
Multilevel Approach for the Treatment of Giardiasis by Targeting Arginine Deiminase

 ,
,  ,
,  , , , ,
, , , ,  and
and 
Abstract
:1. Introduction
2. Results
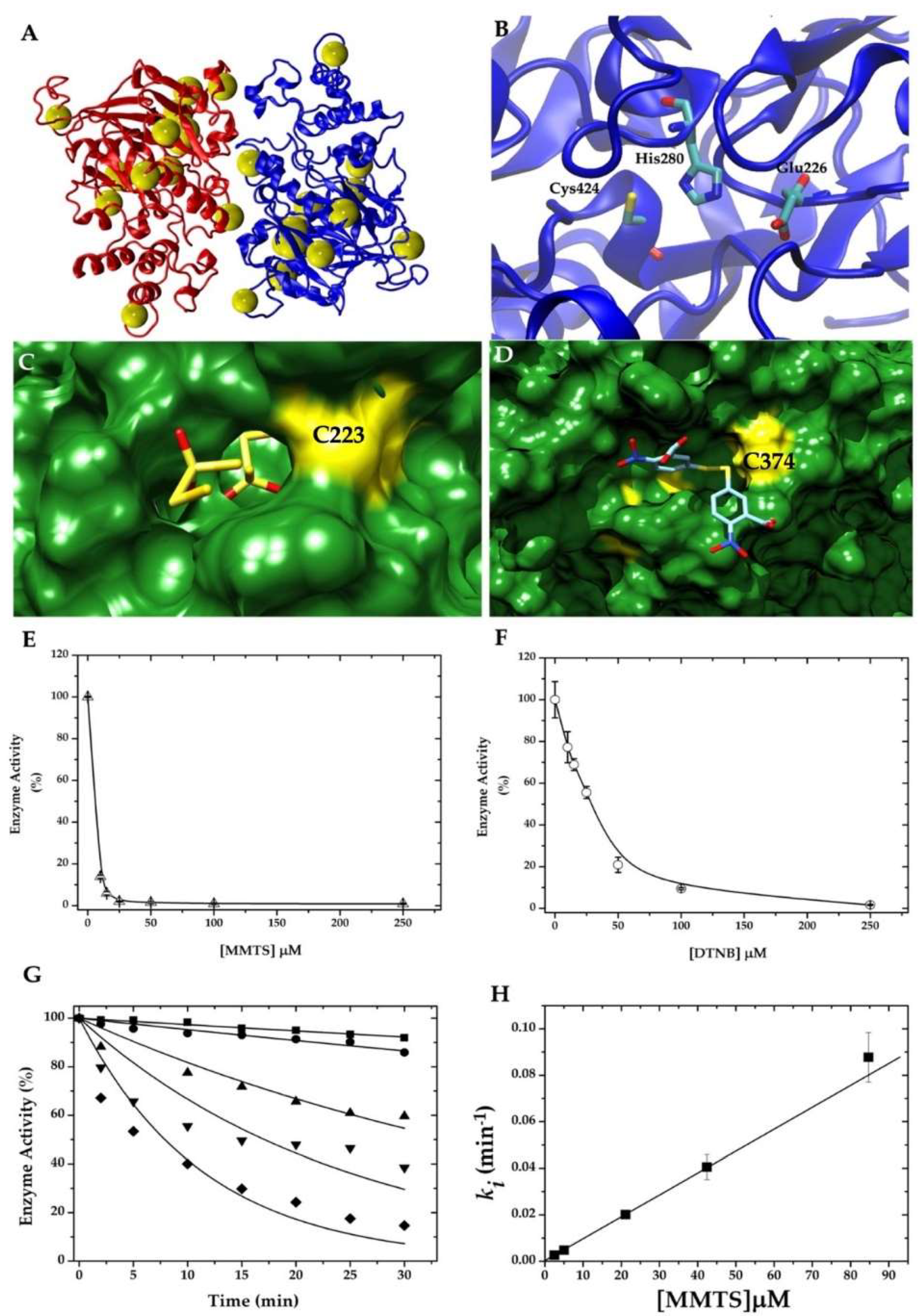
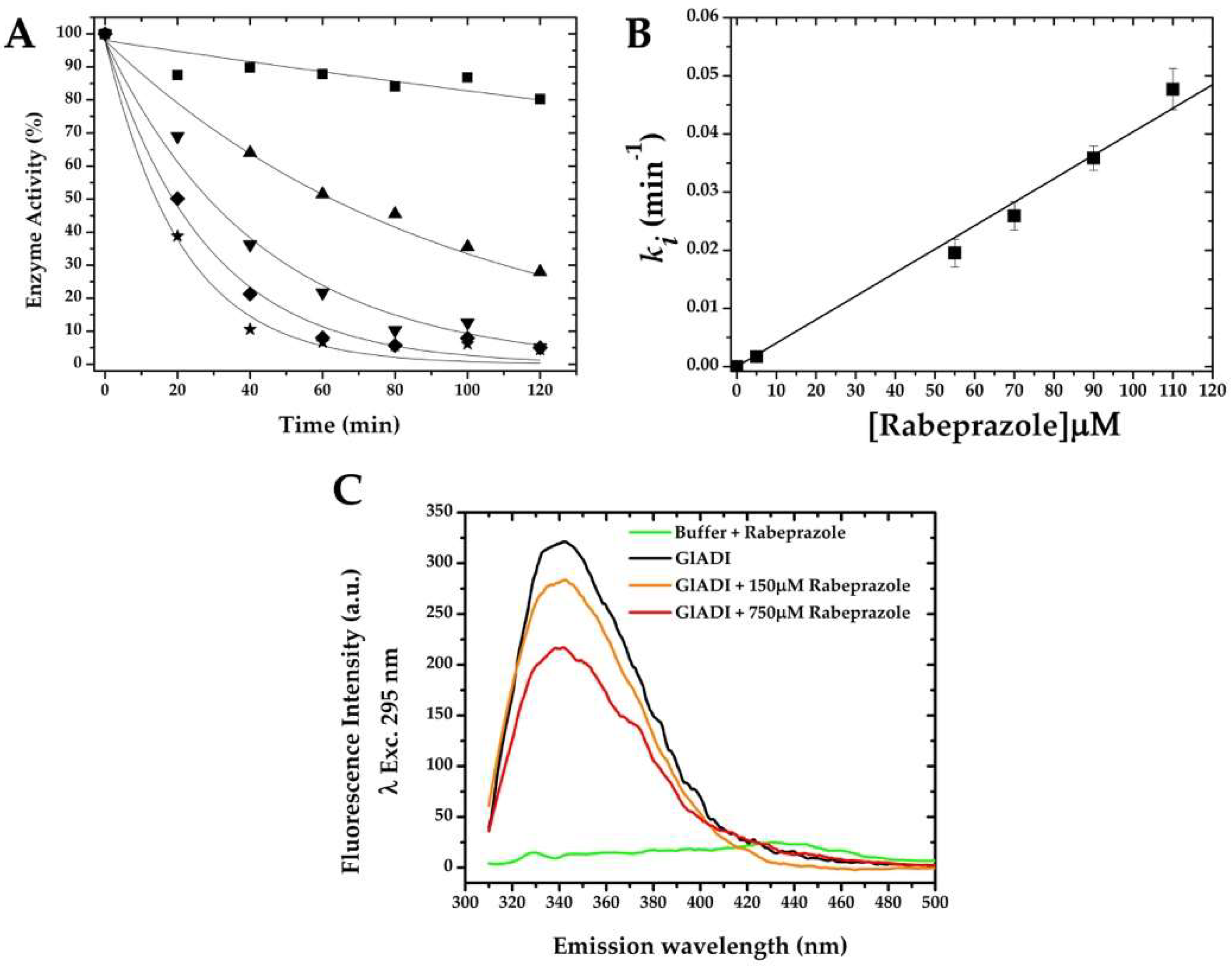
2.1. Cysteine Modification Underlies GlADI Inactivation
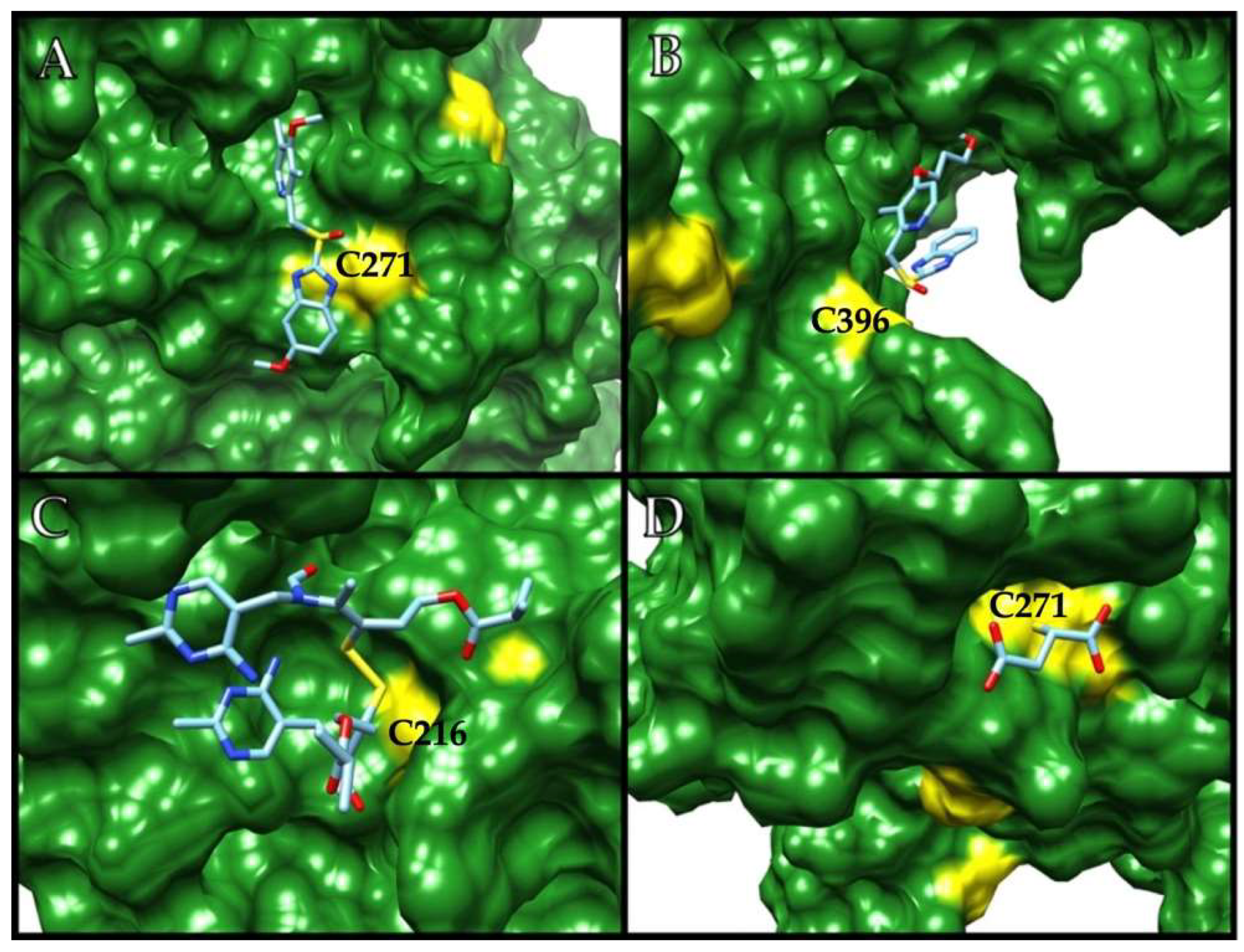
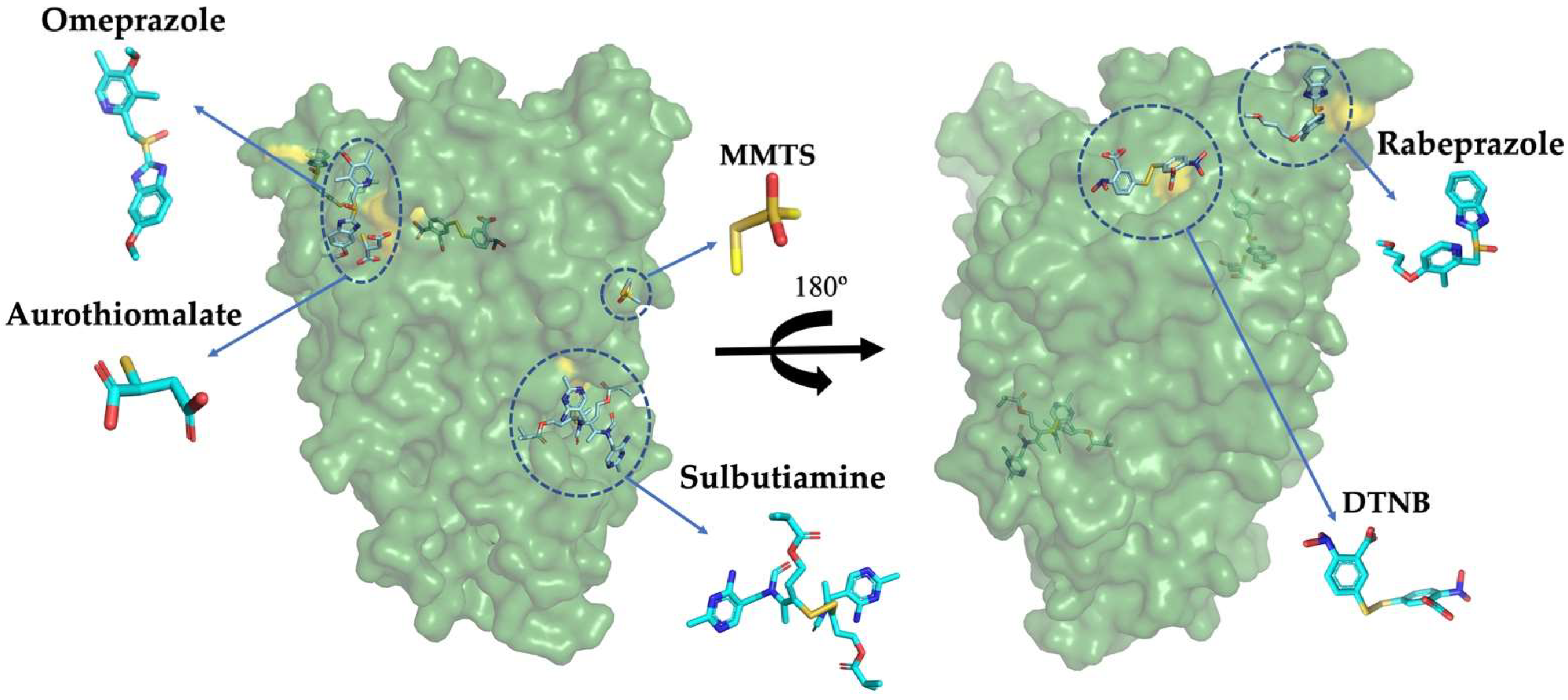
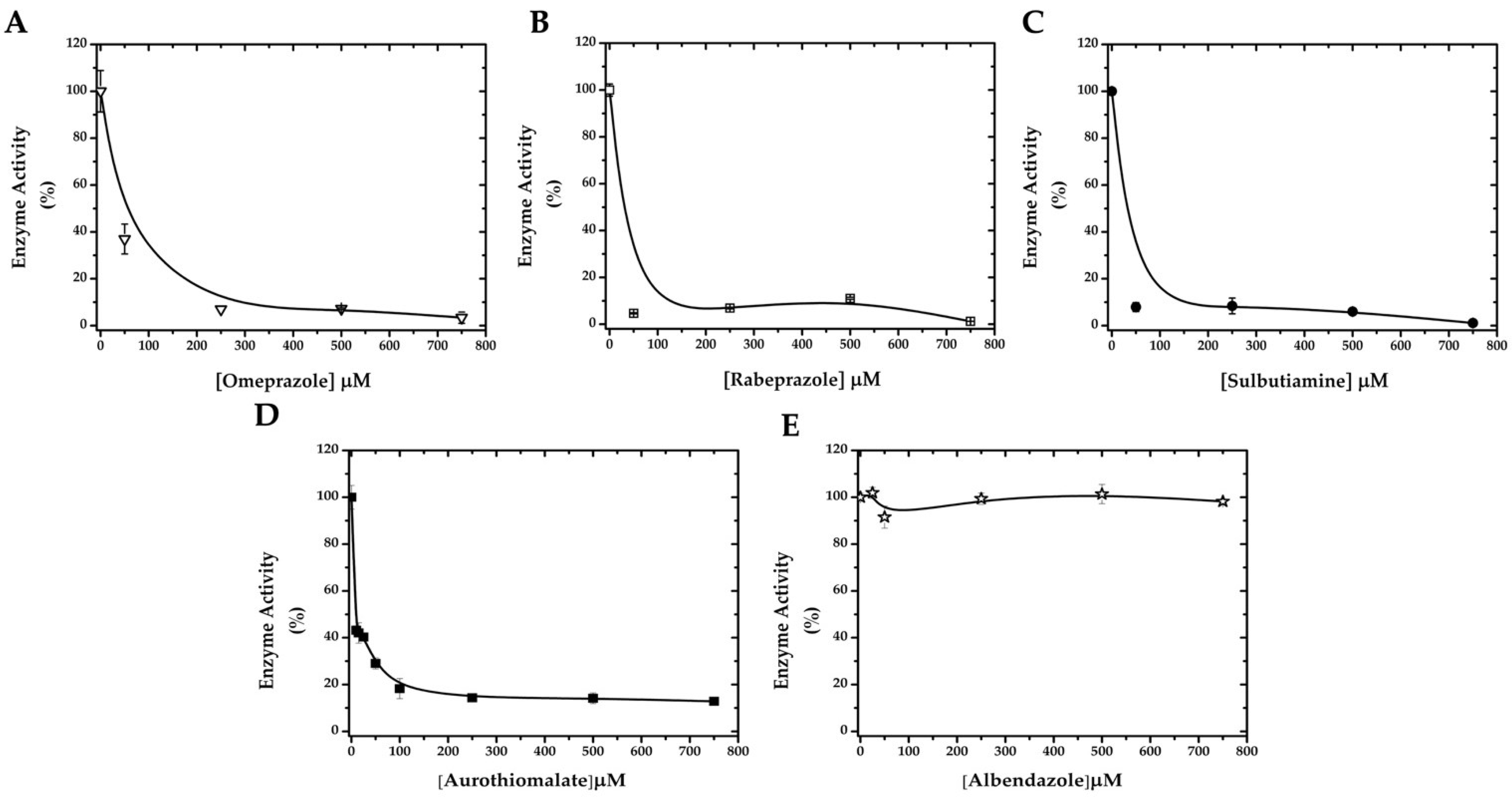
2.2. GlADI Is Inactivated by Thiol-Reactive Drugs Not Previously Applied for Giardiasis
2.3. Targetable Cysteines in GlADI Are Possibly Limited by the Presence of Disulfide Bridges in the Native Structure
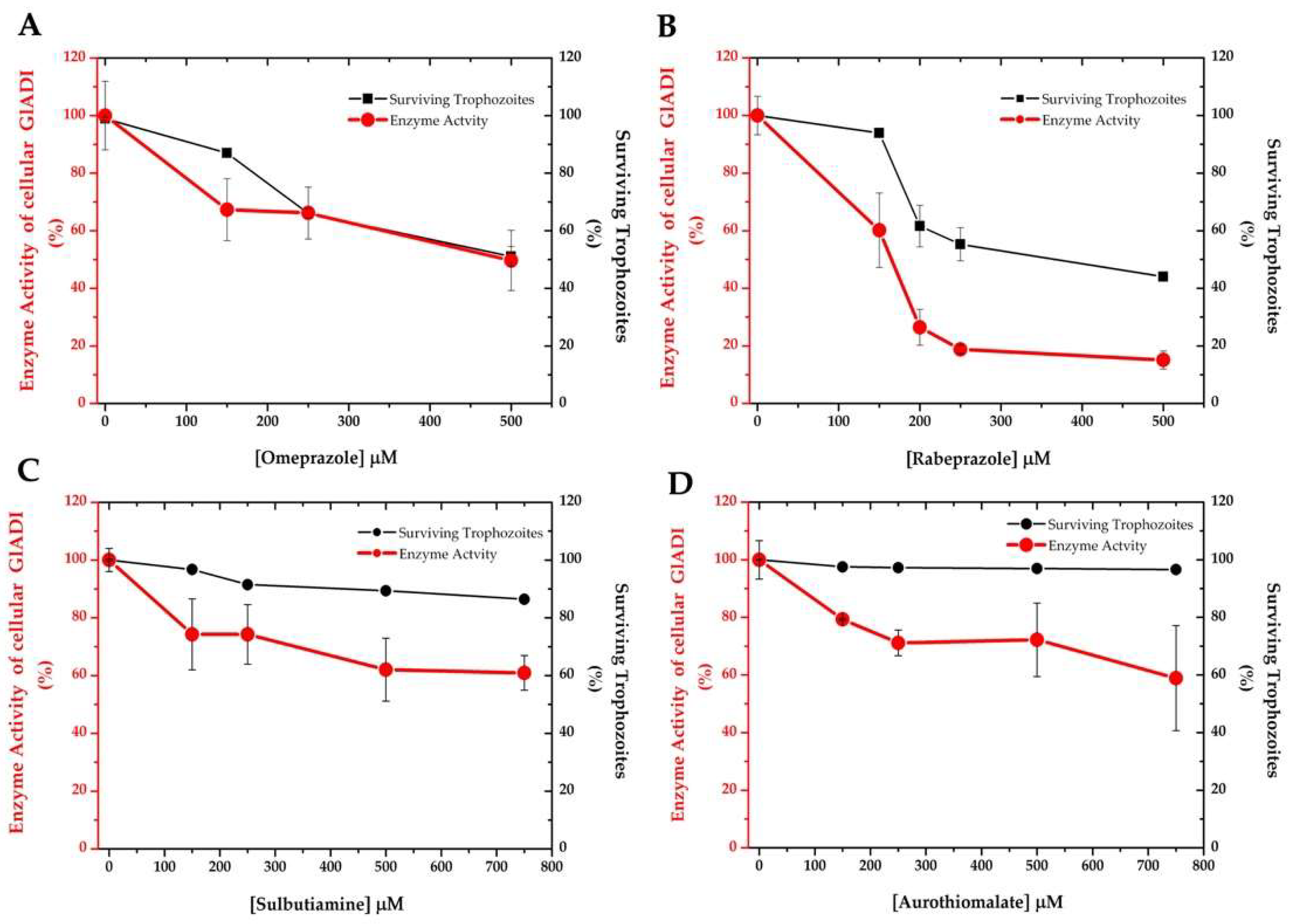
2.4. Drug Susceptibility of Giardia Trophozoites Discloses the Efficacy of the Proposed Drugs
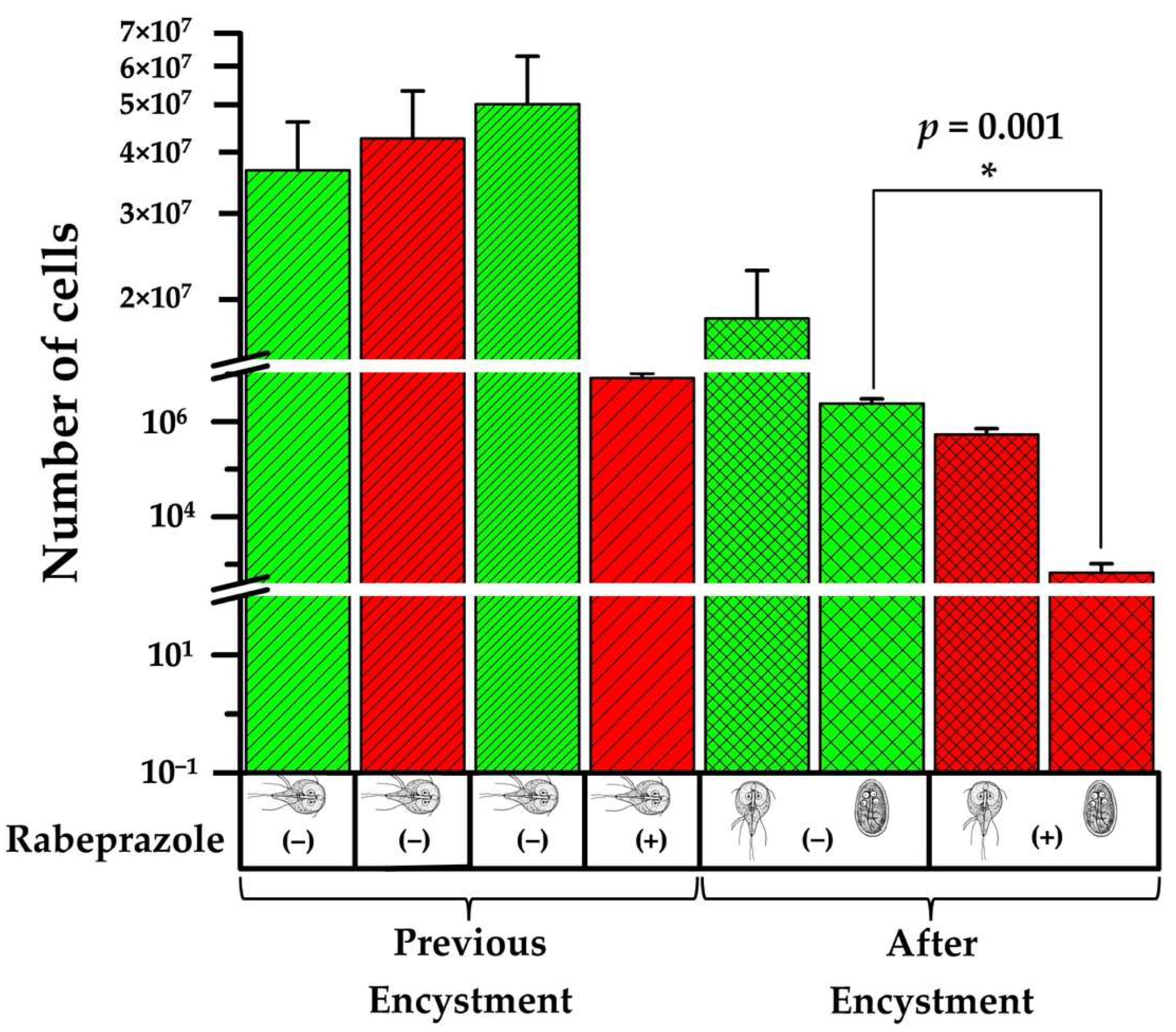
2.5. Rabeprazole Impairs Giardia Encystment
3. Discussion
4. Materials and Methods
4.1. Reagents and Drugs
4.2. Homology Modeling
4.3. Docking
4.4. Expression and Purification of Recombinant GlADI
4.5. Enzyme Activity Assays
4.6. Enzyme Inactivation Assays of Recombinant GlADI and Second-Order Inactivation Constants
4.7. Fluorescence Emission Spectra
4.8. Quantification of Free Cysteines after Drug Treatments
4.9. Drug Susceptibility of Trophozoites and Cellular GlADI Activity Assays
4.10. Induction to Encystment in the Presence of Rabeprazole
4.11. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| DTNB | Dithio-bis-nitrobenzoic acid |
| MMTS | S-methyl methanethiosulfonate |
| GlADI | Arginine deiminase from G. lamblia |
| PPI | Proton pump inhibitor |
| ATP | Adenosine triphosphate |
| ASA | Accessible surface area |
| Cys | Cysteine |
| a.u. | Arbitrary units |
| ki | Pseudo-first-order inactivation rate constant |
| k2 | Second-order rate constant |
| TCEP | Tris(2-carboxyethyl)phosphine |
| SD | Standard deviation |
