
Is It Possible to Create Antimicrobial Peptides Based on the Amyloidogenic Sequence of Ribosomal S1 Protein of P. aeruginosa?

 , , , ,
, , , , 
 , and
, and 
Abstract
:1. Introduction
2. Results
2.1. Prediction of the Secondary Structure and Antimicrobial Propensities of the R23R, R23L, R23R*, and R23L* Peptides
2.2. Experimental Validation of the Antibacterial Activity of Peptides
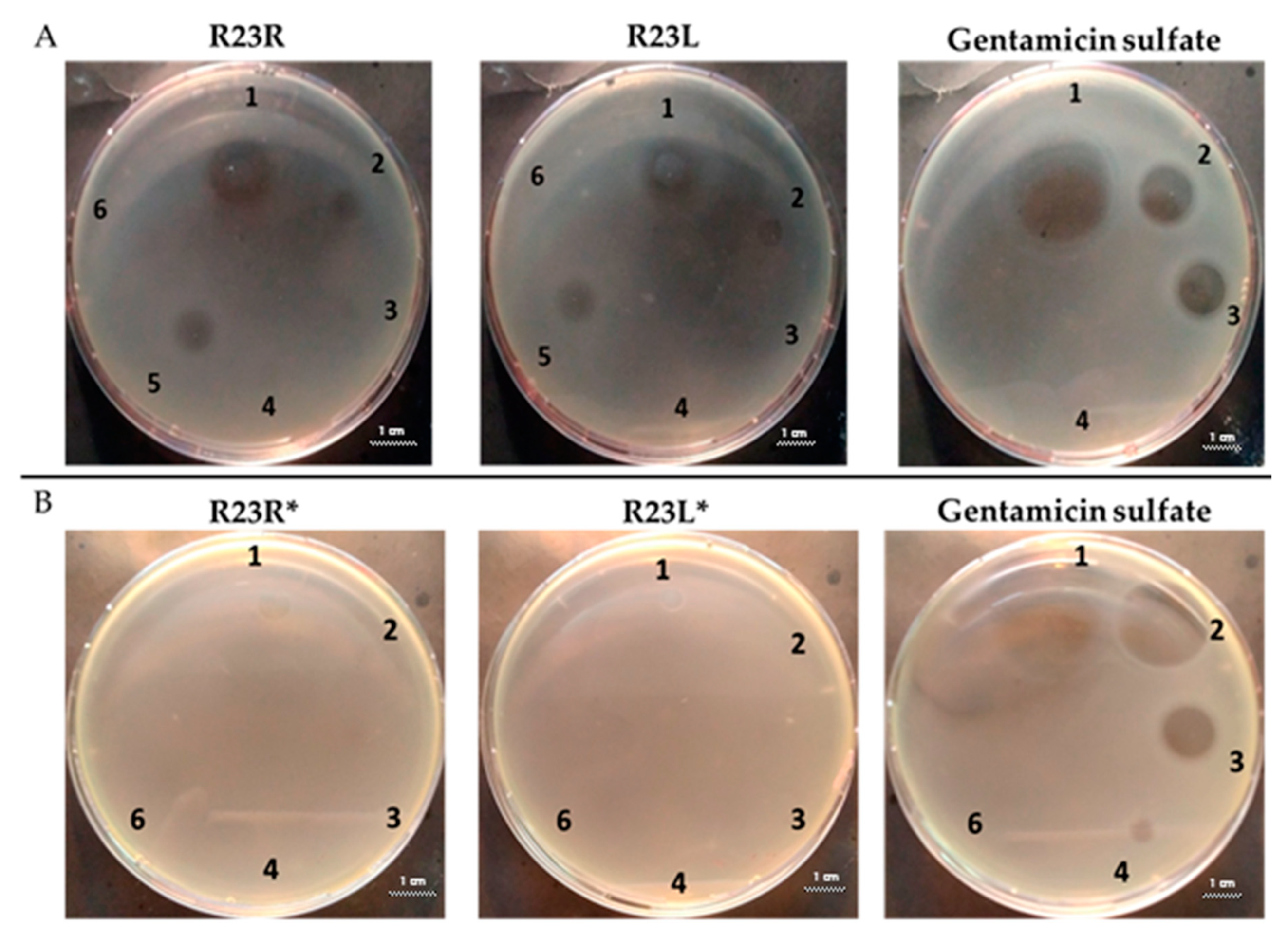
2.2.1. Determination of the Antibacterial Activity of Peptides on Agar
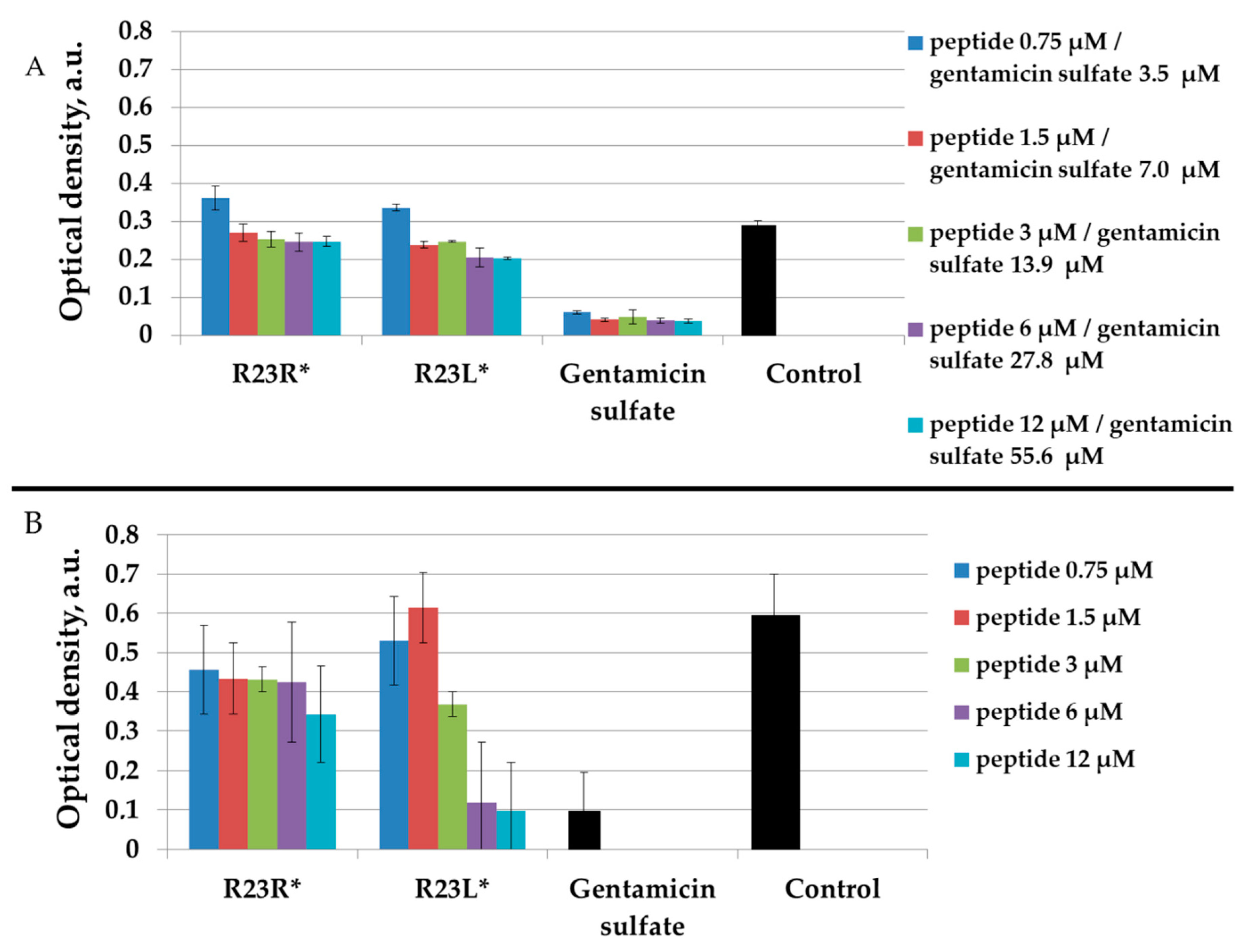
2.2.2. Measurement of the Antibacterial Activity of Peptides by Microdilution Technique
2.3. Toxicity of R23R and R23L
2.4. Amyloidogenic Properties of Synthesized Peptides
3. Discussions
4. Materials and Methods
4.1. Synthesis and Characterization of Peptides
4.1.1. Peptide Synthesis
4.1.2. Bioinformatic Analysis of Peptides
4.2. Antimicrobial Activity of Peptides
4.2.1. Determination of the Antibacterial Properties of Peptides on Agar
4.2.2. Determination of MIC by Broth Dilution Method
4.3. Cytotoxicity Assay
4.4. Thioflavin T Fluorescence Measurement
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AMP | antimicrobial peptide |
| ANN | artificial neural network |
| CPP | cell penetrating peptide |
| DA | discriminant analysis |
| DNN | deep neural network |
| LB | Luria-Bertani (medium) |
| MHB | Mueller-Hinton broth |
| MIC | Minimum inhibitory concentration |
| OB-fold | oligonucleotide/oligosaccharide-binding fold |
| RF | random forest |
| SVM | support vector machine |
| ThT | tioflavin T |
References
- Teixeira, M.C.; Carbone, C.; Sousa, M.C.; Espina, M.; Garcia, M.L.; Sanchez-Lopez, E.; Souto, E.B. Nanomedicines for the Delivery of Antimicrobial Peptides (AMPs). Nanomaterials 2020, 10, 560. [Google Scholar] [CrossRef] [Green Version]
- Honda, M.; Matsumoto, M.; Aizawa, M. Potential Application of Protamine for Antimicrobial Biomaterials in Bone Tissue Engineering. Int. J. Mol. Sci. 2020, 21, 4368. [Google Scholar] [CrossRef]
- Moretta, A.; Scieuzo, C.; Petrone, A.M.; Salvia, R.; Manniello, M.D.; Franco, A.; Lucchetti, D.; Vassallo, A.; Vogel, H.; Sgambato, A.; et al. Antimicrobial Peptides: A New Hope in Biomedical and Pharmaceutical Fields. Front. Cell. Infect. Microbiol. 2021, 11, 453. [Google Scholar] [CrossRef]
- Dey, S.; Gill, K.; Kumar, S.; Xess, I. Novel synthetic anti-fungal tripeptide effective against Candida krusei. Indian J. Med. Microbiol. 2015, 33, 110. [Google Scholar] [CrossRef]
- Ahmed, A.; Siman-Tov, G.; Hall, G.; Bhalla, N.; Narayanan, A. Human Antimicrobial Peptides as Therapeutics for Viral Infections. Viruses 2019, 11, 704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, N.; Wang, C.; Zhang, T.; Zhang, L.; Xue, C.; Feng, X.; Bi, C.; Shan, A. Bioactivity and Bactericidal Mechanism of Histidine-Rich β-Hairpin Peptide Against Gram-Negative Bacteria. Int. J. Mol. Sci. 2019, 20, 3954. [Google Scholar] [CrossRef] [Green Version]
- Burdukiewicz, M.; Sidorczuk, K.; Rafacz, D.; Pietluch, F.; Chilimoniuk, J.; Rödiger, S.; Gagat, P. Proteomic Screening for Prediction and Design of Antimicrobial Peptides with AmpGram. Int. J. Mol. Sci. 2020, 21, 4310. [Google Scholar] [CrossRef]
- Veltri, D.; Kamath, U.; Shehu, A. Deep learning improves antimicrobial peptide recognition. Bioinformatics 2018, 34, 2740–2747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waghu, F.H.; Barai, R.S.; Gurung, P.; Idicula-Thomas, S. CAMPR3: A database on sequences, structures and signatures of antimicrobial peptides. Nucleic Acids Res. 2016, 44, D1094–D1097. [Google Scholar] [CrossRef] [Green Version]
