
Cancer-Nano-Interaction: From Cellular Uptake to Mechanobiological Responses



Abstract
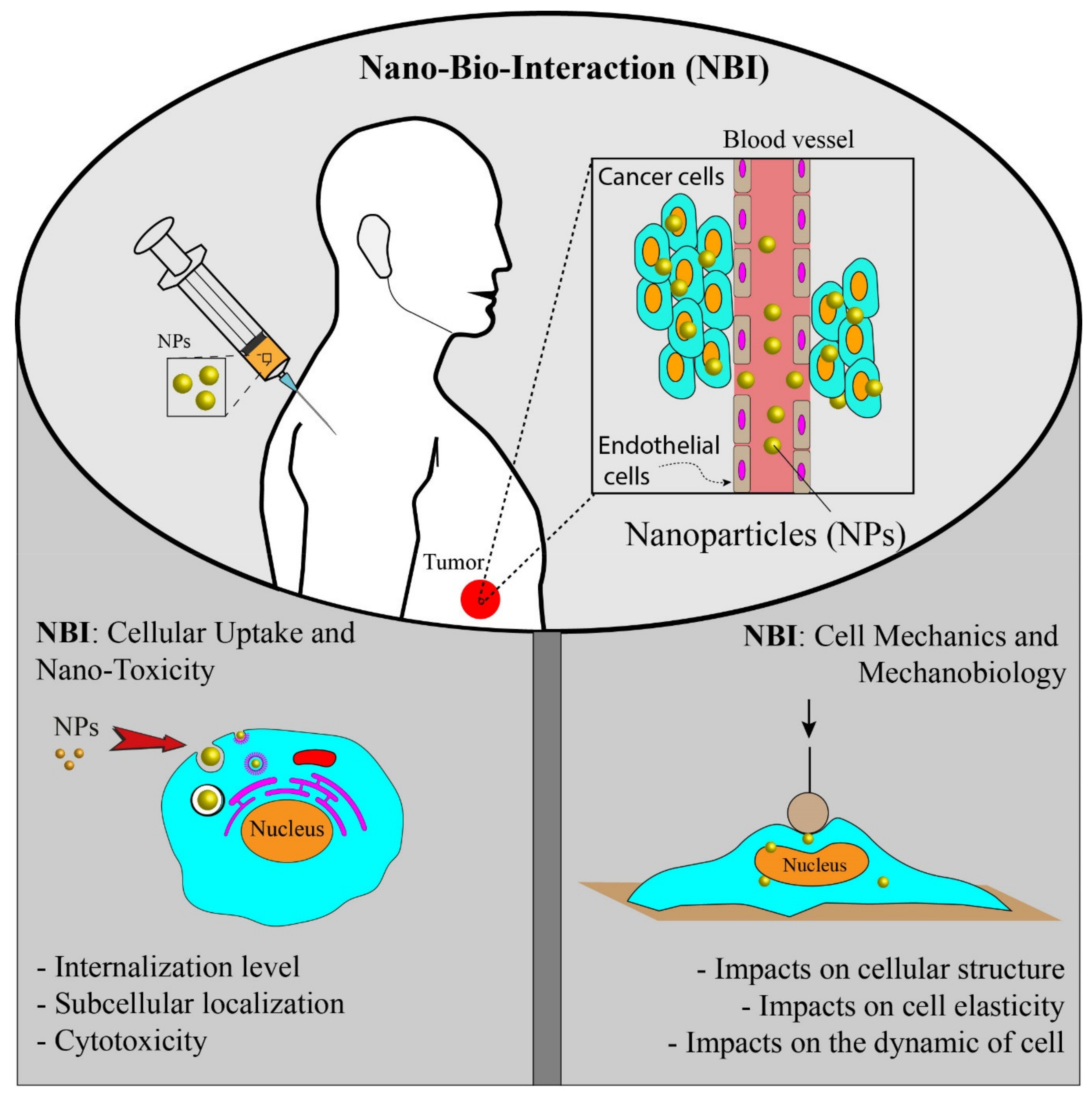
:1. Introduction to Cancer-Nano-Interactions
2. Nano-Bio-Interaction: Cellular Uptake and Toxicity
2.1. Effects of NPs’ Size
2.2. Effects of NPs Shape
2.3. Effects of NPs Surface Charge and Coating
3. Nano-Bio-Interactions: Cell Mechanics and Mechanobiology
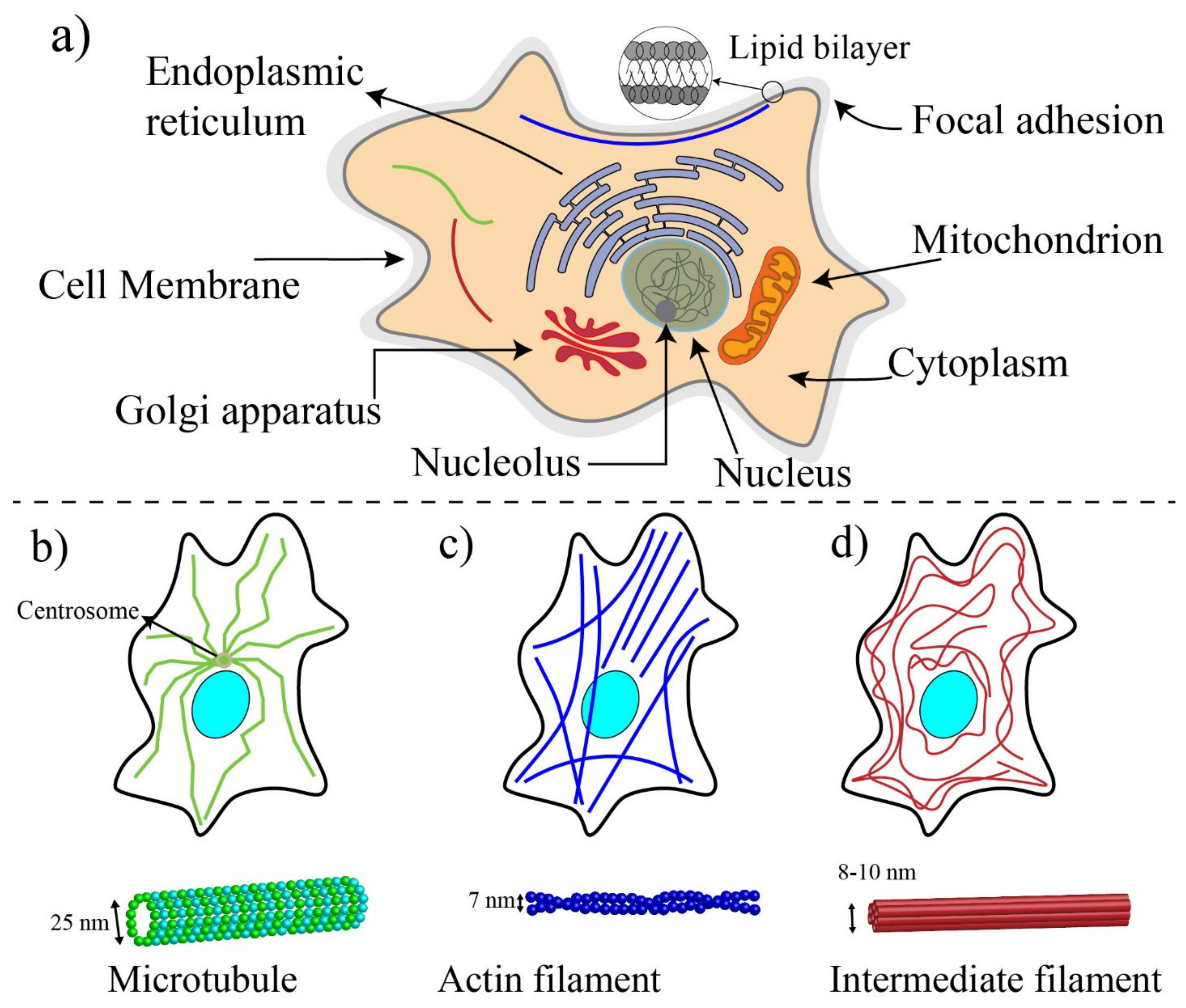
3.1. Basic Components of Cells and Biomechanics
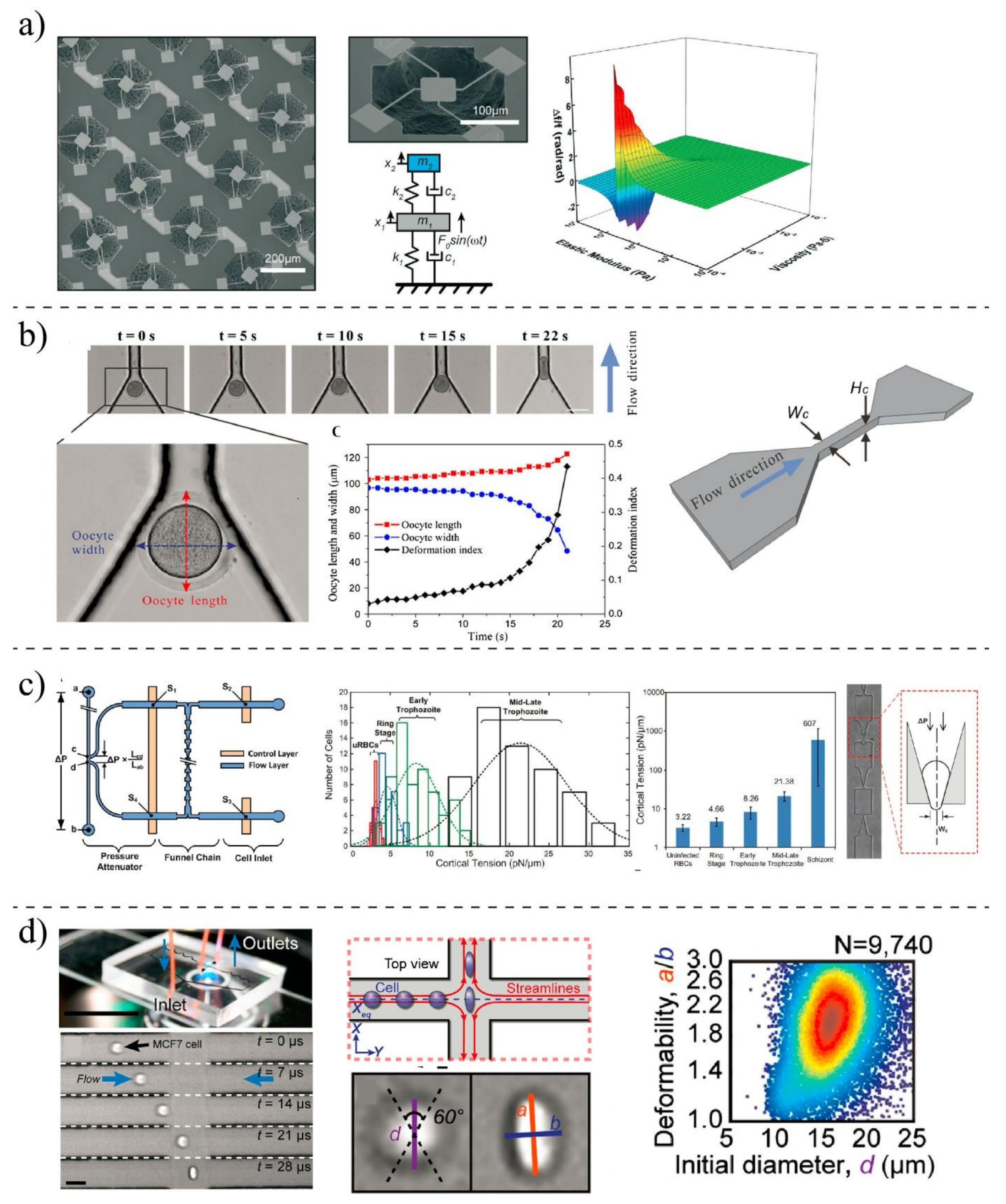
3.2. Techniques for Mechanobiological Characterizations
3.2.1. Classical Methods
3.2.2. MEMS- and Microfluidic-Based Techniques
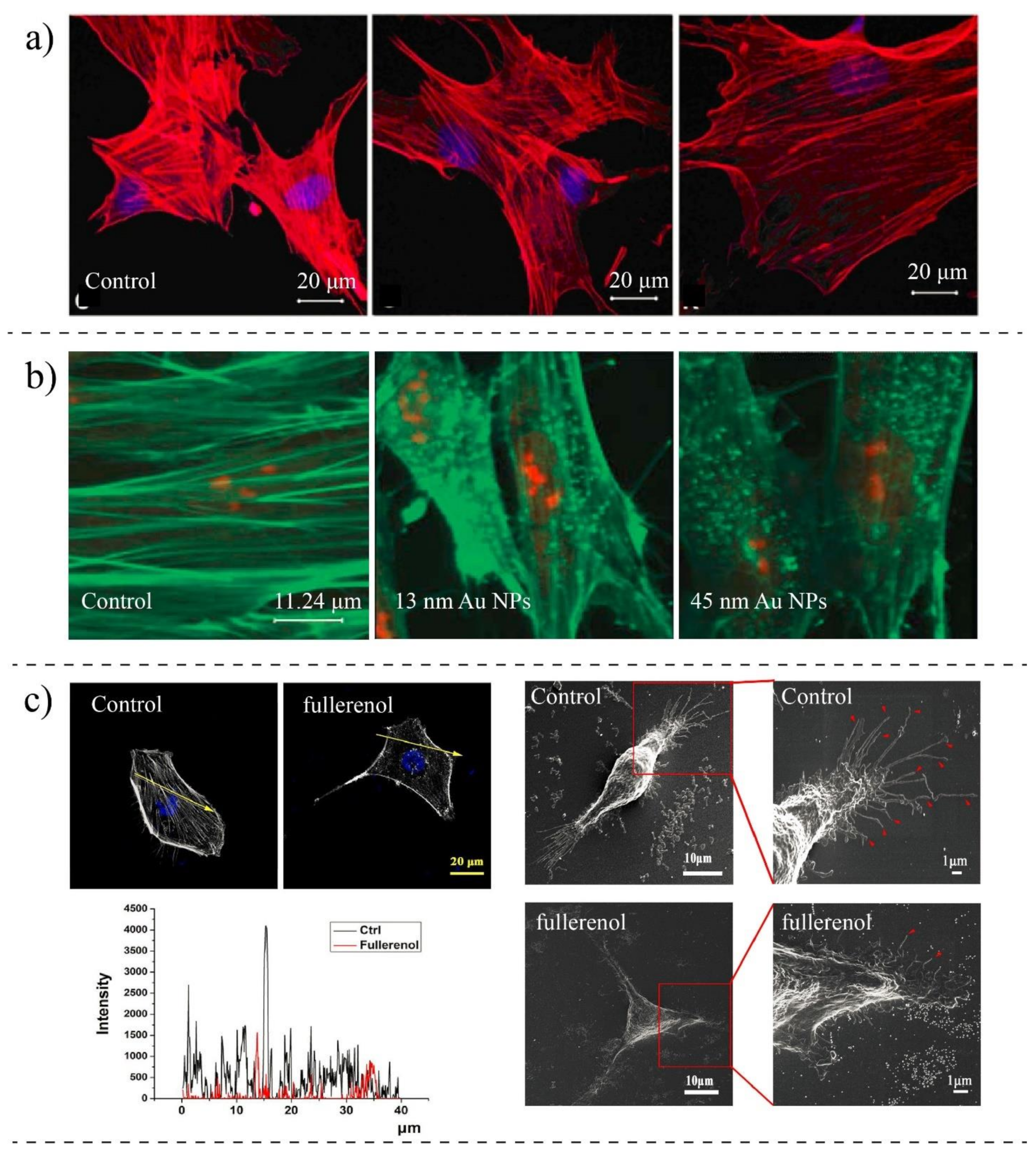
3.3. Impacts of Nanoparticles on Structural Elements and Morphology of Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Author | Cell Type | NPs Type | Methods | Cytoskeleton Changes |
|---|---|---|---|---|
| Pernodet et al., 2007 [162] | CF-31 (human dermal fibroblast) | Gold NPs (13 nm) | TEM, Confocal Imaging, Migration Assay | Modification in actin networks; NPs impaired motility and adhesion |
| Pi et al., 2013 [163] | MCF-7 (breast cancer) | Selenium NPs | AFM, Confocal Microscopy | The organization of F-actin is changed, and they are aggregated; Actin concentration is reduced |
| Choudhury 2013 [150] | A549 (lung cancer) | Citrate-capped Gold NPs (20–60 nm) | Raman, FTIR, TEM, Darkfield Microscopy, UV-Visible Spectroscopy | Inhibiting the polarization of MT; MT structures are damaged, affecting the dynamic equilibrium |
| Qin et al., 2018 [151] | MDA-MB-231 (breast cancer) | Fullerenol NPs | SEM, Fluorescence Imaging, AFM, Scratch Assay | The concentration of actin is reduced, the migration speed is reduced, disturbing actin assembly |
| Hot et al., 2012 [145] | HeLa (cervical cancer) | Single-wall carbon nanotube (1 ± 0.3 nm) | Fluorescence Imaging Microscopy | NPs cause cells to have shorter F-actin; Traction force is reduced; NPs do not affect G-actin and myosin II |
| Huang et al., 2010 [141] | A375 (melanoma) | Silica NPs (MSNs) | TEM, Confocal Microscopy, Western Blot | The actin structure is disorganized and disrupted with NPs; Cell migration is reduced |
| Patra et al., 2007 [159] | A549 (lung cancer) | Gold NPs | Confocal Microscopy | The morphology is changed; Treated cells are rounded compared to non-treated |
| Pisanic et al., 2007 [149] | PC12M (brain) | Fe2O3 NPs | TEM, Western Blot, Fluorescent Microscopy | Reduction in the formation of actin microfilaments; They are less organized; NPs diminish the ability for differentiation |
| Wu et al., 2012 [161] | HAEC (aortic endothelial cells) | Diesel exhaust particles (DEPs) | AFM, Fluorescent Imaging | Cells became degraded; Cellular cytoskeletal structures were impaired |
| Wen et al., 2013 [154] | Acting and tubulin proteins (cell-free system) | Silver NPs | TEM, Hyperspectral Imaging, | Inducing changes in the secondary structures; Silver NPs tend to bind actin vs. tubulin |
| Cooper et al., 2015 [147] | B35 (neuroblastoma) | Silver NPs | Immunocytochemistry | NPs induce F-actin inclusion, disrupting the actin function |
| Rasel et al., 2015 [157] | Osteoblast cells | Boron nitride NPs | AFM, TEM, X-Ray | They do not affect the morphology of cells |
| Liu et al., 2017 [146] | HUVEC (Endothelial cells) | Gold NPs-coated with PEG (20 nm) | Fluorescent Microscopy, Traction Force Microscopy | NPs re-arranged actin filaments; Inhibition of Rock activity reduced the polymerization of actin; Reducing the focal adhesion |
| Vieira et al., 2017 [164] | CCD1072Sk (Normal cells-skin) | Gold NPs and silver NPs | Immunofluorescence Imaging, Cytofluorometry | NPs impair the F-actin;Cytoskeletal reorganization; Cells lose the cell polarization (without losing their viability) |
| Ali et al., 2017 [158] | HSC-3 (tongue cancer) | Gold nanorods coated with PEG and REG | Western Blot, DIC Microscopy, Scratch Assay | The cytoskeletal proteins are rearranged; Cytoskeletal protrusions (filopodia and lamellipoda) are reduced |
| Beaudet et al., 2017 [48] | HeLa (cervical cancer) | AuNPs, Swarna Bhasma | Fluorescent Imaging | Larger particles disrupted the microtubules networks |
| Ibrahim et al., 2018 [144] | SaOS-2 (bone cancer) | TiO2 spherical NPs | Hyperspectral Imaging, Fluorescent Imaging, Western Blot | The actin and microtubule cytoskeletal networks are disorganized |
| Kralovec et al., 2020 [143] | A549 (lung cancer) | Fe3O4@SiO2 | Fluorescent Imaging, Western Blot | Severe disruption of the actin filament and microtubules |
| Kota et al., 2021 [165] | VSMCs (vascular smooth muscle cells) | ZIF-8 NPs | AFM, Fluorescent Imaging, Polymerization Assay | Morphological changes and cytoskeletal disorganization were observed; NPs caused changes in actin filaments at basal and apical surfaces. |
3.4. Impacts of NPs on Cell Stiffness
| Author | Cell Type | Nanoparticles | Parameter | Techniques | Results |
|---|---|---|---|---|---|
| Buyukhatipoglu et al., 2010 [170] | PAEC (endothelial cells) | Iron oxide | Young’s modulus | AFM | The stiffness is increased |
| Yangzhe Wu et al., 2012 [161] | HAEC (endothelial cells) | Diesel exhaust particle (DEP) | Young’s modulus | AFM | Young’s modulus is reduced depending on the doses |
| Jinag Pi et al., 2012 [163] | MCF-7 (breast cancer) | Selenium NPs | Young’s modulus | AFM | The Young’s modulus is reduced. Adhesion is reduced |
| Subbiah et al., 2013 [160] | A549 (lung cancer), NIH3T3 (fibroblasts) HS-5, (fibroblasts) | Hybrid NPs (Silver NPs and single-walled carbon nanotube) | Young’s modulus | AFM | Stiffness is increased |
| Ogneva et al., 2014 [148] | mesenchymal stem cells | Silica-based NPs | Young’s modulus | AFM | Stiffness is increased |
| Rasel et al., 2015 [157] | osteoblast cells | Boron nitride nanoparticle (BN NP) | Young’s modulus | AFM | Stiffness is increased |
| Anna Pietuch et la. 2015 [171] | MDCK II cells (kidney) | Gold nanorods and spherical NPs | Young’s modulus | AFM | Stiffness varies depending on the Au concentration |
| Babhosseini et al., 2015 [173] | MDA-MB-231(breast cancer) | SphKIs with NPs | Deformability-related parameters (passage time and velocity) | Microfluidic (Constriction channel) | Cells became softer (reduced stiffness) |
| Ali et al., 2017 [172] | HEY A8 (ovarian cancer) | Gold nanorods | Young’s modulus of nucleus | AFM | Stiffness is increased and NPs inhibited metastasis |
| Qin et al., 2018 [151] | MDA-MB-231 (breast cancer), MCF7(breast cancer) | Fullerenol NPs | Young’s modulus | AFM | Stiffness is decreased |
| Kashani et al., 2019 [174] | A549 (lung cancer) | Gold nanospheres/nanostars | Young’s modulus | AFM | Stiffness is decreased |
| Rasel et al., 2019 [175] | osteoblast cells | Boron nitride NPs | Young’s modulus | AFM | Stiffness is increased |
