
The Effects of Melatonin Administration on Intestinal Injury Caused by Abdominal Irradiation from Mice



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
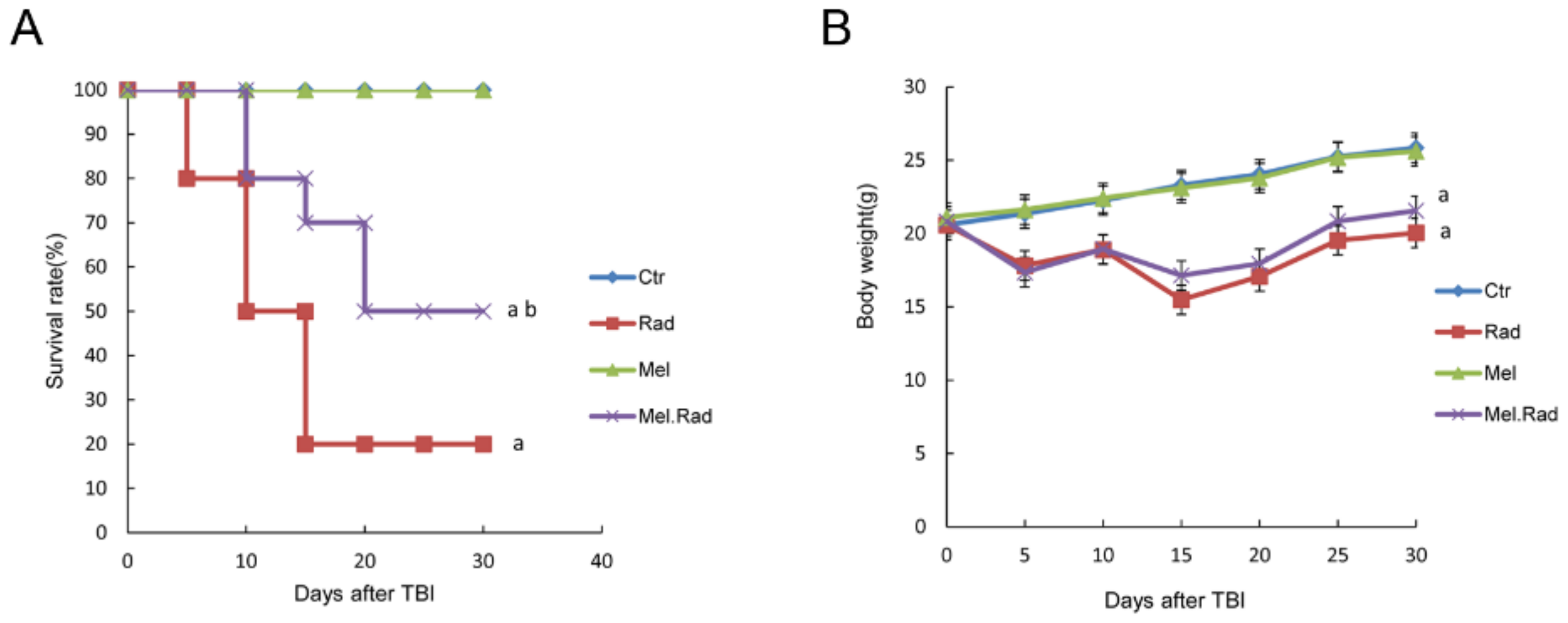
2.1. Melatonin Promoted the Survival of Mice Exposed to Total Body Irradiation (TBI)
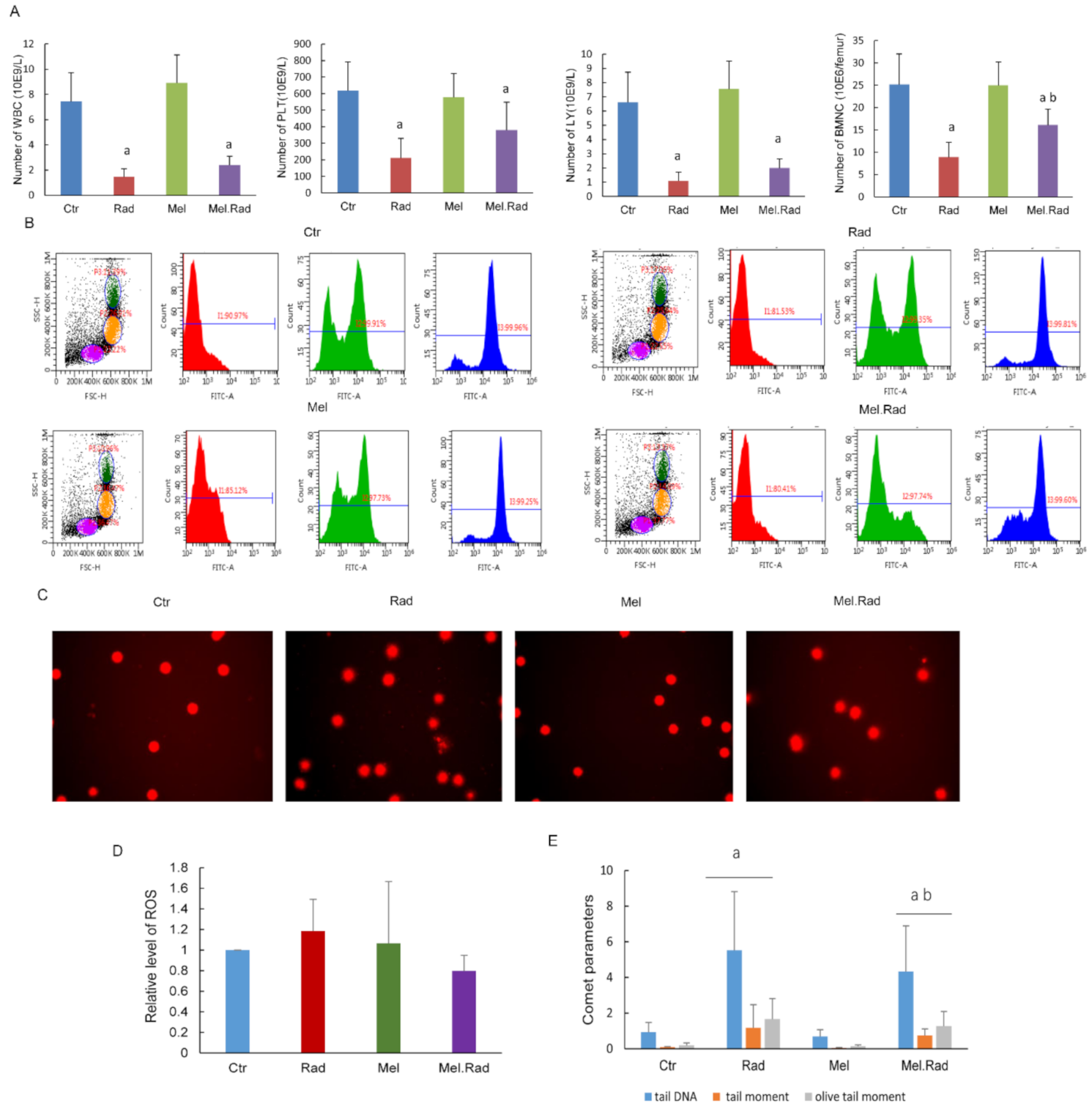
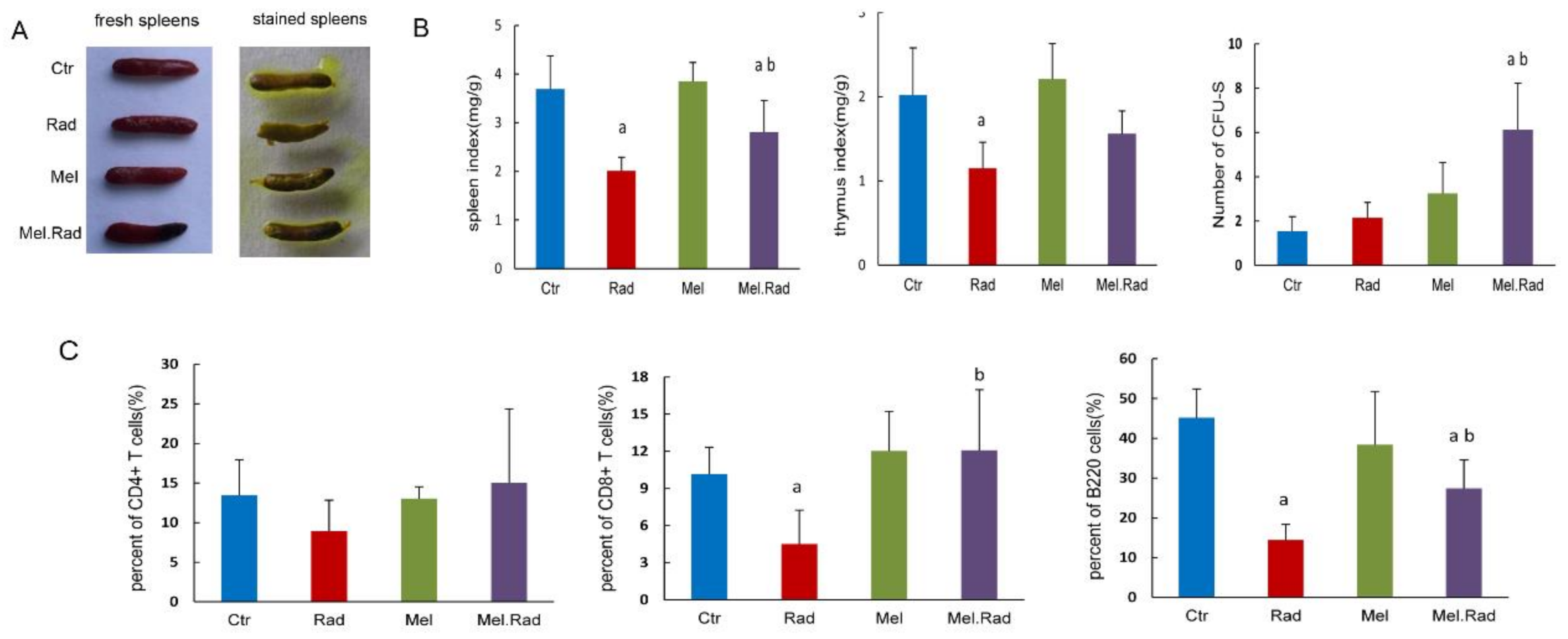
2.2. Melatonin Mitigated the Hematopoietic System Damage of Mice Exposed to TBI
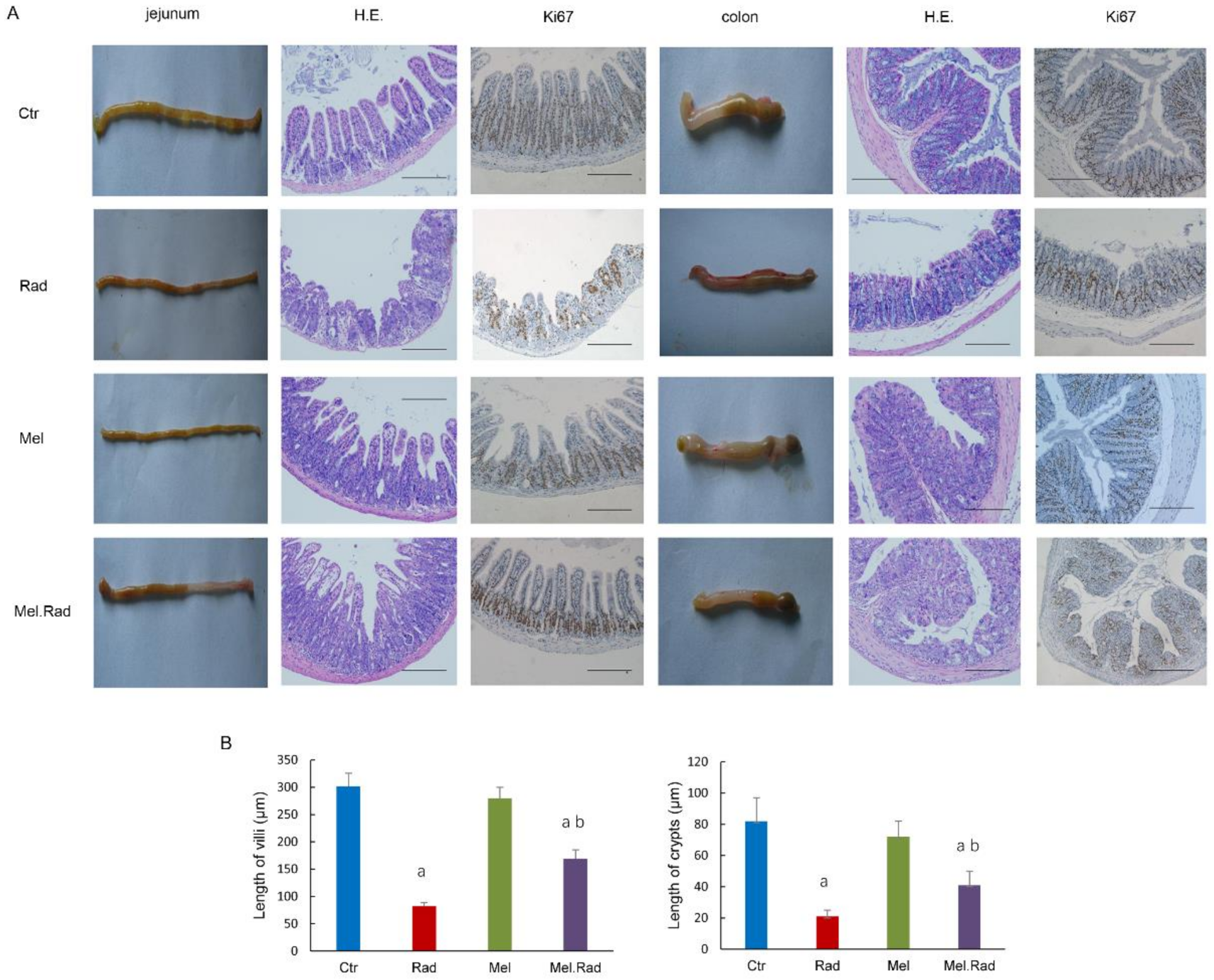
2.3. Melatonin Alleviated the Damage to the Intestines of Mice Exposed to ABI
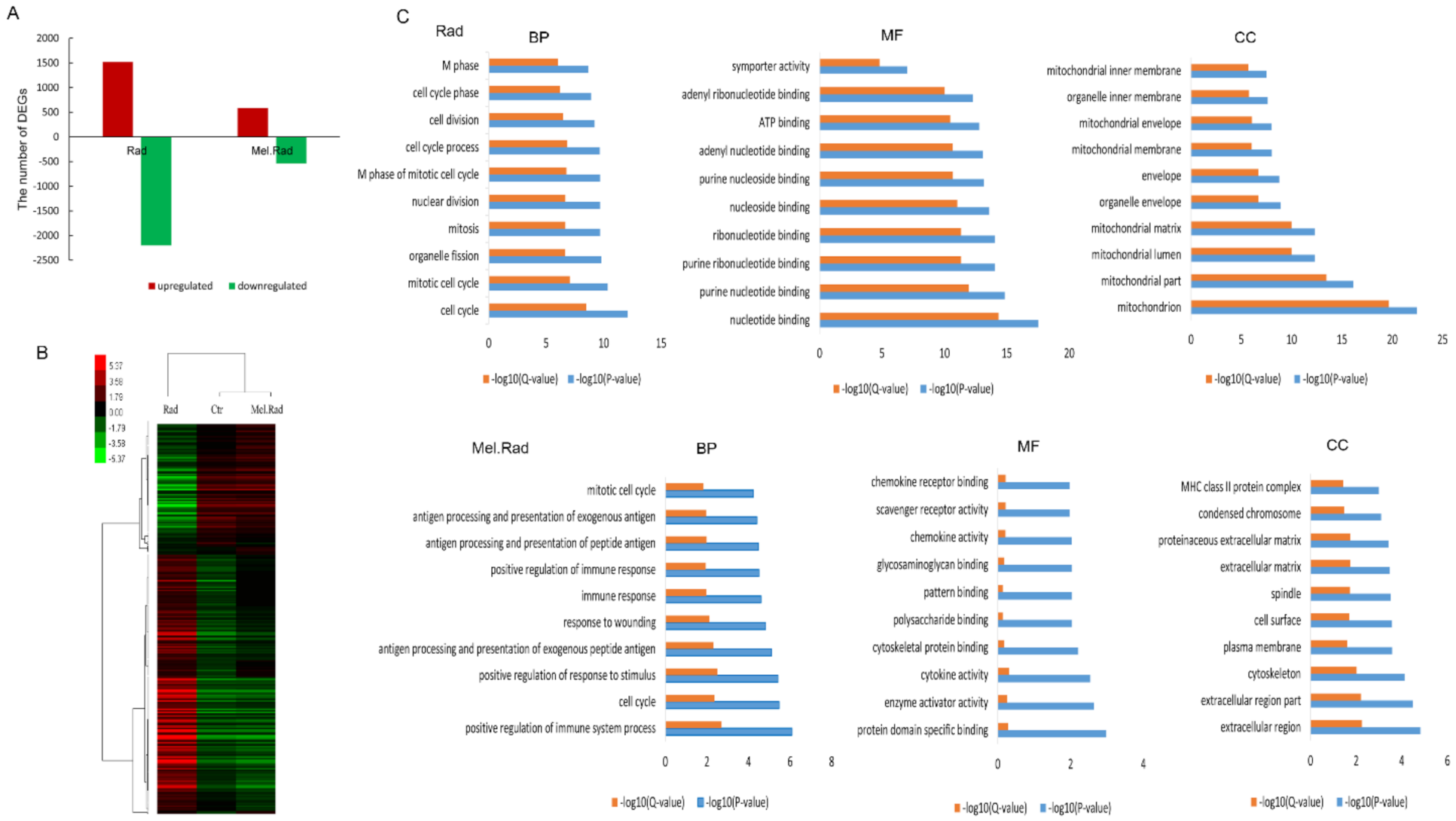
2.4. GO Analysis of Gene Expression of the Small Intestine from Mice Exposed to ABI
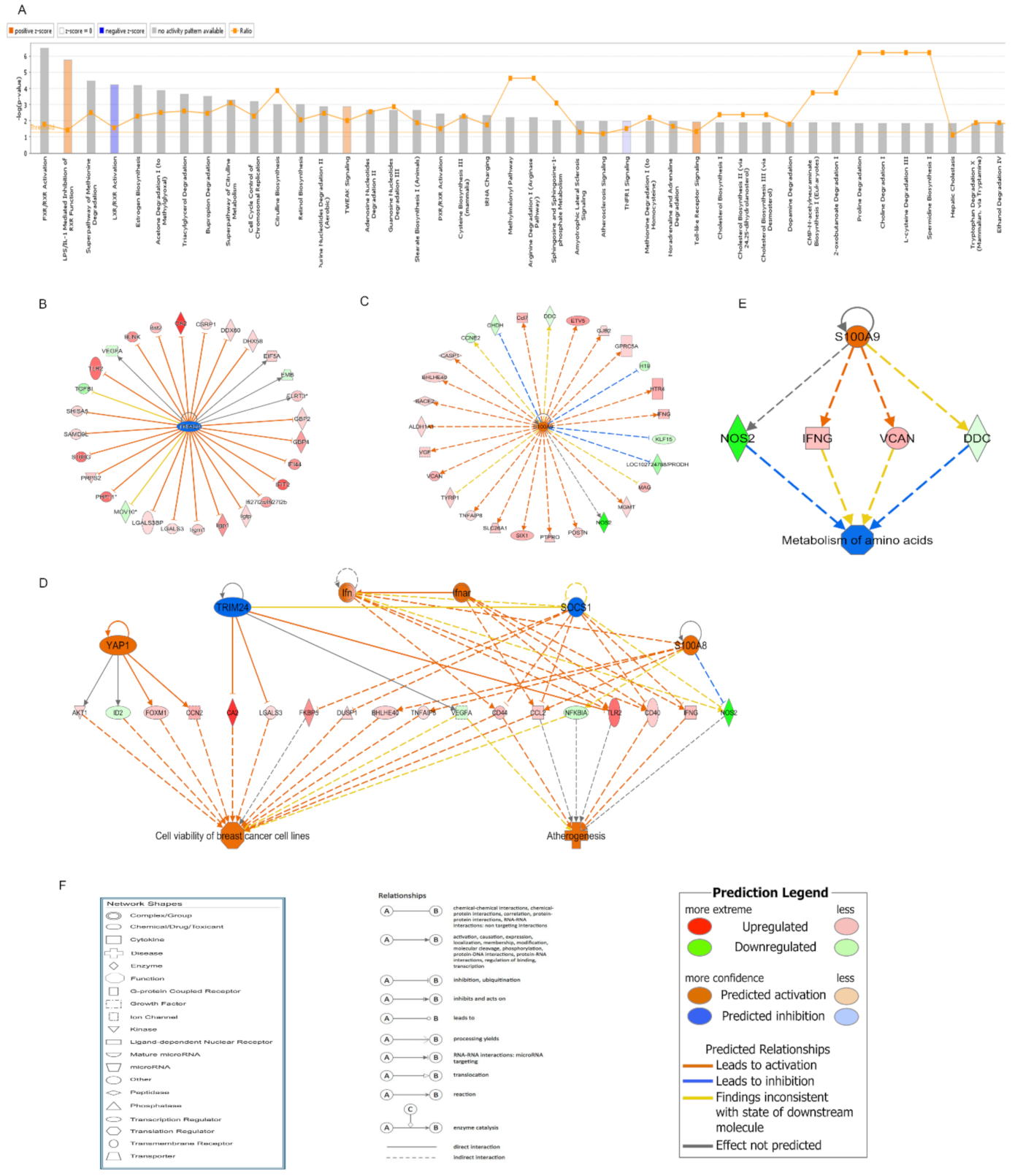
2.5. IPA of the Small Intestine from Mice Exposed to ABI
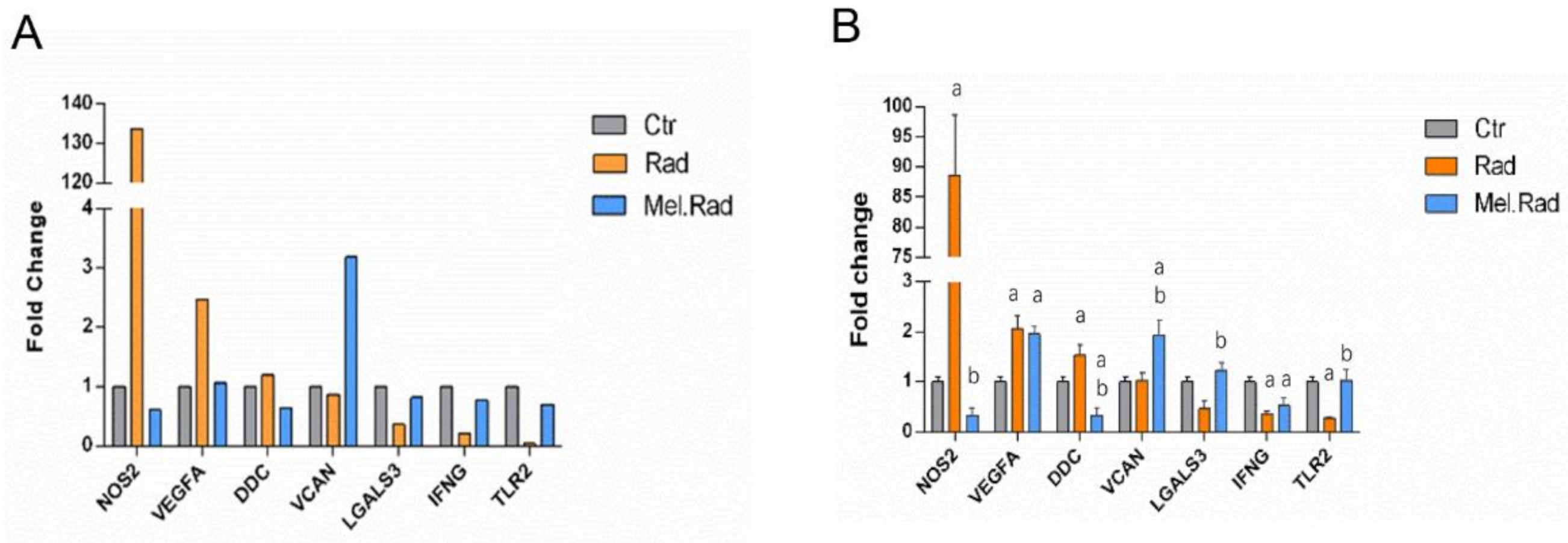
2.6. qRT-PCR Validation of the Candidate DEGs Involved in the Regulatory Network of TRIM24 and S100A9
2.7. Validation of the Effect of Melatonin Treatment on the Immunity
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. Radiation Exposure in Mice
4.3. Administration of Melatonin
4.4. Experimental Design
4.5. Analysis of 30-Day Survival Rate
4.6. Pathological Analysis
4.7. Immunohistochemical Analysis
4.8. Peripheral Blood Cell and Bone-Marrow Cell Counts