| SDS | Sodium dodecyl sulfate |
| ATCC | American Type Culture Collection |
| PBS | Phosphate-buffered saline |
References
- Gavinho, B.; Rossi, I.V.; Evans-Osses, I.; Lange, S.; Ramirez, M.I. Peptidylarginine deiminase inhibition abolishes the production of large extracellular vesicles from Giardia intestinalis, affecting host-pathogen interactions by hindering adhesion to host cells. Biorxiv 2019, 10, 586438. [Google Scholar] [CrossRef]
- Lagunas-Rangel, F.A.; Bermúdez-Cruz, R.M. Epigenetics in the early divergent eukaryotic Giardia duodenalis: An update. Biochimie 2019, 156, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Lindmark, D.G. Energy metabolism of the anaerobic protozoon Giardia lamblia. Mol. Biochem. Parasitol. 1980, 1, 1–12. [Google Scholar] [CrossRef]
- Schofield, P.; Costello, M.; Edwards, M.; O’sullivan, W. The arginine dihydrolase pathway is present in Giardia intestinalis. Int. J. Parasitol. 1990, 20, 697–699. [Google Scholar] [CrossRef]
- Schofield, P.J.; Edwards, M.R.; Matthews, J.; Wilson, J.R. The pathway of arginine catabolism in Giardia intestinalis. Mol. Biochem. Parasitol. 1992, 51, 29–36. [Google Scholar] [CrossRef]
- Adam, R.D. Biology of Giardia lamblia. Clin. Microbiol. Rev. 2001, 14, 447–475. [Google Scholar] [CrossRef]
- Han, J.; Collins, L.J. Reconstruction of sugar metabolic pathways of Giardia lamblia. Int. J. Proteom. 2012, 2012, 1–9. [Google Scholar] [CrossRef]
- Mendis, A.; Thompson, R.; Reynoldson, J.; Armson, A.; Meloni, B.; Gunsberg, S. The uptake and conversion of l-[U14C-] aspartate and l-[U14C-] alanine to 14CO2 by intact trophozoites of Giardia duodenalis. Comp. Biochem. Physiol. B Comp. Biochem. 1992, 102, 235–239. [Google Scholar] [CrossRef]