| Pastrana et al., 2019 [176] | NIH3T3 (fibroblasts) | Multiwall Carbon NPs | Young’s modulus | AFM | Stiffness is decreased |
| Wilhelm et al., 2021 [177] | F9 murine embryonal carcinoma cells | Magnetic NPs (Iron oxide NPs) | Young’s modulus | Parallel plate rheometer | Stiffness is increased |
3.5. Impacts of Nanoparticles on Migratory Ability of Cells
4. Summary, Conclusions and Outlooks
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AFM | Atomic force microscopy |
| AgNPs | Silver nanoparticles |
| CTAB | Cetrimonium bromide |
| DEPs | Diesel exhaust particles (DEPs), |
| FTIR | Fourier transform-infrared |
| HAECs | Human aortic endothelial cells |
| HSC | Human oral squamous cell carcinoma |
| HUVEC | Human umbilical vein endothelial cells |
| MEMS | Micro-electromechanical systems |
| MP | Micropipette |
| MT | Microtubules |
| MTC | Magnetic twisting cytometry |
| NBI | Nano-bio-interaction |
| NPs | Nanoparticles |
| NRs | Nanorods |
| NSs | Nanospheres |
| OP | Optical tweezer |
| PAECs | Porcine aortic endothelial cells |
| PLGA | Poly lactic-co-glycolic acid |
| SEM | Scanning electron microscopy |
| TEM | Transmission electron microscopy |
| VSMCs | Vascular smooth muscle cells |
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Prigerson, H.G.; Bao, Y.; Shah, M.A.; Paulk, M.E.; LeBlanc, T.; Schneider, B.J.; Garrido, M.; Reid, M.C.; Berlin, D.A.; Adelson, K.B.; et al. Chemotherapy Use, Performance Status, and Quality of Life at the End of Life. JAMA Oncol. 2015, 1, 778–784. [Google Scholar] [CrossRef]
- WHO. Guide to Cancer-Guide to Cancer Early Diagnosis; WHO: Geneva, Switzerland, 2017. [Google Scholar]
- Mehlen, P.; Puisieux, A. Metastasis: A question of life or death. Nat. Rev. Cancer 2006, 6, 449–458. [Google Scholar] [CrossRef]
- Stephen, B.J.; Suchanti, S.; Mishra, R.; Singh, A. Cancer Nanotechnology in Medicine: A Promising Approach for Cancer Detection and Diagnosis. Crit. Rev. Ther. Drug Carr. Syst. 2020, 37, 375–405. [Google Scholar] [CrossRef] [PubMed]
- Sutradhar, K.B.; Amin, M.L. Nanotechnology in Cancer Drug Delivery and Selective Targeting. ISRN Nanotechnol. 2014, 2014, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Siamof, C.M.; Goel, S.; Cai, W. Moving Beyond the Pillars of Cancer Treatment: Perspectives from Nanotechnology. Front. Chem. 2020, 8, 8100. [Google Scholar] [CrossRef]
- Pérez-Herrero, E.; Fernández-Medarde, A. Advanced targeted therapies in cancer: Drug nanocarriers, the future of chemotherapy. Eur. J. Pharm. Biopharm. 2015, 93, 52–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics. CA Cancer J. Clin. 2020, 70, 7–30. [Google Scholar] [CrossRef]
- Price, L.S.L.; Stern, S.T.; Deal, A.M.; Kabanov, A.V.; Zamboni, W.C. A reanalysis of nanoparticle tumor delivery using classical pharmacokinetic metrics. Sci. Adv. 2020, 6, 9249. [Google Scholar] [CrossRef] [PubMed]
- Gmeiner, W.H.; Ghosh, S. Nanotechnology for cancer treatment. Nanotechnol. Rev. 2013, 3, 111–122. [Google Scholar] [CrossRef]
- Mu, W.; Chu, Q.; Liu, Y.; Zhang, N. A Review on Nano-Based Drug Delivery System for Cancer Chemoimmunotherapy. Nano-Micro Lett. 2020, 12, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Jurj, A.; Braicu, C.; Pop, L.A.; Tomuleasa, C.; Gherman, C.D.; Berindan-Neagoe, I. The new era of nanotechnology, an al-ternative to change cancer treatment. Drug Des. Devel. Ther. 2017, 11, 2871–2890. [Google Scholar] [CrossRef] [Green Version]
- Sun, T.; Zhang, Y.S.; Pang, B.; Hyun, D.C.; Yang, M.; Xia, Y. Engineered Nanoparticles for Drug Delivery in Cancer Therapy. Angew. Chem. Int. Ed. 2014, 53, 12320–12364. [Google Scholar] [CrossRef] [PubMed]
- Suri, S.S.; Fenniri, H.; Singh, B. Nanotechnology-based drug delivery systems. J. Occup. Med. Toxicol. 2007, 2, 16. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Cai, R.; Chen, C. The Nano–Bio Interactions of Nanomedicines: Understanding the Biochemical Driving Forces and Redox Reactions. Accounts Chem. Res. 2019, 52, 1507–1518. [Google Scholar] [CrossRef] [PubMed]
- Behzadi, S.; Serpooshan, V.; Tao, W.; Hamaly, M.A.; Alkawareek, M.Y.; Dreaden, E.C.; Brown, D.; Alkilany, A.M.; Farokhzad, O.C.; Mahmoudi, M. Cellular uptake of nanoparticles: Journey inside the cell. Chem. Soc. Rev. 2017, 46, 4218–4244. [Google Scholar] [CrossRef] [PubMed]
- Yameen, B.; Choi, W.I.; Vilos, C.; Swami, A.; Shi, J.; Farokhzad, O.C. Insight into nanoparticle cellular uptake and intra-cellular targeting. J. Control. Release 2014, 190, 485–499. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Liu, W.; Sun, L.; Aifantis, K.E.; Yu, B.; Fan, Y.; Feng, Q.; Cui, F.; Watari, F. Effects of physicochemical properties of nanomaterials on their toxicity. J. Biomed. Mater. Res. Part A 2015, 103, 2499–2507. [Google Scholar] [CrossRef] [PubMed]
- Kashani, A.S.; Piekny, A.; Packirisamy, M. Using intracellular plasmonics to characterize nanomorphology in hu-man cells. Microsystems Nanoeng. 2020, 6, 1. [Google Scholar]
- Suresh, S. Biomechanics and biophysics of cancer cells. Acta Mater. 2007, 55, 3989–4014. [Google Scholar] [CrossRef] [Green Version]
- Eluru, G.; Srinivasan, R.; Gorthi, S.S. Deformability Measurement of Single-Cells at High-Throughput with Imaging Flow Cytometry. J. Light. Technol. 2015, 33, 3475–3480. [Google Scholar] [CrossRef]
- Krieg, M.; Flaeschner, G.; Alsteens, D.; Gaub, B.M.; Roos, W.H.; Wuite, G.J.L.; Gaub, H.E.; Gerber, C.; Dufrêne, Y.F.; Müller, D.J. Atomic force microscopy-based mechanobiology. Nat. Rev. Phys. 2018, 1, 41–57. [Google Scholar] [CrossRef]
- Guo, Q.; Park, S.; Ma, H. Microfluidic micropipette aspiration for measuring the deformability of single cells. Lab a Chip 2012, 12, 2687–2695. [Google Scholar] [CrossRef]
- Kashani, A.S.; Packirisamy, M. Cancer cells optimize elasticity for efficient migration: Migratory index. R. Soc. Open Sci. 2020, 7, 10. [Google Scholar] [CrossRef] [PubMed]
- Nematbakhsh, Y.; Lim, C.T. Cell biomechanics and its applications in human disease diagnosis. Acta Mech. Sin. 2015, 31, 268–273. [Google Scholar] [CrossRef]
- Aghebati-Maleki, A.; Dolati, S.; Ahmadi, M.; Baghbanzhadeh, A.; Asadi, M.; Fotouhi, A.; Yousefi, M.; Aghebati-Maleki, L. Nanoparticles and cancer therapy: Perspectives for application of nanoparticles in the treatment of cancers. J. Cell. Physiol. 2020, 235, 1962–1972. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Roy, I.; Yang, C.; Prasad, P.N. Nanochemistry and Nanomedicine for Nanoparticle-based Diagnostics and Therapy. Chem. Rev. 2016, 116, 2826–2885. [Google Scholar] [CrossRef]
- Donahue, N.D.; Acar, H.; Wilhelm, S. Concepts of nanoparticle cellular uptake, intracellular trafficking, and kinetics in nanomedicine. Adv. Drug Deliv. Rev. 2019, 143, 68–96. [Google Scholar] [CrossRef]