4.9. Reactive Oxygen Species (ROS) Level Assay
4.10. Analysis of DNA Damage in Lymphocytes and Lymphocyte Phenotype
4.11. cDNA Microarray Analysis
4.12. GO Annotation and IPA Analysis
4.13. qRT-PCR
4.14. Calculation of CFU-S and Spleen/Thymus Coefficient
4.15. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Citrin, D.E. Recent developments in radiotherapy. N. Engl. J. Med. 2017, 377, 1065–1075. [Google Scholar] [CrossRef] [PubMed]
- Umar, S. Intestinal stem cells. Curr. Gastroenterol. Rep. 2010, 12, 340–348. [Google Scholar] [CrossRef] [PubMed]
- Mettler, F.A., Jr.; Brenner, D.; Coleman, C.N.; Kaminski, J.M.; Kennedy, A.R.; Wagner, L.K. Can radiation risks to patients be reduced without reducing radiation exposure? The status of chemical radioprotectants. AJR Am. J. Roentgenol. 2011, 196, 616–618. [Google Scholar] [CrossRef]
- King, M.; Joseph, S.; Albert, A.; Thomas, T.V.; Nittala, M.R.; Woods, W.C.; Vijayakumar, S.; Packianathan, S. Use of amifostine for cytoprotection during radiation therapy: A Review. Oncology 2020, 98, 61–80. [Google Scholar] [CrossRef] [PubMed]
- Vijayalaxmi; Reiter, R.J.; Tan, D.X.; Herman, T.S.; Thomas, C.R., Jr. Melatonin as a radioprotective agent: A review. Int. J. Radiat. Oncol. Biol. Phys. 2004, 59, 639–653. [Google Scholar] [CrossRef]