- García-Torres, I.; Marcial-Quino, J.; Gómez-Manzo, S.; Vanoye-Carlo, A.; Navarrete-Vázquez, G.; Colín-Lozano, B.; Gutiérrez-Castrellón, P.; Sierra-Palacios, E.; López-Velázquez, G.; Enríquez-Flores, S. Proton pump inhibitors drastically modify triosephosphate isomerase from Giardia lamblia at functional and structural levels, providing molecular leads in the design of new antigiardiasic drugs. Biochim. Et Biophys. Acta (BBA)-Gen. Subj. 2016, 1860, 97–107. [Google Scholar] [CrossRef]
- López-Velázquez, G.; Fernández-Lainez, C.; de la Mora-de, J.I.; de la Portilla, D.C.; Reynoso-Robles, R.; González-Maciel, A.; Ridaura, C.; García-Torres, I.; Gutiérrez-Castrellón, P.; Olivos-García, A. On the molecular and cellular effects of omeprazole to further support its effectiveness as an antigiardial drug. Sci. Rep. 2019, 9, 1–14. [Google Scholar]
- Reyes-Vivas, H.; Castillo-Villanueva, A.; Yépez-Mulia, L.; Hernández-Alcántara, G.; Figueroa-Salazar, R.; García-Torres, I.; Gómez-Manzo, S.; Méndez, S.T.; Vanoye-Carlo, A.; Marcial-Quino, J. Giardial Triosephosphate Isomerase as possible target of the cytotoxic effect of omeprazole in Giardia. Antimicrob. Agents Chemother. 2014, 58, 7072–7082. [Google Scholar] [CrossRef] [PubMed]
- Emery-Corbin, S.J.; Grüttner, J.; Svärd, S. Transcriptomic and proteomic analyses of Giardia intestinalis: Intestinal epithelial cell interactions. Adv. Parasitol. 2020, 107, 139–171. [Google Scholar] [CrossRef] [PubMed]
- Lalle, M.; Hanevik, K. Treatment-refractory giardiasis: Challenges and solutions. Infect. Drug Resist. 2018, 11, 1921. [Google Scholar] [CrossRef]
- Pérez-Villanueva, J.; Romo-Mancillas, A.; Hernández-Campos, A.; Yépez-Mulia, L.; Hernández-Luis, F.; Castillo, R. Antiprotozoal activity of proton-pump inhibitors. Bioorg. Med. Chem. Lett. 2011, 21, 7351–7354. [Google Scholar] [CrossRef]
