
Dual Mkk4 and Mkk7 Gene Deletion in Adult Mouse Causes an Impairment of Hippocampal Immature Granule Cells

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Results
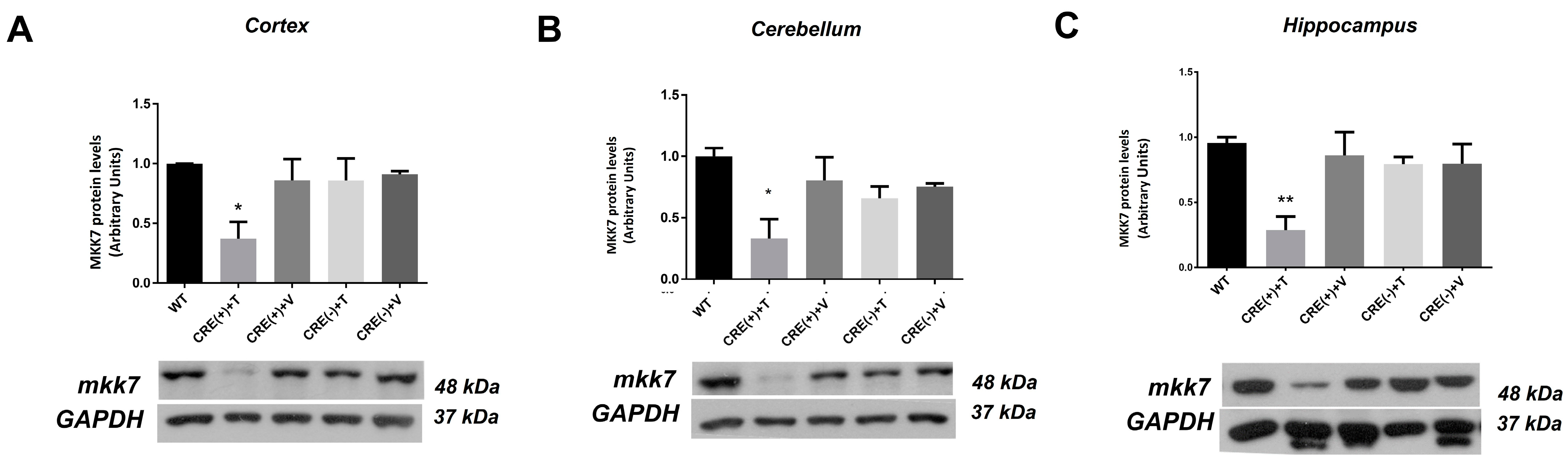
2.1. Characterization of Mkk4 and Mkk7 Gene Deletion and Protein Elimination in the Hippocampus of Actin-CreERT2 (Cre+/−), Mkk4 Δ/Δ, Mkk7Δ/Δ Adult Mice
2.2. Genetic Ablation of Mkk4 and Mkk7 Reduces Phosphorylation of JNK in the Hippocampus
2.3. Immature Hippocampal Neurons in Actin-CreERT2(Cre+/−), Mkk4∆/∆, Mkk7∆/∆ Mice Were Misaligned and Showed Alterations in the Dendritic Pattern
2.4. Reduction of MKK4 and MKK7 Proteins Alters the Late Differentiation of Immature Hippocampal Neurons
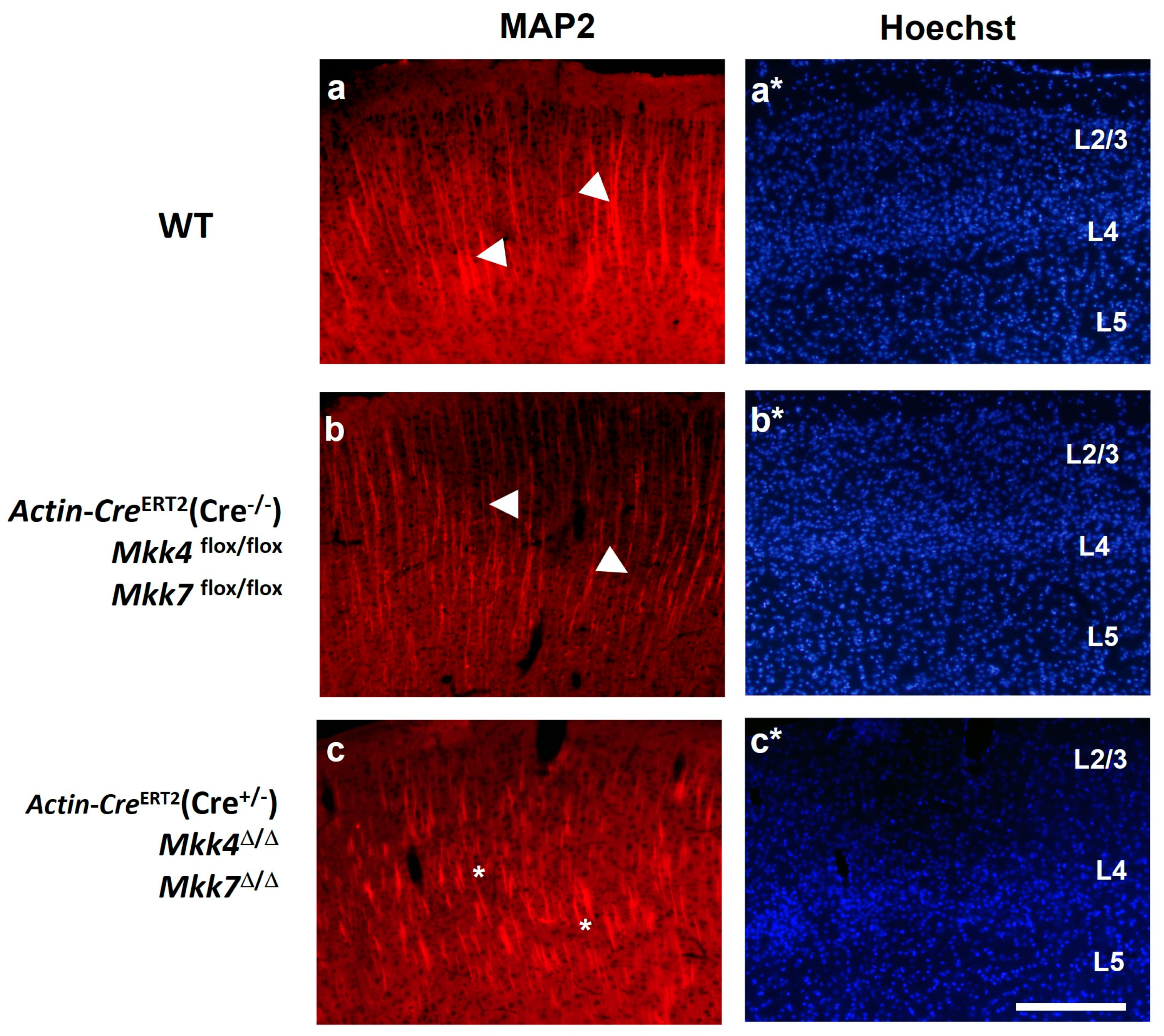
2.5. Neuronal Dendritic Pattern Is Disorganized in Actin-CreERT2(Cre+/−), Mkk4∆/∆, Mkk7∆/∆ Mice
3. Discussion
3.1. The Levels of MKK4 and MKK7 Are Not Equal in the Different Tissues
3.2. MKK7 Plays an Essential Role in Heart and Liver Tissues
3.3. The Levels of JNK Phosphorylation Were Decreased in Actin-CreERT2(Cre+/−), Mkk4∆/∆, Mkk7∆/∆ Mice
3.4. The Deletion of Mkk4 and Mkk7 Gene Alters Immature Hippocampal Neurons
4. Material and Methods
4.1. Animals
4.2. Tamoxifen Treatment
4.3. Genotype Determination in Mice and Deletion on Tissue
4.4. Preparation of Lysates
4.5. Immunoblot Analysis
4.6. Immunofluorescences
4.7. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dong, C.; Davis, R.J.; Flavell, R.A. MAP kinases in the immune response. Annu. Rev. Immunol. 2002, 20, 55–72. [Google Scholar] [CrossRef]
- Kim, E.K.; Choi, E.J. Compromised MAPK signaling in human diseases: An update. Arch. Toxicol. 2015, 89, 867–882. [Google Scholar] [CrossRef]
- Taylor, J.L.; Szmulewitz, R.Z.; Lotan, T.; Hickson, J.; Griend, D.V.; Yamada, S.D.; Macleod, K.; Rinker-Schaeffer, C.W. New paradigms for the function of JNKK1/MKK4 in controlling growth of disseminated cancer cells. Cancer Lett. 2008, 272, 12–22. [Google Scholar] [CrossRef]
- Hirosumi, J.; Tuncman, G.; Chang, L.; Görgün, C.Z.; Uysal, K.T.; Maeda, K.; Karin, M.; Hotamisligil, G.S. A central, role for JNK in obesity and insulin resistance. Nature 2002, 420, 333–336. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Su, B.; Sah, V.P.; Brown, J.H.; Han, J.; Chien, K.R. Cardiac hypertrophy induced by mitogen-activated protein kinase kinase 7, a specific activator for c-Jun NH2-terminal kinase in ventricular muscle cells. J. Biol. Chem. 1998, 273, 5423–5426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antoniou, X. The JNK signalling transduction pathway in the brain. Front. Biosci. 2012, 4, 2110–2120. [Google Scholar] [CrossRef]
