
Geomagnetic Field (GMF)-Dependent Modulation of Iron-Sulfur Interplay in Arabidopsis thaliana


 ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Results
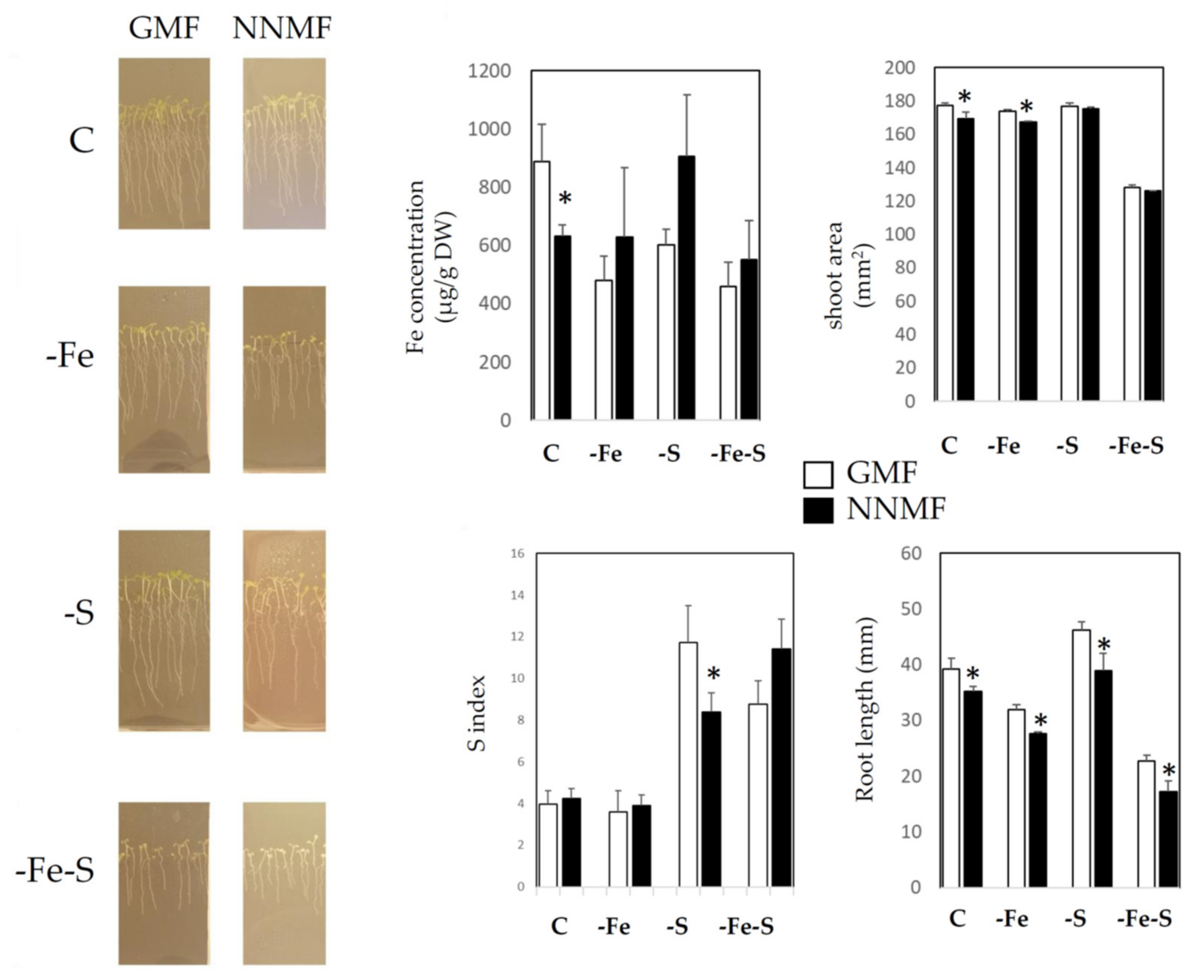
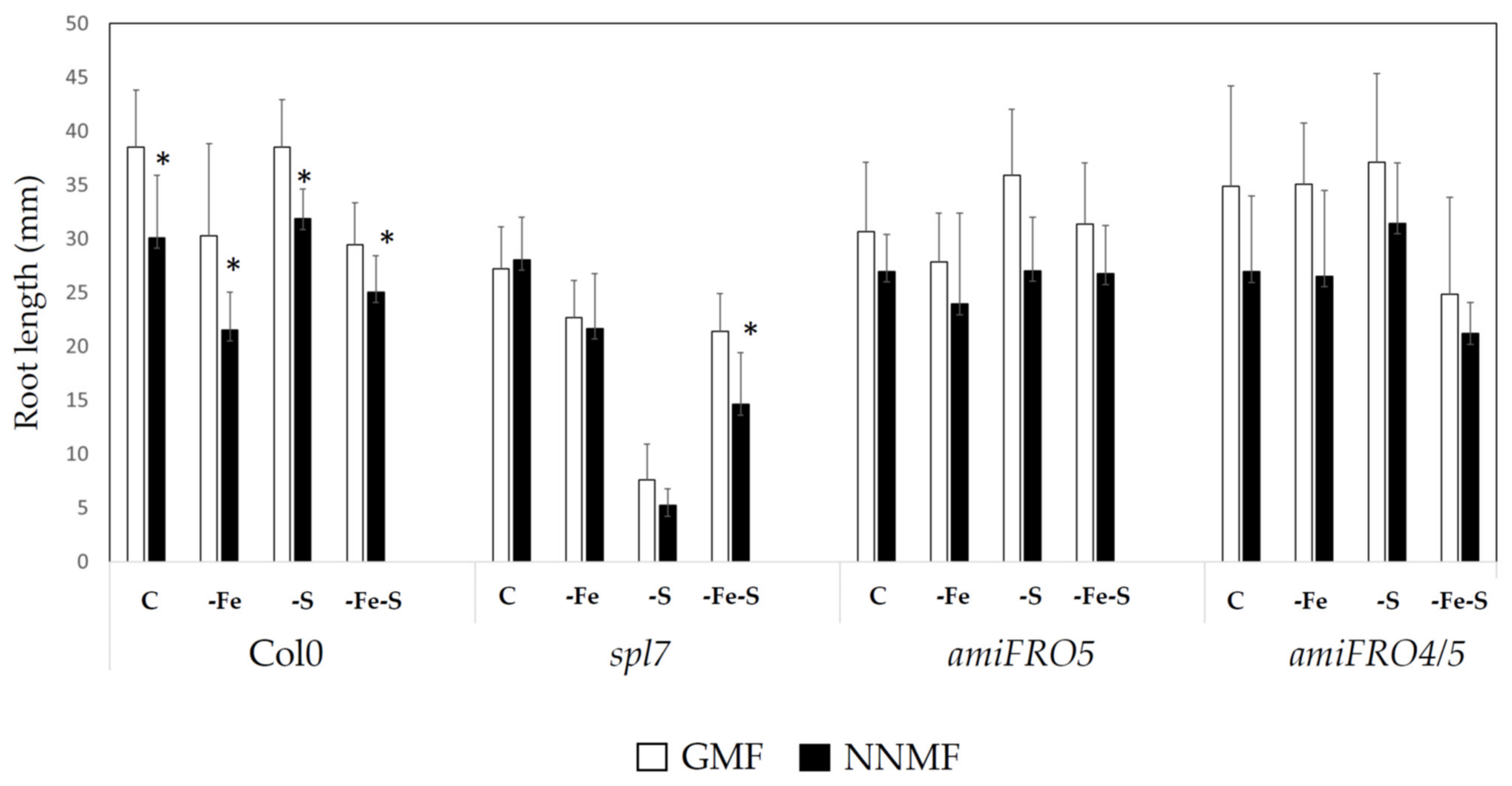
2.1. GMF Differentially Affects Plant Growth under S and Fe Deficiency
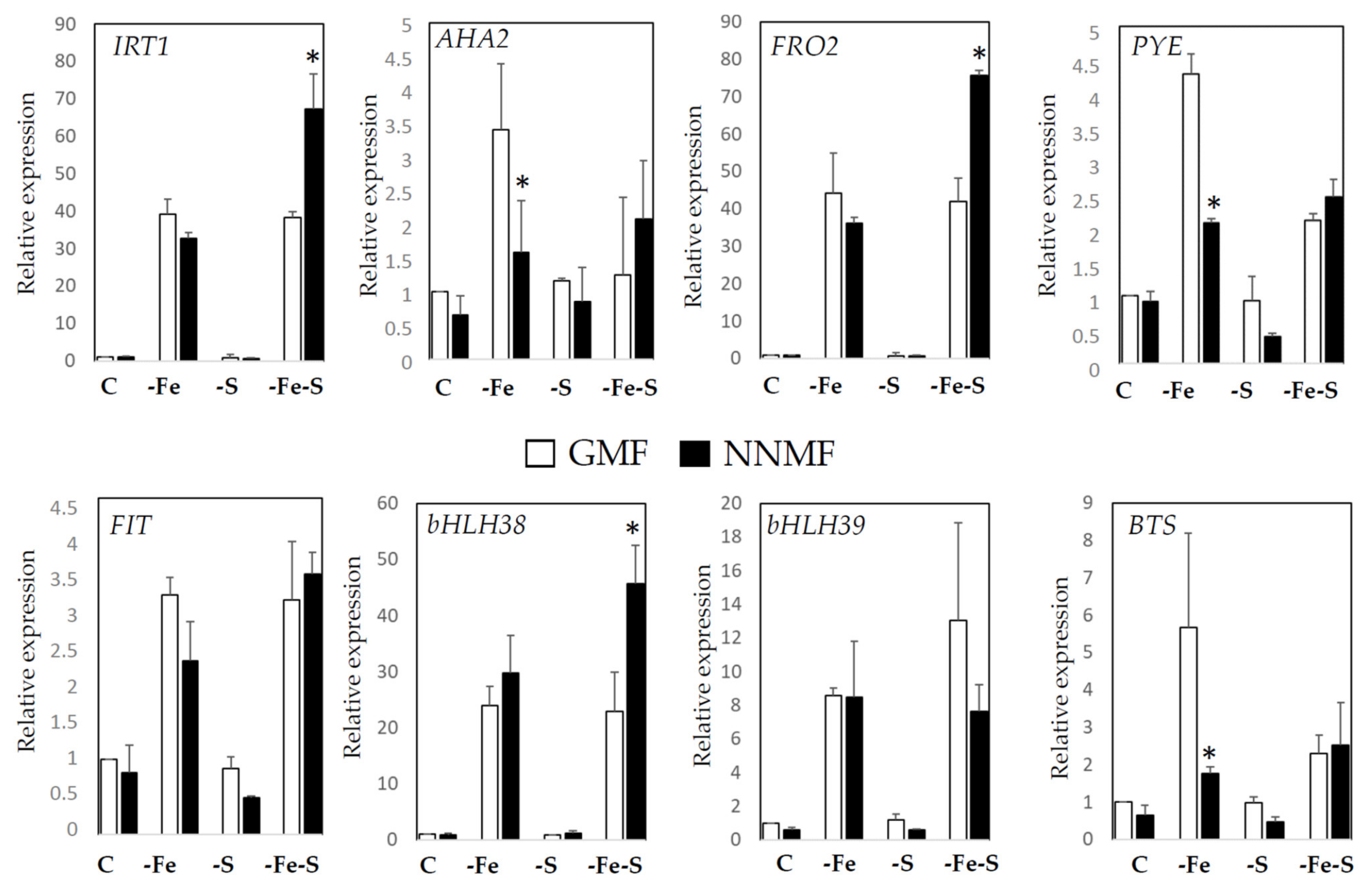
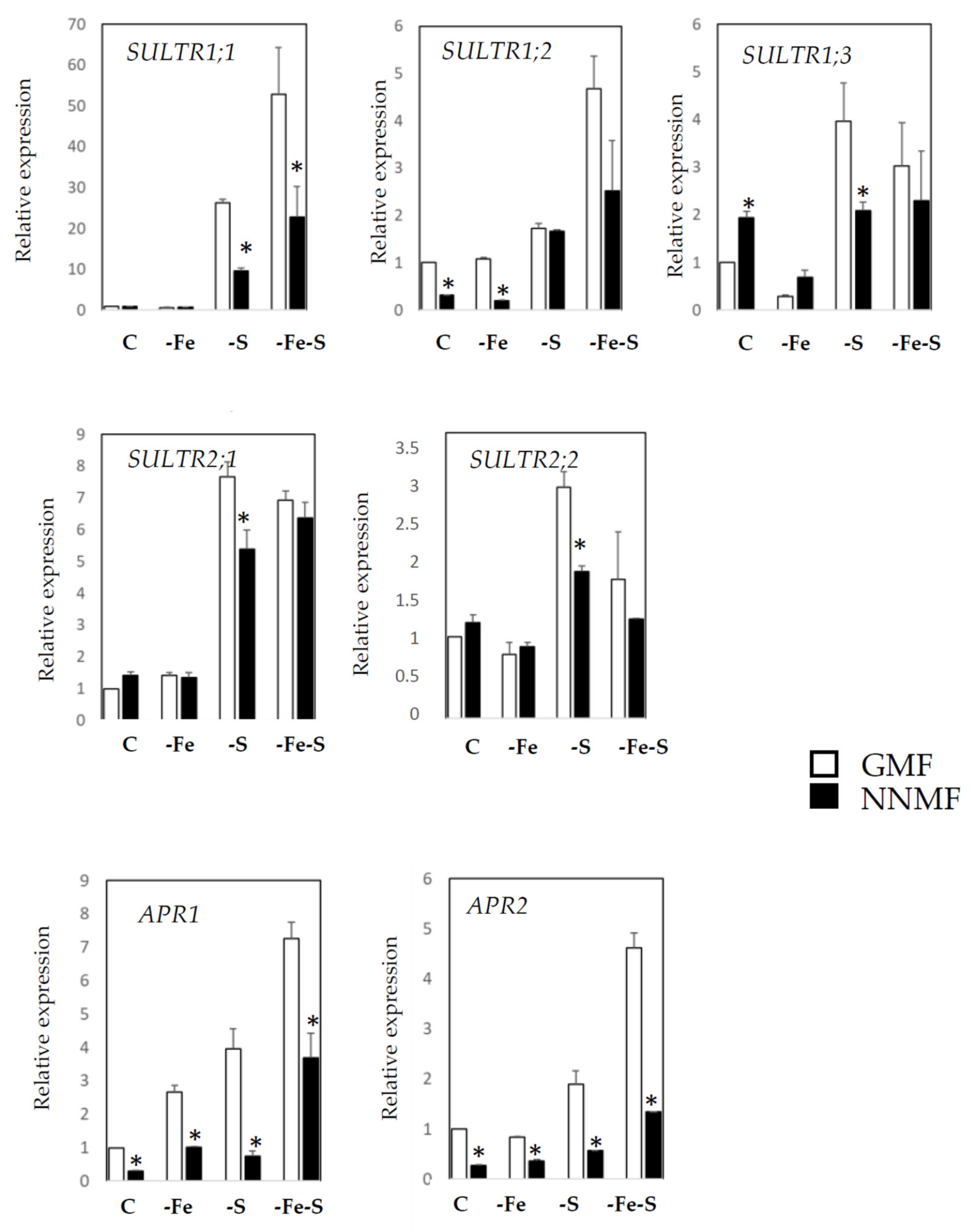
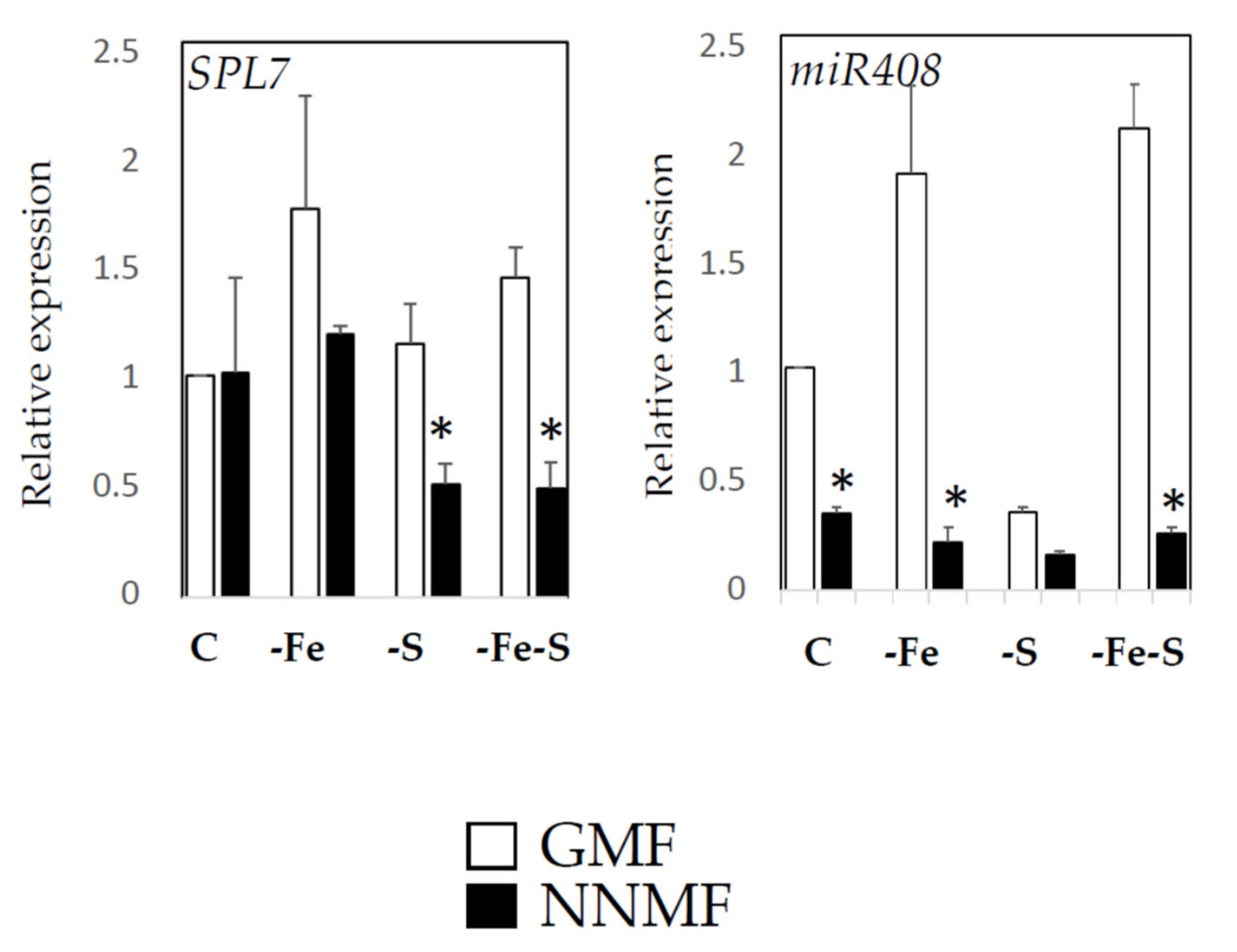
2.2. GMF Affects the Fe/S Interplay as Revealed by the Expression of Fe- and S-Responsive Genes
2.3. GMF Affects the Impact of Fe and S Availability on Metal Accumulation in Plant Tissues
2.4. GMF Affects Cu Homeostasis Depending on the Plant Nutritional Status
3. Discussion
4. Materials and Methods
4.1. Plant Materials, Media Composition and Growing Conditions
4.2. GMF Reduction System and Plant Exposure
4.3. Morphological Measurement
4.4. Metals Analysis in Plant Tissues
4.5. Anions Analysis by Capillary Electrophoresis (CE)
4.6. RNA Purification from Arabidopsis Root Tissues
4.7. Quantitative Real-Time PCR (qPCR)
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ACT1 | ACTIN1 |
| AHA2 | PLASMA MEMBRANE PROTON ATPase 2 |
| amiRNA | artificial microRNA |
| APR1 | ADENOSINE PHOSPHO SULFATE REDUCTASE 1 |
| APR2 | ADENOSINE PHOSPHO SULFATE REDUCTASE 2 |
| BTS | BRUTUS |
| C | Full nutrient condition |
| CE | Capillary Electrophoresis |
| eEF1Balpha2 | ELONGATION FACTOR 1B ALPHA SUBUNIT 2 |
| -Fe | Absence of iron |
| -Fe-S | Combined Fe and S deficiency |
| FIT | FE-DEFICIENCY INDUCED |
| FRO2 | FERRIC REDUCTION OXIDASE 2 |
| GAPC2 | CYTOPLASMIC GLYCERALDEHYDE-3-PHOSPHATE DEHYDROGENASE |
| GMF | Geomagnetic field |
| ICP-MS | Inductively Coupled Plasma-Mass Spectrometry |