- Kyriakides, T.R.; Raj, A.; Tseng, T.H.; Xiao, H.; Nguyen, R.; Mohammed, F.S.; Halder, S.S.; Xu, M.; Wu, M.J.; Bao, S.; et al. Biocompatibility of nanomaterials and their immunological properties. Biomed. Mater. 2021, 16, 042005. [Google Scholar] [CrossRef]
- Naahidi, S.; Jafari, M.; Edalat, F.; Raymond, K.; Khademhosseini, A.; Chen, P. Biocompatibility of engineered nanoparticles for drug delivery. J. Control. Release 2013, 166, 182–194. [Google Scholar] [CrossRef]
- Li, X.; Wang, L.; Fan, Y.; Feng, Q.; Cui, F.-Z. Biocompatibility and Toxicity of Nanoparticles and Nanotubes. J. Nanomater. 2012, 2012, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Williams, D.F. On the mechanisms of biocompatibility. Biomaterials 2008, 29, 2941–2953. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Huo, S.; Mizuhara, T.; Das, R.; Lee, Y.-W.; Hou, S.; Moyano, D.F.; Duncan, B.; Liang, X.-J.; Rotello, V.M. The Interplay of Size and Surface Functionality on the Cellular Uptake of Sub-10 nm Gold Nanoparticles. ACS Nano. 2015, 9, 9986–9993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, X.; Tian, X.; Wu, A.; Li, J.; Tian, J.; Chong, Y.; Chai, Z.; Zhao, Y.; Chen, C.; Ge, C. Protein Corona Influences Cellular Uptake of Gold Nanoparticles by Phagocytic and Nonphagocytic Cells in a Size-Dependent Manner. ACS Appl. Mater. Interfaces 2015, 7, 20568–20575. [Google Scholar] [CrossRef] [PubMed]
- Yah, C.S. The toxicity of gold nanoparticles in relation to their physicochemical properties. Biomed. Res. 2013, 24, 400–413. [Google Scholar]
- Mironava, T.; Hadjiargyrou, M.; Simon, M.; Jurukovski, V.; Rafailovich, M.H. Gold nanoparticles cellular toxicity and re-covery: Effect of size, concentration and exposure time. Nanotoxicology 2010, 4, 120–137. [Google Scholar] [CrossRef]
- Aguilar, Z.P. Nanomaterials for Medical Applications. Newnes 2013. [Google Scholar] [CrossRef]
- Ahn, S.; Seo, E.; Kim, K.H.; Lee, S.J. Physical Property Control on the Cellular Uptake Pathway and Spatial Distribution of Nanoparticles in Cells. J. Biomed. Nanotechnol. 2015, 11, 1051–1070. [Google Scholar] [CrossRef]
- Serpooshan, V.; Sheibani, S.; Pushparaj, P.; Wojcik, M.; Jang, A.Y.; Santoso, M.R.; Jang, J.H.; Huang, H.; Safavi-Sohi, R.; Haghjoo, N.; et al. Effect of Cell Sex on Uptake of Nanoparticles: The Overlooked Factor at the Nanobio Interface. ACS Nano 2018, 12, 2253–2266. [Google Scholar] [CrossRef]
- Hajipour, M.J.; Aghaverdi, H.; Serpooshan, V.; Vali, H.; Sheibani, S.; Mahmoudi, M. Sex as an important factor in nano-medicine. Nat. Commun. 2021, 12, 2984. [Google Scholar] [CrossRef]
- Zhu, Z.J.; Posati, T.; Moyano, D.F.; Tang, R.; Yan, B.; Vachet, R.W.; Rotello, V.M. The interplay of monolayer structure and serum protein interactions on the cellular uptake of gold nanoparticles. Small 2012, 8, 2659–2663. [Google Scholar] [CrossRef] [PubMed]
- Iversen, T.-G.; Skotland, T.; Sandvig, K. Endocytosis and intracellular transport of nanoparticles: Present knowledge and need for future studies. Nano Today 2011, 6, 176–185. [Google Scholar] [CrossRef]
- Zhang, S.; Gao, H.; Bao, G. Physical Principles of Nanoparticle Cellular Endocytosis. ACS Nano 2015, 9, 8655–8671. [Google Scholar] [CrossRef] [Green Version]
- Rennick, J.J.; Johnston, A.P.R.; Parton, R.G. Key principles and methods for studying the endocytosis of biological and nanoparticle therapeutics. Nat. Nanotechnol. 2021, 16, 266–276. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Neuss, S.; Leifert, A.; Fischler, M.; Wen, F.; Simon, U.; Schmid, G.; Brandau, W.; Jahnen-Dechent, W. Size-Dependent Cytotoxicity of Gold Nanoparticles. Small 2007, 3, 1941–1949. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Nie, G.; Meng, H.; Xia, T.; Nel, A.; Zhao, Y. Physicochemical Properties Determine Nanomaterial Cellular Uptake, Transport, and Fate. Accounts Chem. Res. 2012, 46, 622–631. [Google Scholar] [CrossRef] [Green Version]
- Beaudet, D.; Badilescu, S.; Kuruvinashetti, K.; Kashani, A.S.; Jaunky, D.; Ouellette, S.; Piekny, A.; Packirisamy, M. Comparative study on cellular entry of incinerated ancient gold particles (Swarna Bhasma) and chemically synthesized gold particles. Sci. Rep. 2017, 7, 10678. [Google Scholar] [CrossRef] [Green Version]
- Kashani, A.S.; Badilescu, S.; Piekny, A.; Packirisamy, M. Uptake of Medium-Size Gold Particles in the Nucleus of Living Cells. In Proceedings of the 2nd International Conference of Theoretical and Applied Nanoscience and Nanotechnology (TANN’18), Niagara Falls, Canada, 10 June 2018. [Google Scholar] [CrossRef]
- Kashani, A.S.; Kuruvinashetti, K.; Beauet, D.; Badilescu, S.; Piekny, A.; Packirisamy, M. Enhanced Internalization of Indian Ayurvedic Swarna Bhasma (Gold Nanopowder) for Effective Interaction with Human Cells. J. Nanosci. Nanotechnol. 2018, 18, 6791–6798. [Google Scholar] [CrossRef]
- Mayor, S.; Pagano, R.E. Pathways of clathrin-independent endocytosis. Nat. Rev. Mol. Cell Biol. 2007, 8, 603–612. [Google Scholar] [CrossRef]
- Chithrani, D.; Ghazani, A.A.; Chan, W.C.W. Determining the Size and Shape Dependence of Gold Nanoparticle Uptake into Mammalian Cells. Nano Lett. 2006, 6, 662–668. [Google Scholar] [CrossRef]
- Albanese, A.; Chan, W.C. Effect of Gold Nanoparticle Aggregation on Cell Uptake and Toxicity. ACS Nano 2011, 5, 5478–5489. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Kim, B.Y.; Rutka, J.T.; Chan, W.C.W. Nanoparticle-mediated cellular response is size-dependent. Nat. Nanotechnol. 2008, 3, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.; Ma, H.; Liu, J.; Huo, S.; Kumar, A.; Wei, T.; Zhang, X.; Jin, S.; Gan, Y.; Wang, P.C.; et al. Size-Dependent Localization and Penetration of Ultrasmall Gold Nanoparticles in Cancer Cells, Multicellular Spheroids, and Tumors in Vivo. ACS Nano 2012, 6, 4483–4493. [Google Scholar] [CrossRef] [Green Version]
- Ko, W.-K.; Heo, D.N.; Moon, H.-J.; Lee, S.J.; Bae, M.S.; Lee, J.B.; Sun, I.-C.; Jeon, H.B.; Park, H.K.; Kwon, I.K. The effect of gold nanoparticle size on osteogenic differentiation of adipose-derived stem cells. J. Colloid Interface Sci. 2015, 438, 68–76. [Google Scholar] [CrossRef]
- Fratoddi, I.; Venditti, I.; Cametti, C.; Russo, M.V. How toxic are gold nanoparticles? The state-of-the-art. Nano Res. 2015, 8, 1771–1799. [Google Scholar] [CrossRef]
- Kumar, D.; Mutreja, I.; Chitcholtan, K.; Sykes, P. Cytotoxicity and cellular uptake of different sized gold nanoparticles in ovarian cancer cells. Nanotechnology 2017, 28, 475101. [Google Scholar] [CrossRef]
- Lundqvist, M.; Stigler, J.; Cedervall, T.; Berggård, T.; Flanagan, M.B.; Lynch, I.; Elia, G.; Dawson, K.A. The Evolution of the Protein Corona around Nanoparticles: A Test Study. ACS Nano 2011, 5, 7503–7509. [Google Scholar] [CrossRef]
- Li, Y.; Lee, J.-S. Insights into Characterization Methods and Biomedical Applications of Nanoparticle–Protein Corona. Materials 2020, 13, 3093. [Google Scholar] [CrossRef] [PubMed]