- Wang, X.; Nadarajah, B.; Robinson, A.C.C.; McColl, B.W.W.; Jin, J.-W.; Dajas-Bailador, F.; Boot-Handford, R.P.P.; Tournier, C. Targeted deletion of the mitogen-activated protein kinase kinase 4 gene in the nervous system causes severe brain developmental defects and premature death. Mol. Cell. Biol. 2007, 27, 7935–7946. [Google Scholar] [CrossRef] [Green Version]
- Yamasaki, T.; Kawasaki, H.; Arakawa, S.; Shimizu, K.; Shimizu, S.; Reiner, O.; Okano, H.; Nishina, S.; Azuma, N.; Penninger, J.M.; et al. Stress-activated protein kinase MKK7 regulates axon elongation in the developing cerebral cortex. J. Neurosci. 2011, 31, 16872–16883. [Google Scholar] [CrossRef] [PubMed]
- Cazillis, M.; Bringuier, A.F.; Delautier, D.; Buisine, M.; Bernuau, D.; Gespach, C.; Groyer, A. Disruption of MKK4 signaling reveals its tumor-suppressor role in embryonic stem cells. Oncogene 2004, 23, 4735–4744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davies, C.; Tournier, C. Exploring the function of the JNK (c-Jun N-terminal kinase) signalling pathway in physiological and pathological processes to design novel therapeutic strategies. Biochem. Soc. Trans. 2012, 40, 85–89. [Google Scholar] [CrossRef]
- Kennedy, N.J.; Davis, R.J. Role of JNK in tumor development. Cell Cycle 2003, 2, 199–201. [Google Scholar] [PubMed] [Green Version]
- Aouadi, M.; Binetruy, B.; Caron, L.; Le Marchand-Brustel, Y.; Bost, F. Role of MAPKs in development and differentiation: Lessons from knockout mice. Biochimie 2006, 88, 1091–1098. [Google Scholar] [CrossRef]
- Kuan, C.-Y.; Whitmarsh, A.J.; Yang, D.D.; Liao, G.; Schloemer, A.J.; Dong, C.; Bao, J.; Banasiak, K.J.; Haddad, G.G.; Flavell, R.A.; et al. A critical role of neural-specific JNK3 for ischemic apoptosis. Proc. Natl. Acad. Sci. USA 2003, 100, 15184–15189. [Google Scholar] [CrossRef] [Green Version]
- De Lemos, L.; Junyent, F.; Camins, A.; Castro-Torres, R.D.; Folch, J.; Olloquequi, J.; Beas-Zarate, C.; Verdaguer, E.; Auladell, C. Neuroprotective Effects of the Absence of JNK1 or JNK3 Isoforms on Kainic Acid-Induced Temporal Lobe Epilepsy-Like Symptoms. Mol. Neurobiol. 2018, 55, 4437–4452. [Google Scholar] [CrossRef]
- Chang, L.; Jones, Y.; Ellisman, M.H.; Goldstein, L.S.B.; Karin, M. JNK1 is required for maintenance of neuronal microtubules and controls phosphorylation of microtubule-associated proteins. Dev. Cell 2003, 4, 521–533. [Google Scholar] [CrossRef] [Green Version]