| IRT1 | IRON-REGULATED TRANSPORTER 1 |
| MF | Magnetic field |
| MIR408 | microRNA408 |
| NA | Nutrient availability |
| NNMF | Near null magnetic field |
| PHT1 | Phosphate transporter 1 |
| PYE | POPEYE, bHLH47 |
| qPCR | Quantitative Real-Time PCR |
| RL | Root length |
| -S | Absence of sulfur |
| SA | Shoot area |
| SULTR 1;1 | SULPHATE TRANSPORTER 1;1 |
| SULTR 1;2 | SULPHATE TRANSPORTER 1;2 |
| SULTR 1;3 | SULPHATE TRANSPORTER 1;3 |
| SULTR 2;1 | SULPHATE TRANSPORTER 2;1 |
| SULTR 2;2 | SULPHATE TRANSPORTER 2;2 |
| UBP6 | UBIQUITIN SPECIFIC PROTEASE 6 |
References
- Vigani, G.; Costa, A. Harnessing the new emerging imaging technologies to uncover the role of ca2+ signalling in plant nutrient homeostasis. Plant Cell Environ. 2019, 42, 2885–2901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robe, K.; Gao, F.; Bonillo, P.; Tissot, N.; Gaymard, F.; Fourcroy, P.; Izquierdo, E.; Dubos, C. Sulphur availability modulates arabidopsis thaliana responses to iron deficiency. PLoS ONE 2020, 15, e0237998. [Google Scholar] [CrossRef] [PubMed]
- Muneer, S.; Lee, B.-R.; Kim, K.-Y.; Park, S.-H.; Zhang, Q.; Kim, T.-H. Involvement of sulphur nutrition in modulating iron deficiency responses in photosynthetic organelles of oilseed rape (brassica napus l.). Photosynth. Res. 2014, 119, 319–329. [Google Scholar] [CrossRef] [PubMed]
- Zuchi, S.; Cesco, S.; Varanini, Z.; Pinton, R.; Astolfi, S. Sulphur deprivation limits fe-deficiency responses in tomato plants. Planta 2009, 230, 85–94. [Google Scholar] [CrossRef] [Green Version]
- Astolfi, S.; Zuchi, S.; Hubberten, H.-M.; Pinton, R.; Hoefgen, R. Supply of sulphur to s-deficient young barley seedlings restores their capability to cope with iron shortage. J. Exp. Bot. 2009, 61, 799–806. [Google Scholar] [CrossRef] [Green Version]