- Claustrat, B.; Leston, J. Melatonin: Physiological effects in humans. Neurochirurgie 2015, 61, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Zetner, D.; Andersen, L.P.; Rosenberg, J. Melatonin as protection against radiation injury: A systematic review. Drug Res. Stuttg. 2016, 66, 281–296. [Google Scholar] [CrossRef] [Green Version]
- Das, B.; Bennett, P.V.; Cutler, N.C.; Sutherland, J.C.; Sutherland, B.M. Melatonin protects human cells from clustered damages, killing and acquisition of soft agar growth induced by X-rays or 970 MeV/n Fe ions. Int. J. Radiat. Biol. 2011, 87, 545–555. [Google Scholar] [CrossRef]
- Zhou, G.; Kawata, T.; Furusawa, Y.; Aoki, M.; Hirayama, R.; Ando, K.; Ito, H. Protective effects of melatonin against low- and high-LET irradiation. J. Radiat. Res. 2006, 47, 175–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gehart, H.; Clevers, H. Tales from the crypt: New insights into intestinal stem cells. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 19–34. [Google Scholar] [CrossRef] [PubMed]
- Shao, Z.M.; Wang, K.K.; Zhang, S.Y.; Yuan, J.L.; Liao, X.M.; Wu, C.X.; Zou, Y.; Ha, Y.P.; Shen, Z.Y.; Guo, J.L.; et al. Ingenuity pathway analysis of differentially expressed genes involved in signaling pathways and molecular networks in RhoE gene-edited cardiomyocytes. Int. J. Mol. Med. 2020, 46, 1225–1238. [Google Scholar] [CrossRef]
