
Pemafibrate Prevents Retinal Dysfunction in a Mouse Model of Unilateral Common Carotid Artery Occlusion

 ,
,  ,
,  ,
,  , and
, and 
Abstract
:1. Introduction
2. Results
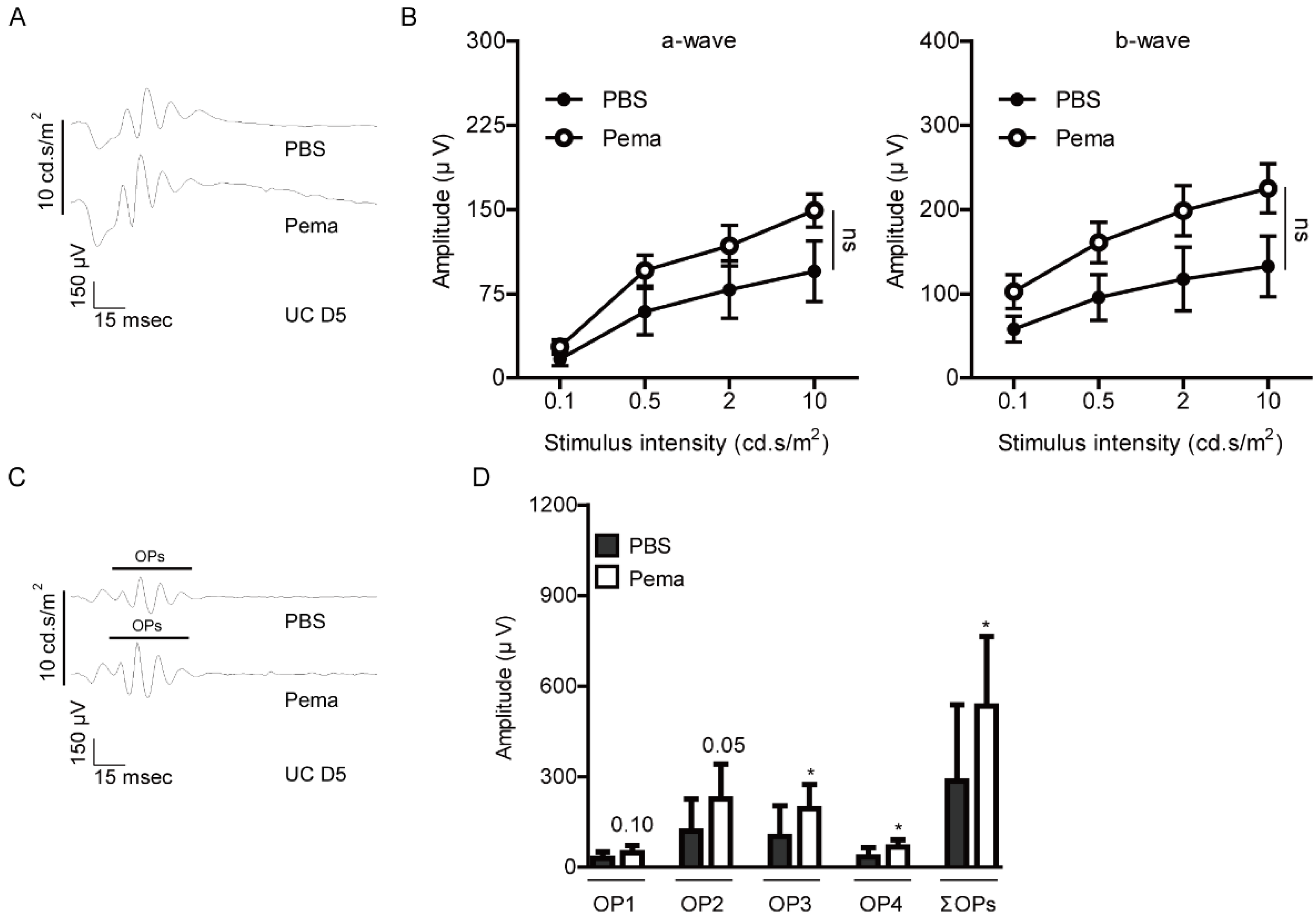
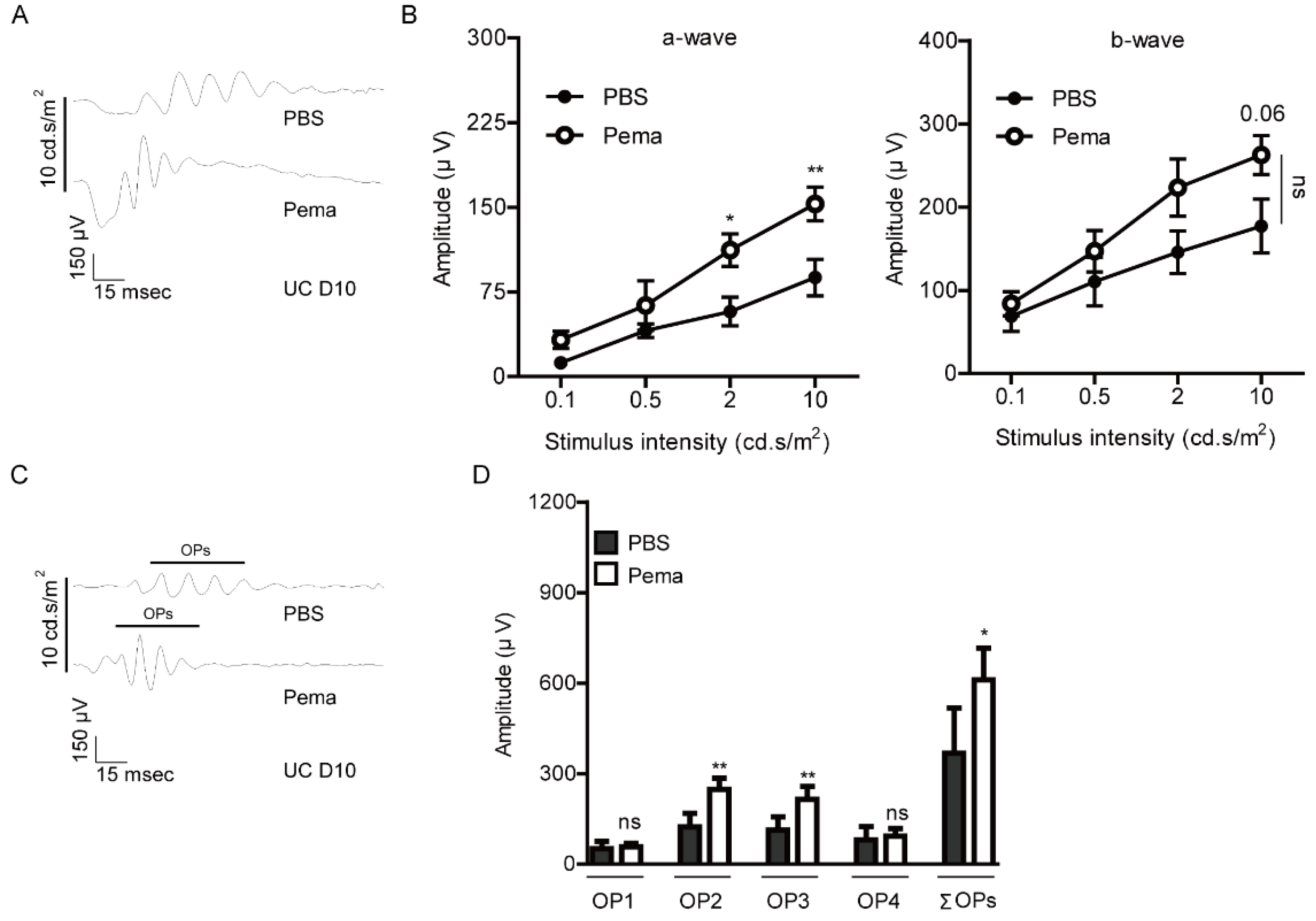
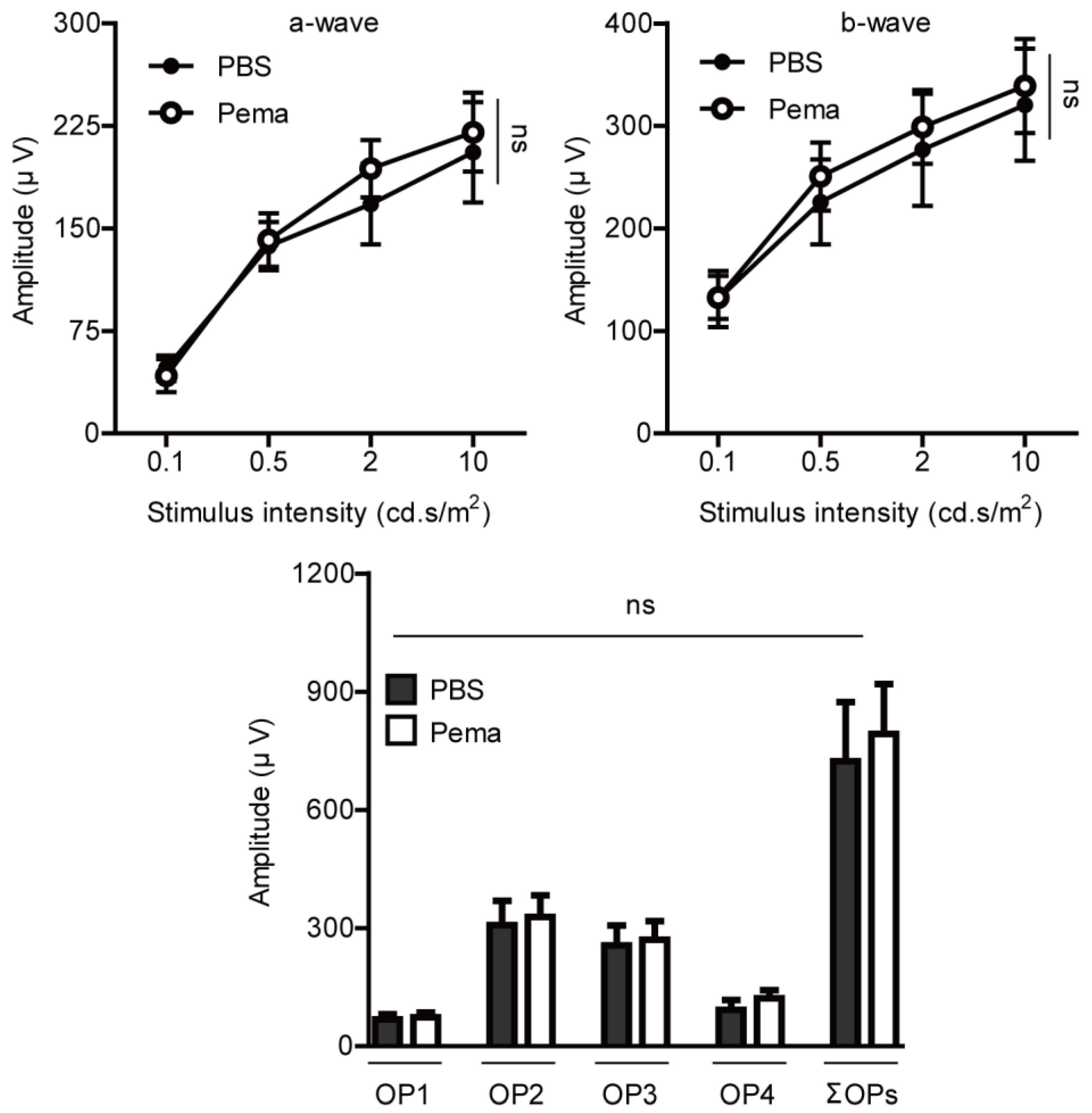
2.1. Suppression of Retinal Dysfunction by Pemafibrate Administration in a Mouse Model of UCCAO-Induced Retinal Ischemia
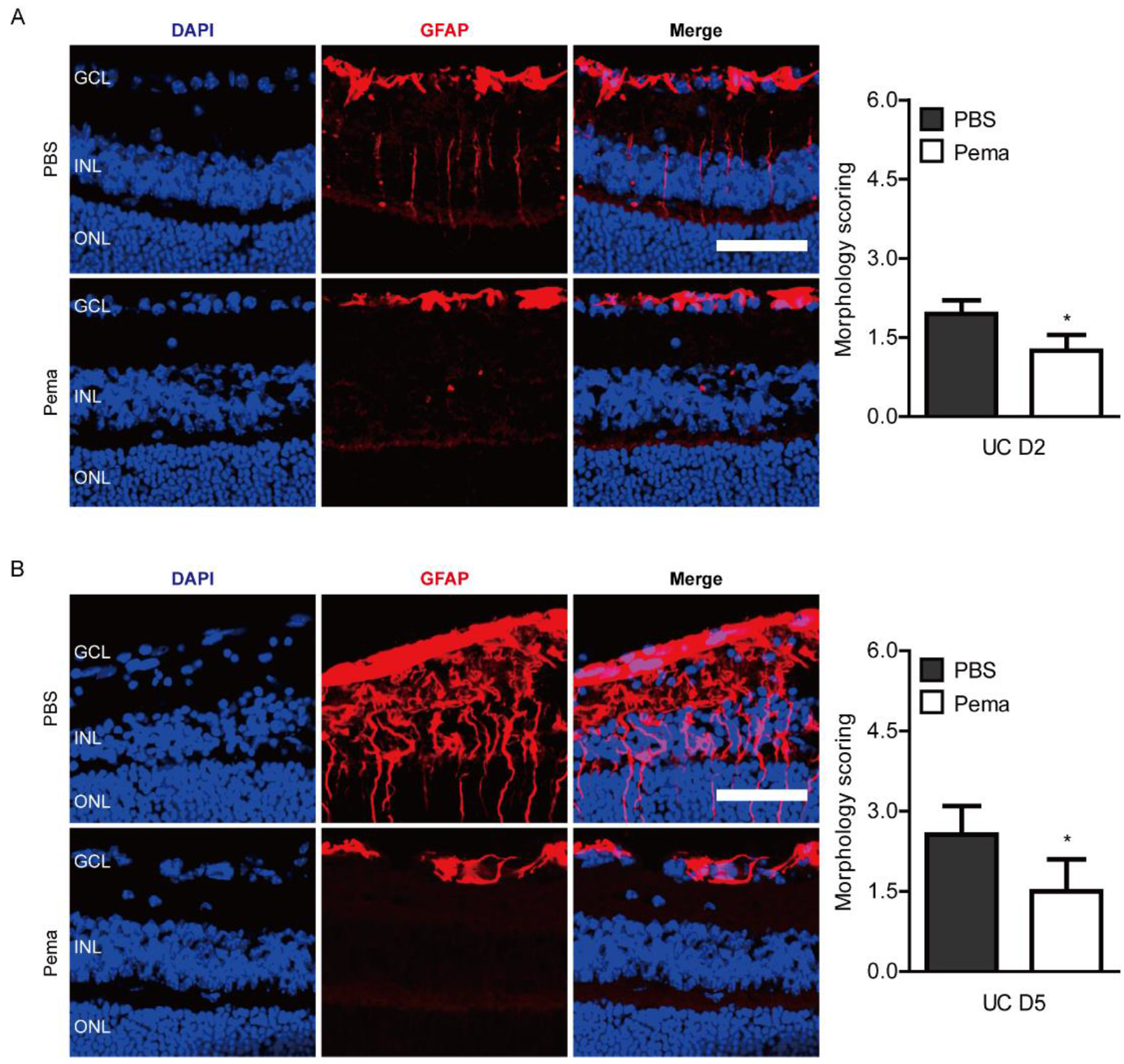
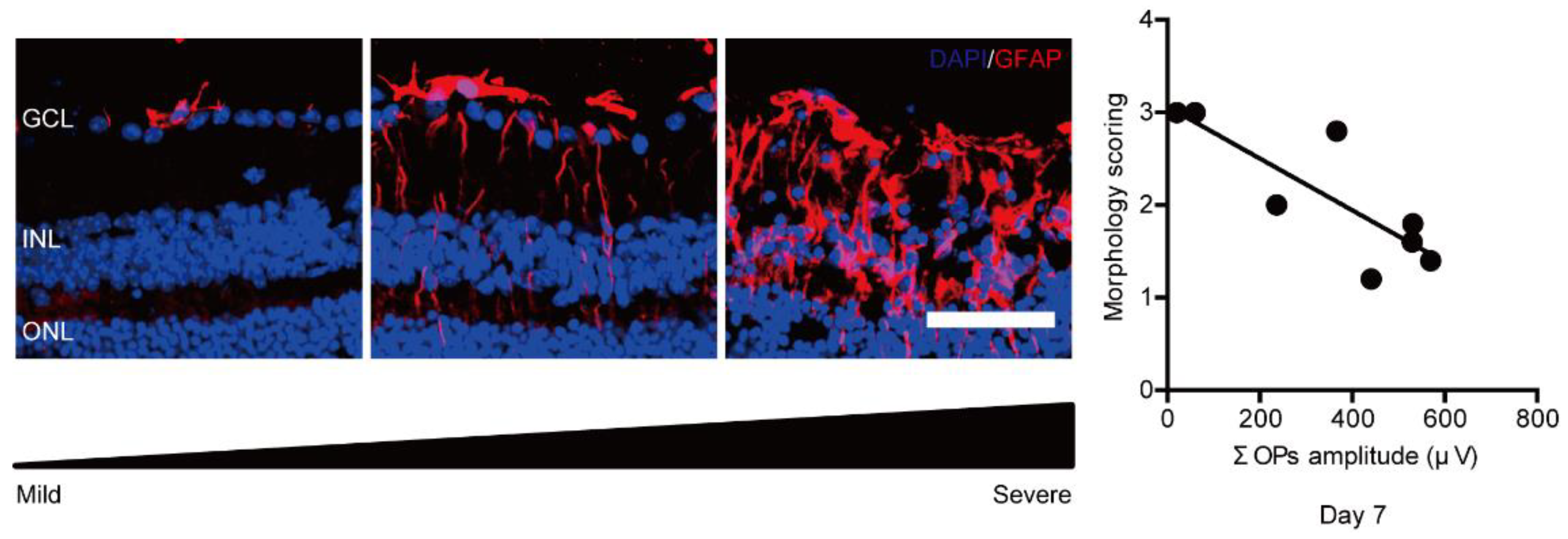
2.2. Suppression of Pathological Retinal Gliosis by Pemafibrate Administration in a Mouse Model of UCCAO-Induced Retinal Ischemia
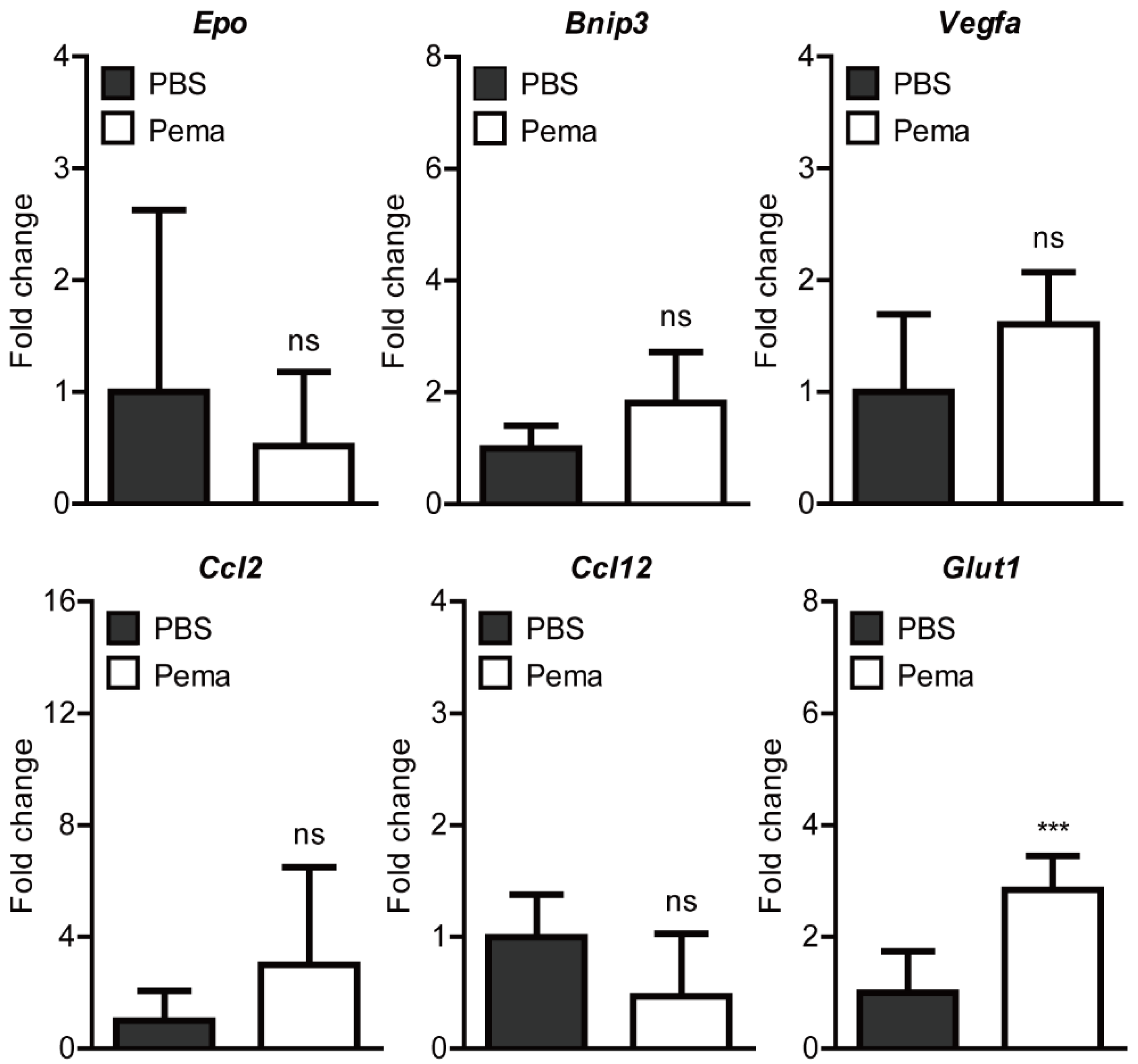
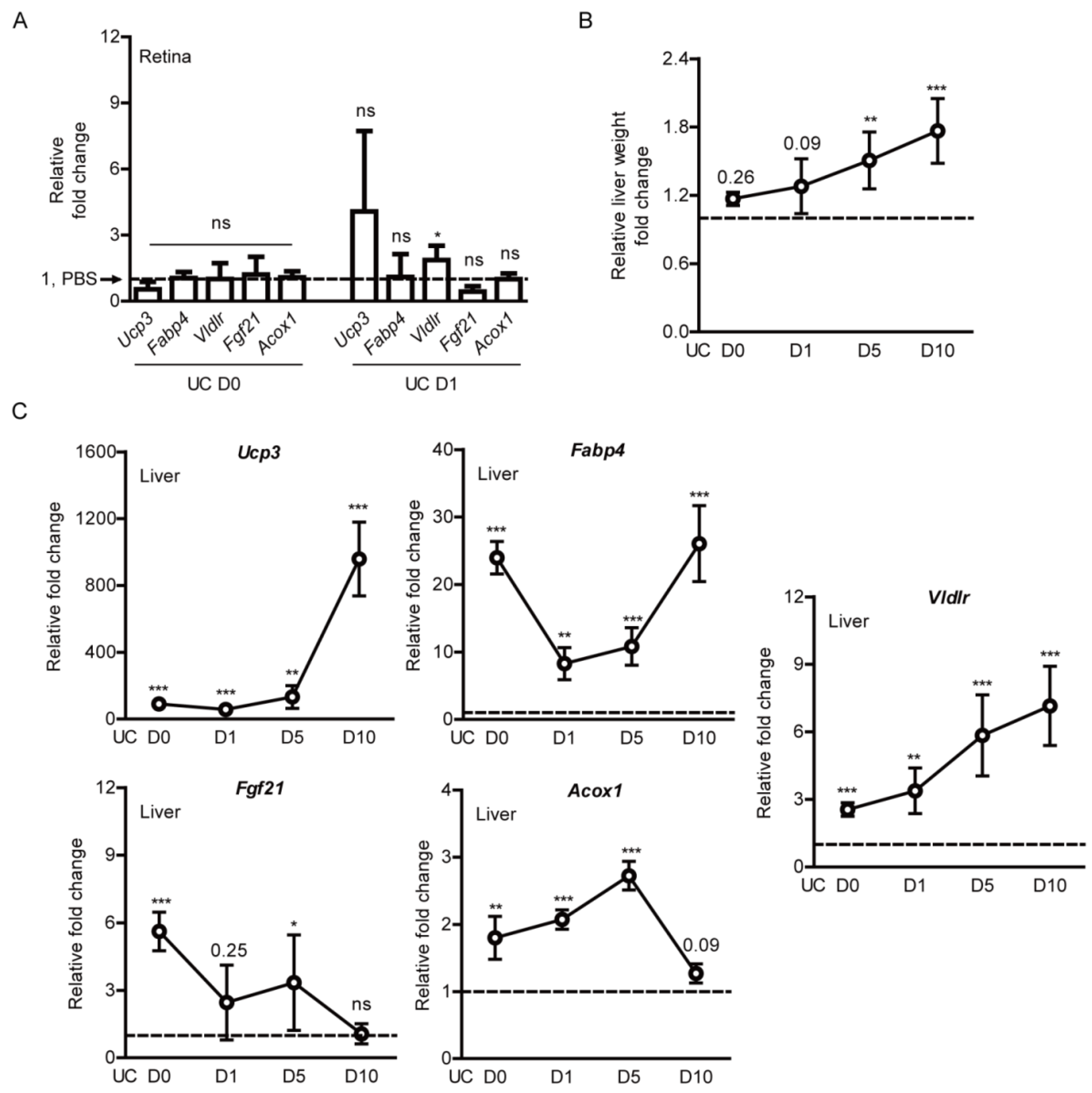
2.3. Screening of Hypoxia-Ischemia-Related Gene Expressions after Pemafibrate Administration in a Mouse Model of UCCAO-Induced Retinal Ischemia
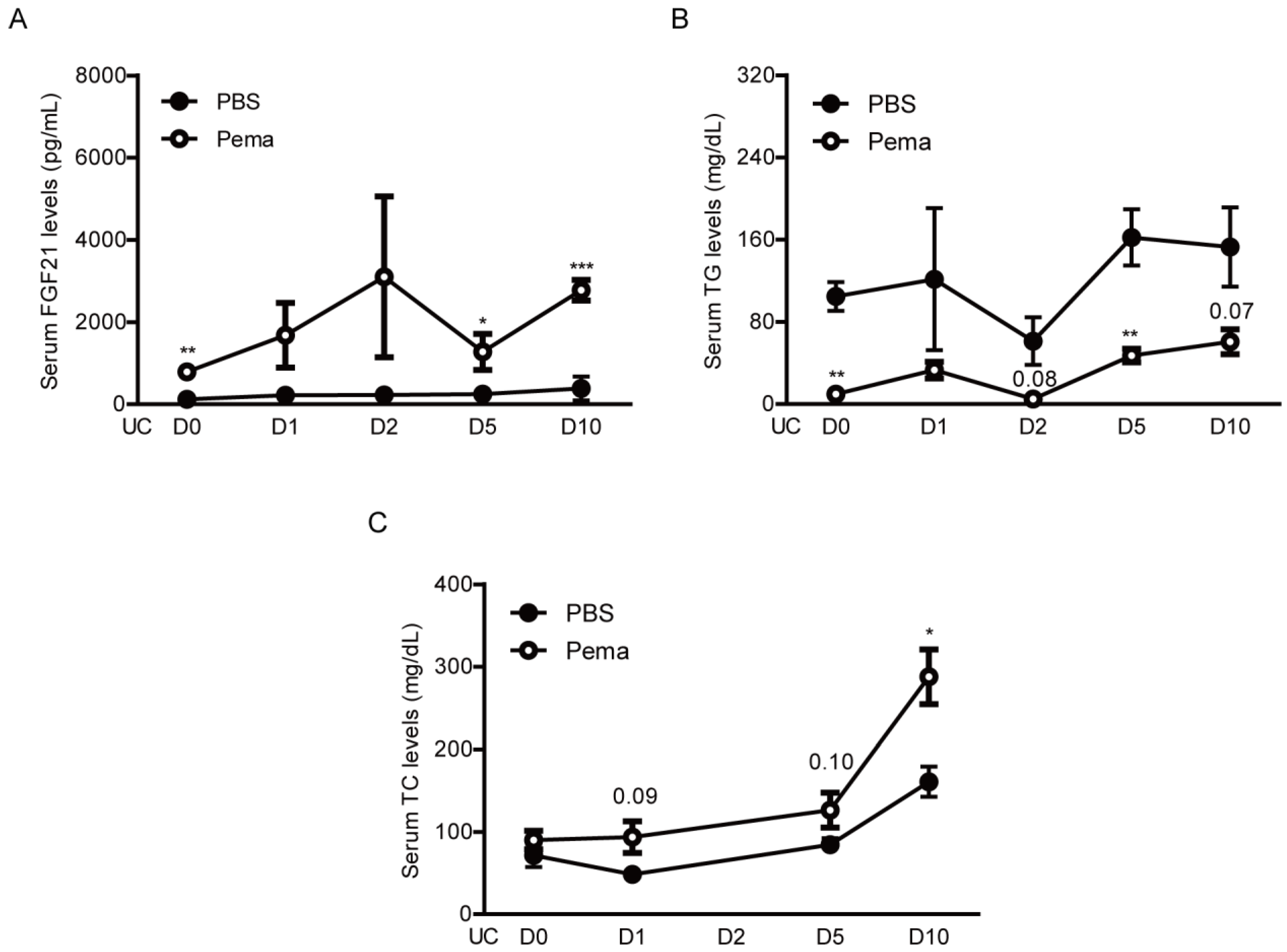
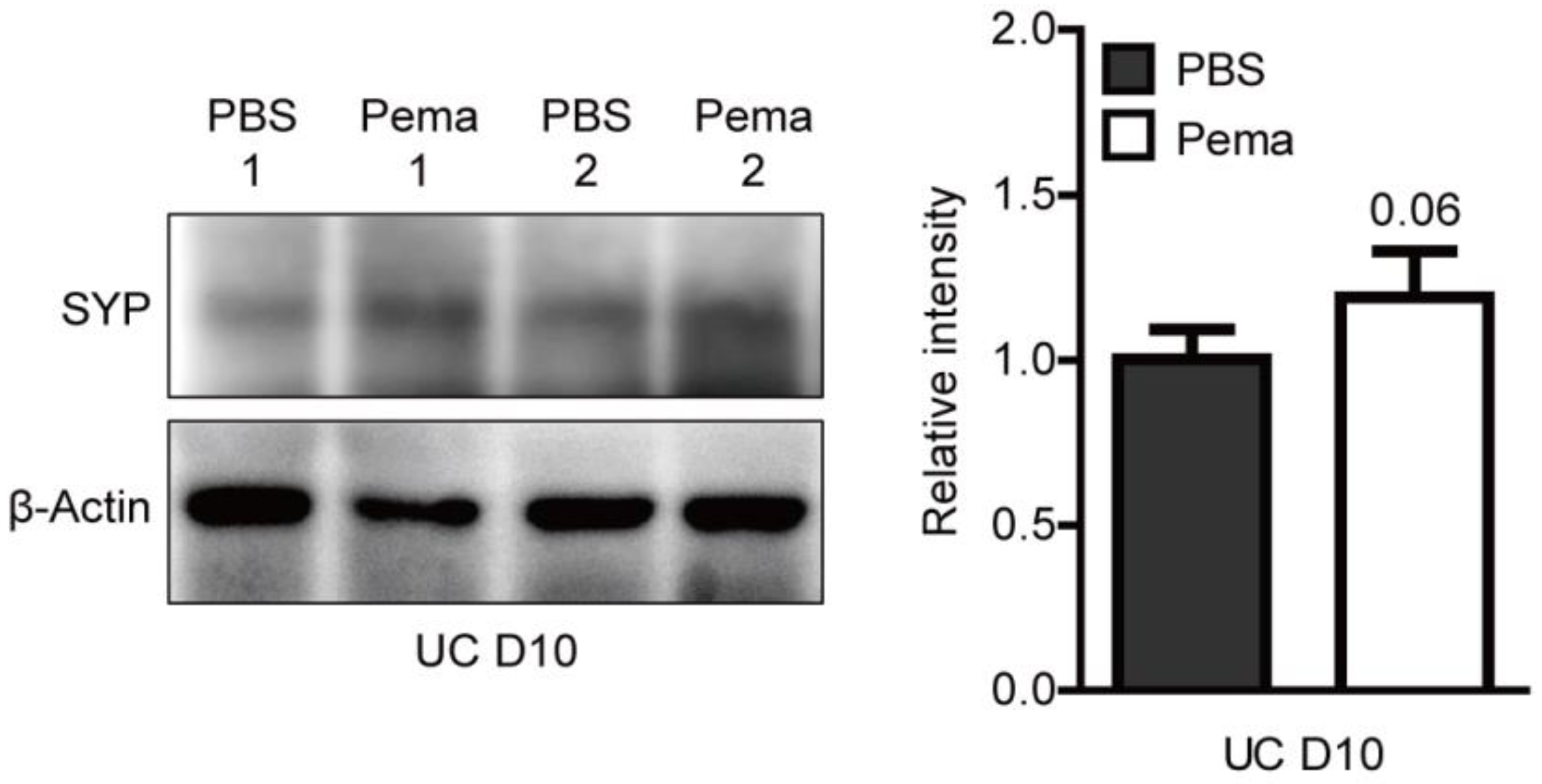
2.4. Induction of PPARα Target Genes by Pemafibrate Administration in a Mouse Model of UCCAO-Induced Retinal Ischemia
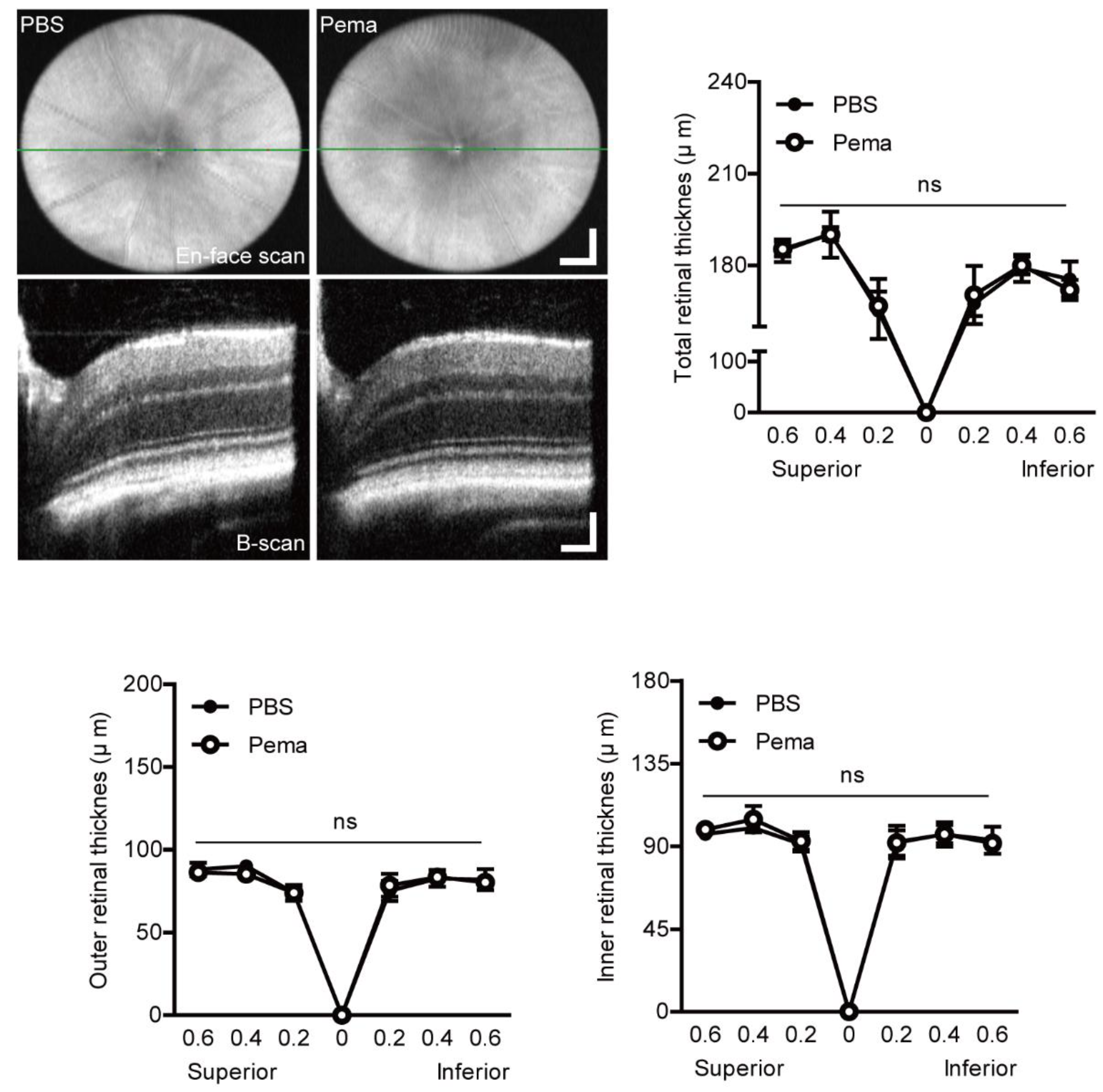
2.5. Observation of Retinal Thickness Changes by Pemafibrate Administration in a Mouse Model of UCCAO-Induced Retinal Ischemia
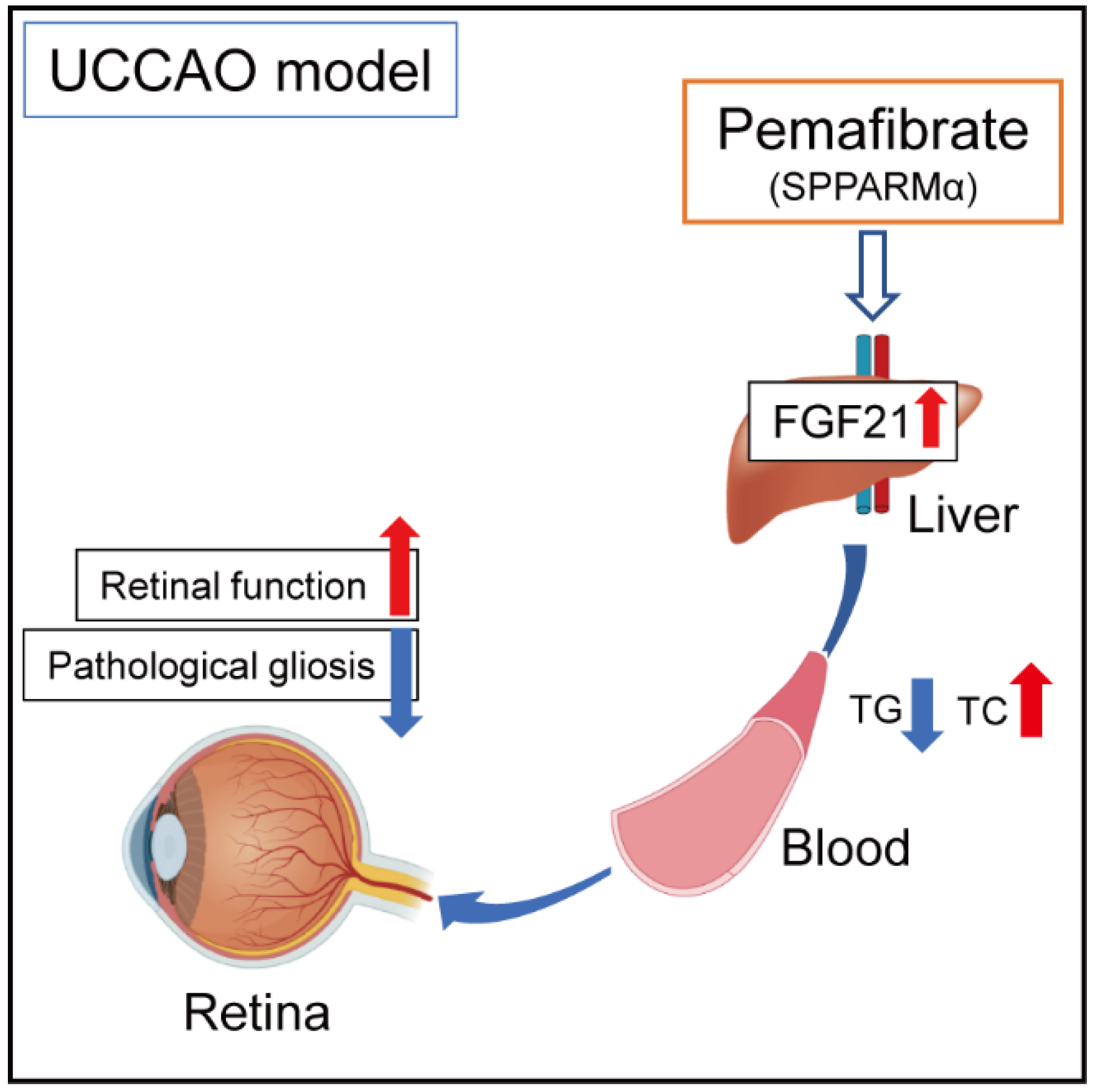
3. Discussion
4. Materials and Methods
4.1. Animal
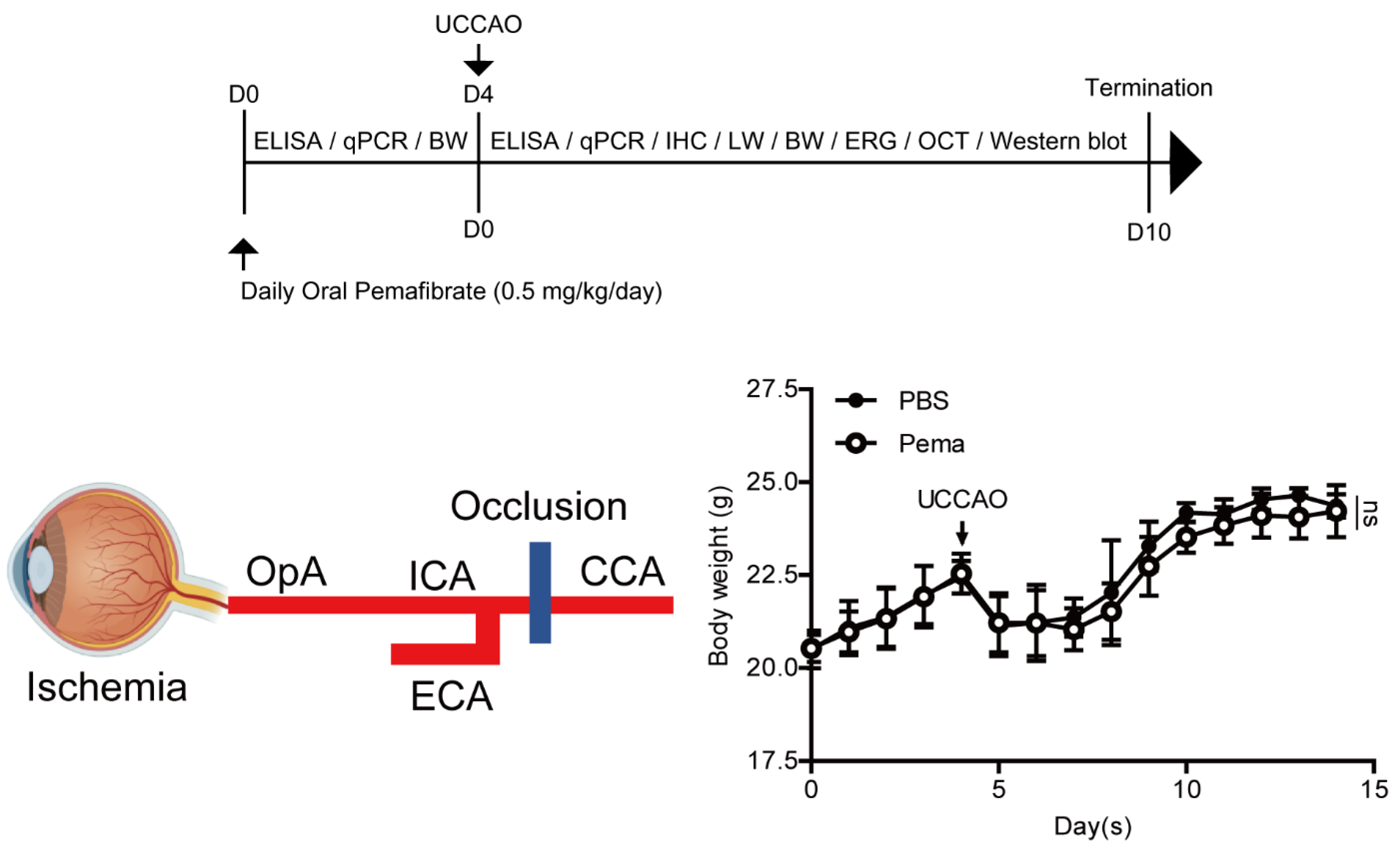
4.2. A Murine Model of UCCAO-Induced Retinal Ischemia and Oral Administration of Pemafibrate
4.3. Optical Coherence Tomography (OCT)
4.4. Electroretinography (ERG)
4.5. Immunohistochemistry (IHC)
4.6. Measurement of Serum FGF21, TC, and TG Levels
4.7. Quantitative PCR
4.8. Western Blotting
4.9. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

Appendix B

Appendix C

Appendix D

Appendix E

References
- Terelak-Borys, B.; Skonieczna, K.; Grabska-Liberek, I. Ocular ischemic syndrome—A systematic review. Med. Sci. Monit. 2012, 18, RA138–RA144. [Google Scholar] [CrossRef] [Green Version]
- Hedges, T.R. Ophthalmoscopic findings in internal carotid artery occlusion. Am. J. Ophthalmol. 1963, 55, 1007–1012. [Google Scholar] [CrossRef]
- Sturrock, G.D.; Mueller, H.R. Chronic ocular ischaemia. Br. J. Ophthalmol. 1984, 68, 716–723. [Google Scholar] [CrossRef]
- Brown, G.C.; Magargal, L.E. The ocular ischemic syndrome. Int. Ophthalmol. 1988, 11, 239–251. [Google Scholar] [CrossRef]
- Duker, J.S.; Belmont, J.B. Ocular ischemic syndrome secondary to carotid artery dissection. Am. J. Ophthalmol. 1988, 106, 750–752. [Google Scholar] [CrossRef]
- Hamed, L.M.; Guy, J.R.; Moster, M.L.; Bosley, T. Giant cell arteritis in the ocular ischemic syndrome. Am. J. Ophthalmol. 1992, 113, 702–705. [Google Scholar] [CrossRef]
- Sadun, A.; Sebag, J.; Bienfang, D.C. Complete bilateral internal carotid artery occlusion in a young man. J. Clin. Neuro-Ophthalmol. 1983, 3, 63–66. [Google Scholar]
- Malhotra, R. Management of ocular ischaemic syndrome. Br. J. Ophthalmol. 2000, 84, 1428–1431. [Google Scholar] [CrossRef] [Green Version]
- Sood, G.; Siddik, A.B. Ocular ischemic syndrome. In StatPearls; StatPearls Publishing LLC: Treasure Island, FL, USA, 2021. [Google Scholar]
- Sivalingam, A.; Brown, G.C.; Magargal, L.E. The ocular ischemic syndrome—III Visual prognosis and the effect of treatment. Int. Ophthalmol. 1991, 15, 15–20. [Google Scholar] [CrossRef]