- Astolfi, S.; Celletti, S.; Vigani, G.; Mimmo, T.; Cesco, S. Interaction between sulfur and iron in plants. Front. Plant Sci. 2021, 12, 670308. [Google Scholar] [CrossRef]
- Islam, M.; Maffei, M.E.; Vigani, G. The geomagnetic field is a contributing factor for an efficient iron uptake in arabidopsis thaliana. Front. Plant Sci. 2020, 11, 325. [Google Scholar] [CrossRef] [Green Version]
- Maffei, M.E. Magnetic field effects on plant growth, development, and evolution. Front. Plant Sci. 2014, 5. [Google Scholar] [CrossRef] [Green Version]
- Occhipinti, A.; De Santis, A.; Maffei, M.E. Magnetoreception: An unavoidable step for plant evolution? Trends Plant Sci 2014, 19, 1–4. [Google Scholar] [CrossRef]
- Bertea, C.M.; Narayana, R.; Agliassa, C.; Rodgers, C.T.; Maffei, M.E. Geomagnetic field (gmf) and plant evolution: Investigating the effects of GMF reversal on Arabidospis thaliana development and gene expression. J. Vis. Exp. 2015, 105, e53286. [Google Scholar] [CrossRef] [Green Version]
- Paponov, I.A.; Fliegmann, J.; Narayana, R.; Maffei, M.E. Differential root and shoot magnetoresponses in arabidopsis thaliana. Sci. Rep. 2021, 11, 9195. [Google Scholar] [CrossRef]
- Agliassa, C.; Narayana, R.; Bertea, C.M.; Rodgers, C.T.; Maffei, M.E. Reduction of the geomagnetic field delays arabidopsis thaliana flowering time through downregulation of flowering-related genes. Bioelectromagnetics 2018, 39, 361–374. [Google Scholar] [CrossRef]
- Narayana, R.; Fliegmann, J.; Paponov, I.; Maffei, M.E. Reduction of geomagnetic field (gmf) to near null magnetic field (nnmf) affects arabidopsis thaliana root mineral nutrition. Life Sci. Space Res. 2018, 19, 43–50. [Google Scholar] [CrossRef]
