
Implementation of a Non-Thermal Atmospheric Pressure Plasma for Eradication of Plant Pathogens from a Surface of Economically Important Seeds

 , , ,
, , ,  ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Results and Discussion
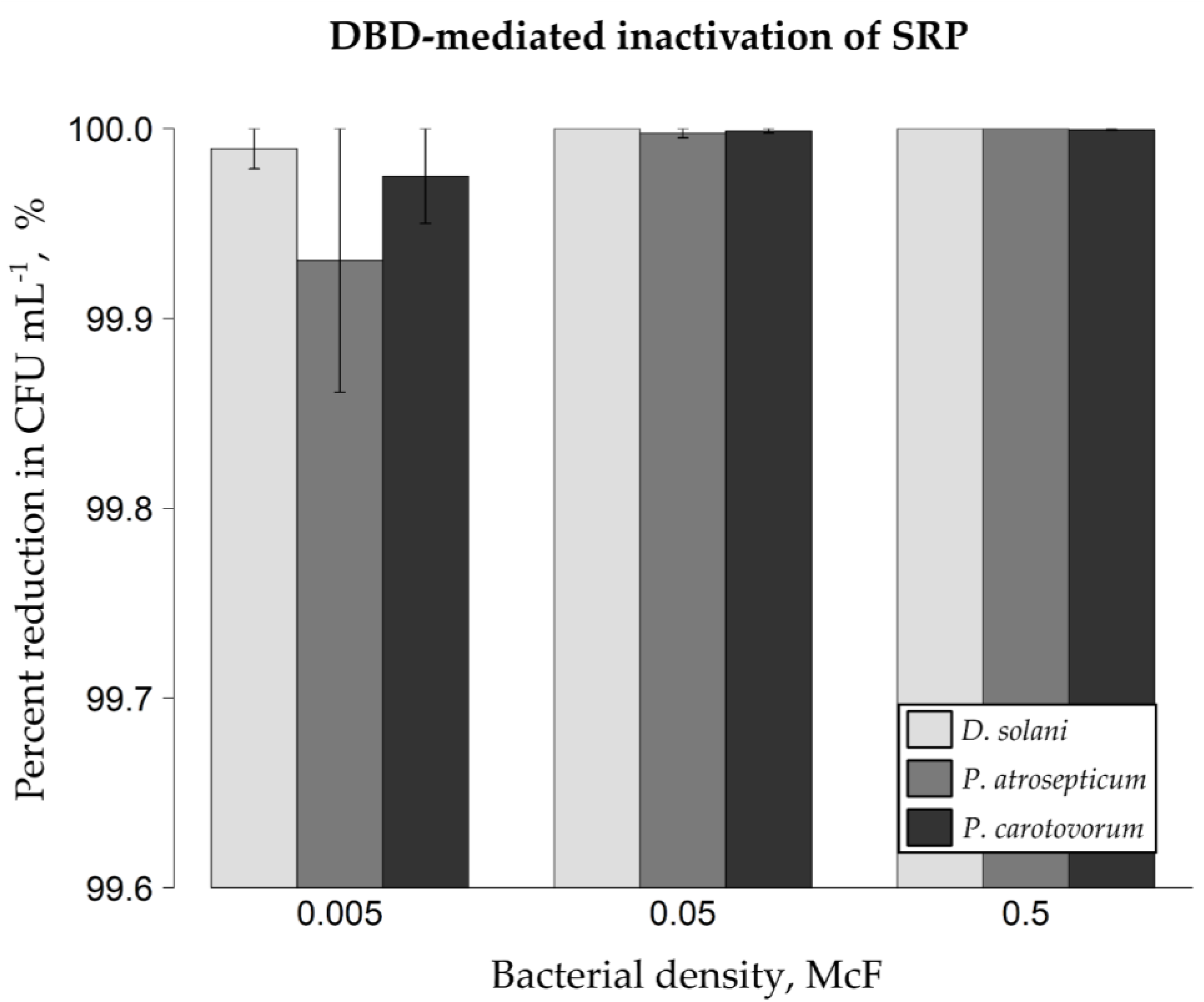
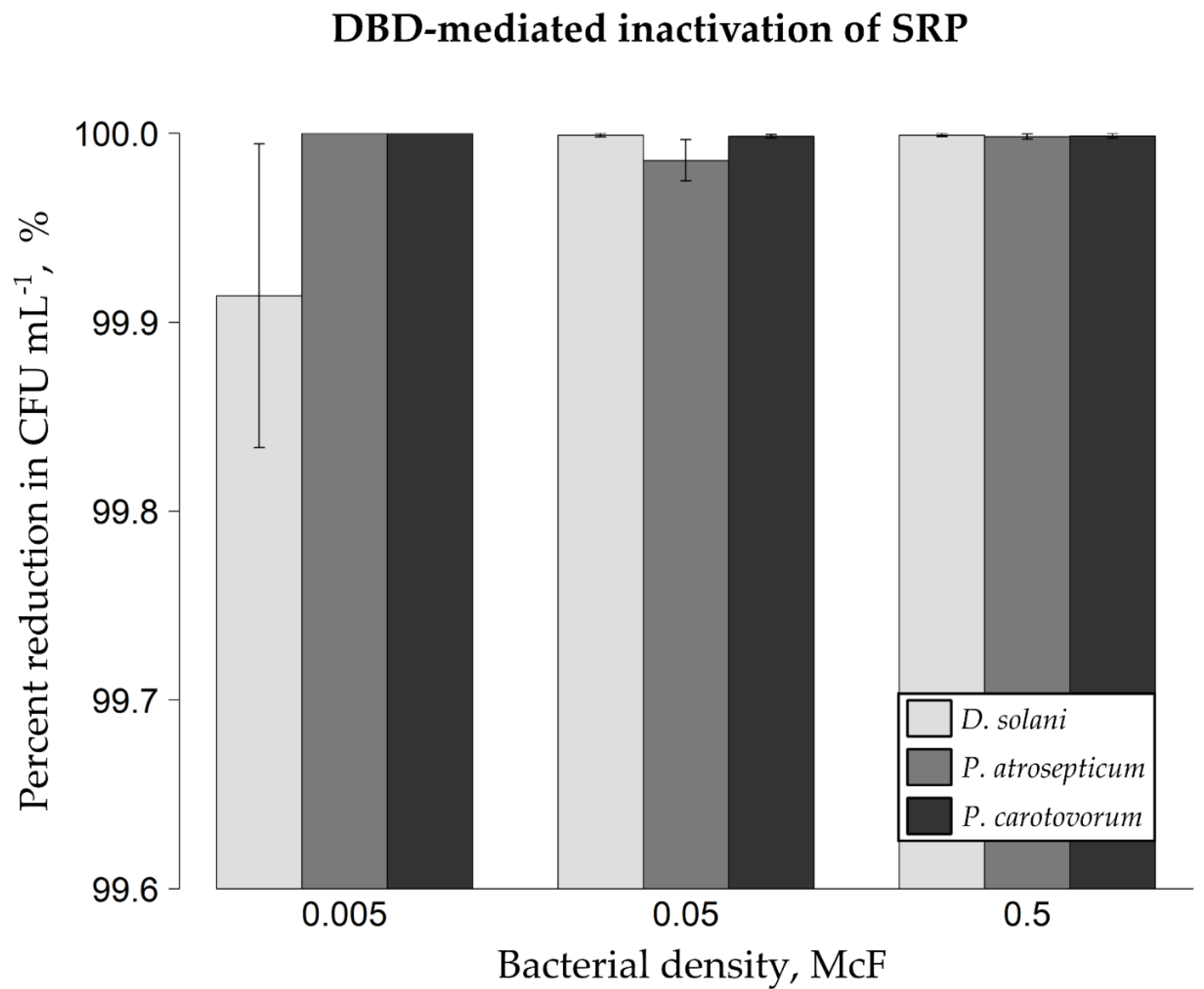
2.1. DBD-Triggered Inactivation of Bacterial Phytopathogens from Glass Spheres and Mung Bean Seeds
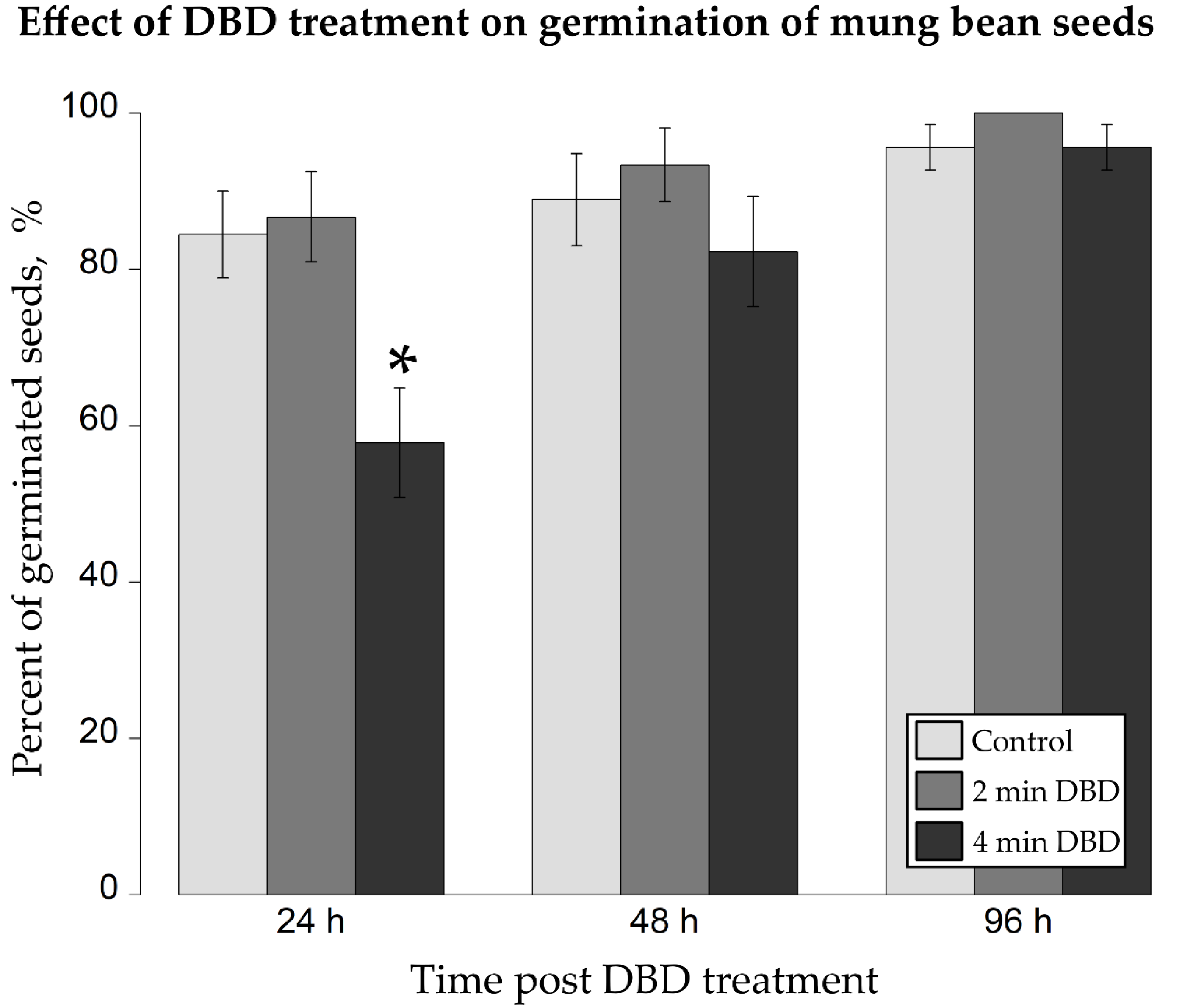
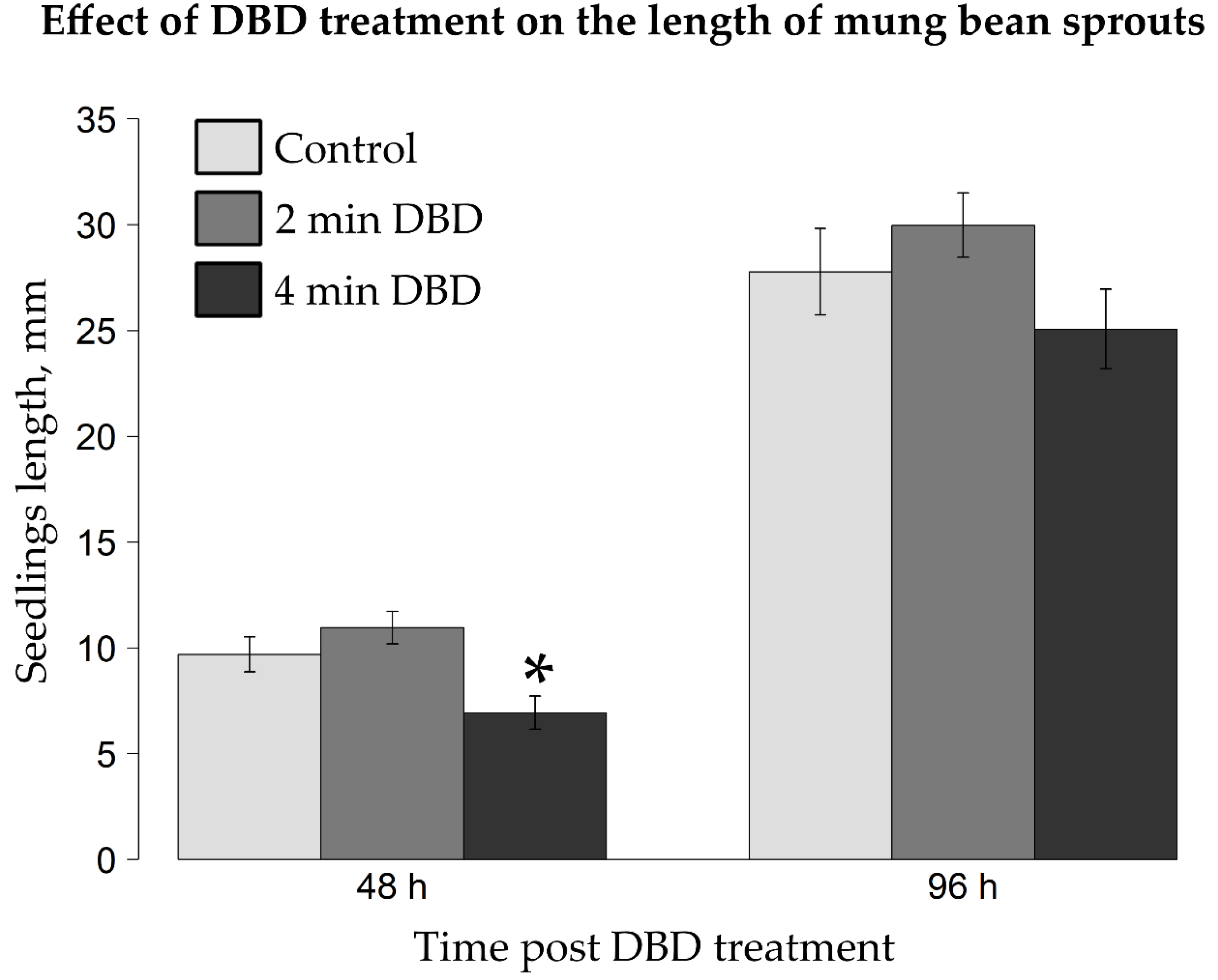
2.2. Impact of DBD Treatment on Germination Rate and Early Growth of Mung Bean Seedlings
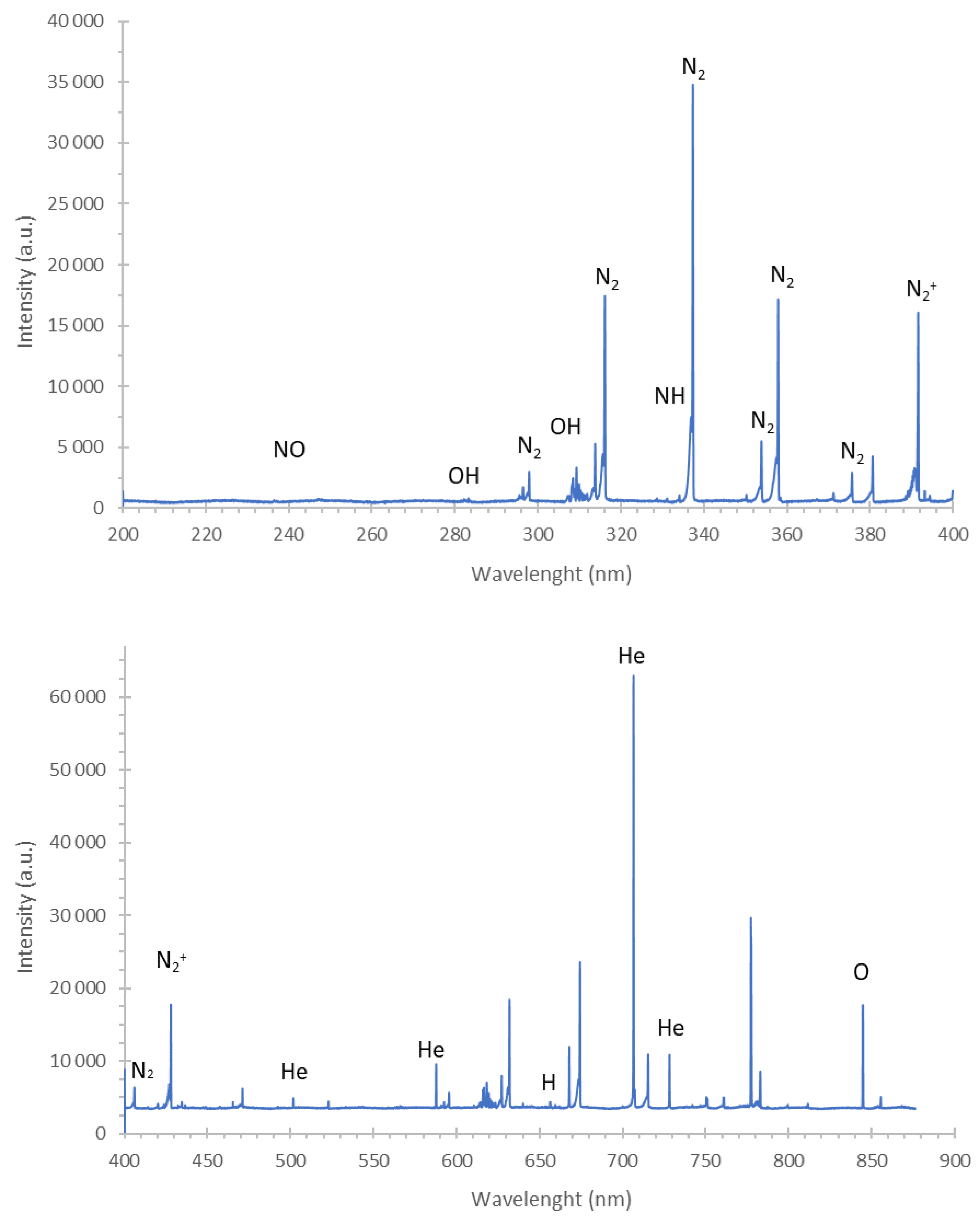
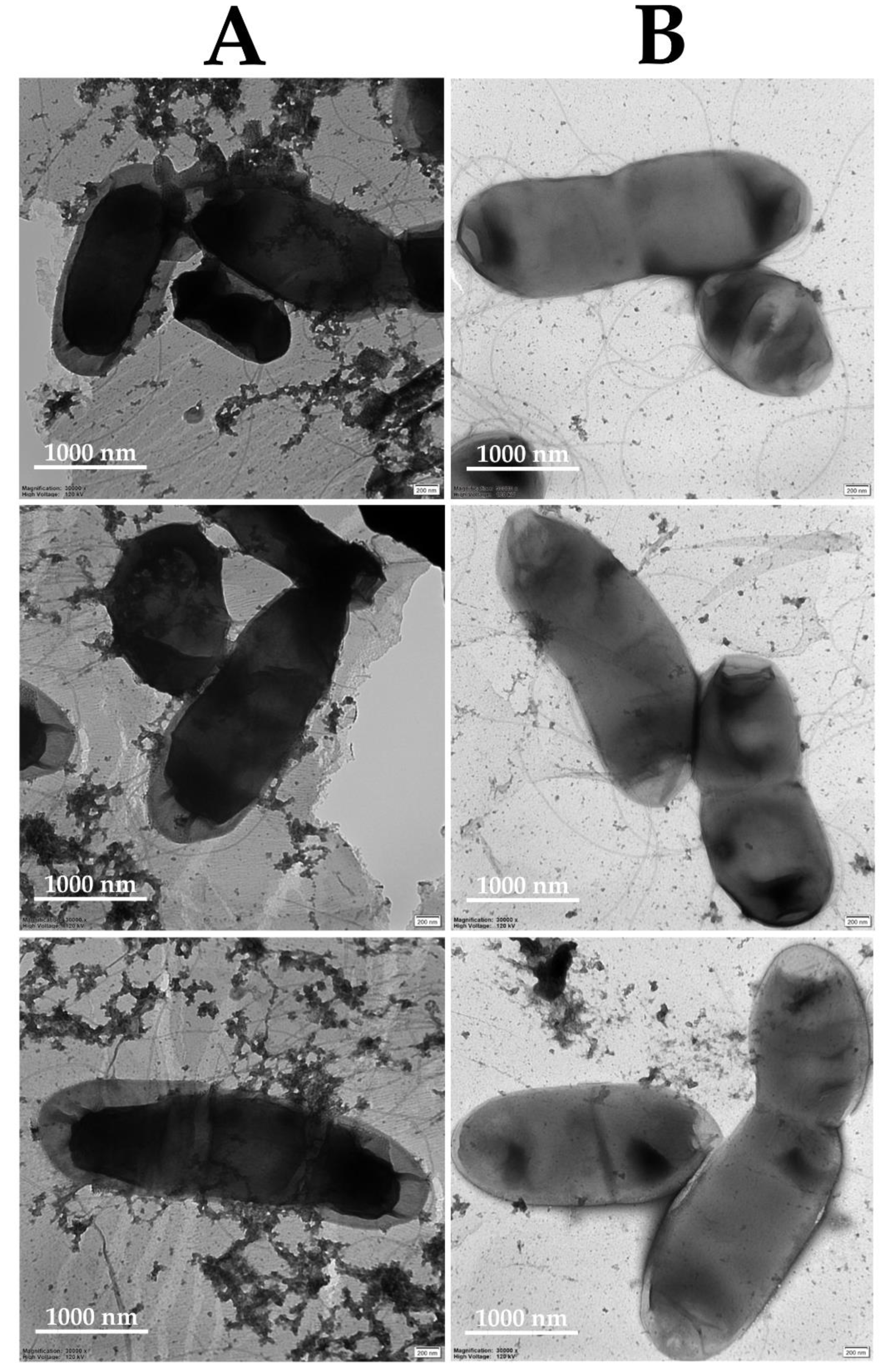
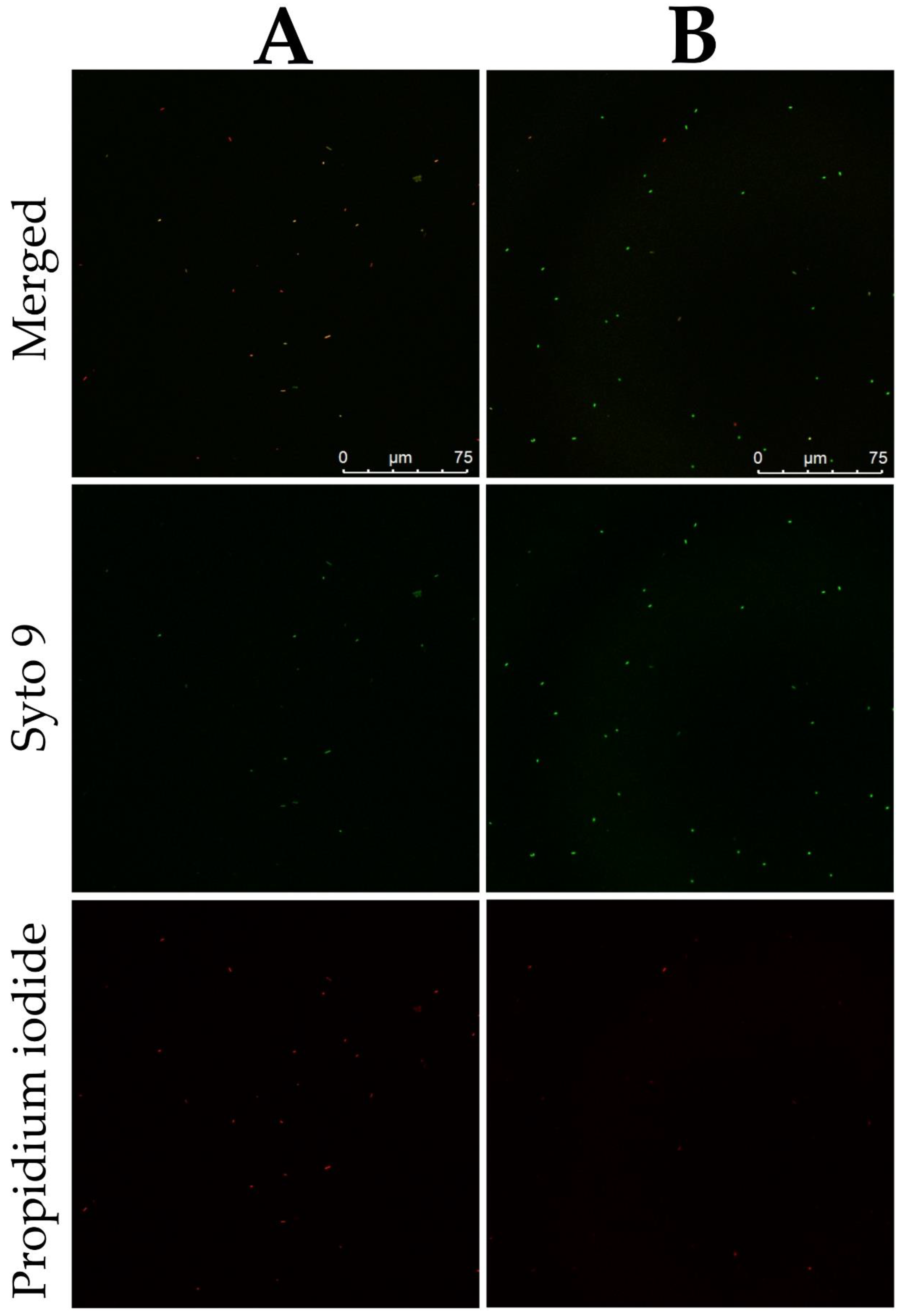
2.3. Mechanism of the Antibacterial Action of DBD
3. Materials and Methods
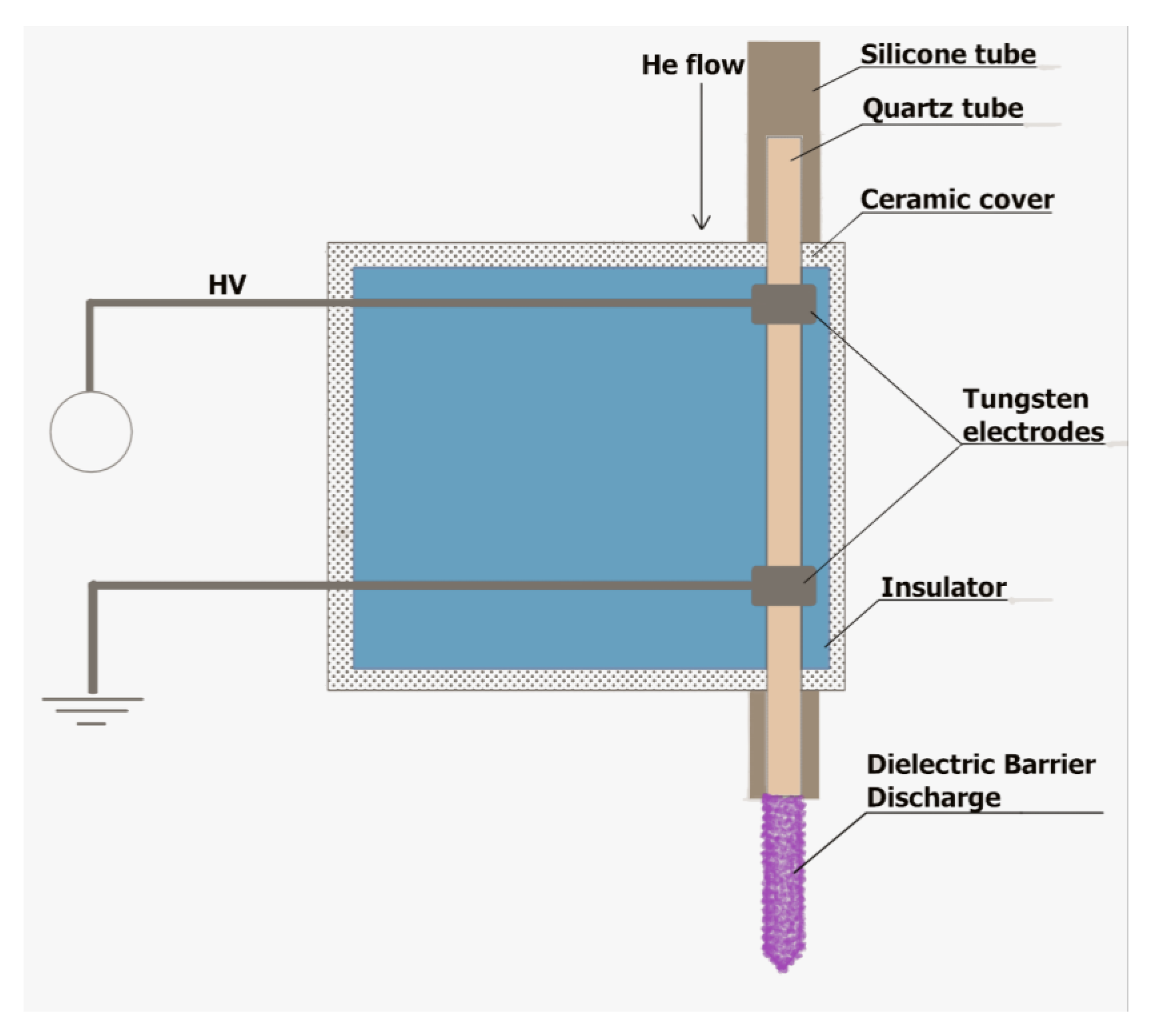
3.1. A DBD-Based Portable Plasma Pencil
3.2. Bacterial Suspensions
3.3. Antibacterial Properties of DBD towards Plant Pathogens Inoculated on the Surface of Glass Spheres
3.4. Antibacterial Properties of DBD towards Plant Pathogens Inoculated on the Surface of Mung Bean Seeds
3.5. Impact of the DBD Treatment on the Mung Bean Seed Germination Rate and the Early Growth of Seedlings
3.6. Mechanism of the Antibacterial Action of DBD
3.7. Data Visualization and Statistical Analysis
4. Conclusions
5. Patents
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ANOVA | Analysis of variance |
| CFUs | Colony-forming units |
| CLSM | Confocal laser scanning microscopy |
| DBD | Dielectric barrier discharge |
| DPPH | 2,2-diphenyl-1-picrylhydrazyl |
| FVB | Full vertical binding |
| McF | McFarland |
| OES | Optical Emission Spectrometry |
| NTAPP | Non-thermal atmospheric pressure plasma |
| PALs | Plasma-activated liquids |
| PCR | Polymerase chain reaction |
| RF | Radio-frequency |
| RNS | Reactive nitrogen species |
| RONS | Reactive oxygen and nitrogen species |
| ROS | Reactive oxygen species |
| SEM | Scanning electron microscopy |
| SMD | Surface microdischarge |
| SRP | Soft rot Pectobacteriaceae |
| TEM | Transmission electron microscopy |
| TSA | Trypticase soy agar |
| TSB | Trypticase soy broth |
References