- Sun, W.; Geng, Y.; Chen, Y.; Tang, X.-H.; Zhang, Y.; Gu, S.-H.; Xie, J.-J.; Zhang, Z.-A.; Tian, X.-S. Differences of brain pathological changes and cognitive function after bilateral common carotid artery occlusion between Sprague-Dawley and Wistar rats. Acta Physiol. Sin. 2019, 71, 705–716. [Google Scholar]
- Yamamoto, H.; Schmidt-Kastner, R.; Hamasaki, D.I.; Yamamoto, H.; Parel, J.-M. Complex neurodegeneration in retina following moderate ischemia induced by bilateral common carotid artery occlusion in Wistar rats. Exp. Eye Res. 2006, 82, 767–779. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.; Ji, M.; Deng, T.; Luo, D.; Zi, Y.; Pan, L.; Wang, Z.; Jin, M. Functional and morphologic study of retinal hypoperfusion injury induced by bilateral common carotid artery occlusion in rats. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Crespo-Garcia, S.; Reichhart, N.; Skosyrski, S.; Foddis, M.; Wu, J.; Figura, A.; Herrspiegel, C.; Fuchtemeier, M.; Sassi, C.; Dirnagl, U.; et al. Individual and temporal variability of the retina after chronic bilateral common carotid artery occlusion (BCCAO). PLoS ONE 2018, 13, e0193961. [Google Scholar]
- Lavinsky, D.; Arterni, N.S.; Achaval, M.; Netto, C.A. Chronic bilateral common carotid artery occlusion: A model for ocular ischemic syndrome in the rat. Graefe’s Arch. Clin. Exp. Ophthalmol. 2005, 244, 199–204. [Google Scholar] [CrossRef] [PubMed]
- Kalesnykas, G.; Tuulos, T.; Uusitalo, H.; Jolkkonen, J. Neurodegeneration and cellular stress in the retina and optic nerve in rat cerebral ischemia and hypoperfusion models. Neuroscience 2008, 155, 937–947. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Kang, H.; Yoon, K.Y.; Chang, Y.Y.; Song, H.B. A mouse model of retinal hypoperfusion injury induced by unilateral common carotid artery occlusion. Exp. Eye Res. 2020, 201, 108275. [Google Scholar] [CrossRef]
- Lee, B.J.; Jun, H.O.; Kim, J.H. Astrocytic cystine/glutamate antiporter is a key regulator of erythropoietin expression in the ischemic retina. FASEB J. 2019, 33, 6045–6054. [Google Scholar] [CrossRef]
- Hayreh, S.S.; Zimmerman, M.B. Ocular arterial occlusive disorders and carotid artery disease. Ophthalmol. Retin. 2016, 1, 12–18. [Google Scholar] [CrossRef] [Green Version]
- Weymouth, W.; Pedersen, C. Central retinal artery occlusion associated with carotid artery occlusion. Clin. Pr. Cases Emerg. Med. 2019, 3, 233–236. [Google Scholar] [CrossRef]
- Yang, G.; Kitagawa, K.; Matsushita, K.; Mabuchi, T.; Yagita, Y.; Yanagihara, T.; Matsumoto, M. C57BL/6 strain is most susceptible to cerebral ischemia following bilateral common carotid occlusion among seven mouse strains: Selective neuronal death in the murine transient forebrain ischemia. Brain Res. 1997, 752, 209–218. [Google Scholar] [CrossRef]
- Lee, D.; Miwa, Y.; Jeong, H.; Ikeda, S.-I.; Katada, Y.; Tsubota, K.; Kurihara, T. A murine model of ischemic retinal injury induced by transient bilateral common carotid artery occlusion. J. Vis. Exp. 2020. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Jeong, H.; Miwa, Y.; Shinojima, A.; Katada, Y.; Tsubota, K.; Kurihara, T. Retinal dysfunction induced in a mouse model of unilateral common carotid artery occlusion. PeerJ 2021, 9, e11665. [Google Scholar] [CrossRef]