- Islam, M.; Vigani, G.; Maffei, M.E. The geomagnetic field (gmf) modulates nutrient status and lipid metabolism during arabidopsis thaliana plant development. Plants 2020, 9, 1729. [Google Scholar]
- Sorin, E.; Etienne, P.; Maillard, A.; Zamarreño, A.M.; Garcia-Mina, J.M.; Arkoun, M.; Jamois, F.; Cruz, F.; Yvin, J.C.; Ourry, A. Effect of sulphur deprivation on osmotic potential components and nitrogen metabolism in oilseed rape leaves: Identification of a new early indicator. J Exp Bot 2015, 66, 6175–6189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernal, M.; Casero, D.; Singh, V.; Wilson, G.T.; Grande, A.; Yang, H.; Dodani, S.C.; Pellegrini, M.; Huijser, P.; Connolly, E.L.; et al. Transcriptome sequencing identifies spl7-regulated copper acquisition genes fro4/fro5 and the copper dependence of iron homeostasis in arabidopsis. Plant Cell 2012, 24, 738–761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrió-Seguí, À.; Ruiz-Rivero, O.; Villamayor-Belinchón, L.; Puig, S.; Perea-García, A.; Peñarrubia, L. The altered expression of microrna408 influences the arabidopsis response to iron deficiency. Front. Plant Sci. 2019, 10, 324. [Google Scholar] [CrossRef] [PubMed]
- Etienne, P.; Sorin, E.; Maillard, A.; Gallardo, K.; Arkoun, M.; Guerrand, J.; Cruz, F.; Yvin, J.C.; Ourry, A. Assessment of sulfur deficiency under field conditions by single measurements of sulfur, chloride and phosphorus in mature leaves. Plants 2018, 7, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaji, N.; Takemoto, Y.; Miyaji, T.; Mitani-Ueno, N.; Yoshida, K.T.; Ma, J.F. Reducing phosphorus accumulation in rice grains with an impaired transporter in the node. Nature 2017, 541, 92–95. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Frank, T.; Tan, Y.; Zhou, C.; Jabnoune, M.; Arpat, A.B.; Cui, H.; Huang, J.; He, Z.; Poirier, Y.; et al. Disruption of ossultr3;3 reduces phytate and phosphorus concentrations and alters the metabolite profile in rice grains. New Phytol. 2016, 211, 926–939. [Google Scholar] [CrossRef] [Green Version]
- Essigmann, B.; Güler, S.; Narang, R.A.; Linke, D.; Benning, C. Phosphate availability affects the thylakoid lipid composition and the expression of sqd1, a gene required for sulfolipid biosynthesis in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 1998, 95, 1950–1955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Härtel, H.; Essigmann, B.; Lokstein, H.; Hoffmann-Benning, S.; Peters-Kottig, M.; Benning, C. The phospholipid-deficient pho1 mutant of arabidopsis thaliana is affected in the organization, but not in the light acclimation, of the thylakoid membrane. Biochim. Biophys. Acta 1998, 1415, 205–218. [Google Scholar] [CrossRef] [Green Version]
- Briat, J.F.; Rouached, H.; Tissot, N.; Gaymard, F.; Dubos, C. Integration of p, s, fe, and zn nutrition signals in arabidopsis thaliana: Potential involvement of phosphate starvation response 1 (phr1). Front. Plant Sci. 2015, 6, 290. [Google Scholar] [CrossRef] [Green Version]
- Oh, Y.J.; Kim, H.; Seo, S.H.; Hwang, B.G.; Chang, Y.S.; Lee, J.; Lee, D.W.; Sohn, E.J.; Lee, S.J.; Lee, Y.; et al. Cytochrome b5 reductase 1 triggers serial reactions that lead to iron uptake in plants. Mol. Plant 2016, 9, 501–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rouached, H. Multilevel coordination of phosphate and sulfate homeostasis in plants. Plant Signal. Behav. 2011, 6, 952–955. [Google Scholar] [CrossRef] [Green Version]