- Favi, P.M.; Gao, M.; Arango, L.J.S.; Ospina, S.P.; Morales, M.; Pavón-Palacio, J.-J.; Webster, T.J. Shape and surface effects on the cytotoxicity of nanoparticles: Gold nanospheres versus gold nanostars. J. Biomed. Mater. Res. Part A 2015, 103, 3449–3462. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Liao, J.; Shao, X.; Li, Q.; Lin, Y. The Effect of shape on Cellular Uptake of Gold Nanoparticles in the forms of Stars, Rods, and Triangles. Sci. Rep. 2017, 7, 1–9. [Google Scholar] [CrossRef]
- Herd, H.; Daum, N.; Jones, A.T.; Huwer, H.; Ghandehari, H.; Lehr, C.-M. Nanoparticle Geometry and Surface Orientation Influence Mode of Cellular Uptake. ACS Nano 2013, 7, 1961–1973. [Google Scholar] [CrossRef] [Green Version]
- Salyers, A.A.; Whitt, D.D. A molecular approach. Bact. Pathog. 2011, 5, 53–100. [Google Scholar]
- Jeon, S.; Clavadetscher, J.; Lee, D.K.; Chankeshwara, S.V.; Bradley, M.; Cho, W.S. Surface charge-dependent cellular up-take of polystyrene nanoparticles. Nanomaterials 2018, 8, 1028. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Gu, N. Thermodynamics of Charged Nanoparticle Adsorption on Charge-Neutral Membranes: A Simulation Study. J. Phys. Chem. B 2010, 114, 2749–2754. [Google Scholar] [CrossRef]
- Cho, E.C.; Xie, J.; Wurm, P.A.; Xia, Y. Understanding the role of surface charges in cellular adsorption versus internaliza-tion by selectively removing gold nanoparticles on the cell surface with a I 2/KI etchant. Nano Lett. 2009, 9, 1080–1084. [Google Scholar]
- Arvizo, R.R.; Miranda, O.R.; Thompson, M.A.; Pabelick, C.M.; Bhattacharya, R.; Robertson, J.; Rotello, V.M.; Prakash, Y.S.; Mukherjee, P. Effect of Nanoparticle Surface Charge at the Plasma Membrane and Beyond. Nano Lett. 2010, 10, 2543–2548. [Google Scholar] [CrossRef] [Green Version]
- Hauck, T.S.; Ghazani, A.A.; Chan, W.C.W. Assessing the Effect of Surface Chemistry on Gold Nanorod Uptake, Toxicity, and Gene Expression in Mammalian Cells. Small 2008, 4, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Yeh, Y.-C.; Saha, K.; Yan, B.; Miranda, O.R.; Yu, X.; Rotello, V.M. The role of ligand coordination on the cytotoxicity of cationic quantum dots in HeLa cells. Nanoscale 2013, 5, 12140–12143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartczak, D.; Kanaras, A. Diacetylene-Containing Ligand as a New Capping Agent for the Preparation of Water-Soluble Colloidal Nanoparticles of Remarkable Stability. Langmuir 2010, 26, 7072–7077. [Google Scholar] [CrossRef] [PubMed]
- Chompoosor, A.; Saha, K.; Ghosh, P.S.; Macarthy, D.J.; Miranda, O.R.; Zhu, Z.J.; Arcaro, K.F.; Rotello, V.M. The Role of Surface Functionality on Acute Cytotoxicity, ROS Generation and DNA Damage by Cationic Gold Nanoparticles. Small 2010, 6, 2246–2249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jansen, K.A.; Donato, D.M.; Balcioglu, H.E.; Schmidt, T.; Danen, E.H.J.; Koenderink, G.H. A guide to mechanobiology: Where biology and physics meet. Biochim. Biophys. Acta Mol. Cell Res. 2015, 1853, 3043–3052. [Google Scholar] [CrossRef] [Green Version]
- Narasimhan, B.N.; Ting, M.S.; Kollmetz, T.; Horrocks, M.S.; Chalard, A.E.; Malmström, J. Mechanical Characterization for Cellular Mechanobiology: Current Trends and Future Prospects. Front. Bioeng. Biotechnol. 2020, 8, 978. [Google Scholar] [CrossRef]
- Kim, D.-H.; Wong, P.K.; Park, J.; Levchenko, A.; Sun, Y. Microengineered Platforms for Cell Mechanobiology. Annu. Rev. Biomed. Eng. 2009, 11, 203–233. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, M.L.; McGarry, P.J.; Sniadecki, N.J. Review on Cell Mechanics: Experimental and Modeling Approaches. Appl. Mech. Rev. 2013, 65, 060801. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Nguyen, J.; Wei, Y.; Sun, Y. Recent advances in microfluidic techniques for single-cell biophysical characterization. Lab. Chip. 2013, 13, 2464–2483. [Google Scholar] [CrossRef] [Green Version]
- Lee, G.Y.; Lim, C.T. Biomechanics approaches to studying human diseases. Trends Biotechnol. 2007, 25, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Krakhmal, N.V.; Zavyalova, M.; Denisov, E.V.; Vtorushin, S.V.; Perelmuter, V. Cancer Invasion: Patterns and Mechanisms. Acta Naturae 2015, 7, 17–28. [Google Scholar] [CrossRef] [Green Version]
- Tao, J.; Li, Y.; Vig, D.K.; Sun, S.X. Cell mechanics: A dialogue. Rep. Prog. Phys. 2010, 80, 036601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fletcher, D.A.; Mullins, R.D. Cell mechanics and the cytoskeleton. Nature 2010, 463, 485–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hohmann, D. The Cytoskeleton—A Complex Interacting Meshwork. Cells 2019, 8, 362. [Google Scholar] [CrossRef] [Green Version]
- Fechheimer, M. Cell and Molecular Biology: Concepts and Experiments. Gerald Karp. Q. Rev. Biol. 2000, 75, 454. [Google Scholar] [CrossRef]
- Lykov, K.; Nematbakhsh, Y.; Shang, M.; Lim, C.T.; Pivkin, I.V. Probing eukaryotic cell mechanics via mesoscopic simulations. PLoS Comput. Biol. 2017, 13, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Dahl, K.N.; Ribeiro, A.J.S.; Lammerding, J. Nuclear shape, mechanics, and mechanotransduction. Circ. Res. 2008, 102, 1307–1318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pullarkat, P.A.; Fernández, P.A.; Ott, A. Rheological properties of the Eukaryotic cell cytoskeleton. Phys. Rep. 2007, 449, 29–53. [Google Scholar] [CrossRef]
- Cotton, F.A.; Wilkinson, G.; Murillo, C.A.; Bochmann, M. Advanced Inorganic Chemistry, 6th ed.; Wiley: Hoboken, NJ, USA, 1999. [Google Scholar]
- Huang, H.; Dai, C.; Shen, H.; Gu, M.; Wang, Y.; Liu, J.; Chen, L.; Sun, L. Recent Advances on the Model, Measurement Technique, and Application of Single Cell Mechanics. Int. J. Mol. Sci. 2020, 21, 6248. [Google Scholar] [CrossRef] [PubMed]
- Sears, D. Protein structure prediction: Concepts and applications. Biochem. Mol. Biol. Educ. 2007, 35, 79–80. [Google Scholar] [CrossRef]
- Pellegrin, S.; Mellor, H. Actin stress fibers. J. Cell Sci. 2007, 120, 3491–3499. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, A.V.; Trompetto, B.; Tan, X.H.M.; Scott, M.B.; Hu, K.H.-H.; Deeds, E.; Butte, M.J.; Chiou, P.Y.; Rowat, A.C. Differential Contributions of Actin and Myosin to the Physical Phenotypes and Invasion of Pancreatic Cancer Cells. Cell. Mol. Bioeng. 2019, 13, 27–44. [Google Scholar] [CrossRef]
- Ford, L.E. Mechanics of Motor Proteins and the Cytoskeleton (review). Perspect. Biol. Med. 2002, 45, 305–307. [Google Scholar] [CrossRef]
- Surcel, A.; Robinson, D.N. Meddling with myosin’s mechanobiology in cancer. Proc. Natl. Acad. of Sci. USA 2006, 116, 15322–15323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mofrad, M.R.K.; Kamm, R.D. Cytoskeletal mechanics: Models and Measurements; Cambridge University Press: Cambridge, UK, 2006. [Google Scholar]