- Tomita, Y.; Lee, D.; Tsubota, K.; Kurihara, T. PPARα agonist oral therapy in diabetic retinopathy. Biomedicines 2020, 8, 433. [Google Scholar] [CrossRef]
- Keech, A.; Mitchell, P.; Summanen, P.; O’Day, J.; Davis, T.; Moffitt, M.; Taskinen, M.-R.; Simes, R.; Tse, D.; Williamson, E.; et al. Effect of fenofibrate on the need for laser treatment for diabetic retinopathy (FIELD study): A randomised controlled trial. Lancet 2007, 370, 1687–1697. [Google Scholar] [CrossRef]
- ACCORD Study Group and ACCORD Eye Study Group. Effects of medical therapies on retinopathy progression in type 2 diabetes. N. Engl. J. Med. 2010, 363, 233–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, D.; Tomita, Y.; Miwa, Y.; Jeong, H.; Mori, K.; Tsubota, K.; Kurihara, T. Fenofibrate protects against retinal dysfunction in a murine model of common carotid artery occlusion-induced ocular ischemia. Pharmaceuticals 2021, 14, 223. [Google Scholar] [CrossRef]
- Noonan, J.E.; Jenkins, A.J.; Ma, J.-X.; Keech, A.C.; Wang, J.J.; Lamoureux, E.L. An update on the molecular actions of fenofibrate and its clinical effects on diabetic retinopathy and other microvascular end points in patients with diabetes. Diabetes 2013, 62, 3968–3975. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Hu, Y.; Lin, M.; Jenkins, A.J.; Keech, A.C.; Mott, R.; Lyons, T.J.; Ma, J.-X. Therapeutic effects of PPARα agonists on diabetic retinopathy in type 1 diabetes models. Diabetes 2012, 62, 261–272. [Google Scholar] [CrossRef] [Green Version]
- Tomita, Y.; Ozawa, N.; Miwa, Y.; Ishida, A.; Ohta, M.; Tsubota, K.; Kurihara, T. Pemafibrate prevents retinal pathological neovascularization by increasing FGF21 level in a murine oxygen-induced retinopathy model. Int. J. Mol. Sci. 2019, 20, 5878. [Google Scholar] [CrossRef] [Green Version]
- Tomita, Y.; Lee, D.; Miwa, Y.; Jiang, X.; Ohta, M.; Tsubota, K.; Kurihara, T. Pemafibrate protects against retinal dysfunction in a murine model of diabetic retinopathy. Int. J. Mol. Sci. 2020, 21, 6243. [Google Scholar] [CrossRef]
- Shiono, A.; Sasaki, H.; Sekine, R.; Abe, Y.; Matsumura, Y.; Inagaki, T.; Tanaka, T.; Kodama, T.; Aburatani, H.; Sakai, J.; et al. PPARα activation directly upregulates thrombomodulin in the diabetic retina. Sci. Rep. 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- Fujita, N.; Sase, K.; Tsukahara, C.; Arizono, I.; Takagi, H.; Kitaoka, Y. Pemafibrate prevents retinal neuronal cell death in NMDA-induced excitotoxicity via inhibition of p-c-Jun expression. Mol. Biol. Rep. 2020, 48, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Wachtmeister, L. Oscillatory potentials in the retina: What do they reveal. Prog. Retin. Eye Res. 1998, 17, 485–521. [Google Scholar] [CrossRef]
- Heynen, H.; Wachtmeister, L.; van Norren, D. Origin of the oscillatory potentials in the primate retina. Vis. Res. 1985, 25, 1365–1373. [Google Scholar] [CrossRef]
- Yonemura, D.; Kawasaki, K. New approaches to ophthalmic electrodiagnosis by retinal oscillatory potential, drug-induced responses from retinal pigment epithelium and cone potential. Doc. Ophthalmol. 1979, 48, 163–222. [Google Scholar] [CrossRef]