- Mulcahy, L.R.; Isabella, V.M.; Lewis, K. Pseudomonas aeruginosa Biofilms in Disease. Microb. Ecol. 2014, 68, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigo-Troyano, A.; Melo, V.; Marcos, P.J.; Laserna, E.; Peiro, M.; Suarez-Cuartin, G.; Perea, L.; Feliu, A.; Plaza, V.; Faverio, P.; et al. Pseudomonas aeruginosa in Chronic Obstructive Pulmonary Disease Patients with Frequent Hospitalized Exacerbations: A Prospective Multicentre Study. Respiration 2018, 96, 417–424. [Google Scholar] [CrossRef] [PubMed]
- Coleman, S.R.; Blimkie, T.; Falsafi, R.; Hancock, R.E.W. Multidrug Adaptive Resistance of Pseudomonas aeruginosa Swarming Cells. Antimicrob. Agents Chemother. 2019, 64, e01999-19. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.; Stapleton, F.; Summers, S.; Rice, S.A.; Willcox, M.D.P. Antibiotic Resistance Characteristics of Pseudomonas aeruginosa Isolated from Keratitis in Australia and India. Antibiotics 2020, 9, 600. [Google Scholar] [CrossRef] [PubMed]
- Shemyakin, I.G.; Firstova, V.V.; Fursova, N.K.; Abaev, I.V.; Filippovich, S.Y.; Ignatov, S.G.; Dyatlov, I.A. Next-Generation Antibiotics, Bacteriophage Endolysins, and Nanomaterials for Combating Pathogens. Biochemistry 2020, 85, 1374–1388. [Google Scholar] [CrossRef]
- Lam, A.K.; Panlilio, H.; Pusavat, J.; Wouters, C.L.; Moen, E.L.; Rice, C.V. Overcoming Multidrug Resistance and Biofilms of Pseudomonas aeruginosa with a Single Dual-Function Potentiator of β-Lactams. ACS Infect. Dis. 2020, 6, 1085–1097. [Google Scholar] [CrossRef]
- Bhagirath, A.Y.; Li, Y.; Patidar, R.; Yerex, K.; Ma, X.; Kumar, A.; Duan, K. Two Component Regulatory Systems and Antibiotic Resistance in Gram-Negative Pathogens. Int. J. Mol. Sci. 2019, 20, 1781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pang, Z.; Raudonis, R.; Glick, B.R.; Lin, T.-J.; Cheng, Z. Antibiotic resistance in Pseudomonas aeruginosa: Mechanisms and alternative therapeutic strategies. Biotechnol. Adv. 2019, 37, 177–192. [Google Scholar] [CrossRef]
- Bukhari, S.I.; Aleanizy, F.S. Association of OprF mutant and disturbance of biofilm and pyocyanin virulence in pseudomonas aeruginosa. Saudi Pharm. J. 2020, 28, 196–200. [Google Scholar] [CrossRef]
- Mishra, B.; Wang, G. Individual and Combined Effects of Engineered Peptides and Antibiotics on Pseudomonas aeruginosa Biofilms. Pharmaceuticals 2017, 10, 58. [Google Scholar] [CrossRef] [Green Version]
- Gupta, V.; Datta, P. Next-generation strategy for treating drug resistant bacteria: Antibiotic hybrids. Indian J. Med. Res. 2019, 149, 97. [Google Scholar] [CrossRef]
- Sarno, R.; Ha, H.; Weinsetel, N.; Tolmasky, M.E. Inhibition of Aminoglycoside 6′-N-Acetyltransferase Type Ib-Mediated Amikacin Resistance by Antisense Oligodeoxynucleotides. Antimicrob. Agents Chemother. 2003, 47, 3296–3304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livermore, D.M. Multiple Mechanisms of Antimicrobial Resistance in Pseudomonas aeruginosa: Our Worst Nightmare? Clin. Infect. Dis. 2002, 34, 634–640. [Google Scholar] [CrossRef] [Green Version]
- Thomsen, T.; Mendel, H.; Al-Mansour, W.; Oddo, A.; Løbner-Olesen, A.; Hansen, P. Analogues of a Cyclic Antimicrobial Peptide with a Flexible Linker Show Promising Activity against Pseudomonas aeruginosa and Staphylococcus aureus. Antibiotics 2020, 9, 366. [Google Scholar] [CrossRef] [PubMed]
- Colque, C.A.; Albarracín Orio, A.G.; Feliziani, S.; Marvig, R.L.; Tobares, A.R.; Johansen, H.K.; Molin, S.; Smania, A.M. Hypermutator Pseudomonas aeruginosa Exploits Multiple Genetic Pathways to Develop Multidrug Resistance during Long-Term Infections in the Airways of Cystic Fibrosis Patients. Antimicrob. Agents Chemother. 2020, 64, e02142-19. [Google Scholar] [CrossRef] [PubMed]