- Shao, L.J.; Luo, Y.; Zhou, D.H. Hematopoietic stem cell injury induced by ionizing radiation. Antioxid. Redox. Signal. 2014, 20, 1447–1462. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.Y.; Jeong, E.K.; Ju, M.K.; Jeon, H.M.; Kim, M.Y.; Kim, C.H.; Park, H.G.; Han, S.I.; Kang, H.S. Induction of metastasis, cancer stem cell phenotype, and oncogenic metabolism in cancer cells by ionizing radiation. Mol. Cancer 2017, 16, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mihandoost, E.; Shirazi, A.; Mahdavi, S.R.; Aliasgharzadeh, A. Consequences of lethal-whole-body gamma radiation and possible ameliorative role of melatonin. Sci. World J. 2014, 2014, 621570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Du, L.Q.; Wang, Y.; Xu, C.; Sun, Z.J.; Fu, Y.; Yang, B.; Wang, Y.Y.; Mu, C.J.; Fan, S.J.; et al. The development and biological characteristics of a novel potentially radioresistant inbred mouse strain. Mol. Med. Rep. 2017, 15, 759–767. [Google Scholar] [CrossRef] [Green Version]
- Bondy, S.C.; Campbell, A. Melatonin and regulation of immune function: Impact on numerous diseases. Curr. Aging Sci. 2020, 13, 92–101. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Wu, R.; Gaspar, J.M.; Sargsyan, D.; Su, Z.Y.; Zhang, C.; Gao, L.; Cheng, D.; Li, W.; Wang, C.; et al. DNA methylome and transcriptome alterations and cancer prevention by curcumin in colitis-accelerated colon cancer in mice. Carcinogenesis 2018, 39, 669–680. [Google Scholar] [CrossRef]