- Yamasaki, H.; Hayashi, M.; Fukazawa, M.; Kobayashi, Y.; Shikanai, T. Squamosa promoter binding protein-like7 is a central regulator for copper homeostasis in arabidopsis. Plant Cell 2009, 21, 347–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Wang, C.; Xu, H.; Shi, X.; Zhen, W.; Hu, Z.; Huang, J.; Zheng, Y.; Huang, P.; Zhang, K.X.; et al. Hy5 contributes to light-regulated root system architecture under a root-covered culture system. Front. Plant Sci. 2019, 10, 1490. [Google Scholar] [CrossRef] [Green Version]
- Koprivova, A.; Calderwood, A.; Lee, B.R.; Kopriva, S. Do pft1 and hy5 interact in regulation of sulfate assimilation by light in arabidopsis? FEBS Lett. 2014, 588, 1116–1121. [Google Scholar] [CrossRef] [Green Version]
- Agliassa, C.; Narayana, R.; Christie, J.M.; Maffei, M.E. Geomagnetic field impacts on cryptochrome and phytochrome signaling. J. Photochem. Photobiol. B Biol. 2018, 185, 32–40. [Google Scholar] [CrossRef]
- Gruber, B.D.; Giehl, R.F.; Friedel, S.; von Wirén, N. Plasticity of the arabidopsis root system under nutrient deficiencies. Plant Physiol. 2013, 163, 161–179. [Google Scholar] [CrossRef] [Green Version]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. Nih image to imagej: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Andersen, C.L.; Jensen, J.L.; Ørntoft, T.F. Normalization of real-time quantitative reverse transcription-pcr data: A model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets. Cancer Res. 2004, 64, 5245–5250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.-Y.; Klatte, M.; Jakoby, M.; Bäumlein, H.; Weisshaar, B.; Bauer, P. Iron deficiency-mediated stress regulation of four subgroup ib bhlh genes in arabidopsis thaliana. Planta 2007, 226, 897–908. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| C | -Fe | -S | -Fe-S | |||||