- Komulainen, E.; Zdrojewska, J.; Freemantle, E.; Mohammad, H.; Kulesskaya, N.; Deshpande, P.; Marchisella, F.; Mysore, R.; Hollos, P.; Michelsen, K.A.; et al. JNK1 controls dendritic field size in L2/3 and l5 of the motor cortex, constrains soma size, and influences fine motor coordination. Front. Cell. Neurosci. 2014, 8, 272. [Google Scholar] [CrossRef] [Green Version]
- Castro-Torres, R.D.; Landa, J.; Rabaza, M.; Busquets, O.; Olloquequi, J.; Ettcheto, M.; Beas-Zarate, C.; Folch, J.; Camins, A.; Auladell, C.; et al. JNK isoforms are involved in the control of adult hippocampal neurogenesis in mice, both in physiological conditions and in an experimental model of temporal lobe epilepsy. Mol. Neurobiol. 2019, 56, 5856–5865. [Google Scholar] [CrossRef] [PubMed]
- Mohammad, H.; Marchisella, F.; Ortega-Martinez, S.; Hollos, P.; Eerola, K.; Komulainen, E.; Kulesskaya, N.; Freemantle, E.; Fagerholm, V.; Savontous, E.; et al. JNK1 controls adult hippocampal neurogenesis and imposes cell-autonomous control of anxiety behaviour from the neurogenic niche. Mol. Psychiatry 2018, 23, 362–374. [Google Scholar] [CrossRef] [PubMed]
- Asaoka, Y.; Nishina, H. Diverse physiological functions of MKK4 and MKK7 during early embryogenesis. J. Biochem. 2010, 148, 393–401. [Google Scholar] [CrossRef]
- Sabapathy, K.; Jochum, W.; Hochedlinger, K.; Chang, L.; Karin, M.; Wagner, E.F. Defective neural tube morphogenesis and altered apoptosis in the absence of both JNK1 and JNK2. Mech. Dev. 1999, 89, 115–124. [Google Scholar] [CrossRef]
- Nishina, H.; Vaz, C.; Billia, P.; Nghiem, M.; Sasaki, T.; De la Pompa, J.L.; Furlonger, K.; Paige, C.; Hui, C.C.; Fischer, K.D.; et al. Defective liver formation and liver cell apoptosis in mice lacking the stress signaling kinase SEK1/MKK4. Development 1999, 126, 505–516. [Google Scholar] [CrossRef] [PubMed]
- Kristianto, J.; Johnson, M.G.; Zastrow, R.K.; Radcliff, A.B.; Blank, R.D. Spontaneous recombinase activity of Cre–ERT2 in vivo. Transgenic Res. 2017, 26, 411–417. [Google Scholar] [CrossRef]
- Wang, X.; Destrument, A.; Tournier, C. Physiological roles of MKK4 and MKK7: Insights from animal models. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2007, 1773, 1349–1357. [Google Scholar] [CrossRef]
- Lee, J.K.; Hwang, W.S.; Lee, Y.D.; Han, P.L. Dynamic expression of SEK1 suggests multiple roles of the gene during embryogenesis and in adult brain of mice. Mol. Brain Res. 1999, 66, 133–140. [Google Scholar] [CrossRef]
- Kuan, C.Y.; Yang, D.D.; Roy, D.R.S.; Davis, R.J.; Rakic, P.; Flavell, R.A. The Jnk1 and Jnk2 protein kinases are required for regional specific apoptosis during early brain development. Neuron 1999, 22, 667–676. [Google Scholar] [CrossRef] [Green Version]