- Delgado-Maldonado, T.; Nogueda-Torres, B.; Espinoza-Hicks, J.C.; Vázquez-Jiménez, L.K.; Paz-González, A.D.; Juárez-Saldívar, A.; Rivera, G. Synthesis and biological evaluation in vitro and in silico of N-propionyl-N′-benzeneacylhydrazone derivatives as cruzain inhibitors of Trypanosoma cruzi. Mol. Divers. 2020, 24, 1–12. [Google Scholar]
- Leitsch, D.; Müller, J.; Müller, N. Evaluation of Giardia lamblia thioredoxin reductase as drug activating enzyme and as drug target. Int. J. Parasitol. Drugs Drug Resist. 2016, 6, 148–153. [Google Scholar] [CrossRef]
- Moreno-Herrera, A.; Cortez-Maya, S.; Bocanegra-Garcia, V.; Banik, B.K.; Rivera, G. Recent advances in the development of broad-spectrum antiprotozoal agents. Curr. Med. Chem. 2021, 28, 583–606. [Google Scholar] [CrossRef]
- Trejo-Soto, P.J.; Aguayo-Ortiz, R.; Yépez-Mulia, L.; Hernández-Campos, A.; Medina-Franco, J.L.; Castillo, R. Insights into the structure and inhibition of Giardia intestinalis arginine deiminase: Homology modeling, docking, and molecular dynamics studies. J. Biomol. Struct. Dyn. 2016, 34, 732–748. [Google Scholar] [CrossRef]
- Li, Z.; Kulakova, L.; Li, L.; Galkin, A.; Zhao, Z.; Nash, T.E.; Mariano, P.S.; Herzberg, O.; Dunaway-Mariano, D. Mechanisms of catalysis and inhibition operative in the arginine deiminase from the human pathogen Giardia lamblia. Bioorg. Chem. 2009, 37, 149–161. [Google Scholar] [CrossRef] [PubMed]
- Aguilar-Díaz, H.; Carrero, J.C.; Argüello-García, R.; Laclette, J.P.; Morales-Montor, J. Cyst and encystment in protozoan parasites: Optimal targets for new life-cycle interrupting strategies? Trends Parasitol. 2011, 27, 450–458. [Google Scholar] [CrossRef] [PubMed]
- Touz, M.C.; Rópolo, A.S.; Rivero, M.R.; Vranych, C.V.; Conrad, J.T.; Svard, S.G.; Nash, T.E. Arginine deiminase has multiple regulatory roles in the biology of Giardia lamblia. J. Cell Sci. 2008, 121, 2930–2938. [Google Scholar] [CrossRef]