|---|---|---|---|---|---|---|---|---|
| ROOT | GMF | NNMF | GMF | NNMF | GMF | NNMF | GMF | NNMF |
| Mn | 24.05 ± 13.80 | 68.51 ± 34.10 | 178.03 ± 24.95 | 895.90 * ± 30.70 | 48.15 ± 10.98 | 72.68 ± 24.71 | 138.35 ± 70.94 | 326.32 * ± 81.27 |
| Fe | 323.46 ± 44.21 | 257.38 * ± 34.78 | 30.68 ± 6.54 | 159.98 * ± 74.49 | 339.40 ± 25.16 | 300.49 ± 12.98 | 72.63 ± 23.32 | 148.84 * ± 20.26 |
| Cu | 14.50 ± 4.32 | 20.36 ± 3.09 | 8.41 ± 3.85 | 33.07 * ± 2.58 | 11.45 ± 3.73 | 19.31 * ± 2.73 | 10.31 ± 1.51 | 30.82 * ± 8.49 |
| Zn | 453.54 ± 203.01 | 552.89 ± 107.70 | 2427.34 ± 399.90 | 5213.27 * ± 607.61 | 538.89 ± 144.71 | 723.83 ± 284.09 | 4887.01 ± 980.71 | 9200.66 * ± 1502.25 |
| Mo | 7.56 ± 3.50 | 5.28 ± 2.61 | 1.01 ± 0.24 | 10.91 * ± 0.16 | 54.46 ± 10.33 | 55.24 ± 12.52 | 25.83 ± 6.41 | 40.80 ± 17.51 |
| SHOOT | ||||||||
| Mn | 13.83 ± 2.22 | 69.26 ± 34.36 | 220.81 ± 25.24 | 112.53 ± 16.95 | 62.90 ± 20.91 | 66.86 ± 14.47 | 37.76 ± 16.56 | 84.49 ± 33.10 |
| Fe | 162.01 ± 23.21 | 134.55 ± 8.89 | 65.72 ± 8.65 | 87.60 * ± 5.39 | 187.49 ± 19.83 | 169.83 ± 15.23 | 83.07 ± 15.47 | 72.40 ± 18.47 |
| Cu | 10.47 ± 1.46 | 8.51 ± 1.25 | 13.02 ± 3.82 | 19.78 ± 3.87 | 7.90 ± 1.01 | 9.89 ± 0.98 | 11.34 ± 2.63 | 11.15 ± 4.14 |
| Zn | 185.05 ± 61.35 | 177.73 ± 21.59 | 995.94 ± 69.21 | 1005.65 ± 101.95 | 259.17 ± 35.12 | 281.31 ± 13.03 | 765.52 ± 404.56 | 710.54 ± 218.09 |
| Mo | 4.47 ± 1.52 | 5.77 ± 1.62 | 6.88 ± 1.77 | 9.92 ± 2.35 | 136.66 ± 20.36 | 120.91 ± 25.37 | 122.94 ± 46.47 | 95.85 ± 30.37 |
| wt | spl7 | ami4/5 | ami5 | |||||
|---|---|---|---|---|---|---|---|---|
| GMF | NNMF | GMF | NNMF | GMF | NNMF | GMF | NNMF | |