- Buttimer, C.; McAuliffe, O.; Ross, R.P.; Hill, C.; O’Mahony, J.; Coffey, A. Bacteriophages and bacterial plant diseases. Front. Microbiol. 2017, 8, 34. [Google Scholar] [CrossRef] [Green Version]
- Borkar, S.G.; Yumlembam, R.A. Bacterial Diseases of Crop Plants; Taylor and Francis Group: London, UK, 2017. [Google Scholar]
- Lindsey, A.P.J.; Murugan, S.; Renitta, R.E. Microbial disease management in agriculture: Current status and future prospects. Biocatal. Agric. Biotechnol. 2020, 23, 101468. [Google Scholar] [CrossRef]
- Czajkowski, R.; Pérombelon, M.C.M.; van Veen, J.A.; van der Wolf, J.M. Control of blackleg and tuber soft rot of potato caused by Pectobacterium and Dickeya species: A review. Plant Pathol. 2011, 60, 999–1013. [Google Scholar] [CrossRef]
- Sundin, G.W.; Castiblanco, L.F.; Yuan, X.; Zeng, Q.; Yang, C.-H. Bacterial disease management: Challenges, experience, innovation and future prospects. Mol. Plant Pathol. 2016, 17, 1506–1518. [Google Scholar] [CrossRef]
- Van der Wolf, J.M.; De Boer, S.H.; Czajkowski, R.; Cahill, G.; Van Gijsegem, F.; Davey, T.; Dupuis, B.; Ellicott, J.; Jafra, S.; Kooman, M.; et al. Management of diseases caused by Pectobacterium and Dickeya species. In Plant Diseases Caused by Dickeya and Pectobacterium Species; Van Gijsegem, F., van der Wolf, J.M., Toth, I.K., Eds.; Springer: Cham, The Switzerland, 2021; pp. 175–214. ISBN 978-3-030-61458-4. [Google Scholar]
- Austin, S.; Lojkowska, E.; Ehlenfeldt, K.; Kelman, A.; Helgeson, J.P. Fertile interspecific somatic hybrids of Solanum: A novel source of resistance to Erwinia soft rot. Phytopathology 1988, 78, 1216–1220. [Google Scholar] [CrossRef]
- Patyka, V.; Buletsa, N.; Pasichnyk, L.; Zhitkevich, N.; Kalinichenko, A.; Gnatiuk, T.; Butsenko, L. Specifics of pesticides effects on the phytopathogenic bacteria. Ecol. Chem. Eng. Soc. 2016, 23, 311–331. [Google Scholar] [CrossRef] [Green Version]
- Rossmann, S.; Dees, M.W.; Perminow, J.; Meadow, R.; Brurberg, M.B. Soft rot Enterobacteriaceae are carried by a large range of insect species in potato fields. Appl. Environ. Microbiol. 2018, 84, e00281-18. [Google Scholar] [CrossRef] [Green Version]