- Tournier, C.; Dong, C.; Turner, T.K.; Jones, S.N.; Flavell, R.A.; Davis, R.J. MKK7 is an essential component of the JNK signal transduction pathway activated by proinflammatory cytokines. Genes Dev. 2001, 15, 1419–1426. [Google Scholar] [CrossRef] [Green Version]
- Ooshio, T.; Yamamoto, M.; Fujii, K.; Xin, B.; Watanabe, K.; Goto, M.; Okada, Y.; Suzuki, A.; Penninger, J.M.; Nishina, H.; et al. Hepatocyte MKK7 contributes to restoration of the liver parenchyma following injury. Hepatology 2020, 73, 2510–2526. [Google Scholar] [CrossRef]
- Liu, W.; Zi, M.; Chi, H.; Jin, J.; Prehar, S.; Neyses, L.; Cartwright, E.J.; Flavell, R.A.; Davis, R.J.; Wang, X. Deprivation of MKK7 in cardiomyocytes provokes heart failure in mice when exposed to pressure overload. J. Mol. Cell. Cardiol. 2011, 50, 702–711. [Google Scholar] [CrossRef]
- Win, S.; Than, T.A.; Zhang, J.; Oo, C.; Min, R.W.M.; Kaplowitz, N. New insights into the role and mechanism of c-Jun-N-terminal kinase signaling in the pathobiology of liver diseases. Hepatology 2018, 67, 2013–2024. [Google Scholar] [CrossRef] [Green Version]
- Seki, T.; Namba, T.; Mochizuki, H.; Onodera, M. Clustering, migration, and neurite formation of neural precursor cells in the adult rat hippocampus. J. Comp. Neurol. 2007, 502, 275–290. [Google Scholar] [CrossRef]
- Sun, G.J.; Zhou, Y.; Stadel, R.P.; Moss, J.; Yong, J.H.A.; Ito, S.; Kawasaki, N.K.; Phan, A.T.; Oh, J.H.; Modak, N.; et al. Tangential migration of neuronal precursors of glutamatergic neurons in the adult mammalian brain. Proc. Natl. Acad. Sci. USA 2015, 112, 9484–9489. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Shen, J.; Kirschen, G.W.; Gu, Y.; Jessberger, S.; Ge, S. Lateral dispersion is required for circuit integration of newly generated dentate granule cells. Nat. Commun. 2019, 10, 3324. [Google Scholar] [CrossRef]
- Smith, S.E.; Coker, N.K.; Tucker, E.S. Jnk signaling regulates cellular mechanics of cortical interneuron migration. eNeuro 2020, 7, 1–21. [Google Scholar] [CrossRef]
- Yamasaki, T.; Deki-Arima, N.; Kaneko, A.; Miyamura, N.; Iwatsuki, M.; Matsuoka, M.; Fujimori-Tonou, N.; Okamoto-Uchida, Y.; Hirayama, J.; Marth, J.D.; et al. Age-dependent motor dysfunction due to neuron-specific disruption of stress-Activated protein kinase MKK7. Sci. Rep. 2017, 7, 7348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castro-Torres, R.D.; Busquets, O.; Parcerisas, A.; Verdaguer, E.; Olloquequi, J.; Etchetto, M.; Beas-Zarate, C.; Folch, J.; Camins, A.; Auladell, C. Involvement of JNK1 in neuronal polarization during brain development. Cells 2020, 9, 1879. [Google Scholar] [CrossRef] [PubMed]