- Lim, K.H.; Staudt, L.M. Toll-like receptor signaling. Cold Spring Harb. Perspect. Biol. 2013, 5, a011247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, M.; Xiao, H.W.; Li, Y.; Zhou, L.X.; Zhao, S.Y.; Luo, D.; Zheng, Q.S.; Dong, J.L.; Zhao, Y.; Zhang, X.; et al. Faecal microbiota transplantation protects against radiation-induced toxicity. EMBO Mol. Med. 2017, 9, 448–461. [Google Scholar] [CrossRef]
- Hatakeyama, S. TRIM family proteins: Roles in autophagy, immunity, and carcinogenesis. Trends Biochem. Sci. 2017, 42, 297–311. [Google Scholar] [CrossRef]
- Appikonda, S.; Thakkar, K.N.; Barton, M.C. Regulation of gene expression in human cancers by TRIM24. Drug Discov. Today Technol. 2016, 19, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Pruenster, M.; Vogl, T.; Roth, J.; Sperandio, M. S100A8/A9: From basic science to clinical application. Pharmacol. Ther. 2016, 167, 120–131. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.W.; Song, R.; Wang, Z.Y.; Jing, Z.C.; Wang, S.X.; Ma, J. S100A8/A9 in inflammation. Front. Immunol. 2018, 9, 1298. [Google Scholar] [CrossRef] [PubMed]
