.; Arima, T. Derivation of Human Trophoblast Stem Cells. Cell Stem Cell 2018, 22, 50–63.e6. [Google Scholar] [CrossRef] [Green Version]
- Gamage, T.K.; Chamley, L.W.; James, J.L. Stem cell insights into human trophoblast lineage differentiation. Hum. Reprod. Update 2016, 23, 77–103. [Google Scholar] [CrossRef] [PubMed]
- Abbas, Y.; Turco, M.Y.; Burton, G.J.; Moffett, A. Investigation of human trophoblast invasion in vitro. Hum. Reprod. Update 2020, 26, 501–513. [Google Scholar] [CrossRef] [PubMed]
- Sokolowska, M.; Chen, L.Y.; Liu, Y.; Martinez-Anton, A.; Qi, H.Y.; Logun, C.; Alsaaty, S.; Park, Y.H.; Kastner, D.L.; Chae, J.J.; et al. Prostaglandin E2 Inhibits NLRP3 Inflammasome Activation through EP4 Receptor and Intracellular Cyclic AMP in Human Macrophages. J. Immunol. 2015, 194, 5472–5487. [Google Scholar] [CrossRef] [Green Version]
- Bogacka, I.; Bogacki, M.; Gaglewska, M.; Kurzynska, A.; Wasielak, M. In vitro effect of peroxisome proliferator activated receptor (PPAR) ligands on prostaglandin E2 synthesis and secretion by porcine endometrium during the estrous cycle and early pregnancy. J. Physiol. Pharmacol. 2013, 64, 47–54. [Google Scholar]
- Niringiyumukiza, J.D.; Cai, H.; Xiang, W. Prostaglandin E2 involvement in mammalian female fertility: Ovulation, fertilization, embryo development and early implantation. Reprod. Biol. Endocrinol. 2018, 16, 43. [Google Scholar] [CrossRef] [PubMed]
- Vilella, F.; Ramirez, L.; Berlanga, O.; Martínez, S.; Alamá, P.; Meseguer, M.; Pellicer, A.; Simón, C. PGE2 and PGF2α concentrations in human endometrial fluid as biomarkers for embryonic implantation. J. Clin. Endocrinol. Metab. 2013, 98, 4123–4132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pakrasi, P.L.; Jain, A.K. Cyclooxygenase-2 derived PGE2 and PGI2 play an important role via EP2 and PPARdelta receptors in early steps of oil induced decidualization in mice. Placenta 2008, 29, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Waclawik, A.; Jabbour, H.N.; Blitek, A.; Ziecik, A.J. Estradiol-17beta, prostaglandin E2 (PGE2), and the PGE2 receptor are involved in PGE2 positive feedback loop in the porcine endometrium. Endocrinology 2009, 150, 3823–3832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, T.A.; Keelan, J.A.; Mitchell, M.D. Critical paracrine interactions between TNF-alpha and IL-10 regulate lipopolysaccharide-stimulated human choriodecidual cytokine and prostaglandin E2 production. J. Immunol. 2003, 170, 158–166. [Google Scholar] [CrossRef] [Green Version]
- Ye, Y.; Vattai, A.; Ditsch, N.; Kuhn, C.; Rahmeh, M.; Mahner, S.; Ripphahn, M.; Immler, R.; Sperandio, M.; Jeschke, U.; et al. Prostaglandin E(2) receptor 3 signaling is induced in placentas with unexplained recurrent pregnancy losses. Endocr. Connect. 2018, 7, 749–761. [Google Scholar] [CrossRef]
- Peng, L.; Ye, Y.; Mullikin, H.; Lin, L.; Kuhn, C.; Rahmeh, M.; Mahner, S.; Jeschke, U.; von Schönfeldt, V. Expression of trophoblast derived prostaglandin E2 receptor 2 (EP2) is reduced in patients with recurrent miscarriage and EP2 regulates cell proliferation and expression of inflammatory cytokines. J. Reprod. Immunol. 2020, 142, 103210. [Google Scholar] [CrossRef]
