
Kisspeptin Receptor on the Sperm Surface Reflects Epididymal Maturation in the Dog

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Anatomic Attributes of the of Testis and Epididymis
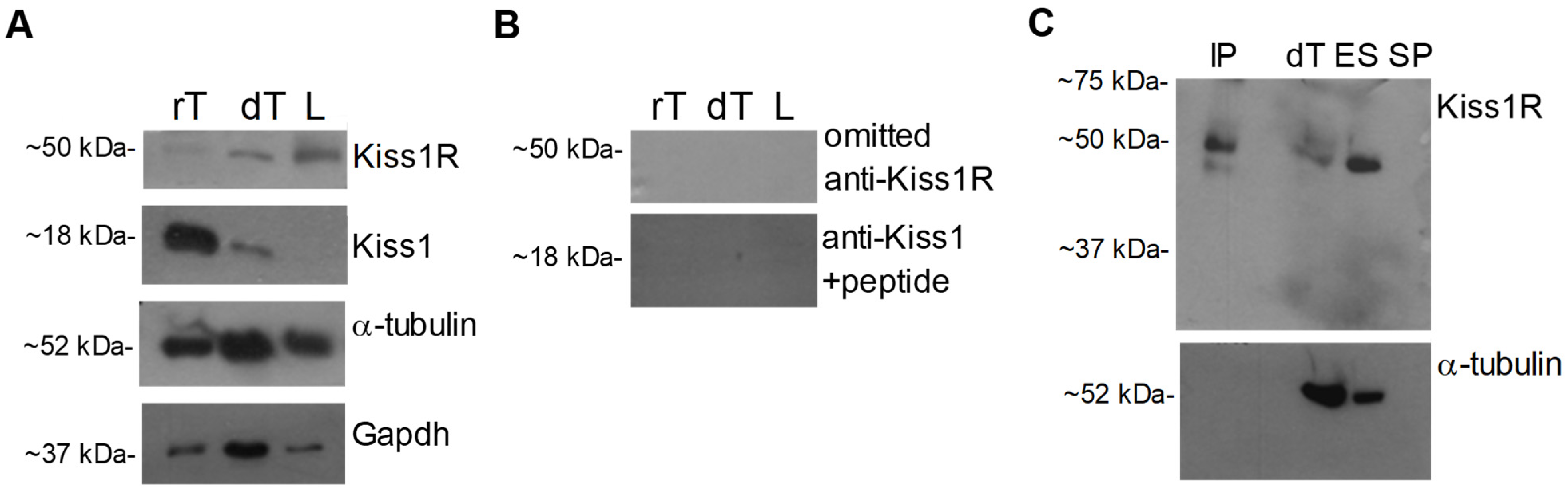
2.2. Evaluation of Kiss1 and Kiss1R in Dog Testis, Spermatozoa and Epididymis
2.2.1. Western Blot
2.2.2. Kisspeptin System in Dog Sperm and Epididymis Fluid
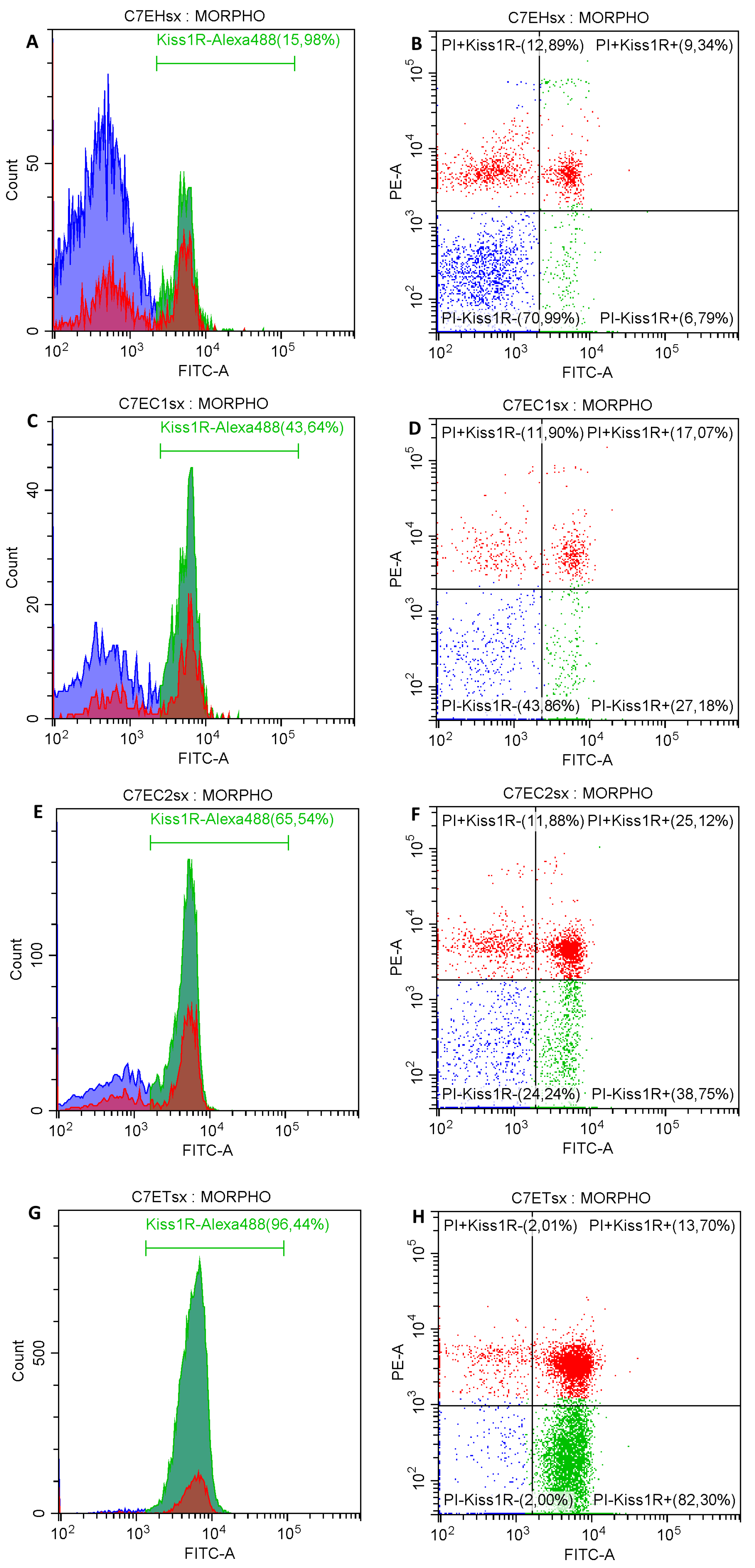
2.2.3. Kiss1R Superficial Localization on Epididymal Spermatozoa
2.3. Sperm Attributes along the Length of the Epididymis
3. Discussion
4. Material and Methods
4.1. Animals
4.2. Surgical Orchiectomy
4.3. Epididymis Collection and Samples Harvest
4.4. Evaluation of KISS1 and Kiss1R in Dog Testis, Spermatozoa and Epididymis
4.4.1. Total Protein Extraction
4.4.2. Sperm Membrane Preparation
4.4.3. Western Blot Analysis
4.4.4. Antisera Specificity Assay
4.4.5. Detection of Kiss1R on Surface of Epididymal Spermatozoa
4.4.6. Dot Blot Analysis
4.5. Sperm Quality Evaluation
4.5.1. Sperm Concentration
4.5.2. Sperm Kinetics
4.5.3. Morphology
4.5.4. Sperm Protamine Deficiency
4.6. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- D’Anglemont de Tassigny, X.; Fagg, L.A.; Carlton, M.B.L.; Colledge, W.H. Kisspeptin can stimulate Gonadotropin-Releasing Hormone (GnRH) release by a direct action at GnRH nerve terminals. Endocrinology 2008, 149, 3926–3932. [Google Scholar] [CrossRef] [PubMed]
- Herbison, A.E.; de Tassigny, X.D.A.; Doran, J.; Colledge, W.H. Distribution and postnatal development of Gpr54 gene expression in mouse brain and gonadotropin-releasing hormone neurons. Endocrinology 2010, 151, 312–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Messager, S.; Chatzidaki, E.E.; Ma, D.; Hendrick, A.G.; Zahn, D.; Dixon, J.; Thresher, R.R.; Malinge, I.; Lomet, D.; Carlton, M.B.L.; et al. Kisspeptin directly stimulates gonadotropin-releasing hormone release via G protein-coupled receptor 54. Proc. Natl. Acad. Sci. USA 2005, 102, 1761–1766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Novaira, H.J.; Ng, Y.; Wolfe, A.; Radovick, S. Kisspeptin increases GnRH mRNA expression and secretion in GnRH secreting neuronal cell lines. Mol. Cell. Endocrinol. 2009, 311, 126–134. [Google Scholar] [CrossRef] [Green Version]
- Pinilla, L.; Aguilar, E.; Dieguez, C.; Millar, R.P.; Tena-Sempere, M. Kisspeptins and Reproduction: Physiological Roles and Regulatory Mechanisms. Physiol. Rev. 2012, 92, 1235–1316. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Miele, M.E.; Hicks, D.J.; Phillips, K.K.; Trent, J.M.; Weissman, B.E.; Welch, D.R. KiSS-1, a novel human malignant melanoma metastasis-suppressor gene. J. Natl. Cancer Inst. 1996, 88, 1731–1737. [Google Scholar] [CrossRef]