- Oshikawa, K.; Sugiyama, Y. Regulation of toll-like receptor 2 and 4 gene expression in murine alveolar macrophages. Exp. Lung Res. 2003, 29, 401–412. [Google Scholar] [CrossRef] [PubMed]
- Anavi, S.; Tirosh, O. iNOS as a metabolic enzyme under stress conditions. Free Radic. Biol. Med. 2020, 146, 16–35. [Google Scholar] [CrossRef]
- Vijayalaxmi; Reiter, R.J.; Herman, T.S.; Meltz, M.L. Melatonin and radioprotection from genetic damage: In vivo/in vitro studies with human volunteers. Mutat. Res. 1996, 371, 221–228. [Google Scholar] [CrossRef]
- Nordlund, J.J.; Lerner, A.B. The effects of oral melatonin on skin color and on the release of pituitary hormones. J. Clin. Endocrinol. Metab. 1977, 45, 768–774. [Google Scholar] [CrossRef]
- Onseng, K.; Johns, N.P.; Khuayjarernpanishk, T.; Subongkot, S.; Priprem, A.; Hurst, C.; Johns, J. Beneficial effects of adjuvant melatonin in minimizing oral mucositis complications in head and neck cancer patients receiving concurrent chemoradiation. J. Altern. Complement. Med. 2017, 23, 957–963. [Google Scholar] [CrossRef]
- Sookprasert, A.; Johns, N.P.; Phunmanee, A.; Pongthai, P.; Cheawchanwattana, A.; Johns, J.; Konsil, J.; Plaimee, P.; Porasuphatana, S.; Jitpimolmard, S. Melatonin in patients with cancer receiving chemotherapy: A randomized, double-blind, placebo-controlled trial. Anticancer Res. 2014, 34, 7327–7337. [Google Scholar]