- Slizen, M.V.; Galzitskaya, O.V. Comparative Analysis of Proteomes of a Number of Nosocomial Pathogens by KEGG Modules and KEGG Pathways. Int. J. Mol. Sci. 2020, 21, 7839. [Google Scholar] [CrossRef]
- Kurpe, S.; Grishin, S.; Surin, A.; Selivanova, O.; Fadeev, R.; Dzhus, U.; Gorbunova, E.; Mustaeva, L.; Azev, V.; Galzitskaya, O. Antimicrobial and Amyloidogenic Activity of Peptides Synthesized on the Basis of the Ribosomal S1 Protein from Thermus Thermophilus. Int. J. Mol. Sci. 2020, 21, 6382. [Google Scholar] [CrossRef]
- Wang, Y.; Kadiyala, U.; Qu, Z.; Elvati, P.; Altheim, C.; Kotov, N.A.; Violi, A.; VanEpps, J.S. Anti-Biofilm Activity of Graphene Quantum Dots via Self-Assembly with Bacterial Amyloid Proteins. ACS Nano 2019, 13, 4278–4289. [Google Scholar] [CrossRef] [PubMed]
- Kagan, B.L.; Jang, H.; Capone, R.; Teran Arce, F.; Ramachandran, S.; Lal, R.; Nussinov, R. Antimicrobial Properties of Amyloid Peptides. Mol. Pharm. 2012, 9, 708–717. [Google Scholar] [CrossRef] [Green Version]
- Lee, E.Y.; Srinivasan, Y.; de Anda, J.; Nicastro, L.K.; Tükel, Ç.; Wong, G.C.L. Functional Reciprocity of Amyloids and Antimicrobial Peptides: Rethinking the Role of Supramolecular Assembly in Host Defense, Immune Activation, and Inflammation. Front. Immunol. 2020, 11, 1629. [Google Scholar] [CrossRef]
- Wang, C.K.; King, G.J.; Conibear, A.C.; Ramos, M.C.; Chaousis, S.; Henriques, S.T.; Craik, D.J. Mirror Images of Antimicrobial Peptides Provide Reflections on Their Functions and Amyloidogenic Properties. J. Am. Chem. Soc. 2016, 138, 5706–5713. [Google Scholar] [CrossRef] [Green Version]
- Vane, E.W.; He, S.; Maibaum, L.; Nath, A. Rapid Formation of Peptide/Lipid Coaggregates by the Amyloidogenic Seminal Peptide PAP248-286. Biophys. J. 2020, 119, 924–938. [Google Scholar] [CrossRef]
- Kurpe, S.R.; Grishin, S.Y.; Surin, A.K.; Panfilov, A.V.; Slizen, M.V.; Chowdhury, S.D.; Galzitskaya, O.V. Antimicrobial and Amyloidogenic Activity of Peptides. Can Antimicrobial Peptides Be Used against SARS-CoV-2? Int. J. Mol. Sci. 2020, 21, 9552. [Google Scholar] [CrossRef] [PubMed]
- Schramm, F.D.; Schroeder, K.; Jonas, K. Protein aggregation in bacteria. FEMS Microbiol. Rev. 2020, 44, 54–72. [Google Scholar] [CrossRef] [PubMed]
- Giraldo, R. Optogenetic Navigation of Routes Leading to Protein Amyloidogenesis in Bacteria. J. Mol. Biol. 2019, 431, 1186–1202. [Google Scholar] [CrossRef] [Green Version]
- Khodaparast, L.; Wu, G.; Khodaparast, L.; Schmidt, B.Z.; Rousseau, F.; Schymkowitz, J. Bacterial Protein Homeostasis Disruption as a Therapeutic Intervention. Front. Mol. Biosci. 2021, 8, 434. [Google Scholar] [CrossRef]
- Bednarska, N.G.; van Eldere, J.; Gallardo, R.; Ganesan, A.; Ramakers, M.; Vogel, I.; Baatsen, P.; Staes, A.; Goethals, M.; Hammarström, P.; et al. Protein aggregation as an antibiotic design strategy. Mol. Microbiol. 2016, 99, 849–865. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.; Khodaparast, L.; Khodaparast, L.; De Vleeschouwer, M.; Housmans, J.; Houben, B.; Schymkowitz, J.; Rousseau, F. Investigating the mechanism of action of aggregation-inducing antimicrobial Pept-ins. Cell Chem. Biol. 2021, 28, 524–536.e4. [Google Scholar] [CrossRef] [PubMed]
- Matilla-Cuenca, L.; Toledo-Arana, A.; Valle, J. Anti-Biofilm Molecules Targeting Functional Amyloids. Antibiotics 2021, 10, 795. [Google Scholar] [CrossRef]
- Jain, N.; Ådén, J.; Nagamatsu, K.; Evans, M.L.; Li, X.; McMichael, B.; Ivanova, M.I.; Almqvist, F.; Buxbaum, J.N.; Chapman, M.R. Inhibition of curli assembly and Escherichia coli biofilm formation by the human systemic amyloid precursor transthyretin. Proc. Natl. Acad. Sci. USA 2017, 114, 12184–12189. [Google Scholar] [CrossRef] [Green Version]