- Gottsch, M.L.; Clifton, D.K.; Steiner, R.A. From KISS1 to kisspeptins: An historical perspective and suggested nomenclature. Peptides 2009, 30, 4–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bilban, M.; Ghaffari-Tabrizi, N.; Hintermann, E.; Bauer, S.; Molzer, S.; Zoratti, C.; Malli, R.; Sharabi, A.; Hiden, U.; Graier, W. Kisspeptin-10, a KiSS-1/metastin-derived decapeptide, is a physiological invasion inhibitor of primary human trophoblasts. J. Cell Sci. 2004, 117, 1319–1328. [Google Scholar] [CrossRef] [Green Version]
- Kotani, M.; Detheux, M.; Vandenbogaerde, A.; Communi, D.; Vanderwinden, J.M.; Le Poul, E.; Brézillon, S.; Tyldesley, R.; Suarez-Huerta, N.; Vandeput, F. The Metastasis Suppressor Gene KiSS-1 Encodes Kisspeptins, the Natural Ligands of the Orphan G Protein-coupled Receptor GPR54. J. Biol. Chem. 2001, 276, 34631–34636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navarro, V.M. Metabolic regulation of kisspeptin—The link between energy balance and reproduction. Nat. Rev. Endocrinol. 2020, 16, 407–420. [Google Scholar] [CrossRef]
- Uenoyama, Y.; Inoue, N.; Nakamura, S.; Tsukamura, H. Central Mechanism Controlling Pubertal Onset in Mammals: A Triggering Role of Kisspeptin. Front. Endocrinol. 2019, 10, 312. [Google Scholar] [CrossRef] [Green Version]
- Tanyapanyachon, P.; Amelkina, O.; Chatdarong, K. The expression of kisspeptin and its receptor in the domestic cat ovary and uterus in different stages of the ovarian cycle. Theriogenology 2018, 117, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Cielesh, M.; McGrath, B.; Scott, C.; Norman, S.; Stephen, C. The localization of kisspeptin and kisspeptin receptor in the canine ovary during different stages of the reproductive cycle. Reprod. Domest. Anim. 2017, 52, 24–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clarke, I.J.; Smith, J.T.; Caraty, A.; Goodman, R.L.; Lehman, M.N. Kisspeptin and seasonality in sheep. Peptides 2009, 30, 154–163. [Google Scholar] [CrossRef] [Green Version]
- Smith, J.T.; Clay, C.M.; Caraty, A.; Clarke, I.J. KiSS-1 messenger ribonucleic acid expression in the hypothalamus of the ewe is regulated by sex steroids and season. Endocrinology 2007, 148, 1150–1157. [Google Scholar] [CrossRef]
- Revel, F.G.; Saboureau, M.; Masson-Pévet, M.; Pévet, P.; Mikkelsen, J.D.D.; Simonneaux, V. Kisspeptin Mediates the Photoperiodic Control of Reproduction in Hamsters. Curr. Biol. 2006, 16, 1730–1735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chianese, R.; Ciaramella, V.; Fasano, S.; Pierantoni, R.; Meccariello, R. Kisspeptin Receptor, GPR54, as a Candidate for the Regulation of Testicular Activity in the Frog Rana esculenta. Biol. Reprod. 2013, 88. [Google Scholar] [CrossRef]
- Chianese, R.; Ciaramella, V.; Fasano, S.; Pierantoni, R.; Meccariello, R. Kisspeptin drives germ cell progression in the anuran amphibian Pelophylax esculentus: A study carried out in ex vivo testes. Gen. Comp. Endocrinol. 2015, 211, 81–91. [Google Scholar] [CrossRef]