- Nykyri, J.; Fang, X.; Dorati, F.; Bakr, R.; Pasanen, M.; Niemi, O.; Palva, E.T.; Jackson, R.W.; Pirhonen, M. Evidence that nematodes may vector the soft rot-causing enterobacterial phytopathogens. Plant Pathol. 2014, 63, 747–757. [Google Scholar] [CrossRef]
- Gitaitis, R.; Walcott, R. The epidemiology and management of seedborne bacterial diseases. Annu. Rev. Phytopathol. 2008, 45, 371–397. [Google Scholar] [CrossRef]
- Ding, H.; Fu, T.-J.; Smith, M.A. Microbial Contamination in Sprouts: How Effective Is Seed Disinfection Treatment? J. Food Sci. 2013, 78, R495–R501. [Google Scholar] [CrossRef] [Green Version]
- Nishioka, T.; Takai, Y.; Mishima, T.; Kawaradani, M.; Tanimoto, H.; Okada, K.; Misawa, T.; Kusakari, S. Low-Pressure Plasma Application for the Inactivation of the Seed-borne Pathogen Xanthomonas campestris. Biocontrol Sci. 2016, 21, 37–43. [Google Scholar] [CrossRef] [Green Version]
- Bruggeman, P.J.; Kushner, M.J.; Locke, B.R.; Gardeniers, J.G.E.; Graham, W.G.; Graves, D.B.; Hofman-Caris, R.C.H.M.; Maric, D.; Reid, J.P.; Ceriani, E.; et al. Plasma-liquid interactions: A review and roadmap. Plasma Sources Sci. Technol. 2016, 25, 053002. [Google Scholar] [CrossRef]
- Adhikari, B.; Pangomm, K.; Veerana, M.; Mitra, S.; Park, G. Plant Disease Control by Non-Thermal Atmospheric-Pressure Plasma. Front. Plant Sci. 2020, 11, 77. [Google Scholar] [CrossRef] [PubMed]
- Ito, M.; Oh, J.-S.; Ohta, T.; Shiratani, M.; Hori, M. Current status and future prospects of agricultural applications using atmospheric-pressure plasma technologies. Plasma Process. Polym. 2018, 15, 1700073. [Google Scholar] [CrossRef]
- Selcuk, M.; Oksuz, L.; Basaran, P. Decontamination of grains and legumes infected with Aspergillus spp. and Penicillum spp. by cold plasma treatment. Bioresour. Technol. 2008, 99, 5104–5109. [Google Scholar] [CrossRef]
- Nishioka, T.; Takai, Y.; Kawaradani, M.; Okada, K.; Tanimoto, H.; Misawa, T.; Kusakari, S. Seed Disinfection Effect of Atmospheric Pressure Plasma and Low Pressure Plasma on Rhizoctonia solani. Biocontrol Sci. 2014, 19, 99–102. [Google Scholar] [CrossRef] [Green Version]