- de Hoz, R.; Rojas, B.; Ramírez, A.; Salazar, J.J.; Gallego, B.I.; Triviño, A.; Ramírez, J.M. Retinal macroglial responses in health and disease. BioMed Res. Int. 2016, 2016, 2954721. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, Y.; Raza-Iqbal, S.; Tanaka, T.; Murakami, K.; Anai, M.; Osawa, T.; Matsumura, Y.; Sakai, J.; Kodama, T. Gene expression profiles induced by a novel selective peroxisome proliferator-activated receptor α modulator (SPPARMα) pemafibrate. Int. J. Mol. Sci. 2019, 20, 5682. [Google Scholar] [CrossRef] [Green Version]
- Badman, M.K.; Pissios, P.; Kennedy, A.R.; Koukos, G.; Flier, J.S.; Maratos-Flier, E. Hepatic fibroblast growth factor 21 is regulated by PPARα and is a key mediator of hepatic lipid metabolism in ketotic states. Cell Metab. 2007, 5, 426–437. [Google Scholar] [CrossRef] [Green Version]
- Ong, K.-L.; Januszewski, A.S.; O’Connell, R.; Jenkins, A.; Xu, A.; Sullivan, D.R.; Barter, P.J.; Hung, W.-T.; Scott, R.S.; Taskinen, M.-R.; et al. The relationship of fibroblast growth factor 21 with cardiovascular outcome events in the fenofibrate intervention and event lowering in diabetes study. Diabetologia 2014, 58, 464–473. [Google Scholar] [CrossRef]
- Ong, K.-L.; O’Connell, R.; Januszewski, A.S.; Jenkins, A.J.; Xu, A.; Sullivan, D.R.; Barter, P.J.; Scott, R.S.; Taskinen, M.-R.; Waldman, B.; et al. Baseline circulating FGF21 concentrations and increase after fenofibrate treatment predict more rapid glycemic progression in type 2 diabetes: Results from the FIELD study. Clin. Chem. 2017, 63, 1261–1270. [Google Scholar] [CrossRef] [PubMed]
- Ong, K.L.; Rye, K.-A.; O’Connell, R.; Jenkins, A.J.; Brown, C.; Xu, A.; Sullivan, D.R.; Barter, P.J.; Keech, A.C.; FIELD Study Investigators. Long-term fenofibrate therapy increases fibroblast growth factor 21 and retinol-binding protein 4 in subjects with type 2 diabetes. J. Clin. Endocrinol. Metab. 2012, 97, 4701–4708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sairyo, M.; Kobayashi, T.; Masuda, D.; Kanno, K.; Zhu, Y.; Okada, T.; Koseki, M.; Ohama, T.; Nishida, M.; Sakata, Y.; et al. A novel selective PPARα modulator (SPPARMα), K-877 (pemafibrate), attenuates postprandial hypertriglyceridemia in mice. J. Atheroscler. Thromb. 2018, 25, 142–152. [Google Scholar] [CrossRef] [Green Version]
- Bando, H.; Taneda, S.; Manda, N. Efficacy and safety of low-dose pemafibrate therapy for hypertriglyceridemia in patients with type 2 diabetes. JMA J. 2021, 4, 135–140. [Google Scholar] [CrossRef] [PubMed]
- Komiya, I.; Yamamoto, A.; Sunakawa, S.; Wakugami, T. Pemafibrate decreases triglycerides and small, dense LDL, but increases LDL-C depending on baseline triglycerides and LDL-C in type 2 diabetes patients with hypertriglyceridemia: An observational study. Lipids Heal. Dis. 2021, 20, 1–11. [Google Scholar] [CrossRef]
- Macosko, E.Z.; Basu, A.; Satija, R.; Nemesh, J.; Shekhar, K.; Goldman, M.; Tirosh, I.; Bialas, A.R.; Kamitaki, N.; Martersteck, E.M.; et al. Highly parallel genome-wide expression profiling of individual cells using nanoliter droplets. Cell 2015, 161, 1202–1214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lundåsen, T.; Hunt, M.C.; Nilsson, L.-M.; Sanyal, S.; Angelin, B.; Alexson, S.E.; Rudling, M. PPARα is a key regulator of hepatic FGF21. Biochem. Biophys. Res. Commun. 2007, 360, 437–440. [Google Scholar] [CrossRef] [Green Version]