- Ohga, H.; Selvaraj, S.; Matsuyama, M. The Roles of Kisspeptin System in the Reproductive Physiology of Fish With Special Reference to Chub Mackerel Studies as Main Axis. Front. Endocrinol. 2018, 9, 147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, Y.; Li, Z.; Jiang, W.; Ling, Y.; Kuang, H. Reproductive functions of Kisspeptin/KISS1R Systems in the Periphery. Reprod. Biol. Endocrinol. 2019, 17, 65. [Google Scholar] [CrossRef]
- Cejudo Roman, A.; Pinto, F.M.; Dorta, I.; Almeida, T.A.; Hernández, M.; Illanes, M.; Tena-Sempere, M.; Candenas, L. Analysis of the expression of neurokinin B, kisspeptin, and their cognate receptors NK3R and KISS1R in the human female genital tract. Fertil. Steril. 2012, 97, 1213–1219. [Google Scholar] [CrossRef]
- Gaytan, F.; Garcia-Galiano, D.; Dorfman, M.D.; Manfredi-Lozano, M.; Castellano, J.M.; Dissen, G.A.; Ojeda, S.R.; Tena-Sempere, M. Kisspeptin receptor haplo-insufficiency causes premature ovarian failure despite preserved gonadotropin secretion. Endocrinology 2014, 155, 3088–3097. [Google Scholar] [CrossRef]
- Chianese, R.; Cobellis, G.; Chioccarelli, T.; Ciaramella, V.; Migliaccio, M.; Fasano, S.; Pierantoni, R.; Meccariello, R. Kisspeptins, Estrogens and male fertility. Curr. Med. Chem. 2016, 23, 4070–4091. [Google Scholar] [CrossRef]
- Meccariello, R.; Fasano, S.; Pierantoni, R. Kisspeptins, new local modulators of male reproduction: A comparative overview. Gen. Comp. Endocrinol. 2020, 299, 113618. [Google Scholar] [CrossRef]
- Sharma, A.; Thaventhiran, T.; Minhas, S.; Dhillo, W.S.; Jayasena, C.N. Kisspeptin and testicular function—Is it necessary? Int. J. Mol. Sci. 2020, 21, 2958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wahab, F.; Atika, B.; Shahab, M.; Behr, R. Kisspeptin signalling in the physiology and pathophysiology of the urogenital system. Nat. Rev. Urol. 2016, 13, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Chianese, R.; Ciaramella, V.; Fasano, S.; Pierantoni, R.; Meccariello, R. Kisspeptin regulates steroidogenesis and spermiation in anuran amphibian. Reproduction 2017, 154, 403–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, M.C.; Wang, J.Y.; Lee, Y.J.; Jong, D.S.; Tsui, K.H.; Chiu, C.H. Kisspeptin modulates fertilization capacity of mouse spermatozoa. Reproduction 2014, 147, 835–845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinto, F.M.; Cejudo-Román, A.; Ravina, C.G.; Fernández-Sánchez, M.; Martín-Lozano, D.; Illanes, M.; Tena-Sempere, M.; Candenas, M.L. Characterization of the kisspeptin system in human spermatozoa. Int. J. Androl. 2012, 35, 63–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gervasi, M.G.; Visconti, P.E. Molecular changes and signaling events occurring in spermatozoa during epididymal maturation. Andrology 2017, 5, 204–218. [Google Scholar] [CrossRef]