| Mn | 94.99 ± 21.41 | 224.91 ± 45.66 | 7.93 ± 1.44 | 16.21 * ± 3.45 | 60.46 ± 11.78 | 121.89 * ± 39.85 | 58.51 ± 15.47 | 98.27 ± 28.12 |
| Fe | 390.01 ± 40.44 | 317.11 * ± 21.44 | 184.23 ± 19.84 | 61.4 * ± 11.74 | 344.18 ± 58.74 | 424.28 ± 69.45 | 188.86 ± 36.75 | 716.72 * ± 120.56 |
| Cu | 11.13 ± 2.12 | 13.89 ± 1.85 | 6.2 ± 0.87 | 19.6 * ± 3.66 | 20.54 ± 4.83 | 8.70 * ± 2.14 | 19.34 ± 4.36 | 48.33 * ± 19.45 |
| Zn | 327.97 ± 59.74 | 353.90 ± 78.63 | 448.17 ± 44.62 | 162.80 ± 63.14 | 579.30 ± 31.56 | 252.07 ± 98.12 | 540.31 ± 65.12 | 947.73 ± 105.74 |
| Mo | 1.35 ± 0.12 | 8.01 ± 1.87 | 1.93 ± 0.65 | 2.00 ± 0.57 | 4.69 ± 1.63 | 1.81 ± 0.9 | 27.36 ± 5.44 | 7.35 ± 2.85 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vigani, G.; Islam, M.; Cavallaro, V.; Nocito, F.F.; Maffei, M.E. Geomagnetic Field (GMF)-Dependent Modulation of Iron-Sulfur Interplay in Arabidopsis thaliana. Int. J. Mol. Sci. 2021, 22, 10166. https://doi.org/10.3390/ijms221810166
Vigani G, Islam M, Cavallaro V, Nocito FF, Maffei ME. Geomagnetic Field (GMF)-Dependent Modulation of Iron-Sulfur Interplay in Arabidopsis thaliana. International Journal of Molecular Sciences. 2021; 22(18):10166. https://doi.org/10.3390/ijms221810166
Chicago/Turabian StyleVigani, Gianpiero, Monirul Islam, Viviana Cavallaro, Fabio F. Nocito, and Massimo E. Maffei. 2021. "Geomagnetic Field (GMF)-Dependent Modulation of Iron-Sulfur Interplay in Arabidopsis thaliana" International Journal of Molecular Sciences 22, no. 18: 10166. https://doi.org/10.3390/ijms221810166
APA StyleVigani, G., Islam, M., Cavallaro, V., Nocito, F. F., & Maffei, M. E. (2021). Geomagnetic Field (GMF)-Dependent Modulation of Iron-Sulfur Interplay in Arabidopsis thaliana. International Journal of Molecular Sciences, 22(18), 10166. https://doi.org/10.3390/ijms221810166