- Kolben, T.M.; Rogatsch, E.; Vattai, A.; Hester, A.; Kuhn, C.; Schmoeckel, E.; Mahner, S.; Jeschke, U.; Kolben, T. PPARγ Expression Is Diminished in Macrophages of Recurrent Miscarriage Placentas. Int. J. Mol. Sci. 2018, 19, 1872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, F.; Rouault, C.; Guesnon, M.; Zhu, W.; Clément, K.; Degrelle, S.A.; Fournier, T. Comparative Study of PPARγ Targets in Human Extravillous and Villous Cytotrophoblasts. PPAR Res. 2020, 2020, 9210748. [Google Scholar] [CrossRef] [PubMed]
- Darashchonak, N.; Koepsell, B.; Bogdanova, N.; von Versen-Höynck, F. Adenosine A2B receptors induce proliferation, invasion and activation of cAMP response element binding protein (CREB) in trophoblast cells. BMC Pregnancy Childbirth 2014, 14, 2. [Google Scholar] [CrossRef] [Green Version]
- Nicola, C.; Timoshenko, A.V.; Dixon, S.J.; Lala, P.K.; Chakraborty, C. EP1 receptor-mediated migration of the first trimester human extravillous trophoblast: The role of intracellular calcium and calpain. J. Clin. Endocrinol. Metab. 2005, 90, 4736–4746. [Google Scholar] [CrossRef] [Green Version]
- Dhanasekaran, N.; Tsim, S.T.; Dermott, J.M.; Onesime, D. Regulation of cell proliferation by G proteins. Oncogene 1998, 17, 1383–1394. [Google Scholar] [CrossRef] [Green Version]
- Sirotkin, A.V.; Ben, O.A.; Tandlmajerová, A.; Lauková, M.; Vaší Ek, D.; Laurin Ik, J.; Kornhauser, J.; Alwasel, S.; Harrath, A.H. cAMP response element-binding protein 1 controls porcine ovarian cell proliferation, apoptosis, and FSH and insulin-like growth factor 1 response. Reprod. Fertil. Dev. 2018, 30, 1145–1153. [Google Scholar] [CrossRef]
- Hsu, H.H.; Lin, Y.M.; Shen, C.Y.; Shibu, M.A.; Li, S.Y.; Chang, S.H.; Lin, C.C.; Chen, R.J.; Viswanadha, V.P.; Shih, H.N.; et al. Prostaglandin E2-Induced COX-2 Expressions via EP2 and EP4 Signaling Pathways in Human LoVo Colon Cancer Cells. Int. J. Mol. Sci. 2017, 18, 1132. [Google Scholar] [CrossRef] [PubMed]
- Parida, S.; Pal, I.; Parekh, A.; Thakur, B.; Bharti, R.; Das, S.; Mandal, M. GW627368X inhibits proliferation and induces apoptosis in cervical cancer by interfering with EP4/EGFR interactive signaling. Cell Death Dis. 2016, 7, e2154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, J.; Zeng, B.; Zhong, M.; Li, H.; Xu, L.; Shu, J.; Wang, Y.; Yang, F.; Zhong, C.; Ye, X.; et al. Scutellarin inhibits caspase-11 activation and pyroptosis in macrophages via regulating PKA signaling. Acta Pharm. Sin. B 2021, 11, 112–126. [Google Scholar] [CrossRef] [PubMed]
- Tao, J.L.; Zhang, X.; Zhou, J.Q.; Li, C.Y.; Yang, M.H.; Liu, Z.J.; Zhang, L.L.; Deng, S.L.; Zhang, L.; Shen, M.; et al. Melatonin Alleviates Hypoxia-Induced Apoptosis of Granulosa Cells by Reducing ROS and Activating MTNR1B-PKA-Caspase8/9 Pathway. Antioxidants 2021, 10, 184. [Google Scholar] [CrossRef]