- Bedford, J.M. Report of a workshop: Maturation of the fertilizing ability of mammalian spermatozoa in the male and female reproductive tract. Biol. Reprod. 1974, 11, 346–362. [Google Scholar] [CrossRef] [PubMed]
- Cooper, T.G. Role of the epididymis in mediating changes in the male gamete during maturation. Adv. Exp. Med. Biol. 1995, 377. [Google Scholar] [CrossRef]
- Contri, A.; Gloria, A.; Robbe, D.; De Amicis, I.; Carluccio, A. Characteristics of donkey spermatozoa along the length of the epididymis. Theriogenology 2012, 77, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Jones, R. Plasma Membrane Composition and Organisation During Maturation of Spermatozoa in the Epididymis. In The Epididymis: From Molecules to Clinical Practice; Springer: Boston, MA, USA, 2002; pp. 405–416. [Google Scholar] [CrossRef]
- Ostrander, E.A.; Giniger, E. Semper Fidelis: What Man’s Best Friend Can Teach Us about Human Biology and Disease. Am. J. Hum. Genet. 1997, 61, 475–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirchhoff, C. The dog as a model to study human epididymal function at a molecular level. Mol. Hum. Reprod. 2002, 8, 695–701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schäfer-Somi, S.; Ay, S.; Kaya, D.; Sözmen, M.; Beceriklisoy, H.; Ağaoğlu, A.; Fındık, M.; van Haeften, T.; Aslan, S. Kisspeptin-10 and the G protein-coupled receptor 54 are differentially expressed in the canine pregnant uterus and trophoblast cells. Reprod. Domest. Anim. 2017, 52, 123–129. [Google Scholar] [CrossRef]
- Albers-Wolthers, C.H.J.; de Gier, J.; Rutten, V.P.M.G.; van Kooten, P.J.S.; Leegwater, P.A.J.; Schaefers-Okkens, A.C.; Kooistra, H.S. The effects of kisspeptin agonist canine KP-10 and kisspeptin antagonist p271 on plasma LH concentrations during different stages of the estrous cycle and anestrus in the bitch. Theriogenology 2016, 86, 589–595. [Google Scholar] [CrossRef] [PubMed]
- Albers-Wolthers, C.H.J.; de Gier, J.; Walen, M.; van Kooten, P.J.S.; Lambalk, C.B.; Leegwater, P.A.J.; Roelen, B.A.J.; Schaefers-Okkens, A.C.; Rutten, V.P.M.G.; Millar, R.P.M.; et al. In vitro and in vivo effects of kisspeptin antagonists p234, p271, p354, and p356 on GPR54 activation. PLoS ONE 2017, 12, e0179156. [Google Scholar] [CrossRef] [Green Version]
- Amann, R.; Hammerstedt, R.; Veeramachaneni, D. The epididymis and sperm maturation: A perspective. Reprod. Fertil. Dev. 1993, 5, 361–381. [Google Scholar] [CrossRef]
- Bianco, S.D.; Vandepas, L.; Correa-Medina, M.; Gereben, B.; Mukherjee, A.; Kuohung, W.; Carroll, R.; Teles, M.G.; Latronico, A.C.; Kaiser, U.B. KISS1R Intracellular Trafficking and Degradation: Effect of the Arg386Pro Disease-Associated Mutation. Endocrinology 2011, 152, 1616–1626. [Google Scholar] [CrossRef] [Green Version]