- Sengupta, J.; Agrawal, R.K.; Frank, J. Visualization of protein S1 within the 30S ribosomal subunit and its interaction with messenger RNA. Proc. Natl. Acad. Sci. USA 2001, 98, 11991–11996. [Google Scholar] [CrossRef] [Green Version]
- Machulin, A.V.; Deryusheva, E.I.; Selivanova, O.M.; Galzitskaya, O.V. The number of domains in the ribosomal protein S1 as a hallmark of the phylogenetic grouping of bacteria. PLoS ONE 2019, 14, e0221370. [Google Scholar] [CrossRef] [Green Version]
- Machulin, A.; Deryusheva, E.; Lobanov, M.; Galzitskaya, O. Repeats in S1 Proteins: Flexibility and Tendency for Intrinsic Disorder. Int. J. Mol. Sci. 2019, 20, 2377. [Google Scholar] [CrossRef] [Green Version]
- Deryusheva, E.I.; Machulin, A.V.; Selivanova, O.M.; Galzitskaya, O.V. Taxonomic distribution, repeats, and functions of the S1 domain-containing proteins as members of the OB-fold family. Proteins Struct. Funct. Bioinforma 2017, 85, 602–613. [Google Scholar] [CrossRef]
- Heinemann, U.; Roske, Y. Cold-Shock Domains—Abundance, Structure, Properties, and Nucleic-Acid Binding. Cancers 2021, 13, 190. [Google Scholar] [CrossRef] [PubMed]
- Deryusheva, E.I.; Machulin, A.V.; Matyunin, M.A.; Galzitskaya, O.V. Investigation of the Relationship between the S1 Domain and Its Molecular Functions Derived from Studies of the Tertiary Structure. Molecules 2019, 24, 3681. [Google Scholar] [CrossRef] [Green Version]
- Deryusheva, E.; Machulin, A.; Matyunin, M.; Galzitskaya, O. Sequence and evolutionary analysis of bacterial ribosomal S1 proteins. Proteins Struct. Funct. Bioinforma. 2021, 84, 1111–1124. [Google Scholar] [CrossRef] [PubMed]
- Garbuzynskiy, S.O.; Lobanov, M.Y.; Galzitskaya, O.V. FoldAmyloid: A method of prediction of amyloidogenic regions from protein sequence. Bioinformatics 2010, 26, 326–332. [Google Scholar] [CrossRef]
- Oliveberg, M. Waltz, an exciting new move in amyloid prediction. Nat. Methods 2010, 7, 187–188. [Google Scholar] [CrossRef]
- Conchillo-Solé, O.; de Groot, N.S.; Avilés, F.X.; Vendrell, J.; Daura, X.; Ventura, S. AGGRESCAN: A server for the prediction and evaluation of “hot spots” of aggregation in polypeptides. BMC Bioinform. 2007, 8, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walsh, I.; Seno, F.; Tosatto, S.C.E.; Trovato, A. PASTA 2.0: An improved server for protein aggregation prediction. Nucleic Acids Res. 2014, 42, W301–W307. [Google Scholar] [CrossRef]
- Grishin, S.Y.; Dzhus, U.F.; Glukhov, A.S.; Selivanova, O.M.; Surin, A.K.; Galzitskaya, O.V. Identification of Amyloidogenic Regions in Pseudomonas aeruginosa Ribosomal S1 Protein. Int. J. Mol. Sci. 2021, 22, 7291. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, Y.; Tang, Y.; Zhang, D.; He, H.; Wu, J.; Zheng, J. Antimicrobial α-defensins as multi-target inhibitors against amyloid formation and microbial infection. Chem. Sci. 2021, 12, 9124–9139. [Google Scholar] [CrossRef] [PubMed]
- Stepanenko, O.V.; Sulatsky, M.I.; Mikhailova, E.V.; Stepanenko, O.V.; Kuznetsova, I.M.; Turoverov, K.K.; Sulatskaya, A.I. Trypsin Induced Degradation of Amyloid Fibrils. Int. J. Mol. Sci. 2021, 22, 4828. [Google Scholar] [CrossRef] [PubMed]
- Surin, A.K.; Grishin, S.Y.; Galzitskaya, O.V. Determination of amyloid core regions of insulin analogues fibrils. Prion 2020, 14, 149–162. [Google Scholar] [CrossRef]
- Sarkar, T.; Chetia, M.; Chatterjee, S. Antimicrobial Peptides and Proteins: From Nature’s Reservoir to the Laboratory and Beyond. Front. Chem. 2021, 9, 432. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Zou, C.; Wen, N.; Liu, X.; Meng, Z.; Feng, S.; Zheng, Z.; Meng, Q.; Wang, C. The effect of structural modification of antimicrobial peptides on their antimicrobial activity, hemolytic activity, and plasma stability. J. Pept. Sci. 2021, 27, e3306. [Google Scholar] [CrossRef]