- Bjorkblom, B.; Ostman, N.; Hongisto, V.; Komarovski, V.; Filen, J.-J.; Nyman, T.A.; Kallunki, T.; Courtney, M.J.; Coffey, E.T. Constitutively active cytoplasmic c-Jun N-terminal kinase 1 is a dominant regulator of dendritic architecture: Role of microtubule-associated protein 2 as an effector. J. Neurosci. 2005, 25, 6350–6361. [Google Scholar] [CrossRef]
- Tararuk, T.; Östman, N.; Li, W.; Björkblom, B.; Padzik, A.; Zdrojewska, J.; Hongisto, V.; Herdegen, T.; Konopka, W.; Courtney, M.J.; et al. JNK1 phosphorylation of SCG10 determines microtubule dynamics and axodendritic length. J. Cell Biol. 2006, 173, 265–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komulainen, E.; Varidaki, A.; Kulesskaya, N.; Mohammad, H.; Sourander, C.; Rauvala, H.; Coffey, E.T. Impact of JNK and its substrates on dendritic spine morphology. Cells 2020, 9, 440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGill, B.E.; Barve, R.A.; Maloney, S.E.; Strickland, A.; Rensing, N.; Wang, P.L.; Wong, M.; Head, R.; Wozniak, D.F.; Milbrandt, J. Abnormal microglia and enhanced inflammation-related gene transcription in mice with conditional deletion of Ctcf in Camk2a-Cre-expressing neurons. J. Neurosci. 2018, 38, 200–219. [Google Scholar] [CrossRef] [Green Version]
- Dragatsis, I.; Zeitlin, S. CaMKIIalpha-Cre transgene expression and recombination patterns in the mouse brain. Genesis 2000, 26, 133–135. [Google Scholar] [CrossRef]
- Rempe, D.; Vangeison, G.; Hamilton, J.; Li, Y.; Jepson, M.; Federoff, H.J. Synapsin I Cre transgene expression in male mice produces germline recombination in progeny. Genesis 2006, 44, 44–49. [Google Scholar] [CrossRef] [PubMed]
- Ganat, Y.M.; Silbereis, J.; Cave, C.; Ngu, H.; Anderson, G.M.; Ohkubo, Y.; Ment, L.R.; Vaccarino, F.M. Early postnatal astroglial cells produce multilineage precursors and neural stem cells In Vivo. J. Neurosci. 2006, 26, 8609–8621. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Wang, X.; Xiao, J.; Wang, Y.; Lu, H.; Teng, J.; Wang, W. Early postnatal GFAP-expressing cells produce multilineage progeny in cerebrum and astrocytes in cerebellum of adult mice. Brain Res. 2013, 1532, 14–20. [Google Scholar] [CrossRef]