- Enright, J.M.; Zhang, S.; Thebeau, C.; Siebert, E.; Jin, A.; Gadiraju, V.; Zhang, X.; Chen, S.; Semenkovich, C.F.; Rajagopal, R. Fenofibrate reduces the severity of neuroretinopathy in a type 2 model of diabetes without inducing peroxisome proliferator-activated receptor alpha-dependent retinal gene expression. J. Clin. Med. 2020, 10, 126. [Google Scholar] [CrossRef]
- Eitoh, N. FGF21 as a hepatokine, adipokine, and myokine in metabolism and diseases. Front. Endocrinol. 2014, 5, 107. [Google Scholar] [CrossRef] [Green Version]
- IItoh, N.; Nakayama, Y.; Konishi, M. Roles of FGFs as paracrine or endocrine signals in liver development, health, and disease. Front. Cell Dev. Biol. 2016, 4, 30. [Google Scholar] [CrossRef] [Green Version]
- Nishimura, T.; Nakatake, Y.; Konishi, M.; Itoh, N. Identification of a novel FGF, FGF-21, preferentially expressed in the liver. Biochim. Biophys. Acta Gene Struct. Expr. 2000, 1492, 203–206. [Google Scholar] [CrossRef]
- Tezze, C.; Romanello, V.; Sandri, M. FGF21 as modulator of metabolism in health and disease. Front. Physiol. 2019, 10, 419. [Google Scholar] [CrossRef] [PubMed]
- Talukdar, S.; Zhou, Y.; Li, D.; Rossulek, M.; Dong, J.; Somayaji, V.; Weng, Y.; Clark, R.; Lanba, A.; Owen, B.; et al. A long-acting FGF21 molecule, PF-05231023, decreases body weight and improves lipid profile in non-human primates and type 2 diabetic subjects. Cell Metab. 2016, 23, 427–440. [Google Scholar] [CrossRef] [Green Version]
- Fu, Z.; Gong, Y.; Liegl, R.; Wang, Z.; Liu, C.-H.; Meng, S.S.; Burnim, S.B.; Saba, N.J.; Fredrick, T.W.; Morss-Walton, P.; et al. FGF21 administration suppresses retinal and choroidal neovascularization in mice. Cell Rep. 2017, 18, 1606–1613. [Google Scholar] [CrossRef] [PubMed]
- Fu, Z.; Wang, Z.; Liu, C.-H.; Gong, Y.; Cakir, B.; Liegl, R.; Sun, Y.; Meng, S.S.; Burnim, S.B.; Arellano, I.; et al. Fibroblast growth factor 21 protects photoreceptor function in type 1 diabetic mice. Diabetes 2018, 67, 974–985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomita, Y.; Fu, Z.; Wang, Z.; Cakir, B.; Cho, S.S.; Britton, W.; Sun, Y.; Hellström, A.; Talukdar, S.; Smith, L.E. Long-acting FGF21 inhibits retinal vascular leakage in in vivo and in vitro models. Int. J. Mol. Sci. 2020, 21, 1188. [Google Scholar] [CrossRef] [Green Version]
- Fu, Z.; Qiu, C.; Cagnone, G.; Tomita, Y.; Huang, S.; Cakir, B.; Kotoda, Y.; Allen, W.; Bull, E.; Akula, J.D.; et al. Retinal glial remodeling by FGF21 preserves retinal function during photoreceptor degeneration. iScience 2021, 24, 102376. [Google Scholar] [CrossRef]
- Staiger, H.; Keuper, M.; Berti, L.; de Angelis, M.H.; Häring, H.-U. Fibroblast growth factor 21—Metabolic role in mice and men. Endocr. Rev. 2017, 38, 468–488. [Google Scholar] [CrossRef]
- Liu, M.; Cao, H.; Hou, Y.; Sun, G.; Li, D.; Wang, W. Liver plays a major role in FGF-21 mediated glucose homeostasis. Cell. Physiol. Biochem. 2018, 45, 1423–1433. [Google Scholar] [CrossRef]
- Ye, X.; Qi, J.; Yu, D.; Wu, Y.; Zhu, S.; Li, S.; Wu, Q.; Ren, G.; Li, D. Pharmacological efficacy of FGF21 analogue, liraglutide and insulin glargine in treatment of type 2 diabetes. J. Diabetes Complicat. 2017, 31, 726–734. [Google Scholar] [CrossRef]