- Chen, X.; Petranovic, D. Amyloid-β peptide-induced cytotoxicity and mitochondrial dysfunction in yeast. FEMS Yeast Res. 2015, 15, fov061. [Google Scholar] [CrossRef] [Green Version]
- Fakhranurova, L.; Balobanov, V.; Ryabova, N.; Glukhov, A.; Ilyina, N.; Markelova, N.; Marchenkov, V.; Katina, N. The presence of cross-β-structure as a key determinant of carbonic anhydrase amyloid fibrils cytotoxicity. Biochem. Biophys. Res. Commun. 2020, 524, 453–458. [Google Scholar] [CrossRef]
- Zhang, Y.; Tang, Y.; Liu, Y.; Zhang, D.; Zheng, J. Design and Engineering of Amyloid Aggregation-Prone Fragments and Their Antimicrobial Conjugates with Multi-Target Functionality. Adv. Funct. Mater. 2021, 31, 2102978. [Google Scholar] [CrossRef]
- Buccini, D.F.; Cardoso, M.H.; Franco, O.L. Antimicrobial Peptides and Cell-Penetrating Peptides for Treating Intracellular Bacterial Infections. Front. Cell. Infect. Microbiol. 2021, 10, 612931. [Google Scholar] [CrossRef]
- Beyermann, M.; Henklein, P.; Klose, A.; Sohr, R.; Bienert, M. Effect of tertiary amine on the carbodiimide-mediated peptide synthesis. Int. J. Pept. Protein Res. 2009, 37, 252–256. [Google Scholar] [CrossRef]
- Wu, J.; Gao, B.; Zhu, S. Single-point mutation-mediated local amphipathic adjustment dramatically enhances antibacterial activity of a fungal defensin. FASEB J. 2016, 30, 2602–2614. [Google Scholar] [CrossRef] [Green Version]
- Gaglione, R.; Smaldone, G.; Cesaro, A.; Rumolo, M.; De Luca, M.; Di Girolamo, R.; Petraccone, L.; Del Vecchio, P.; Oliva, R.; Notomista, E.; et al. Impact of a Single Point Mutation on the Antimicrobial and Fibrillogenic Properties of Cryptides from Human Apolipoprotein B. Pharmaceuticals 2021, 14, 631. [Google Scholar] [CrossRef] [PubMed]
- Yokoo, H.; Hirano, M.; Ohoka, N.; Misawa, T.; Demizu, Y. Structure–activity relationship study of amphipathic antimicrobial peptides using helix-destabilizing sarcosine. J. Pept. Sci. 2021, e3360. [Google Scholar] [CrossRef]
- Drozdetskiy, A.; Cole, C.; Procter, J.; Barton, G.J. JPred4: A protein secondary structure prediction server. Nucleic Acids Res. 2015, 43, W389–W394. [Google Scholar] [CrossRef]
- Rost, B.; Yachdav, G.; Liu, J. The PredictProtein server. Nucleic Acids Res. 2004, 32, W321–W326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pirtskhalava, M.; Amstrong, A.A.; Grigolava, M.; Chubinidze, M.; Alimbarashvili, E.; Vishnepolsky, B.; Gabrielian, A.; Rosenthal, A.; Hurt, D.E.; Tartakovsky, M. DBAASP v3: Database of antimicrobial/cytotoxic activity and structure of peptides as a resource for development of new therapeutics. Nucleic Acids Res. 2021, 49, D288–D297. [Google Scholar] [CrossRef] [PubMed]
- Pirtskhalava, M.; Vishnepolsky, B.; Grigolava, M.; Managadze, G. Physicochemical Features and Peculiarities of Interaction of AMP with the Membrane. Pharmaceuticals 2021, 14, 471. [Google Scholar] [CrossRef]
- Pandit, G.; Chowdhury, N.; Abdul Mohid, S.; Bidkar, A.P.; Bhunia, A.; Chatterjee, S. Effect of Secondary Structure and Side Chain Length of Hydrophobic Amino Acid Residues on the Antimicrobial Activity and Toxicity of 14-Residue-Long de novo AMPs. ChemMedChem 2021, 16, 355–367. [Google Scholar] [CrossRef]
- Wang, K.F.; Nagarajan, R.; Camesano, T.A. Differentiating antimicrobial peptides interacting with lipid bilayer: Molecular signatures derived from quartz crystal microbalance with dissipation monitoring. Biophys. Chem. 2015, 196, 53–67. [Google Scholar] [CrossRef]
- Liu, Y.; Xia, X.; Xu, L.; Wang, Y. Design of hybrid β-hairpin peptides with enhanced cell specificity and potent anti-inflammatory activity. Biomaterials 2013, 34, 237–250. [Google Scholar] [CrossRef]