- Riches, A.; Hart, C.J.S.; Trenholme, K.R.; Skinner-Adams, T.S. Anti-Giardia Drug Discovery: Current Status and Gut Feelings. J. Med. Chem. 2020, 63, 13330–13354. [Google Scholar] [CrossRef] [PubMed]
- Rópolo, A.S.; Feliziani, C.; Touz, M.C. Unusual proteins in Giardia duodenalis and their role in survival. Adv. Parasitol. 2019, 106, 1–50. [Google Scholar] [CrossRef]
- Hajduk, P.J.; Huth, J.R.; Tse, C. Predicting protein druggability. Drug Discov. Today 2005, 10, 1675–1682. [Google Scholar] [CrossRef]
- Papanastasiou, P.; Bruderer, T.; Li, Y.; Bommeli, C.; Köhler, P. Primary structure and biochemical properties of a variant-specific surface protein of Giardia. Mol. Biochem. Parasitol. 1997, 86, 13–27. [Google Scholar] [CrossRef]
- López-Velázquez, G.; Molina-Ortiz, D.; Cabrera, N.; Hernández-Alcántara, G.; Peon-Peralta, J.; Yépez-Mulia, L.; Pérez-Montfort, R.; Reyes-Vivas, H. An unusual triosephosphate isomerase from the early divergent eukaryote Giardia lamblia. Proteins 2004, 55, 824–834. [Google Scholar] [CrossRef]
- Reyes-Vivas, H.; Diaz, A.; Peon, J.; Mendoza-Hernandez, G.; Hernandez-Alcantara, G.; De la Mora-De la Mora, I.; Enriquez-Flores, S.; Dominguez-Ramirez, L.; Lopez-Velazquez, G. Disulfide bridges in the mesophilic triosephosphate isomerase from Giardia lamblia are related to oligomerization and activity. J. Mol. Biol. 2007, 365, 752–763. [Google Scholar] [CrossRef] [PubMed]
- Ladokhin, A.S. Fluorescence spectroscopy in peptide and protein analysis. Encycl. Anal. Chem. Appl. Theory Instrum. 2006. [Google Scholar]
- Hernández-Alcántara, G.; Rodríguez-Romero, A.; Reyes-Vivas, H.; Peon, J.; Cabrera, N.; Ortiz, C.; Enríquez-Flores, S.; López-Velázquez, G. Unraveling the mechanisms of tryptophan fluorescence quenching in the triosephosphate isomerase from Giardia lamblia. Biochim. Et Biophys. Acta (BBA)-Proteins Proteom. 2008, 1784, 1493–1500. [Google Scholar] [CrossRef]