- Yu, D.; Ye, X.; Wu, Q.; Li, S.; Yang, Y.; He, J.; Liu, Y.; Zhang, X.; Yuan, Q.; Liu, M.; et al. Insulin sensitizes FGF21 in glucose and lipid metabolisms via activating common AKT pathway. Endocrine 2015, 52, 527–540. [Google Scholar] [CrossRef]
- Li, K.; Li, L.; Yang, M.; Liu, H.; Boden, G.; Yang, G. The effects of fibroblast growth factor-21 knockdown and over-expression on its signaling pathway and glucose-lipid metabolism in vitro. Mol. Cell. Endocrinol. 2012, 348, 21–26. [Google Scholar] [CrossRef]
- Hu, S.; Cao, S.; Liu, J. Role of angiopoietin-2 in the cardioprotective effect of fibroblast growth factor 21 on ischemia/reperfusion-induced injury in H9c2 cardiomyocytes. Exp. Ther. Med. 2017, 14, 771–779. [Google Scholar] [CrossRef] [Green Version]
- Joyal, J.-S.; Sun, Y.; Gantner, M.L.; Shao, Z.; Evans, L.P.; Saba, N.; Fredrick, T.; Burnim, S.; Kim, J.S.; Patel, G.; et al. Retinal lipid and glucose metabolism dictates angiogenesis through the lipid sensor Ffar1. Nat. Med. 2016, 22, 439–445. [Google Scholar] [CrossRef] [Green Version]
- Mohammad, H.M.; Sami, M.M.; Makary, S.; Toraih, E.A.; Mohamed, A.O.; El-Ghaiesh, S.H. Neuroprotective effect of levetiracetam in mouse diabetic retinopathy: Effect on glucose transporter-1 and GAP43 expression. Life Sci. 2019, 232, 116588. [Google Scholar] [CrossRef]
- You, Z.-P.; Zhang, Y.-L.; Shi, K.; Shi, L.; Zhang, Y.-Z.; Zhou, Y.; Wang, C.-Y. Suppression of diabetic retinopathy with GLUT1 siRNA. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badr, G.A.; Tang, J.; Ismail-Beigi, F.; Kern, T.S. Diabetes downregulates GLUT1 expression in the retina and its microvessels but not in the cerebral cortex or its microvessels. Diabetes 2000, 49, 1016–1021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandes, R.; Suzuki, K.-I.; Kumagai, A.K. Inner blood-retinal barrier GLUT1 in long-term diabetic rats: An immunogold electron microscopic study. Investig. Opthalmology Vis. Sci. 2003, 44, 3150–3154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandes, R.; Carvalho, A.L.; Kumagai, A.; Seica, R.; Hosoya, K.-I.; Terasaki, T.; Murta, J.; Pereira, P.; Faro, C. Downregulation of retinal GLUT1 in diabetes by ubiquitinylation. Mol. Vis. 2004, 10, 618–628. [Google Scholar]
- Sergeys, J.; Etienne, I.; Van Hove, I.; Lefevere, E.; Stalmans, I.; Feyen, J.H.M.; Moons, L.; Van Bergen, T. Longitudinal in vivo characterization of the streptozotocin-induced diabetic mouse model: Focus on early inner retinal responses. Investig. Opthalmology Vis. Sci. 2019, 60, 807–822. [Google Scholar] [CrossRef] [Green Version]
- Bogdanov, P.; Hernández, C.; Corraliza, L.; Carvalho, A.R.; Simó, R. Effect of fenofibrate on retinal neurodegeneration in an experimental model of type 2 diabetes. Acta Diabetol. 2014, 52, 113–122. [Google Scholar] [CrossRef]
- Brown, K.T. The electroretinogram: Its components and their origins. Vis. Res. 1968, 8, 633-IN6. [Google Scholar] [CrossRef]
- Robson, J.G.; Saszik, S.M.; Ahmed, J.; Frishman, L.J. Rod and cone contributions to the A-wave of the electroretinogram of the macaque. J. Physiol. 2003, 547, 509–530. [Google Scholar] [CrossRef] [PubMed]
- Vecino, E.; Rodriguez, F.D.; Ruzafa, N.; Pereiro, X.; Sharma, S.C. Glia-neuron interactions in the mammalian retina. Prog. Retin. Eye Res. 2016, 51, 1–40. [Google Scholar] [CrossRef] [Green Version]