- Aquila, S.; Sisci, D.; Gentile, M.; Middea, E.; Catalano, S.; Carpino, A.; Rago, V.; Andò, S. Estrogen Receptor (ER)α and ERβ Are Both Expressed in Human Ejaculated Spermatozoa: Evidence of Their Direct Interaction with Phosphatidylinositol-3-OH Kinase/Akt Pathway. J. Clin. Endocrinol. Metab. 2004, 89, 1443–1451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blackmore, P.F.; Lattanzio, F.A. Cell surface localization of a novel non-genomic progesterone receptor on the head of human sperm. Biochem. Biophys. Res. Commun. 1991, 181, 331–336. [Google Scholar] [CrossRef]
- Sirivaidyapong, S.; Bevers, M.M.; Gadella, B.M.; Colenbrander, B. Induction of the acrosome reaction in dog sperm cells is dependent on epididymal maturation: The generation of a functional progesterone receptor is involved. Mol. Reprod. Dev. 2001, 58, 451–459. [Google Scholar] [CrossRef]
- Costello, S.; Michelangeli, F.; Nash, K.; Lefievre, L.; Morris, J.; Machado-Oliveira, G.; Barratt, C.; Kirkman-Brown, J.; Publicover, S. Ca2+-stores in sperm: Their identities and functions. Reproduction 2009, 138, 425–437. [Google Scholar] [CrossRef] [Green Version]
- Bedu-Addo, K.; Barratt, C.L.R.; Kirkman-Brown, J.C.; Publicover, S.J. Patterns of [Ca2+]i mobilization and cell response in human spermatozoa exposed to progesterone. Dev. Biol. 2007, 302, 324–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kay, V.J.; Coutts, J.R.T.; Robertson, L. Andrology: Effects of pentoxifylline and progesterone on human sperm capacitation and acrosome reaction. Hum. Reprod. 1994, 9, 2318–2323. [Google Scholar] [CrossRef]
- Sagare-Patil, V.; Galvankar, M.; Satiya, M.; Bhandari, B.; Gupta, S.K.; Modi, D. Differential concentration and time dependent effects of progesterone on kinase activity, hyperactivation and acrosome reaction in human spermatozoa. Int. J. Androl. 2012, 35, 633–644. [Google Scholar] [CrossRef] [PubMed]
- Sumigama, S.; Mansell, S.; Miller, M.; Lishko, P.V.; Cherr, G.N.; Meyers, S.A.; Tollner, T. Progesterone Accelerates the Completion of Sperm Capacitation and Activates CatSper Channel in Spermatozoa from the Rhesus Macaque. Biol. Reprod. 2015, 93, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Deppe, M.; Risopatrón, J.; Sánchez, R. Trypsin and chymotrypsin are involved in the progesterone-induced acrosome reaction in canine spermatozoa. Reprod. Domest. Anim. 2010, 45, 453–457. [Google Scholar] [CrossRef] [PubMed]
- Baldi, E.; Casano, R.; Falsetti, C.; Krausz, C.; Maggi, M.; Forti, G. Intracellular Calcium Accumulation and Responsiveness to Progesterone in Capacitating Human Spermatozoa. J. Androl. 1991, 12, 323–330. [Google Scholar] [CrossRef]
- Luque, G.M.; Dalotto-Moreno, T.; Martín-Hidalgo, D.; Ritagliati, C.; Molina, L.C.P.; Romarowski, A.; Balestrini, P.A.; Schiavi-Ehrenhaus, L.J.; Gilio, N.; Krapf, D.; et al. Only a subpopulation of mouse sperm displays a rapid increase in intracellular calcium during capacitation. J. Cell. Physiol. 2018, 233, 9685–9700. [Google Scholar] [CrossRef]