- Tobinick, E.L. The value of drug repositioning in the current pharmaceutical market. Drug News Perspect. 2009, 22, 119–125. [Google Scholar] [CrossRef] [PubMed]
- Starling-Soares, B.; Carrera-Bastos, P.; Bettendorff, L. Role of the Synthetic B1 Vitamin Sulbutiamine on Health. J. Nutr. Metab. 2020, 2020, 9349063. [Google Scholar] [CrossRef] [PubMed]
- García-Torres, I.; De la Mora-De la Mora, I.; Hernández-Alcántara, G.; Molina-Ortiz, D.; Caballero-Salazar, S.; Olivos-García, A.; Nava, G.; López-Velázquez, G.; Enríquez-Flores, S. First characterization of a microsporidial triosephosphate isomerase and the biochemical mechanisms of its inactivation to propose a new druggable target. Sci. Rep. 2018, 8, 8591. [Google Scholar] [CrossRef] [PubMed]
- Wood, P.L.; Khan, M.A.; Moskal, J.R. Mechanism of action of the disease-modifying anti-arthritic thiol agents D-penicillamine and sodium aurothiomalate: Restoration of cellular free thiols and sequestration of reactive aldehydes. Eur. J. Pharmacol. 2008, 580, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Morrison, H.G.; McArthur, A.G.; Gillin, F.D.; Aley, S.B.; Adam, R.D.; Olsen, G.J.; Best, A.A.; Cande, W.Z.; Chen, F.; Cipriano, M.J. Genomic minimalism in the early diverging intestinal parasite Giardia lamblia. Science 2007, 317, 1921–1926. [Google Scholar] [CrossRef]
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The Protein Data Bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual molecular dynamics. J. Mol. Graph. 1996, 14, 33–38. [Google Scholar] [CrossRef]
- Chimera, U. A visualization system for exploratory research and analysis. Pettersen EF, Goddard TD, Huang CC, Couch GS, Greenblatt DM, Meng EC, Ferrin TE. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar]