- Frederiksen, N.; Hansen, P.R.; Zabicka, D.; Tomczak, M.; Urbas, M.; Domraceva, I.; Björkling, F.; Franzyk, H. Alternating Cationic-Hydrophobic Peptide/Peptoid Hybrids: Influence of Hydrophobicity on Antibacterial Activity and Cell Selectivity. ChemMedChem 2020, 15, 2544–2561. [Google Scholar] [CrossRef]
- Gong, H.; Liao, M.; Hu, X.; Fa, K.; Phanphak, S.; Ciumac, D.; Hollowell, P.; Shen, K.; Clifton, L.A.; Campana, M.; et al. Aggregated Amphiphilic Antimicrobial Peptides Embedded in Bacterial Membranes. ACS Appl. Mater. Interfaces 2020, 12, 44420–44432. [Google Scholar] [CrossRef]
- Dathe, M.; Meyer, J.; Beyermann, M.; Maul, B.; Hoischen, C.; Bienert, M. General aspects of peptide selectivity towards lipid bilayers and cell membranes studied by variation of the structural parameters of amphipathic helical model peptides. Biochim. Biophys. Acta Biomembr. 2002, 1558, 171–186. [Google Scholar] [CrossRef] [Green Version]
- Meikle, T.G.; Dharmadana, D.; Hoffmann, S.V.; Jones, N.C.; Drummond, C.J.; Conn, C.E. Analysis of the structure, loading and activity of six antimicrobial peptides encapsulated in cubic phase lipid nanoparticles. J. Colloid Interface Sci. 2021, 587, 90–100. [Google Scholar] [CrossRef] [PubMed]
- Souza, P.F.N.; Marques, L.S.M.; Oliveira, J.T.A.; Lima, P.G.; Dias, L.P.; Neto, N.A.S.; Lopes, F.E.S.; Sousa, J.S.; Silva, A.F.B.; Caneiro, R.F.; et al. Synthetic antimicrobial peptides: From choice of the best sequences to action mechanisms. Biochimie 2020, 175, 132–145. [Google Scholar] [CrossRef] [PubMed]
- Singh, O.; Hsu, W.-L.; Su, E.C.-Y. Co-AMPpred for in silico-aided predictions of antimicrobial peptides by integrating composition-based features. BMC Bioinform. 2021, 22, 389. [Google Scholar] [CrossRef]
- Sheehan, D.J.; Janda, J.M.; Bottone, E.J. Pseudomonas aeruginosa: Changes in antibiotic susceptibility, enzymatic activity, and antigenicity among colonial morphotypes. J. Clin. Microbiol. 1982, 15, 926–930. [Google Scholar] [CrossRef] [Green Version]
- Domnin, P.; Arkhipova, A.; Petrov, S.; Sysolyatina, E.; Parfenov, V.; Karalkin, P.; Mukhachev, A.; Gusarov, A.; Moisenovich, M.; Khesuani, Y.; et al. An In Vitro Model of Nonattached Biofilm-Like Bacterial Aggregates Based on Magnetic Levitation. Appl. Environ. Microbiol. 2020, 86, e01074-20. [Google Scholar] [CrossRef]
- Soundrarajan, N.; Park, S.; Le Van Chanh, Q.; Cho, H.; Raghunathan, G.; Ahn, B.; Song, H.; Kim, J.-H.; Park, C. Protegrin-1 cytotoxicity towards mammalian cells positively correlates with the magnitude of conformational changes of the unfolded form upon cell interaction. Sci. Rep. 2019, 9, 11569. [Google Scholar] [CrossRef] [Green Version]
- Grishin, S.Y.; Deryusheva, E.I.; Machulin, A.V.; Selivanova, O.M.; Glyakina, A.V.; Gorbunova, E.Y.; Mustaeva, L.G.; Azev, V.N.; Rekstina, V.V.; Kalebina, T.S.; et al. Amyloidogenic Propensities of Ribosomal S1 Proteins: Bioinformatics Screening and Experimental Checking. Int. J. Mol. Sci. 2020, 21, 5199. [Google Scholar] [CrossRef]
- Vasilchenko, A.S.; Vasilchenko, A.V.; Valyshev, A.V.; Rogozhin, E.A. A Novel High-Molecular-Mass Bacteriocin Produced by Enterococcus faecium: Biochemical Features and Mode of Action. Probiotics Antimicrob. Proteins 2018, 10, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Suvorina, M.Y.; Selivanova, O.M.; Grigorashvili, E.I.; Nikulin, A.D.; Marchenkov, V.V.; Surin, A.K.; Galzitskaya, O.V. Studies of Polymorphism of Amyloid-β 42 Peptide from Different Suppliers. J. Alzheimer’s Dis. 2015, 47, 583–593. [Google Scholar] [CrossRef] [PubMed]
- Stefani, M. Biochemical and biophysical features of both oligomer/fibril and cell membrane in amyloid cytotoxicity. FEBS J. 2010, 277, 4602–4613. [Google Scholar] [CrossRef] [Green Version]