- Hoshino, T.; Tsutsumi, S.; Tomisato, W.; Hwang, H.J.; Tsuchiya, T.; Mizushima, T. Prostaglandin E2 protects gastric mucosal cells from apoptosis via EP2 and EP4 receptor activation. J. Biol. Chem. 2003, 278, 12752–12758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fournier, T. Human chorionic gonadotropin: Different glycoforms and biological activity depending on its source of production. Ann. Endocrinol. 2016, 77, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Vondra, S.; Kunihs, V.; Eberhart, T.; Eigner, K.; Bauer, R.; Haslinger, P.; Haider, S.; Windsperger, K.; Klambauer, G.; Schütz, B.; et al. Metabolism of cholesterol and progesterone is differentially regulated in primary trophoblastic subtypes and might be disturbed in recurrent miscarriages. J. Lipid Res. 2019, 60, 1922–1934. [Google Scholar] [CrossRef]
- Kusama, K.; Bai, R.; Imakawa, K. Regulation of human trophoblast cell syncytialization by transcription factors STAT5B and NR4A3. J. Cell Biochem. 2018, 119, 4918–4927. [Google Scholar] [CrossRef]
- Sullivan, M.H. Endocrine cell lines from the placenta. Mol. Cell Endocrinol. 2004, 228, 103–119. [Google Scholar] [CrossRef]
- Tuckey, R.C. Progesterone synthesis by the human placenta. Placenta 2005, 26, 273–281. [Google Scholar] [CrossRef]
- Mayr, B.; Montminy, M. Transcriptional regulation by the phosphorylation-dependent factor CREB. Nat. Rev. Mol. Cell Biol. 2001, 2, 599–609. [Google Scholar] [CrossRef]
- Knöfler, M.; Saleh, L.; Strohmer, H.; Husslein, P.; Wolschek, M.F. Cyclic AMP- and differentiation-dependent regulation of the proximal alphaHCG gene promoter in term villous trophoblasts. Mol. Hum. Reprod. 1999, 5, 573–580. [Google Scholar] [CrossRef] [PubMed]
- Gawriluk, T.R.; Ko, C.; Hong, X.; Christenson, L.K.; Rucker, E.B., 3rd. Beclin-1 deficiency in the murine ovary results in the reduction of progesterone production to promote preterm labor. Proc. Natl. Acad. Sci. USA 2014, 111, E4194–E4203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waclawik, A.; Kaczmarek, M.M.; Blitek, A.; Kaczynski, P.; Ziecik, A.J. Embryo-maternal dialogue during pregnancy establishment and implantation in the pig. Mol. Reprod. Dev. 2017, 84, 842–855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziecik, A.J.; Waclawik, A.; Kaczmarek, M.M.; Blitek, A.; Jalali, B.M.; Andronowska, A. Mechanisms for the establishment of pregnancy in the pig. Reprod. Domest. Anim. 2011, 46, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Waclawik, A.; Kaczynski, P.; Jabbour, H.N. Autocrine and paracrine mechanisms of prostaglandin E₂ action on trophoblast/conceptus cells through the prostaglandin E₂ receptor (PTGER2) during implantation. Endocrinology 2013, 154, 3864–3876. [Google Scholar] [CrossRef] [Green Version]
- Kowalewski, M.P.; Fox, B.; Gram, A.; Boos, A.; Reichler, I. Prostaglandin E2 functions as a luteotrophic factor in the dog. Reproduction 2013, 145, 213–226. [Google Scholar] [CrossRef] [Green Version]
- Cole, L.A. Biological functions of hCG and hCG-related molecules. Reprod. Biol. Endocrinol. 2010, 8, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tarrade, A.; Schoonjans, K.; Guibourdenche, J.; Bidart, J.M.; Vidaud, M.; Auwerx, J.; Rochette-Egly, C.; Evain-Brion, D. PPAR gamma/RXR alpha heterodimers are involved in human CG beta synthesis and human trophoblast differentiation. Endocrinology 2001, 142, 4504–4514. [Google Scholar] [CrossRef]