- Calzado-Martín, A.; Encinar, M.; Tamayo, J.; Calleja, M.; Paulo, A.S. Effect of Actin Organization on the Stiffness of Living Breast Cancer Cells Revealed by Peak-Force Modulation Atomic Force Microscopy. ACS Nano 2016, 10, 3365–3374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lekka, M.; Laidler, P.; Gil, D.; Lekki, J.; Stachura, Z.; Hrynkiewicz, A.Z. Elasticity of normal and cancerous human bladder cells studied by scanning force microscopy. Eur. Biophys. J. 1999, 28, 312–316. [Google Scholar] [CrossRef]
- Li, M.; Dang, D.; Liu, L.; Xi, N.; Wang, Y. Atomic force microscopy in characterizing cell mechanics for biomedical applications: A review. IEEE Trans. Nanobiosci. 2017, 16, 523–540. [Google Scholar] [CrossRef]
- Trickey, W.R.; Lee, G.M.; Guilak, F. Viscoelastic properties of chondrocytes from normal and osteoarthritic human cartilage. J. Orthop. Res. 2000, 18, 891–898. [Google Scholar] [CrossRef]
- Bellini, N.; Vishnubhatla, K.C.; Bragheri, F.; Ferrara, L.; Minzioni, P.; Ramponi, R.; Cristiani, I.; Osellame, R. Femtosecond laser fabricated monolithic chip for optical trapping and stretching of single cells. Opt. Express 2010, 18, 4679–4688. [Google Scholar] [CrossRef]
- Coughlin, M.F.; Puig-De-Morales, M.; Bursac, P.; Mellema, M.; Millet, E.; Fredberg, J.J. Filamin-A and Rheological Properties of Cultured Melanoma Cells. Biophys. J. 2006, 90, 2199–2205. [Google Scholar] [CrossRef] [Green Version]
- Byun, S.; Son, S.; Amodei, D.; Cermak, N.; Shaw, J.; Kang, J.H.; Hecht, V.C.; Winslow, M.M.; Jacks, T.; Mallick, P.; et al. Characterizing deformability and surface friction of cancer cells. Proc. Natl. Acad. Sci. USA 2013, 110, 7580–7585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gossett, D.R.; Tse, H.T.K.; Lee, S.A.; Ying, Y.; Lindgren, A.G.; Yang, O.O.; Rao, J.; Clark, A.T.; di Carlo, D. Hydrodynamic stretching of single cells for large population mechanical phenotyping. Proc. Natl. Acad. Sci. USA 2012, 109, 7630–7635. [Google Scholar] [CrossRef] [Green Version]
- Rajagopalan, J.; Saif, M.T.A. MEMS sensors and microsystems for cell mechanobiology. J. Micromechanics Microengineering 2011, 21, 054002–054012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; Lee, G.; Ong, C.N.; Lim, C. AFM indentation study of breast cancer cells. Biochem. Biophys. Res. Commun. 2008, 374, 609–613. [Google Scholar] [CrossRef]
- Pachenari, M.; Seyedpour, S.; Janmaleki, M.; Shayan, S.B.; Taranejoo, S.; Hosseinkhani, H. Mechanical properties of cancer cytoskeleton depend on actin filaments to microtubules content: Investigating different grades of colon cancer cell lines. J. Biomech. 2014, 47, 373–379. [Google Scholar] [CrossRef]
- Wu, P.-H.; Aroush, D.R.-B.; Asnacios, A.; Chen, W.-C.; Dokukin, M.E.; Doss, B.L.; Durand-Smet, P.; Ekpenyong, A.; Guck, J.; Guz, N.V.; et al. A comparison of methods to assess cell mechanical properties. Nat. Methods 2018, 15, 491–498. [Google Scholar] [CrossRef]
- Dao, M.; Lim, C.T.; Suresh, S. Mechanics of the human red blood cell deformed by optical tweezers. J. Mech. Phys. Solids 2003, 51, 2259–2280. [Google Scholar] [CrossRef]
- Kashani, A.S.; Packirisamy, M. Cellular deformation characterization of human breast cancer cells under hydrodynamic forces. AIMS Biophys. 2017, 4, 400–414. [Google Scholar] [CrossRef]
- Lange, J.R.; Steinwachs, J.; Kolb, T.; Lautscham, L.A.; Harder, I.; Whyte, G.; Fabry, B. Microconstriction Arrays for High-Throughput Quantitative Measurements of Cell Mechanical Properties. Biophys. J. 2015, 109, 26–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mak, M.; Erickson, D. A serial micropipette microfluidic device with applications to cancer cell repeated deformation studies. Integr. Biol. 2013, 5, 1374–1384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guck, J.; Schinkinger, S.; Lincoln, B.; Wottawah, F.; Ebert, S.; Romeyke, M.; Lenz, D.; Erickson, H.M.; Ananthakrishnan, R.; Mitchell, D.; et al. Optical deformability as an inherent cell marker for testing malignant transformation and metastatic compe-tence. Biophys. J. 2005, 88, 3689–3698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, T.; Paie, P.; Nava, G.; Bragheri, F.; Vázquez, R.M.; Minzioni, P.; Veglione, M.; DI Tano, M.; Mondello, C.; Osellame, R.; et al. An integrated optofluidic device for single-cell sorting driven by mechanical properties. Lab. Chip. 2015, 15, 1262–1266. [Google Scholar] [CrossRef] [PubMed]
- Bao, N.; Zhan, Y.; Lu, C. Microfluidic Electroporative Flow Cytometry for Studying Single-Cell Biomechanics. Anal. Chem. 2008, 80, 7714–7719. [Google Scholar] [CrossRef]
- Bagnall, J.S.; Byun, S.; Begum, S.; Miyamoto, D.T.; Hecht, V.C.; Maheswaran, S.; Stott, S.L.; Toner, M.; Hynes, R.O.; Manalis, S.R. Deformability of Tumor Cells versus Blood Cells. Sci. Rep. 2015, 5, 18542. [Google Scholar] [CrossRef] [Green Version]
- Corbin, E.A.; Kong, F.; Lim, C.T.; King, W.P.; Bashir, R. Biophysical properties of human breast cancer cells measured using silicon MEMS resonators and atomic force microscopy. Lab. Chip. 2015, 15, 839–847. [Google Scholar] [CrossRef] [PubMed]
- Puig-De-Morales, M.; Grabulosa, M.; Alcaraz, J.; Mullol, J.; Maksym, G.N.; Fredberg, J.J.; Navajas, D. Measurement of cell microrheology by magnetic twisting cytometry with frequency domain de-modulation. J. Appl. Physiol. 2001, 91, 1152–1159. [Google Scholar] [CrossRef]
- Unal, M.; Alapan, Y.; Jia, H.; Varga, A.G.; Angelino, K.; Aslan, M.; Sayin, I.; Han, C.; Jiang, Y.; Zhang, Z.; et al. Micro and Nano-Scale Technologies for Cell Mechanics. Nanobiomedicine 2014, 1, 5. [Google Scholar] [CrossRef] [PubMed]
- Vigmostad, S.C.; Krog, B.L.; Nauseef, J.T.; Henry, M.; Keshav, V. Alterations in cancer cell mechanical properties after fluid shear stress exposure: A micropipette aspiration study. Cell Heal. Cytoskelet. 2015, 7, 25–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shojaei-Baghini, E.; Zheng, Y.; Sun, Y. Automated Micropipette Aspiration of Single Cells. Ann. Biomed. Eng. 2013, 41, 1208–1216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rebelo, L.M.; de Sousa, J.S.; Filho, J.M.; Radmacher, M. Comparison of the viscoelastic properties of cells from different kidney cancer phenotypes measured with atomic force microscopy. Nanotechnology 2013, 24, 055102. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Ramos, J.; Pabijan, J.; García, R.G.; Lekka, M. The softening of human bladder cancer cells happens at an early stage of the malignancy process. Beilstein J. Nanotechnol. 2014, 5, 447–457. [Google Scholar] [CrossRef]
- Ketene, A.N.; Schmelz, E.M.; Roberts, P.C.; Agah, M. The effects of cancer progression on the viscoelasticity of ovarian cell cytoskeleton structures. Nanomed. Nanotechnol. Biol. Med. 2012, 8, 93–102. [Google Scholar] [CrossRef]