- Wang, Q.; Sun, Z.J.; Du, L.Q.; Xu, C.; Wang, Y.; Yang, B.; He, N.N.; Wang, J.H.; Ji, K.H.; Liu, Y.; et al. Melatonin sensitizes human colorectal cancer cells to γ-ray ionizing radiation in vitro and in vivo. Int. J. Mol. Sci. 2018, 19, 3974. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Schulte, B.A.; LaRue, A.C.; Ogawa, M.; Zhou, D. Total body irradiation selectively induces murine hematopoietic stem cell senescence. Blood 2006, 107, 358–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, X.H.; Wang, Q.; Wang, Y.; Du, L.Q.; Xu, C.; Liu, Q. Brusatol enhances theradiosensitivity of A549 cells by promoting ROS production and enhancing DNA damage. Int. J. Mol. Sci. 2016, 17, 997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Końca, K.; Lankoff, A.; Banasik, A.; Lisowska, H.; Kuszewski, T.; Góźdź, S.; Koza, Z.; Wojcik, A. A cross-platform public domain PC image-analysis program for the comet assay. Mutat. Res. 2003, 534, 15–20. [Google Scholar] [CrossRef]
- Li, D.G.; Wang, Y.Y.; Wu, H.Y.; Lu, L.; Wang, X.C.; Zhang, J.L.; Zhang, H.; Fan, S.J.; Fan, F.Y.; Zhou, D.H.; et al. The effects of p38 MAPK inhibition combined with G-CSF administration on the hematoimmune system in mice with irradiation injury. PLoS ONE 2013, 