- Peng, L.; Yang, H.; Ye, Y.; Ma, Z.; Kuhn, C.; Rahmeh, M.; Mahner, S.; Makrigiannakis, A.; Jeschke, U.; von Schönfeldt, V. Role of Peroxisome Proliferator-Activated Receptors (PPARs) in Trophoblast Functions. Int. J. Mol. Sci. 2021, 22, 433. [Google Scholar] [CrossRef] [PubMed]
- Mitsunari, M.; Yoshida, S.; Shoji, T.; Tsukihara, S.; Iwabe, T.; Harada, T.; Terakawa, N. Macrophage-activating lipopeptide-2 induces cyclooxygenase-2 and prostaglandin E(2) via toll-like receptor 2 in human placental trophoblast cells. J. Reprod. Immunol. 2006, 72, 46–59. [Google Scholar] [CrossRef] [PubMed]
- Loch-Caruso, R.; Korte, C.S.; Hogan, K.A.; Liao, S.; Harris, C. Tert-Butyl Hydroperoxide Stimulated Apoptosis Independent of Prostaglandin E(2) and IL-6 in the HTR-8/SVneo Human Placental Cell Line. Reprod. Sci. 2020, 27, 2104–2114. [Google Scholar] [CrossRef]
- Diehl, S.; Rincón, M. The two faces of IL-6 on Th1/Th2 differentiation. Mol. Immunol. 2002, 39, 531–536. [Google Scholar] [CrossRef]
- Saito, S. Cytokine network at the feto-maternal interface. J. Reprod. Immunol. 2000, 47, 87–103. [Google Scholar] [CrossRef]
- Von Wolff, M.; Thaler, C.J.; Strowitzki, T.; Broome, J.; Stolz, W.; Tabibzadeh, S. Regulated expression of cytokines in human endometrium throughout the menstrual cycle: Dysregulation in habitual abortion. Mol. Hum. Reprod. 2000, 6, 627–634. [Google Scholar] [CrossRef]
- Koumantaki, Y.; Matalliotakis, I.; Sifakis, S.; Kyriakou, D.; Neonaki, M.; Goymenou, A.; Koumantakis, E. Detection of interleukin-6, interleukin-8, and interleukin-11 in plasma from women with spontaneous abortion. Eur. J. Obstet. Gynecol. Reprod. Biol. 2001, 98, 66–71. [Google Scholar] [CrossRef]
- Lim, K.J.; Odukoya, O.A.; Ajjan, R.A.; Li, T.C.; Weetman, A.P.; Cooke, I.D. The role of T-helper cytokines in human reproduction. Fertil Steril 2000, 73, 136–142. [Google Scholar] [CrossRef]
- Raychaudhuri, N.; Douglas, R.S.; Smith, T.J. PGE2 induces IL-6 in orbital fibroblasts through EP2 receptors and increased gene promoter activity: Implications to thyroid-associated ophthalmopathy. PLoS ONE 2010, 5, e15296. [Google Scholar] [CrossRef] [Green Version]
- Hershko, D.D.; Robb, B.W.; Luo, G.; Hasselgren, P.O. Multiple transcription factors regulating the IL-6 gene are activated by cAMP in cultured Caco-2 cells. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2002, 283, R1140–R1148. [Google Scholar] [CrossRef] [Green Version]
- Jovanović, M.; Vićovac, L. Interleukin-6 stimulates cell migration, invasion and integrin expression in HTR-8/SVneo cell line. Placenta 2009, 30, 320–328. [Google Scholar] [CrossRef] [PubMed]
- Suman, P.; Gupta, S.K. STAT3 and ERK1/2 cross-talk in leukaemia inhibitory factor mediated trophoblastic JEG-3 cell invasion and expression of mucin 1 and Fos. Am. J. Reprod. Immunol. 2014, 72, 65–74. [Google Scholar] [CrossRef]
- Ding, J.; Yang, C.; Cheng, Y.; Wang, J.; Zhang, S.; Yan, S.; He, F.; Yin, T.; Yang, J. Trophoblast-derived IL-6 serves as an important factor for normal pregnancy by activating Stat3-mediated M2 macrophages polarization. Int. Immunopharmacol. 2021, 90, 106788. [Google Scholar] [CrossRef]