- Ono, R.; Uchida, S.; Hayashi, N.; Kosaka, R.; Soeda, Y. Inactivation of bacteria on plant seed surface by low-pressure RF plasma using a vibrating stirring device. Vacuum 2017, 136, 214–220. [Google Scholar] [CrossRef]
- Ochi, A.; Konishi, H.; Ando, S.; Sato, K.; Yokoyama, K.; Tsushima, S.; Yoshida, S.; Morikawa, T.; Kaneko, T.; Takahashi, H. Management of bakanae and bacterial seedling blight diseases in nurseries by irradiating rice seeds with atmospheric plasma. Plant Pathol. 2017, 66, 67–76. [Google Scholar] [CrossRef] [Green Version]
- Jiang, J.; Lu, Y.; Li, J.; Li, L.; He, X.; Shao, H.; Dong, Y. Effect of Seed Treatment by Cold Plasma on the Resistance of Tomato to Ralstonia solanacearum (Bacterial Wilt). PLoS ONE 2014, 9, e97753. [Google Scholar] [CrossRef] [Green Version]
- Adeolu, M.; Alnajar, S.; Naushad, S.; Gupta, R.S. Genome-based phylogeny and taxonomy of the ‘Enterobacteriales’: Proposal for Enterobacterales ord. nov. divided into the families Enterobacteriaceae, Erwiniaceae fam. nov., Pectobacteriaceae fam. nov., Yersiniaceae fam. nov., Hafniaceae fam. nov., Morganellaceae fam. nov., and Budviciaceae fam. nov. Int. J. Syst. Evol. Microbiol. 2016, 66, 5575–5599. [Google Scholar] [CrossRef]
- Motyka, A.; Dzimitrowicz, A.; Jamroz, P.; Lojkowska, E.; Sledz, W.; Pohl, P. Rapid eradication of bacterial phytopathogens by atmospheric pressure glow discharge generated in contact with a flowing liquid cathode. Biotechnol. Bioeng. 2018, 115, 1581–1593. [Google Scholar] [CrossRef]
- Moreau, M.; Feuilloley, M.G.J.; Veron, W.; Meylheuc, T.; Chevalier, S.; Brisset, J.L.; Orange, N. Gliding arc discharge in the potato pathogen Erwinia carotovora subsp. atroseptica: Mechanism of lethal action and effect on membrane-associated molecules. Appl. Environ. Microbiol. 2007, 73, 5904–5910. [Google Scholar] [CrossRef] [Green Version]
- Schnabel, U.; Niquet, R.; Schlüter, O.; Gniffke, H.; Ehlbeck, J. Decontamination and Sensory Properties of Microbiologically Contaminated Fresh Fruits and Vegetables by Microwave Plasma Processed Air (PPA). J. Food Process. Preserv. 2015, 39, 653–662. [Google Scholar] [CrossRef]