- Muñoz-Cruz, S.; Gomez-García, A.; Matadamas-Martínez, F.; Alvarado-Torres, J.A.; Meza-Cervantez, P.; Arriaga-Pizano, L.; Yépez-Mulia, L. Giardia lamblia: Identification of molecules that contribute to direct mast cell activation. Parasitol. Res. 2018, 117, 2555–2567. [Google Scholar] [CrossRef]
- Pace, C.N.; Vajdos, F.; Fee, L.; Grimsley, G.; Gray, T. How to measure and predict the molar absorption coefficient of a protein. Protein Sci. 1995, 4, 2411–2423. [Google Scholar] [CrossRef] [PubMed]
- Weickmann, J.L.; Fahrney, D.E. Arginine deiminase from Mycoplasma arthritidis. Evidence for multiple forms. J. Biol. Chem. 1977, 252, 2615–2620. [Google Scholar] [CrossRef]
- Enríquez-Flores, S.; Rodríguez-Romero, A.; Hernández-Alcántara, G.; Oria-Hernández, J.; Gutiérrez-Castrellón, P.; Pérez-Hernández, G.; Mora, I.d.l.M.d.l.; Castillo-Villanueva, A.; García-Torres, I.; Mendez, S.T. Determining the molecular mechanism of inactivation by chemical modification of triosephosphate isomerase from the human parasite Giardia lamblia: A study for antiparasitic drug design. Proteins Struct. Funct. Bioinform. 2011, 79, 2711–2724. [Google Scholar] [CrossRef] [PubMed]
- Ellman, G.L. A colorimetric method for determining low concentrations of mercaptans. Arch. Biochem. Biophys. 1958, 74, 443–450. [Google Scholar] [CrossRef]
- Boucher, S.E.; Gillin, F.D. Excystation of in vitro-derived Giardia lamblia cysts. Infect. Immun. 1990, 58, 3516–3522. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cys Position | ASA (%) | Chain |
|---|---|---|
| 329 | 71.3/46.5 | A/B |
| 396 | 66.2/31.4 | A/B |
| 300 | 63.3/58.1 | A/B |
| 374 | 24.1/28.0 | A/B |
| 271 | 25.1 | A |
| 223 | 21.1 | B |
| 118 | 16.4/15.8 | A/B |
| 216 | 15.4 | B |
| 52 | 13.7 | A |
| Giardial Arginine Deiminase (GlADI) | Number of Modified Cys/Monomer |
|---|---|
| +DTNB (250 µM) | 6 ± 0.1 |