- Vicente-Carrillo, A.; Álvarez-Rodríguez, M.; Rodríguez-Martínez, H. The CatSper channel modulates boar sperm motility during capacitation. Reprod. Biol. 2017, 17, 69–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeung, C.H.; Cooper, T.G.; Oberpenning, F.; Schulze, H.; Nieschlag, E. Changes in Movement Characteristics of Human Spermatozoa along the Length of the Epididymis. Biol. Reprod. 1993, 49, 274–280. [Google Scholar] [CrossRef] [Green Version]
- Soler, C.; Yeung, C.H.; Cooper, T.G. Development of sperm motility patterns in the murine epididymis. Int. J. Androl. 1994, 17, 271–278. [Google Scholar] [CrossRef] [PubMed]
- Yeung, C.H.; Oberländer, G.; Cooper, T.G. Characterization of the motility of maturing rat spermatozoa by computer-aided objective measurement. Reproduction 1992, 96, 427–441. [Google Scholar] [CrossRef] [Green Version]
- Sutovsky, P.; Moreno, R.; Ramalho-Santos, J.; Dominko, T.; Thompson, W.E.; Schatten, G. A putative, ubiquitin-dependent mechanism for the recognition and elimination of defective spermatozoa in the mammalian epididymis. J. Cell Sci. 2001, 114, 1665–1675. [Google Scholar] [CrossRef] [PubMed]
- Hermo, L.; Oliveira, R.L.; Smith, C.E.; Au, C.E.; Bergeron, J.J.M. Dark side of the epididymis: Tails of sperm maturation. Andrology 2019, 7, 566–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esterhuizen, A.D.; Franken, D.R.; Lourens, J.G.H.; Prinsloo, E.; Van Rooyen, L.H. Sperm chromatin packaging as an indicator of in-vitro fertilization rates. Hum. Reprod. 2000, 15, 657–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fathi, Z.; Tavalaee, M.; Kiani, A.; Deemeh, M.R.; Modaresi, M.; Nasr-Esfahani, M.H. Flow cytometry: A Novel approach for Indirect Assessment of Protamine Deficiency by CMA3 staining, Taking into Account the Presence of M540 or Apoptotic Bodies. Int. J. Fertil. Steril. 2011, 5, 128–133. [Google Scholar]
- Fortes, M.R.S.; Satake, N.; Corbet, D.H.; Corbet, N.J.; Burns, B.M.; Moore, S.S.; Boe-Hansen, G.B. Sperm protamine deficiency correlates with sperm DNA damage in Bos indicus bulls. Andrology 2014, 2, 370–378. [Google Scholar] [CrossRef]
- Rahman, M.B.; Vandaele, L.; Rijsselaere, T.; Maes, D.; Hoogewijs, M.; Frijters, A.; Noordman, J.; Granados, A.; Dernelle, E.; Shamsuddin, M. Scrotal insulation and its relationship to abnormal morphology, chromatin protamination and nuclear shape of spermatozoa in Holstein-Friesian and Belgian Blue bulls. Theriogenology 2011, 76, 1246–1257. [Google Scholar] [CrossRef] [PubMed]
- Amann, R.P.; Veeramachaneni, D.N.R. Cryptorchidism in common eutherian mammals. Reproduction 2007, 133, 541–561. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; De Iuliis, G.N.; Dun, M.D.; Nixon, B. Characteristics of the Epididymal Luminal Environment Responsible for Sperm Maturation and Storage. Front. Endocrinol. 2018, 9, 59. [Google Scholar] [CrossRef] [Green Version]