- Dzimitrowicz, A.; Jamroz, P.; Pohl, P.; Babinska, W.; Terefinko, D.; Sledz, W.; Motyka-Pomagruk, A. Multivariate Optimization of the FLC-dc-APGD-Based Reaction-Discharge System for Continuous Production of a Plasma-Activated Liquid of Defined Physicochemical and Anti-Phytopathogenic Properties. Int. J. Mol. Sci. 2021, 22, 4813. [Google Scholar] [CrossRef]
- Schnabel, U.; Niquet, R.; Krohmann, U.; Winter, J.; Schlüter, O.; Weltmann, K.-D.; Ehlbeck, J. Decontamination of Microbiologically Contaminated Specimen by Direct and Indirect Plasma Treatment. Plasma Process. Polym. 2012, 9, 569–575. [Google Scholar] [CrossRef]
- Yu, H.; Perni, S.; Shi, J.; Wang, D.; Kong, M.; Shama, G. Effects of cell surface loading and phase of growth in cold atmospheric gas plasma inactivation of Escherichia coli K12. J. Appl. Microbiol. 2006, 101, 1323–1330. [Google Scholar] [CrossRef] [PubMed]
- Dzimitrowicz, A.; Bielawska-Pohl, A.; Jamroz, P.; Dora, J.; Krawczenko, A.; Busco, G.; Grillon, C.; Kieda, C.; Klimczak, A.; Terefinko, D. Activation of the Normal Human Skin Cells by a Portable Dielectric Barrier Discharge-Based Reaction-Discharge System of a Defined Gas Temperature. Plasma Chem. Plasma Process. 2020, 40, 79–97. [Google Scholar] [CrossRef] [Green Version]
- Mitra, A.; Li, Y.F.; Klämpfl, T.G.; Shimizu, T.; Jeon, J.; Morfill, G.E.; Zimmermann, J.L. Inactivation of Surface-Borne Microorganisms and Increased Germination of Seed Specimen by Cold Atmospheric Plasma. Food Bioprocess Technol. 2014, 7, 645–653. [Google Scholar] [CrossRef] [Green Version]
- Puligundla, P.; Kim, J.W.; Mok, C. Effect of corona discharge plasma jet treatment on decontamination and sprouting of rapeseed (Brassica napus L.) seeds. Food Control 2017, 71, 376–382. [Google Scholar] [CrossRef]
- Li, L.; Jiang, J.; Li, J.; Shen, M.; He, X.; Shao, H.; Dong, Y. Effects of cold plasma treatment on seed germination and seedling growth of soybean. Sci. Rep. 2014, 4, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Hoon Park, J.; Kumar, N.; Hoon Park, D.; Yusupov, M.; Neyts, E.C.; Verlackt, C.C.W.; Bogaerts, A.; Kang, M.H.; Uhm, H.S.; Choi, E.H.; et al. A comparative study for the inactivation of multidrug resistance bacteria using dielectric barrier discharge and nano-second pulsed plasma. Sci. Rep. 2015, 5, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Olatunde, O.O.; Benjakul, S.; Vongkamjan, K. Dielectric barrier discharge cold atmospheric plasma: Bacterial inactivation mechanism. J. Food Saf. 2019, 39, e12705. [Google Scholar] [CrossRef]