8, e62921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, W.Q.; Yang, F.J.; Shen, X.; Fan, S.J.; Liu, Q.; Wang, D.Z. Polysaccharide isolated from parmelia tinctorum ameliorates ionizing irradiation-induced damage in mice. J. Radiat. Res. 2014, 55, 641–647. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Q.; Wang, Y.; Du, L.; Xu, C.; Liu, Q.; Fan, S. The Effects of Melatonin Administration on Intestinal Injury Caused by Abdominal Irradiation from Mice. Int. J. Mol. Sci. 2021, 22, 9715. https://doi.org/10.3390/ijms22189715
Wang Q, Wang Y, Du L, Xu C, Liu Q, Fan S. The Effects of Melatonin Administration on Intestinal Injury Caused by Abdominal Irradiation from Mice. International Journal of Molecular Sciences. 2021; 22(18):9715. https://doi.org/10.3390/ijms22189715
Chicago/Turabian StyleWang, Qin, Yan Wang, Liqing Du, Chang Xu, Qiang Liu, and Saijun Fan. 2021. "The Effects of Melatonin Administration on Intestinal Injury Caused by Abdominal Irradiation from Mice" International Journal of Molecular Sciences 22, no. 18: 9715. https://doi.org/10.3390/ijms22189715
APA StyleWang, Q., Wang, Y., Du, L., Xu, C., Liu, Q., & Fan, S. (2021). The Effects of Melatonin Administration on Intestinal Injury Caused by Abdominal Irradiation from Mice. International Journal of Molecular Sciences, 22(18), 9715. https://doi.org/10.3390/ijms22189715