- Schramek, D.; Kotsinas, A.; Meixner, A.; Wada, T.; Elling, U.; Pospisilik, J.A.; Neely, G.G.; Zwick, R.H.; Sigl, V.; Forni, G.; et al. The stress kinase MKK7 couples oncogenic stress to p53 stability and tumor suppression. Nat. Genet. 2011, 43, 212–219. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Kim, M.; Im, S.-K.; Fang, S. Mouse Cre-LoxP system: General principles to determine tissue-specific roles of target genes. Lab. Anim. Res. 2018, 34, 147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paxinos, G.; Watson, C.; Diego, S.; Boston, L.; York, N. The Rat Brain in Stereotaxic Coordinates; Academic Press: Cambridge, MA, USA, 2006. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype | Treatment | Purpose: | Group Name |
|---|---|---|---|
| HEMIZYGOUS Actin-CreERT2(Cre+/−), Mkk4flox/flox, Mkk7flox/flox | Tamoxifen | Mkk4∆/∆, Mkk7∆/∆ genotype | CRE (+) +T |
| HEMIZYGOUS Actin-CreERT2(Cre+/−), Mkk4flox/flox, Mkk7flox/flox | Vehicle | Control | CRE (+) +V |
| NEGATIVE Actin-CreERT2(Cre−/−), Mkk4flox/flox, Mkk7flox/flox | Tamoxifen | Control | CRE (−) +T |
| NEGATIVE Actin-CreERT2(Cre−/−), Mkk4flox/flox, Mkk7flox/flox | Vehicle | Control | CRE (−) +V |
| NEGATIVE Actin-CreERT2(Cre−/−), Mkk4flox/flox, Mkk7flox/flox | None | Control | CRE (−) |
| C57BL/6 | None | Control | WT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castro-Torres, R.D.; Olloquequi, J.; Etchetto, M.; Caruana, P.; Steele, L.; Leighton, K.-M.; Ureña, J.; Beas-Zarate, C.; Camins, A.; Verdaguer, E.; et al. Dual Mkk4 and Mkk7 Gene Deletion in Adult Mouse Causes an Impairment of Hippocampal Immature Granule Cells. Int. J. Mol. Sci. 2021, 22, 9545. https://doi.org/10.3390/ijms22179545
Castro-Torres RD, Olloquequi J, Etchetto M, Caruana P, Steele L, Leighton K-M, Ureña J, Beas-Zarate C, Camins A, Verdaguer E, et al. Dual Mkk4 and Mkk7 Gene Deletion in Adult Mouse Causes an Impairment of Hippocampal Immature Granule Cells. International Journal of Molecular Sciences. 2021; 22(17):9545. https://doi.org/10.3390/ijms22179545
Chicago/Turabian StyleCastro-Torres, Rubén Darío, Jordi Olloquequi, Miren Etchetto, Pablo Caruana, Luke Steele, Kyra-Mae Leighton, Jesús Ureña, Carlos Beas-Zarate, Antoni Camins, Ester Verdaguer, and et al. 2021. "Dual Mkk4 and Mkk7 Gene Deletion in Adult Mouse Causes an Impairment of Hippocampal Immature Granule Cells" International Journal of Molecular Sciences 22, no. 17: 9545. https://doi.org/10.3390/ijms22179545
APA StyleCastro-Torres, R. D., Olloquequi, J., Etchetto, M., Caruana, P., Steele, L., Leighton, K. -M., Ureña, J., Beas-Zarate, C., Camins, A., Verdaguer, E., & Auladell, C. (2021). Dual Mkk4 and Mkk7 Gene Deletion in Adult Mouse Causes an Impairment of Hippocampal Immature Granule Cells. International Journal of Molecular Sciences, 22(17), 9545. https://doi.org/10.3390/ijms22179545