- Miao, H.; Yun, G. The sterilization of Escherichia coli by dielectric-barrier discharge plasma at atmospheric pressure. Appl. Surf. Sci. 2011, 257, 7065–7070. [Google Scholar] [CrossRef]
- Lee, M.J.; Kwon, J.S.; Jiang, H.B.; Choi, E.H.; Park, G.; Kim, K.M. The antibacterial effect of non-thermal atmospheric pressure plasma treatment of titanium surfaces according to the bacterial wall structure. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Xia, C.; Guo, Y.; Yang, C.; Cheng, C.; Zhao, J.; Yang, X.; Cao, Z. Bactericidal efficacy of cold atmospheric plasma treatment against multidrug-resistant Pseudomonas aeruginosa. Future Microbiol. 2020, 15, 115–125. [Google Scholar] [CrossRef]
- Feng, Q.L.; Wu, J.; Chen, G.Q.; Cui, F.Z.; Kim, T.N.; Kim, J.O. A mechanistic study of the antibacterial effect of silver ions on Escherichia coli and Staphylococcus aureus. J. Biomed. Mater. Res. 2000, 52, 662–668. [Google Scholar] [CrossRef]
- Shaikh, S.; Nazam, N.; Rizvi, S.M.D.; Ahmad, K.; Baig, M.H.; Lee, E.J.; Choi, I. Mechanistic insights into the antimicrobial actions of metallic nanoparticles and their implications for multidrug resistance. Int. J. Mol. Sci. 2019, 20, 2468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Y.; Zhang, G.J.; Shi, X.M.; Xu, G.M.; Yang, Y. Chemical mechanisms of bacterial inactivation using dielectric barrier discharge plasma in atmospheric air. IEEE Trans. Plasma Sci. 2008, 36, 1615–1620. [Google Scholar] [CrossRef]
- Kelly-Wintenberg, K.; Hodge, A.; Montie, T.C.; Deleanu, L.; Sherman, D.; Reece Roth, J.; Tsai, P.; Wadsworth, L. Use of a one atmosphere uniform glow discharge plasma to kill a broad spectrum of microorganisms. J. Vac. Sci. Technol. A Vac. Surf. Films 1999, 17, 1539–1544. [Google Scholar] [CrossRef]