- Hennuyer, N.; Duplan, I.; Paquet, C.; Vanhoutte, J.; Woitrain, E.; Touche, V.; Colin, S.; Vallez, E.; Lestavel, S.; Lefebvre, P.; et al. The novel selective PPARα modulator (SPPARMα) pemafibrate improves dyslipidemia, enhances reverse cholesterol transport and decreases inflammation and atherosclerosis. Atherosclerosis 2016, 249, 200–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, A.; Singh, R. Triglyceride and cardiovascular risk: A critical appraisal. Indian J. Endocrinol. Metab. 2016, 20, 418–428. [Google Scholar] [CrossRef] [PubMed]
- Ye, X.; Kong, W.; Zafar, M.I.; Chen, L.-L. Serum triglycerides as a risk factor for cardiovascular diseases in type 2 diabetes mellitus: A systematic review and meta-analysis of prospective studies. Cardiovasc. Diabetol. 2019, 18, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarwar, N.; Danesh, J.; Eiriksdottir, G.; Sigurdsson, G.; Wareham, N.; Bingham, S.; Boekholdt, S.M.; Khaw, K.-T.; Gudnason, V. Triglycerides and the risk of coronary heart disease. Circulation 2007, 115, 450–458. [Google Scholar] [CrossRef]
- Zoppini, G.; Negri, C.; Stoico, V.; Casati, S.; Pichiri, I.; Bonora, E. Triglyceride-high-density lipoprotein cholesterol is associated with microvascular complications in type 2 diabetes mellitus. Metabolism 2012, 61, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.-C.; Wu, W.-C. Dyslipidemia and diabetic retinopathy. Rev. Diabet. Stud. 2013, 10, 121–132. [Google Scholar] [CrossRef]
- Mohamed, Q.; Gillies, M.C.; Wong, T.Y. Management of diabetic retinopathy. JAMA 2007, 298, 902–916. [Google Scholar] [CrossRef] [Green Version]
- Fu, Z.; Chen, C.T.; Cagnone, G.; Heckel, E.; Sun, Y.; Cakir, B.; Tomita, Y.; Huang, S.; Li, Q.; Britton, W.; et al. Dyslipidemia in retinal metabolic disorders. EMBO Mol. Med. 2019, 11, e10473. [Google Scholar] [CrossRef] [PubMed]
- Jacob, J.J.; Chopra, R.; Chander, A. Ocular associations of metabolic syndrome. Indian J. Endocrinol. Metab. 2012, 16, S6–S11. [Google Scholar] [CrossRef] [PubMed]
- Naiman, S.; Huynh, F.; Gil, R.; Glick, Y.; Shahar, Y.; Touitou, N.; Nahum, L.; Avivi, M.Y.; Roichman, A.; Kanfi, Y.; et al. SIRT6 promotes hepatic beta-oxidation via activation of PPARα. Cell Rep. 2019, 29, 4127–4143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raza-Iqbal, S.; Tanaka, T.; Anai, M.; Inagaki, T.; Matsumura, Y.; Ikeda, K.; Taguchi, A.; Gonzalez, F.J.; Sakai, J.; Kodama, T. Transcriptome analysis of K-877 (a novel selective PPARα modulator (SPPARMα))-regulated genes in primary human hepatocytes and the mouse liver. J. Atheroscler. Thromb. 2015, 22, 754–772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasaki, Y.; Asahiyama, M.; Tanaka, T.; Yamamoto, S.; Murakami, K.; Kamiya, W.; Matsumura, Y.; Osawa, T.; Anai, M.; Fruchart, J.-C.; et al. Pemafibrate, a selective PPARα modulator, prevents non-alcoholic steatohepatitis development without reducing the hepatic triglyceride content. Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Giovannitti, J.A.; Trapp, L.D. Adult sedation: Oral, rectal, IM, IV. Anesth. Prog. 1991, 38, 154–171. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Author | Year of Publication | Journal | In Vitro | Effect | In Vivo | Effect |
|---|---|---|---|---|---|---|
| Cell Type | Experimental Model | |||||