- Omidvar, R.; Tafazzoli-Shadpour, M.; Shokrgozar, M.A.; Rostami, M. Atomic force microscope-based single cell force spectroscopy of breast cancer cell lines: An approach for evaluating cellular invasion. J. Biomech. 2014, 47, 3373–3379. [Google Scholar] [CrossRef]
- Binnig, G.; Quate, C.F.; Gerber, C. Atomic Force Microscope. Phys. Rev. Lett. 1986, 56, 930–933. [Google Scholar] [CrossRef] [Green Version]
- Senapati, S.; Lindsay, S. Recent Progress in Molecular Recognition Imaging Using Atomic Force Microscopy. Accounts Chem. Res. 2016, 49, 503–510. [Google Scholar] [CrossRef]
- Vinckier, A.; Semenza, G. Measuring elasticity of biological materials by atomic force microscopy. FEBS Lett. 1998, 430, 12–16. [Google Scholar] [CrossRef]
- Nguyen, T.D.; Gu, Y. Investigation of Cell-Substrate Adhesion Properties of Living Chondrocyte by Measuring Adhesive Shear Force and Detachment Using AFM and Inverse FEA. Sci. Rep. 2016, 6, 1–13. [Google Scholar]
- Thomas, G.; Burnham, N.A.; Camesano, T.A.; Wen, Q. Measuring the Mechanical Properties of Living Cells Using Atomic Force Microscopy. J. Vis. Exp. 2013, 76, e50497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosen, Y.; Gurman, P. MEMS and Microfluidics for Diagnostics Devices. Curr. Pharm. Biotechnol. 2011, 11, 366–375. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Sun, Y. Microfluidic devices for mechanical characterisation of single cells in suspension. Micro Nano Lett. 2011, 6, 327–331. [Google Scholar] [CrossRef] [Green Version]
- Godin, M.; Bryan, A.K.; Burg, T.P.; Babcock, K.; Manalis, S.R. Measuring the mass, density, and size of particles and cells using a suspended microchannel resonator. Appl. Phys. Lett. 2007, 91, 123121. [Google Scholar] [CrossRef] [Green Version]
- Weibel, D.B.; DiLuzio, W.R.; Whitesides, G.M. Microfabrication meets microbiology. Nat. Rev. Genet. 2007, 5, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Shelby, J.P.; White, J.; Ganesan, K.; Rathod, P.K.; Chiu, D.T. A microfluidic model for single-cell capillary obstruction by Plasmodium falciparum-infected erythrocytes. Proc. Natl. Acad. Sci. USA 2003, 100, 14618–14622. [Google Scholar] [CrossRef] [Green Version]
- Valizadeh, A.; Khosroushahi, A.Y. Single-cell analysis based on lab on a chip fluidic system. Anal. Methods 2015, 7, 8524–8533. [Google Scholar] [CrossRef]
- Luo, Z.; Guven, S.; Gözen, I.; Chen, P.; Tasoglu, S.; Anchan, R.M.; Bai, B.; Demirci, U. Deformation of a single mouse oocyte in a constricted microfluidic channel. Microfluid. Nanofluidics 2015, 19, 883–890. [Google Scholar] [CrossRef] [Green Version]
- Guo, Q.; Reiling, S.; Rohrbach, P.; Ma, H. Microfluidic biomechanical assay for red blood cells parasitized by Plasmodium falciparum. Lab. Chip. 2012, 12, 1143–1150. [Google Scholar] [CrossRef]
- Geiser, M.; Rothen-Rutishauser, B.; Kapp, N.; Schürch, S.; Kreyling, W.; Schulz, H.; Semmler, M.; Hof, V.I.; Heyder, J.; Gehr, P. Ultrafine Particles Cross Cellular Membranes by Nonphagocytic Mechanisms in Lungs and in Cultured Cells. Environ. Heal. Perspect. 2005, 113, 1555–1560. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.-L.; Putcha, N.; Ng, K.W.; Leong, D.T.; Lim, C.T.; Loo, S.C.J.; Chen, X. Biophysical Responses upon the Interaction of Nanomaterials with Cellular Interfaces. Accounts Chem. Res. 2012, 46, 782–791. [Google Scholar] [CrossRef]
- Panariti, A.; Miserocchi, G.; Rivolta, I. The effect of nanoparticle uptake on cellular behavior: Disrupting or enabling functions? Nanotechnol. Sci. Appl. 2012, 5, 87–100. [Google Scholar] [PubMed] [Green Version]
- Tian, F.; Cui, D.; Schwarz, H.; Estrada, G.G.; Kobayashi, H. Cytotoxicity of single-wall carbon nanotubes on human fibroblasts. Toxicol. Vitr. 2006, 20, 1202–1212. [Google Scholar] [CrossRef]
- Huang, X.; Teng, X.; Chen, D.; Tang, F.; He, J. The effect of the shape of mesoporous silica nanoparticles on cellular uptake and cell function. Biomaterials 2010, 31, 438–448. [Google Scholar] [CrossRef]
- Meng, H.; Yang, S.; Li, Z.; Xia, T.; Chen, J.; Ji, Z.; Zhang, H.; Wang, X.; Lin, S.; Huang, C.; et al. Aspect Ratio Determines the Quantity of Mesoporous Silica Nanoparticle Uptake by a Small GTPase-Dependent Macropinocytosis Mechanism. ACS Nano 2011, 5, 4434–4447. [Google Scholar] [CrossRef] [PubMed]
- Královec, K.; Melounková, L.; Slováková, M.; Mannová, N.; Sedlák, M.; Bartáček, J.; Havelek, R. Disruption of cell adhesion and cytoskeletal networks by thiol-functionalized silica-coated iron oxide nanoparticles. Int. J. Mol. Sci. 2020, 21, 9350. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, M.; Schoelermann, J.; Mustafa, K.; Cimpan, M.R. TiO2 nanoparticles disrupt cell adhesion and the architecture of cytoskeletal networks of human osteoblast-like cells in a size dependent manner. J. Biomed. Mater. Res. Part A 2018, 106, 2582–2593. [Google Scholar] [CrossRef]
- Holt, B.D.; Shams, H.; Horst, T.A.; Basu, S.; Rape, A.D.; Wang, Y.L.; Rohde, G.K.; Mofrad, M.R.; Islam, M.F.; Dahl, K.N. Altered Cell Mechanics from the Inside: Dispersed Single Wall Carbon Nanotubes Integrate with and Re-structure Actin. J. Funct. Biomater. 2012, 3, 398–417. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Rogel, N.; Harada, K.; Jarett, L.; Maiorana, C.H.; German, G.K.; Mahler, G.J.; Doiron, A.L. Nanoparticle size-specific actin rearrangement and barrier dysfunction of endothelial cells. Nanotoxicology 2017, 11, 846–856. [Google Scholar] [CrossRef]
- Cooper, R.J.; Spitzer, N. Silver nanoparticles at sublethal concentrations disrupt cytoskeleton and neurite dynamics in cultured adult neural stem cells. NeuroToxicology 2015, 48, 231–238. [Google Scholar] [CrossRef] [Green Version]
- Ogneva, I.V.; Buravkov, S.V.; Shubenkov, A.N.; Buravkova, L.B. Mechanical characteristics of mesenchymal stem cells under impact of silica-based nanoparticles. Nanoscale Res. Lett. 2014, 9, 284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pisanic, T.R.; Blackwell, J.D.; Shubayev, V.I.; Fiñones, R.R.; Jin, S. Nanotoxicity of iron oxide nanoparticle internalization in growing neurons. Biomaterials 2007, 28, 2572–2581. [Google Scholar] [CrossRef]
- Choudhury, D.; Xavier, P.L.; Chaudhari, K.; John, R.; Dasgupta, A.K.; Pradeep, T.; Chakrabarti, G. Unprecedented inhibition of tubulin polymerization directed by gold nanoparticles inducing cell cycle arrest and apoptosis. Nanoscale 2013, 5, 4476–4489. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.; Chen, K.; Gu, W.; Dong, X.; Lei, R.; Chang, Y.; Bai, X.; Xia, S.; Zeng, L.; Zhang, J.; et al. Small size fullerenol nanoparticles suppress lung metastasis of breast cancer cell by disrupting actin dynamics. J. Nanobiotechnol. 2018, 16, 54. [Google Scholar] [CrossRef]