- Ziuzina, D.; Patil, S.; Cullen, P.J.; Boehm, D.; Bourke, P. Dielectric barrier discharge atmospheric cold plasma for inactivation of Pseudomonas aeruginosa biofilms. Plasma Med. 2014, 4, 137–152. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.-S.; Lee, E.-J.; Kim, Y.-J. Inactivation of Campylobacter jejuni with Dielectric Barrier Discharge Plasma Using Air and Nitrogen Gases. Foodborne Pathog. Dis. 2014, 11, 645–651. [Google Scholar] [CrossRef]
- Smolková, B.; Uzhytchak, M.; Lynnyk, A.; Kubinová, Š.; Dejneka, A.; Lunov, O. A critical review on selected external physical cues and modulation of cell behavior: Magnetic Nanoparticles, Non-thermal Plasma and Lasers. J. Funct. Biomater. 2019, 10, 2. [Google Scholar] [CrossRef] [Green Version]
- Mendis, D.A.; Rosenberg, M.; Azam, F. A note on the possible electrostatic disruption of bacteria. IEEE Trans. Plasma Sci. 2000, 28, 1304–1306. [Google Scholar] [CrossRef]
- Ito, M.; Ohta, T.; Hori, M. Plasma agriculture. J. Korean Phys. Soc. 2012, 60, 937–943. [Google Scholar] [CrossRef]
- Puač, N.; Gherardi, M.; Shiratani, M. Plasma agriculture: A rapidly emerging field. Plasma Process. Polym. 2018, 15, 1700174. [Google Scholar] [CrossRef]
- Attri, P.; Ishikawa, K.; Okumura, T.; Koga, K.; Shiratani, M. Plasma agriculture from laboratory to farm: A review. Processes 2020, 8, 1002. [Google Scholar] [CrossRef]
- Guo, J.; Huang, K.; Wang, J. Bactericidal effect of various non-thermal plasma agents and the influence of experimental conditions in microbial inactivation: A review. Food Control 2015, 50, 482–490. [Google Scholar] [CrossRef]
- Cockerill, F.R.; Wikler, M.A.; Alder, J.; Dudley, M.N.; Eliopoulos, G.M.; Ferraro, M.J.; Hardy, D.J.; Hecht, D.W.; Hindler, J.A.; Patel, J.B.; et al. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically, Approved Standard, 9th ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2012. [Google Scholar]
- Potrykus, M.; Sledz, W.; Golanowska, M.; Slawiak, M.; Binek, A.; Motyka, A.; Zoledowska, S.; Czajkowski, R.; Lojkowska, E. Simultaneous detection of major blackleg and soft rot bacterial pathogens in potato by multiplex polymerase chain reaction. Ann. Appl. Biol. 2014, 165, 474–487. [Google Scholar] [CrossRef]
- Slawiak, M.; Łojkowska, E.; van der Wolf, J.M. First report of bacterial soft rot on potato caused by Dickeya sp. (syn. Erwinia chrysanthemi) in Poland. Plant Pathol. 2009, 58, 794. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Pathogenic Species | Strain Nos. a | Disease | Country of Isolation | Year of Isolation | Reference |