| Fu et al. [54] | 2017 | Cell Reports | HRMEC | Promotes cell migration | OIR; VldlrKO; Laser-induced CNV | Suppresses NV via decreasing TNF-α expression |
| Fu et al. [55] | 2018 | Diabetes | 661 W | Inhibits oxidative stress-induced inflammation | STZ; Akita mouse | Rescues retinal morphology and function |
| Tomita and Ozawa et al. [30] | 2019 | IJMS | 661 W | Inhibits a HIF activity | - | - |
| Tomita et al. [56] | 2020 | IJMS | HRMEC | Prevents vascular permeability | mVEGF164-induced retinal vascular leakage mouse | Preserves an expression of tight junction protein |
| Tomita and Lee et al. [31] | 2020 | IJMS | PC12D | Increases synaptophysin protein expression | - | - |
| Fu and Qiu et al. [57] | 2021 | iScience | rMC-1 | Increases SRF protein expression | P23H mutation mouse | Modulates retinal glial responses |
| Name | Direction | Sequence (5′ → 3′) | Accession Number |
|---|---|---|---|
| Hprt | Forward | TCAGTCAACGGGGGACATAAA | NM_013556.2 |
| Reverse | GGGGCTGTACTGCTTAACCAG | ||
| Epo | Forward | GGCCATAGAAGTTTGGCAAG | NM_007942 |
| Reverse | CCTCTCCCGTGTACAGCTTC | ||
| Bnip3 | Forward | GCTCCCAGACACCACAAGAT | NM_009760.4 |
| Reverse | TGAGAGTAGCTGTGCGCTTC | ||
| Vegfa | Forward | CCTGGTGGACATCTTCCAGGAGTACC | AY707864.1 |
| Reverse | GAAGCTCATCTCTCCTATGTGCTGGC | ||
| Glut1 | Forward | CAGTTCGGCTATAACACTGGTG | NM_011400.3 |
| Reverse | GCCCCCGACAGAGAAGATG | ||
| Ccl2 | Forward | CCCAATGAGTAGGCTGGAGA | NM_011333.3 |
| Reverse | TCTGGACCCATTCCTTCTTG | ||
| Ccl12 | Forward | GCTACAGGAGAATCACAAGCAGC | NM_011331.3 |
| Reverse | ACGTCTTATCCAAGTGGTTTATGG | ||
| Ucp3 | Forward | GGAGTCTCACCTGTTTACTGACAACT | NM_009464.3 |
| Reverse | GCACAGAAGCCAGCTCCAA | ||
| Fabp4 | Forward | CCGCAGACGACAGGA | NM_024406.3 |
| Reverse | CTCATGCCCTTTCATAAACT | ||
| Fgf21 | Forward | AACAGCCATTCACTTTGCCTGAGC | NM_020013.4 |
| Reverse | GGCAGCTGGAATTGTGTTCTGACT | ||
| Vldlr | Forward | GAGCCCCTGAAGGAATGCC | NM_001161420.1 |
| Reverse | CCTATAACTAGGTCTTTGCAGATATGG | ||
| Acox1 | Forward | TCTTCTTGAGACAGGGCCCAG | AF006688.1 |
| Reverse | GTTCCGACTAGCCAGGCATG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, D.; Tomita, Y.; Jeong, H.; Miwa, Y.; Tsubota, K.; Negishi, K.; Kurihara, T. Pemafibrate Prevents Retinal Dysfunction in a Mouse Model of Unilateral Common Carotid Artery Occlusion. Int. J. Mol. Sci. 2021, 22, 9408. https://doi.org/10.3390/ijms22179408
Lee D, Tomita Y, Jeong H, Miwa Y, Tsubota K, Negishi K, Kurihara T. Pemafibrate Prevents Retinal Dysfunction in a Mouse Model of Unilateral Common Carotid Artery Occlusion. International Journal of Molecular Sciences. 2021; 22(17):9408. https://doi.org/10.3390/ijms22179408
Chicago/Turabian StyleLee, Deokho, Yohei Tomita, Heonuk Jeong, Yukihiro Miwa, Kazuo Tsubota, Kazuno Negishi, and Toshihide Kurihara. 2021. "Pemafibrate Prevents Retinal Dysfunction in a Mouse Model of Unilateral Common Carotid Artery Occlusion" International Journal of Molecular Sciences 22, no. 17: 9408. https://doi.org/10.3390/ijms22179408
APA StyleLee, D., Tomita, Y., Jeong, H., Miwa, Y., Tsubota, K., Negishi, K., & Kurihara, T. (2021). Pemafibrate Prevents Retinal Dysfunction in a Mouse Model of Unilateral Common Carotid Artery Occlusion. International Journal of Molecular Sciences, 22(17), 9408. https://doi.org/10.3390/ijms22179408