- Mwangi, J.; Yin, Y.; Wang, G.; Yang, M.; Li, Y.; Zhang, Z.; Lai, R. The antimicrobial peptide ZY4 combats multidrug-resistant Pseudomonas aeruginosa and Acinetobacter baumannii infection. Proc. Natl. Acad. Sci. USA 2019, 116, 26516–26522. [Google Scholar] [CrossRef] [Green Version]
- Yasir, M.; Dutta, D.; Willcox, M.D.P. Comparative mode of action of the antimicrobial peptide melimine and its derivative Mel4 against Pseudomonas aeruginosa. Sci. Rep. 2019, 9, 7063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yasir, M.; Dutta, D.; Hossain, K.R.; Chen, R.; Ho, K.K.K.; Kuppusamy, R.; Clarke, R.J.; Kumar, N.; Willcox, M.D.P. Mechanism of Action of Surface Immobilized Antimicrobial Peptides Against Pseudomonas aeruginosa. Front. Microbiol. 2020, 10, 3053. [Google Scholar] [CrossRef] [PubMed]
- Bernhofer, M.; Dallago, C.; Karl, T.; Satagopam, V.; Heinzinger, M.; Littmann, M.; Olenyi, T.; Qiu, J.; Schütze, K.; Yachdav, G.; et al. PredictProtein—Predicting Protein Structure and Function for 29 Years. bioRxiv 2021. [Google Scholar] [CrossRef]
- Moon, C.P.; Fleming, K.G. Side-chain hydrophobicity scale derived from transmembrane protein folding into lipid bilayers. Proc. Natl. Acad. Sci. USA 2011, 108, 10174–10177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waghu, F.H.; Gopi, L.; Barai, R.S.; Ramteke, P.; Nizami, B.; Idicula-Thomas, S. CAMP: Collection of sequences and structures of antimicrobial peptides. Nucleic Acids Res. 2014, 42, D1154–D1158. [Google Scholar] [CrossRef] [Green Version]
- Kurpe, S.R.; Grishin, S.Y.; Glyakina, A.V.; Slizen, M.V.; Panfilov, A.V.; Kochetov, A.P.; Surin, A.K.; Kobyakova, M.I.; Fadeev, R.S.; Galzitskaya, O.V. Antibacterial effects of peptides synthesized based on the sequence of ribosome protein S1. Biomed. Khim. 2021, 67, 231–243. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide | JPred4 (Jnetpred) | Predict Protein (RePROF) | DBAASP v3.0 (MF Scale) | |||
|---|---|---|---|---|---|---|
| µHn | Hn | Z | I0 | |||
| R23R vs. R23R* | ||||||
| RKKRRQRRRGGGGLHITDMAWKR | β-strand 14–18 a.a. (LHITD) | Helix 2–9 a.a. (KKRRQRRR), β-strand 14–18 a.a. (LHITD) | 0.36 | 0.91 | +9 | 12.51 |
| RKKRRQRRRGG-Sar §§(A)-GLHITD-Nle §§§(M)-AWKR | Helix 2–9 a.a. (KKRRQRRR), other 10–14 a.a. (GGAGL), helix 18–22 (DMAWK) | 0.32 | 0.84 | +9 | 12.51 | |
| RKKRRQRRRGG-Sar(A)-GLHITD-Nle(L)-AWKR | β-strand 14–18 a.a. (LHITD) | Helix 2–9 a.a. (KKRRQRRR), other 10–14 a.a. (GGAGL), helix 18–22 (DMAWK) | 0.29 | 0.79 | +9 | 12.51 |
| RKKRRQRRRGG-Sar(P)-GLHITD-Nle(M)-AWKR | Helix 2–9 a.a. (KKRRQRRR), other 10–14 a.a. (GGAGL) | 0.29 | 0.77 | +9 | 12.51 | |
| RKKRRQRRRGG-Sar(P)-GLHITD-Nle(L)-AWKR | Helix 2–9 a.a. (KKRRQRRR), other 10–14 a.a. (GGPGL) | 0.27 | 0.73 | +9 | 12.51 | |
| R23L vs. R23L* | ||||||
| RKKRRQRRRGGGGITDFGIFIGL | β-strand 17–21 a.a. (FGIFI) | Helix 3–9 a.a. (KRRQRRR), other 10–15 a.a. (GGGGIT), β-strand 16–21 a.a. (DFGIFI) | 0.24 | 0.18 | +7 | 12.41 |
| RKKRRQRRRGG-Sar(A)-GITDFGIFIGL | β-strand 17–21 a.a. (FGIFI) | Helix 3–9 a.a. (KKRRQRRR), other 10–15 a.a. (GGAGIT) | 0.18 | 0.11 | +7 | 12.41 |
| RKKRRQRRRGG-Sar(P)-GITDFGIFIGL | β-strand 17–21 a.a. (FGIFI) | Helix 3–9 a.a. (KKRRQRRR), other 10–15 a.a. (GGPGIT) | 0.14 | 0.04 | +7 | 12.41 |
| Peptide | CAMPR3 | AmpGram | AMP Scanner | |||
|---|---|---|---|---|---|---|
| RF | SVM | ANN | DA | RF and n-Grams | DNN | |
| R23R vs. R23R* | ||||||
| RKKRRQRRRGGGGLHITDMAWKR | 0.48 (non-AMP §) | 0.03 (non-AMP) | AMP §§ | 0.93 (AMP) | 0.59 (AMP) | 0.07 (non-AMP) |
| RKKRRQRRRGG-Sar §§§(A)-GLHITD-Nle §§§§(M)-AWKR | 0.49 (non-AMP) | 0.03 (non-AMP) | AMP | 0.95 (AMP) | 0.54 (AMP) | 0.05 (non-AMP) |
| RKKRRQRRRGG-Sar(A)-GLHITD-Nle(L)-AWKR | 0.58 (AMP) | 0.05 (non-AMP) | AMP | 0.96 (AMP) | 0.63 (AMP) | 0.03 (non-AMP) |