|---|---|---|---|---|---|
| Pectobacterium carotovorum | IFB5118 SCRI136 | Blackleg | USA | NA | SCRI collection [51] |
| Pectobacterium atrosepticum | IFB5103 SCRI1086 | Blackleg and soft rot | Canada | 1985 | SCRI collection [51] |
| Dickeya solani | IFB0099 IPO2276 LMG28824 | Blackleg and soft rot | Poland | 2005 | Slawiak et al. [52] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Motyka-Pomagruk, A.; Dzimitrowicz, A.; Orlowski, J.; Babinska, W.; Terefinko, D.; Rychlowski, M.; Prusinski, M.; Pohl, P.; Lojkowska, E.; Jamroz, P.; et al. Implementation of a Non-Thermal Atmospheric Pressure Plasma for Eradication of Plant Pathogens from a Surface of Economically Important Seeds. Int. J. Mol. Sci. 2021, 22, 9256. https://doi.org/10.3390/ijms22179256
Motyka-Pomagruk A, Dzimitrowicz A, Orlowski J, Babinska W, Terefinko D, Rychlowski M, Prusinski M, Pohl P, Lojkowska E, Jamroz P, et al. Implementation of a Non-Thermal Atmospheric Pressure Plasma for Eradication of Plant Pathogens from a Surface of Economically Important Seeds. International Journal of Molecular Sciences. 2021; 22(17):9256. https://doi.org/10.3390/ijms22179256
Chicago/Turabian StyleMotyka-Pomagruk, Agata, Anna Dzimitrowicz, Jakub Orlowski, Weronika Babinska, Dominik Terefinko, Michal Rychlowski, Michal Prusinski, Pawel Pohl, Ewa Lojkowska, Piotr Jamroz, and et al. 2021. "Implementation of a Non-Thermal Atmospheric Pressure Plasma for Eradication of Plant Pathogens from a Surface of Economically Important Seeds" International Journal of Molecular Sciences 22, no. 17: 9256. https://doi.org/10.3390/ijms22179256
APA StyleMotyka-Pomagruk, A., Dzimitrowicz, A., Orlowski, J., Babinska, W., Terefinko, D., Rychlowski, M., Prusinski, M., Pohl, P., Lojkowska, E., Jamroz, P., & Sledz, W. (2021). Implementation of a Non-Thermal Atmospheric Pressure Plasma for Eradication of Plant Pathogens from a Surface of Economically Important Seeds. International Journal of Molecular Sciences, 22(17), 9256. https://doi.org/10.3390/ijms22179256