- Issaad, D.; Moustaoui, H.; Medjahed, A.; Lalaoui, L.; Spadavecchia, J.; Bouafia, M.; de la Chapelle, M.L.; Djaker, N. Scattering Correlation Spectroscopy and Raman Spectroscopy of Thiophenol on Gold Nanoparticles: Comparative Study between Nanospheres and Nanourchins. J. Phys. Chem. C 2017, 121, 18254–18262. [Google Scholar] [CrossRef]
- Kashani, A.S.; Badilescu, S.; Piekny, A.; Packirisamy, M. Differing Affinities of Gold Nanostars and Nanospheres toward HeLa and HepG2 Cells: Implications for Cancer Therapy. ACS Appl. Nano Mater. 2020, 3, 4114–4126. [Google Scholar] [CrossRef]
- Wen, Y.; Geitner, N.; Chen, R.; Ding, F.; Chen, P.; Andorfer, R.E.; Govindan, P.N.; Ke, P.C. Binding of cytoskeletal proteins with silver nanoparticles. RSC Adv. 2013, 3, 22002–22007. [Google Scholar] [CrossRef]
- Sinclair, W.E.; Chang, H.H.; Dan, A.; Kenis, P.J.A.; Murphy, C.J.; Leckband, D.E. Gold nanoparticles disrupt actin organization and pulmonary endothelial barriers. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef]
- Septiadi, D.; Crippa, F.; Moore, T.L.; Rothen-Rutishauser, B.; Petri-Fink, A. Nanoparticle–Cell Interaction: A Cell Mechanics Perspective. Adv. Mater. 2018, 30, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Rasel, M.A.; Li, T.; Nguyen, T.D.; Singh, S.; Zhou, Y.; Xiao, Y.; Gu, Y. Biophysical response of living cells to boron nitride nanoparticles: Uptake mechanism and biomechanical characterization. J. Nanopart. Res. 2015, 17, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Ali, M.R.K.; Wu, Y.; Tang, Y.; Xiao, H.; Chen, K.; Han, T.; Fang, N.; Wu, R.; El-Sayed, M.A. Targeting cancer cell integrins using gold nanorods in photothermal therapy inhibits migration through affecting cytoskeletal proteins. Proc. Natl. Acad. Sci. USA 2017, 114, E5655–E5663. [Google Scholar] [CrossRef] [Green Version]
- Patra, H.; Banerjee, S.; Chaudhuri, U.; Lahiri, P.; Dasgupta, A.K. Cell selective response to gold nanoparticles. Nanomed. Nanotechnol. Biol. Med. 2007, 3, 111–119. [Google Scholar] [CrossRef]
- Subbiah, R.; Ramasundaram, S.; Du, P.; Hyojin, K.; Sung, D.; Park, K.; Lee, N.E.; Yun, K.; Choi, K.J. Evaluation of cytotoxicity, biophysics and biomechanics of cells treated with functionalized hybrid nano-materials. J. R. Soc. Interface 2013, 10, 20130694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.; Yu, T.; Gilbertson, T.A.; Zhou, A.; Xu, H.; Nguyen, K.T. Biophysical Assessment of Single Cell Cytotoxicity: Diesel Exhaust Particle-Treated Human Aortic Endothelial Cells. PLoS ONE 2012, 7, e36885. [Google Scholar] [CrossRef] [Green Version]
- Pernodet, N.; Fang, X.; Sun, Y.; Bakhtina, A.; Ramakrishnan, A.; Sokolov, J.; Ulman, A.; Rafailovich, M. Adverse Effects of Citrate/Gold Nanoparticles on Human Dermal Fibroblasts. Small 2006, 2, 766–773. [Google Scholar] [CrossRef]
- Pi, J.; Yang, F.; Jin, H.; Huang, X.; Liu, R.; Yang, P.; Cai, J. Selenium nanoparticles induced membrane bio-mechanical property changes in MCF-7 cells by disturbing mem-brane molecules and F-actin. Bioorganic Med. Chem. Lett. 2013, 23, 6296–6303. [Google Scholar] [CrossRef]
- Vieira, L.F.d.; Lins, M.P.; Viana, I.M.M.N.; Santos, J.E.d.; Smaniotto, S.; Reis, M.D.d.S. Metallic nanoparticles re-duce the migration of human fibroblasts in vitro. Nanoscale Res. Lett. 2017, 12, 200. [Google Scholar] [CrossRef] [Green Version]
- Kota, D.; Kang, L.; Rickel, A.; Liu, J.; Smith, S.; Hong, Z.; Wang, C. Low doses of zeolitic imidazolate framework-8 nanoparticles alter the actin organization and contractility of vascular smooth muscle cells. J. Hazard. Mater. 2021, 414, 125514. [Google Scholar] [CrossRef] [PubMed]
- Plodinec, M.; Loparic, M.; Monnier, C.; Obermann, E.C.; Zanetti-Dallenbach, R.; Oertle, P.; Hyotyla, J.T.; Aebi, U.; Bentires-Alj, M.; Lim, R.; et al. The nanomechanical signature of breast cancer. Nat. Nanotechnol. 2012, 7, 757–765. [Google Scholar] [CrossRef]
- Xu, W.; Mezencev, R.; Kim, B.; Wang, L.; McDonald, J.F.; Sulchek, T. Cell Stiffness Is a Biomarker of the Metastatic Potential of Ovarian Cancer Cells. PLoS ONE 2012, 7, e46609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tojkander, S.; Gateva, G.; Lappalainen, P. Actin stress fibers-Assembly, dynamics and biological roles. J. Cell Sci. 2012, 125, 1855–1864. [Google Scholar] [CrossRef] [Green Version]
- Rotsch, C.; Radmacher, M. Drug-Induced Changes of Cytoskeletal Structure and Mechanics in Fibroblasts: An Atomic Force Microscopy Study. Biophys. J. 2000, 78, 520–535. [Google Scholar] [CrossRef] [Green Version]
- Buyukhatipoglu, K.; Clyne, A. Superparamagnetic iron oxide nanoparticles change endothelial cell morphology and mechanics via reactive oxygen species formation. J. Biomed. Mater. Res. Part A 2010, 96, 186–195. [Google Scholar] [CrossRef] [PubMed]
- Pietuch, A.; Brückner, B.R.; Schneider, D.; Tarantola, M.; Rosman, C.; Sönnichsen, C.; Janshoff, A. Mechanical properties of MDCK II cells exposed to gold nanorods. Beilstein J. Nanotechnol. 2015, 6, 223–231. [Google Scholar] [CrossRef] [Green Version]
- Ali, M.R.K.; Wu, Y.; Ghosh, D.; Do, B.H.; Chen, K.; Dawson, M.; Fang, N.; Sulchek, T.A.; El-Sayed, M.A. Nuclear Membrane-Targeted Gold Nanoparticles Inhibit Cancer Cell Migration and Invasion. ACS Nano 2017, 11, 3716–3726. [Google Scholar] [CrossRef] [Green Version]
- Babahosseini, H.; Srinivasaraghavan, V.; Zhao, Z.; Gillam, F.; Childress, E.; Strobl, J.S.; Santos, W.L.; Zhang, C.; Agah, M. The impact of sphingosine kinase inhibitor-loaded nanoparticles on bioelectrical and biomechanical properties of cancer cells. Lab. Chip. 2015, 16, 188–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kashani, A.S.; Badilescu, S.; Piekny, A.; Packirisamy, M. Abstract format–Gold nanoparticles modulate the mechanobiological properties of human lung cancer cells. In Proceedings of the 35th Annual Meeting of the Canadian Biomaterials Society 2019, Quebec City, QC, Canada, 24 May 2019; Volume 30, p. 10678. [Google Scholar]
- Rasel, A.I.; Singh, S.; Nguyen, T.D.; Afara, I.O.; Gu, Y. Impact of Nanoparticle Uptake on the Biophysical Properties of Cell for Biomedical Engineering Applications. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Pastrana, H.F.; Rivera, A.X.C.; Raman, A.; Ávila, A. Evaluation of the elastic Young’ s modulus and cytotoxicity variations in fibroblasts exposed to carbon-based nanomaterials. J. Nanobiotechnol. 2019, 17, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Perez, J.E.; Fage, F.; Pereira, D.; Abou-Hassan, A.; Asnacios, S.; Asnacios, A.; Wilhelm, C. Transient cell stiffening triggered by magnetic nanoparticle exposure. J. Nanobiotechnol. 2021, 19, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Cai, K.; Hou, Y.; Li, J.; Chen, X.; Hu, Y.; Luo, Z.; Ding, X.; Xu, D.; Lai, M. Effects of titanium nanoparticles on adhesion, migration, proliferation, and differentiation of mesenchymal stem cells. Int. J. Nanomed. 2013, 8, 3619–3630. [Google Scholar] [CrossRef] [Green Version]