| RKKRRQRRRGG-Sar(P)-GLHITD-Nle(M)-AWKR | 0.48 (non-AMP) | 0.04 (non-AMP) | non-AMP | 0.95 (AMP) | 0.47 (non-AMP) | 0.13 (non-AMP) |
| RKKRRQRRRGG-Sar(P)-GLHITD-Nle(L)-AWKR | 0.53 (AMP) | 0.07 (non-AMP) | AMP | 0.97 (AMP) | 0.62 (AMP) | 0.07 (non-AMP) |
| R23L vs. R23L* | ||||||
| RKKRRQRRRGGGGITDFGIFIGL | 0.59 (AMP) | 0.06 (non-AMP) | AMP | 1.00 (AMP) | 0.37 (non-AMP) | 0.93 (AMP) |
| RKKRRQRRRGG-Sar(A)-GITDFGIFIGL | 0.59 (AMP) | 0.06 (non-AMP) | AMP | 1.00 (AMP) | 0.22 (non-AMP) | 0.44 (non-AMP) |
| RKKRRQRRRGG-Sar(P)-GITDFGIFIGL | 0.58 (AMP) | 0.10 (non-AMP) | AMP | 1.00 (AMP) | 0.45 (non-AMP) | 0.95 (AMP) |
| Photo of Results | Scheme |
|---|---|
 | Test of peptide R23R 1—R23R, 300 µM 2—R23R, 250 µM 3—R23R, 200 µM 4—R23R, 150 µM 5—DMSO, 2% (volume/volume) 6—Gentamicin sulfate, 1700 µM |
 | Test of peptide R23L 1—R23L, 300 µM 2—R23L, 150 µM 3—R23L, 75 µM 4—R23L, 37.5 µM 5—DMSO, 2% (volume/volume) 6—Gentamicin sulfate, 1700 µM |
 | Test of peptide R23R* 1—R23R*, 300 µM 2—R23R*, 150 µM 3—R23R*, 75 µM 4—R23R*, 37.5 µM 5—R23R*, 18.8 µM 6—DMSO, 2% (volume/volume) 7—Gentamicin sulfate, 1700 µM |
 | Test of peptide R23L* 1—R23L*, 300 µM 2—R23L*, 150 µM 3—R23L*, 75 µM 4—R23L*, 37.5 µM 5—R23L*, 18.8 µM 6—DMSO, 2% (volume/volume) 7—Gentamicin sulfate, 1700 µM |
| Photo of Results | Scheme |
|---|---|
 | Test of peptide R23R 1—R23R, 3750 µM 2—R23R, 375 µM 3—R23R, 37.5 µM 4—R23R, 3.75 µM 5—DMSO, 20% (volume/volume) 6—LB medium |
 | Test of peptide R23L 1—R23L, 3750 µM 2—R23L, 375 µM 3—R23L, 37.5 µM 4—R23L, 3.75 µM 5—DMSO, 20% (volume/volume) 6—Luria-Bertani (LB) medium |
 | 1—LB medium 2—DMSO, 20% (volume/volume) 3—Gentamicin sulfate, 17 µM |
 | Test of peptide R23R* 1—R23R*, 3750 µM 2—R23R*, 375 µM 3—R23R*, 37.5 µM 4—R23R*, 3.75 µM 5—DMSO, 20% (volume/volume) 6—LB medium 7—Gentamicin sulfate, 17 µM |
 | Test of peptide R23L* 1—R23L*, 3750 µM 2—R23L*, 375 µM 3—R23L*, 37.5 µM 4—R23L*, 3.75 µM 5—DMSO, 20% (volume/volume) 6—LB medium 7—Gentamicin sulfate, 17 µM |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grishin, S.Y.; Domnin, P.A.; Kravchenko, S.V.; Azev, V.N.; Mustaeva, L.G.; Gorbunova, E.Y.; Kobyakova, M.I.; Surin, A.K.; Makarova, M.A.; Kurpe, S.R.; et al. Is It Possible to Create Antimicrobial Peptides Based on the Amyloidogenic Sequence of Ribosomal S1 Protein of P. aeruginosa? Int. J. Mol. Sci. 2021, 22, 9776. https://doi.org/10.3390/ijms22189776
Grishin SY, Domnin PA, Kravchenko SV, Azev VN, Mustaeva LG, Gorbunova EY, Kobyakova MI, Surin AK, Makarova MA, Kurpe SR, et al. Is It Possible to Create Antimicrobial Peptides Based on the Amyloidogenic Sequence of Ribosomal S1 Protein of P. aeruginosa? International Journal of Molecular Sciences. 2021; 22(18):9776. https://doi.org/10.3390/ijms22189776
Chicago/Turabian StyleGrishin, Sergei Y., Pavel A. Domnin, Sergey V. Kravchenko, Viacheslav N. Azev, Leila G. Mustaeva, Elena Y. Gorbunova, Margarita I. Kobyakova, Alexey K. Surin, Maria A. Makarova, Stanislav R. Kurpe, and et al. 2021. "Is It Possible to Create Antimicrobial Peptides Based on the Amyloidogenic Sequence of Ribosomal S1 Protein of P. aeruginosa?" International Journal of Molecular Sciences 22, no. 18: 9776. https://doi.org/10.3390/ijms22189776
APA StyleGrishin, S. Y., Domnin, P. A., Kravchenko, S. V., Azev, V. N., Mustaeva, L. G., Gorbunova, E. Y., Kobyakova, M. I., Surin, A. K., Makarova, M. A., Kurpe, S. R., Fadeev, R. S., Vasilchenko, A. S., Firstova, V. V., Ermolaeva, S. A., & Galzitskaya, O. V. (2021). Is It Possible to Create Antimicrobial Peptides Based on the Amyloidogenic Sequence of Ribosomal S1 Protein of P. aeruginosa? International Journal of Molecular Sciences, 22(18), 9776. https://doi.org/10.3390/ijms22189776