- Ye, Y.; Peng, L.; Vattai, A.; Deuster, E.; Kuhn, C.; Dannecker, C.; Mahner, S.; Jeschke, U.; von Schönfeldt, V.; Heidegger, H.H. Prostaglandin E2 receptor 3 (EP3) signaling promotes migration of cervical cancer via urokinase-type plasminogen activator receptor (uPAR). J. Cancer. Res. Clin. Oncol. 2020, 146, 2189–2203. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | Normal Pregnancy n = 19 | uRM n = 19 | p Value |
|---|---|---|---|
| maternal age (years) | 35.78 ± 5.88 (25–46) | 37.76 ± 4.88 (30–44) | 0.41 |
| gestational age (weeks) | 9.71 ± 1.88 (6–13) | 9.09 ± 2.17 (4–12) | 0.66 |
| gravidity | 3.42 ± 1.90 (1–7) | 3.11 ± 1.08 (2–5) | 0.78 |
| parity | 1.63 ± 1.12 (0–4) | 0.94 ± 0.94 (0–3) | 0.06 |
| Antibody | Species | Clone | Dilution | Company (Catalog#) | Concentration (μg/mL) |
|---|---|---|---|---|---|
| EP4 | rabbit | polyclonal | 1:100 | Abcam. ab45295 | 400 |
| CREB | rabbit | monoclonal | 1:500 | Abcam. ab32515 | 166 |
| pCREB | rabbit | monoclonal | 1:1000 | Abcam. ab32096 | 150 |
| PPAR-γ | mouse | monoclonal | 1:500 | Abnova. MAB8316 | 1000 |
| HLA-G | mouse | monoclonal | 1:200 | Novus. NBP1-43123 | 1000 |
| HLA-G biotin | mouse | monoclonal | 1:200 | LSBio. LS-C204117 | NA |
| Cy-2 | goat | polyclonal | 1:500 | Dianova. 115-225-146 | 1500 |
| Cy-3 | goat | polyclonal | 1:100 | Dianova.115-167-003 | 1000 |
| Cy-5 | Streptavidin | NA | 1:200 | Dianova.016-170-084 | 1000 |
| PKA | rabbit | monoclonal | 1:5000 | Abcam, ab32514 | 200 |
| pPKA | rabbit | polyclonal | 1:500 | Abcam, ab226754 | 300 |
| β-actin | mouse | monoclonal | 1:1000 | Sigma, A5441 | 200 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peng, L.; Chelariu-Raicu, A.; Ye, Y.; Ma, Z.; Yang, H.; Ishikawa-Ankerhold, H.; Rahmeh, M.; Mahner, S.; Jeschke, U.; von Schönfeldt, V. Prostaglandin E2 Receptor 4 (EP4) Affects Trophoblast Functions via Activating the cAMP-PKA-pCREB Signaling Pathway at the Maternal-Fetal Interface in Unexplained Recurrent Miscarriage. Int. J. Mol. Sci. 2021, 22, 9134. https://doi.org/10.3390/ijms22179134
Peng L, Chelariu-Raicu A, Ye Y, Ma Z, Yang H, Ishikawa-Ankerhold H, Rahmeh M, Mahner S, Jeschke U, von Schönfeldt V. Prostaglandin E2 Receptor 4 (EP4) Affects Trophoblast Functions via Activating the cAMP-PKA-pCREB Signaling Pathway at the Maternal-Fetal Interface in Unexplained Recurrent Miscarriage. International Journal of Molecular Sciences. 2021; 22(17):9134. https://doi.org/10.3390/ijms22179134
Chicago/Turabian StylePeng, Lin, Anca Chelariu-Raicu, Yao Ye, Zhi Ma, Huixia Yang, Hellen Ishikawa-Ankerhold, Martina Rahmeh, Sven Mahner, Udo Jeschke, and Viktoria von Schönfeldt. 2021. "Prostaglandin E2 Receptor 4 (EP4) Affects Trophoblast Functions via Activating the cAMP-PKA-pCREB Signaling Pathway at the Maternal-Fetal Interface in Unexplained Recurrent Miscarriage" International Journal of Molecular Sciences 22, no. 17: 9134. https://doi.org/10.3390/ijms22179134
APA StylePeng, L., Chelariu-Raicu, A., Ye, Y., Ma, Z., Yang, H., Ishikawa-Ankerhold, H., Rahmeh, M., Mahner, S., Jeschke, U., & von Schönfeldt, V. (2021). Prostaglandin E2 Receptor 4 (EP4) Affects Trophoblast Functions via Activating the cAMP-PKA-pCREB Signaling Pathway at the Maternal-Fetal Interface in Unexplained Recurrent Miscarriage. International Journal of Molecular Sciences, 22(17), 9134. https://doi.org/10.3390/ijms22179134