- Zhou, T.; Yu, M.; Zhang, B.; Wang, L.; Wu, X.; Zhou, H.; Du, Y.; Hao, J.; Tu, Y.; Chen, C.; et al. Inhibition of cancer cell migration by gold nanorods: Molecular mechanisms and implications for cancer therapy. Adv. Funct. Mater. 2014, 24, 6922–6932. [Google Scholar] [CrossRef]
- Pan, Y.; Wu, Q.; Qin, L.; Cai, J.; Du, B. Gold nanoparticles inhibit VEGF165-induced migration and tube formation of endothelial cells via the Akt pathway. Biomed Res. Int. 2014, 2014, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Chan, C.-M.; Hsiao, C.-Y.; Li, H.-J.; Fang, J.-Y.; Chang, D.-C.; Hung, C.-F. The Inhibitory Effects of Gold Nanoparticles on VEGF-A-Induced Cell Migration in Choroid-Retina Endothelial Cells. Int. J. Mol. Sci. 2019, 21, 109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Wu, Y.; Guo, Z.; Liu, Y.; Shen, Y.; Zhou, P.; Lu, X. Effects of Internalized Gold Nanoparticles with Respect to Cytotoxicity and Invasion Activity in Lung Cancer Cells. PLoS ONE 2014, 9, e99175. [Google Scholar] [CrossRef] [Green Version]
- Shahhoseini, E.; Nakayama, M.; Piva, T.; Geso, M. Differential Effects of Gold Nanoparticles and Ionizing Radiation on Cell Motility between Primary Human Colonic and Melanocytic Cells and Their Cancerous Counterparts. Int. J. Mol. Sci. 2021, 22, 1418. [Google Scholar] [CrossRef]
- Kashani, A.S.; Badilescu, S.; Piekny, A.; Packirisamy, M. Perspective—Bio-Nano-Interaction in Treatment and Management of Cancer. J. Electrochem. Soc. 2018, 166, B3007–B3011. [Google Scholar] [CrossRef]
- Wang, M.D.; Shin, D.M.; Simons, J.W.; Nie, S. Nanotechnology for targeted cancer therapy. Expert Rev. Anticancer. Ther. 2007, 7, 833–837. [Google Scholar] [CrossRef] [PubMed]
- Sechi, M.; Sanna, V.; Pala, N. Targeted therapy using nanotechnology: Focus on cancer. Int. J. Nanomed. 2014, 9, 467–483. [Google Scholar] [CrossRef] [PubMed] [Green Version]








| Techniques | Cell Type | Mechanical Stimuli | Important Parameters | Advantages | Limitations | |
|---|---|---|---|---|---|---|
| Classical Techniques | Atomic Force Microscopy (AFM) | MCF7 [104]; Human bladder [96] | Cantilever micro indention | Tip deflection, Young’s modulus | High-resolution measurement; Provids both structural and mechanical information for local, whole, and interior measurements [23,97] | Low throughput; Mechanical hitting of AFM tip may affect cell activities and position of probe; Requires a high-resolution microscope |
| Micropipette aspiration (MA) | Human cartilage [98]; Colon cancer cells [105] | Negative force | Young’s modulus | Low-cost and well-established method | Limited spatial resolution; Low throughput; For suspended cells only | |
| Magnetic twisting cytometry (MTC) | Melanoma [100]; MCF7 [106] | Force is applied by magnetic beads | Stiffness and Young’s modulus | Inducing little heat and photodamages compared to optical tweezer [10] | Resolution limitation; Inducing non-uniform stress; Beads are localized randomly on cell; Attachment angle affects the displacement | |
| Optical tweezers (OP) | RBC [99,107] | Laser-induced surface force | Deformation index | Without physical contact | Only for suspended cells; Damaging consequence of optical heating on cells; Limited magnitude of forces | |
| Parallel plate | Epithelial ovarian cancer [23]; MCF7 [106,108] | Shear stress | Aspect ratio | Homogeneity of the applied shear stress; Simplicity; Ability to study cell population | Need bulky devices; Large amount of reagents; Difficult to visualize deformation | |
| Microfluidic Techniques | Fluid-induced deformation | PBMCs [102] | Fluid shear stress | Deformation index, size | High throughput; Simultaneously, other chemical assays can be done; The measurment can be done continuously; Contactless deformation; Applicable for both suspended and adhered cells | Needing expensive high-speed camera for imaging |
| Constriction-induced deformation | K562 [109]; MDA-MB-231 [110] | Mechanical squeezing | Passage time, entry times, stiffness | Wide-ranging applications in cell deformation; Applicable for different geometry structures; Adjustable dimension for different cell types | Clogging and channel blockage; Possible effects of friction between cell and channel’s wall on measurements; Ignoring the effects of membrane rigidity and viscosity | |
| Aspiration-induced deformation | Neutrophils [24] | Negative pressure | Young’s modulus, cortical tension | Straightforward method; Well-established mathematical model | Leaking problem; Rectangle-like cross-section of microfluidic channels; Time-consuming process; Requiring high-vacuum pressure | |
| Optical stretcher | MCF7 [106]; MCF-7, MCF-10, MDA-MB-231 [111]; Red blood cells [99]; Melanoma cells [112] | Optically-induced surface forces | Deformation index, cell elasticity | No physical contact; Relatively high-throughput measurements | Alignment problem; Optical heating; Thermal damage | |
| Electrical-induced deformation | MCF-10A, MCF-7 [113] | Electroporation-induced swelling | Deformation index, size of cells | Fast heat dissipation; Better resolution; Automation and parallelization of test with reduced amount of samples | High energy consumption and high voltage | |
| MEMS Techniques | Suspended microcantilever | Circulating tumor cells [114]; Fibroblast [101] | External actuator | Frequency of cantilever, passage time, transit time | All-inclusive systems; Parallel analysis; Better quality factor; Automation | Fabrication is expensive; Non-transparent channels; High stiffness of silicon; calibration process |
| MEMS resonator | MCF7 [115] | External actuator | Frequency of cantilever | High throughput | Expensive fabrication; Requiring external electrical system; Only for adherent cells | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sohrabi Kashani, A.; Packirisamy, M. Cancer-Nano-Interaction: From Cellular Uptake to Mechanobiological Responses. Int. J. Mol. Sci. 2021, 22, 9587. https://doi.org/10.3390/ijms22179587
Sohrabi Kashani A, Packirisamy M. Cancer-Nano-Interaction: From Cellular Uptake to Mechanobiological Responses. International Journal of Molecular Sciences. 2021; 22(17):9587. https://doi.org/10.3390/ijms22179587
Chicago/Turabian StyleSohrabi Kashani, Ahmad, and Muthukumaran Packirisamy. 2021. "Cancer-Nano-Interaction: From Cellular Uptake to Mechanobiological Responses" International Journal of Molecular Sciences 22, no. 17: 9587. https://doi.org/10.3390/ijms22179587
APA StyleSohrabi Kashani, A., & Packirisamy, M. (2021). Cancer-Nano-Interaction: From Cellular Uptake to Mechanobiological Responses. International Journal of Molecular Sciences, 22(17), 9587. https://doi.org/10.3390/ijms22179587