- Salehi, S.; Adeshina, I.; Chen, H.; Zirkin, B.R.; Hussain, M.A.; Wondisford, F.; Wolfe, A.; Radovick, S. Developmental and Endocrine Regulation of Kisspeptin Expression in Mouse Leydig Cells. Endocrinology 2015, 156, 1514–1522. [Google Scholar] [CrossRef] [Green Version]
- Abou Khalil, N.S.; Mahmoud, G.B. Reproductive, antioxidant and metabolic responses of Ossimi rams to kisspeptin. Theriogenology 2020, 142, 414–420. [Google Scholar] [CrossRef]
- Güvenç, M.; Aksakal, M. Ameliorating effect of kisspeptin-10 on methotrexate-induced sperm damages and testicular oxidative stress in rats. Andrologia 2018, 50, e13057. [Google Scholar] [CrossRef]
- Zou, P.; Wang, X.; Chen, Q.; Yang, H.; Zhou, N.; Sun, L.; Chen, H.; Liu, J.; Ao, L.; Cui, Z. Kisspeptin Protein in Seminal Plasma Is Positively Associated with Semen Quality: Results from the MARHCS Study in Chongqing, China. Biomed Res. Int. 2019, 2019, 5129263. [Google Scholar] [CrossRef]
- Rota, A.; Ström, B.; Linde-Forsberg, C.; Rodriguez-Martinez, H. Effects of equex stm paste on viability of frozen-thawed dog spermatozoa during in vitro incubation at 38 °C. Theriogenology 1997, 47, 1093–1101. [Google Scholar] [CrossRef]
- Lowry, O.; Rosenbrough, N.; Farr, A.; Randall, R. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Thaler, C.D.; Thomas, M.; Ramalie, J.R. Reorganization of mouse sperm lipid rafts by capacitation. Mol. Reprod. Dev. 2006, 73, 1541–1549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, S.; Li, G.; Qu, L.; Cao, Y.; Wang, Q.; Zhou, J.; Zhong, R.; Guo, X.; Zhu, Y. Identification of New Epididymal Luminal Fluid Proteins Involved in Sperm Maturation in Infertile Rats Treated by Dutasteride Using iTRAQ. Molecules 2016, 21, 602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rijsselaere, T.; Van Soom, A.; Maes, D.; De Kruif, A. Effect of technical settings on canine semen motility parameters measured by the Hamilton-Thorne analyzer. Theriogenology 2003, 60. [Google Scholar] [CrossRef]
- Gloria, A.; Di Francesco, L.; Marruchella, G.; Robbe, D.; Contri, A. Pulse-wave Doppler pulsatility and resistive indexes of the testicular artery increase in canine testis with abnormal spermatogenesis. Theriogenology 2020, 158. [Google Scholar] [CrossRef]
- Hancock, J.L. The morphology of boar spermatozoa. J. R. Microsc. Soc. 1957, 76, 84–97. [Google Scholar] [CrossRef] [PubMed]
- Barth, A.D.; Oko, R.J. Abnormal Morphology of Bovine Spermatozoa; Iowa State University Press: Ames, IA, USA, 1989. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| EH (Mean ± SEM) | EC1 (Mean ± SEM) | EC2 (Mean ± SEM) | ET (Mean ± SEM) | |
|---|---|---|---|---|
| Kiss1R-/PI− (%) | 54.76 ± 3.48 a | 24.37 ± 4.15 b | 29.27 ± 4.98 b | 10.19 ± 2.97 c |