| +MMTS (250 µM) | 6 ± 0.3 |
| +Omeprazole (750 µM) | 2 ± 0.9 |
| +Rabeprazole (750 µM) | 5 ± 0.3 |
| +Sulbutiamine (750 µM) | 2 ± 0.9 |
| +Aurothiomalate (750 µM) | 5 ± 0.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernández-Lainez, C.; de la Mora-de la Mora, I.; García-Torres, I.; Enríquez-Flores, S.; Flores-López, L.A.; Gutiérrez-Castrellón, P.; Yépez-Mulia, L.; Matadamas-Martínez, F.; de Vos, P.; López-Velázquez, G. Multilevel Approach for the Treatment of Giardiasis by Targeting Arginine Deiminase. Int. J. Mol. Sci. 2021, 22, 9491. https://doi.org/10.3390/ijms22179491
Fernández-Lainez C, de la Mora-de la Mora I, García-Torres I, Enríquez-Flores S, Flores-López LA, Gutiérrez-Castrellón P, Yépez-Mulia L, Matadamas-Martínez F, de Vos P, López-Velázquez G. Multilevel Approach for the Treatment of Giardiasis by Targeting Arginine Deiminase. International Journal of Molecular Sciences. 2021; 22(17):9491. https://doi.org/10.3390/ijms22179491
Chicago/Turabian StyleFernández-Lainez, Cynthia, Ignacio de la Mora-de la Mora, Itzhel García-Torres, Sergio Enríquez-Flores, Luis A. Flores-López, Pedro Gutiérrez-Castrellón, Lilian Yépez-Mulia, Felix Matadamas-Martínez, Paul de Vos, and Gabriel López-Velázquez. 2021. "Multilevel Approach for the Treatment of Giardiasis by Targeting Arginine Deiminase" International Journal of Molecular Sciences 22, no. 17: 9491. https://doi.org/10.3390/ijms22179491
APA StyleFernández-Lainez, C., de la Mora-de la Mora, I., García-Torres, I., Enríquez-Flores, S., Flores-López, L. A., Gutiérrez-Castrellón, P., Yépez-Mulia, L., Matadamas-Martínez, F., de Vos, P., & López-Velázquez, G. (2021). Multilevel Approach for the Treatment of Giardiasis by Targeting Arginine Deiminase. International Journal of Molecular Sciences, 22(17), 9491. https://doi.org/10.3390/ijms22179491