| Kiss1R+/PI− (%) | 19.11 ± 3.37 a | 31.99 ± 4.83 b | 31.73 ± 5.78 b | 75.37 ± 4.61 c |
| Kiss1R+/PI+ (%) | 15.58 ± 1.25 a | 32.98 ± 1.89 b | 30.74 ± 2.67 b | 11.68 ± 1.38 a |
| Kiss1R−/PI+ (%) | 10.55 ± 1.08 a | 10.67 ± 1.81 a | 8.27 ± 1.19 a | 2.76 ± 1.09 b |
| TRACT | ||||
|---|---|---|---|---|
| EH (Mean ± SEM) | EC1 (Mean ± SEM) | EC2 (Mean ± SEM) | ET (Mean ± SEM) | |
| Concentration (×106/mL) | 6.4 ± 1.2 a | 22.8 ± 4.3 b | 27.3 ± 3.9 b | 368.6 ± 53.9 c |
| Total motility (%) | 2 ± 0.9 a | 22.9 ± 4 b | 51.9 ± 6.3 c | 83.8 ± 4.8 d |
| Progressive motility (%) | 0 | 2.8 ± 2.4 a | 9.2 ± 1.9 a | 39.7 ± 7.1 b |
| VAP (µm/s) | 13 ± 3.8 a | 38.4 ± 5.3 b | 58.4 ± 5.6 c | 99.2 ± 4.4 d |
| VSL (µm/s) | 11.2 ± 3.3 a | 24.9 ± 4.4 b | 37.7 ± 3.8 c | 75.2 ± 5.5 d |
| VCL (µm/s) | 25.4 ± 8.3 a | 90.2 ± 10.8 b | 136.5 ± 13.7 c | 174.5 ± 9.6 d |
| ALH (µm) | 0.6 ± 0.6 a | 6.5 ± 1.2 b | 9 ± 0.7 c | 8.2 ± 0.5 c |
| BCF (Hz) | 16.6 ± 4.7 a | 30.9 ± 2.3 b | 30.3 ± 2.2 b | 35 ± 0.8 b |
| STR (%) | 49.5 ± 11.3 a | 61.3 ± 4.8 b | 59.6 ± 4.4 b | 73.1 ± 3.1 c |
| LIN (%) | 30.8 ± 8.2 a | 29.4 ± 2.8 a | 27.6 ± 2.3 a | 45.3 ± 4.1 b |
| EH (Mean ± SEM) | EC1 (Mean ± SEM) | EC2 (Mean ± SEM) | ET (Mean ± SEM) | |
|---|---|---|---|---|
| Normal spermatozoa | 89.9 ± 1.4 a | 57.3 ± 2.9 b | 59.7 ± 2.9 b | 90.1 ± 1.4 a |
| Primary abnormalities (%) | 5.5 ± 1.6 a | 6.2 ± 1.8 a | 5.9 ± 2.2 a | 4.8 ± 1.7 a |
| Secondary abnormalities (%) | 4.6 ± 1.2 a | 36.5 ± 3.7 b | 34.4 ± 3.5 b | 5.2 ± 1.2 a |
| Proximal cytoplasmic droplet (%) | 39.3 ± 2.2 a | 20.4 ± 8.6 b | 3.5 ± 1.9 c | 0.8 ± 0.3 d |
| Distal cytoplasmic droplet (%) | 0 | 14.3 ± 13.5 a | 41.3 ± 8.1 b | 27.7 ± 6.8 c |
| Protamine deficiency (%) | 45.4 ± 1.2 a | 20.3 ± 1.4 b | 14.6 ± 1.1 c | 4.2 ± 0.8 d |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gloria, A.; Contri, A.; Mele, E.; Fasano, S.; Pierantoni, R.; Meccariello, R. Kisspeptin Receptor on the Sperm Surface Reflects Epididymal Maturation in the Dog. Int. J. Mol. Sci. 2021, 22, 10120. https://doi.org/10.3390/ijms221810120
Gloria A, Contri A, Mele E, Fasano S, Pierantoni R, Meccariello R. Kisspeptin Receptor on the Sperm Surface Reflects Epididymal Maturation in the Dog. International Journal of Molecular Sciences. 2021; 22(18):10120. https://doi.org/10.3390/ijms221810120
Chicago/Turabian StyleGloria, Alessia, Alberto Contri, Elena Mele, Silvia Fasano, Riccardo Pierantoni, and Rosaria Meccariello. 2021. "Kisspeptin Receptor on the Sperm Surface Reflects Epididymal Maturation in the Dog" International Journal of Molecular Sciences 22, no. 18: 10120. https://doi.org/10.3390/ijms221810120
APA StyleGloria, A., Contri, A., Mele, E., Fasano, S., Pierantoni, R., & Meccariello, R. (2021). Kisspeptin Receptor on the Sperm Surface Reflects Epididymal Maturation in the Dog. International Journal of Molecular Sciences, 22(18), 10120. https://doi.org/10.3390/ijms221810120