
Procoagulant Extracellular Vesicles Alter Trophoblast Differentiation in Mice by a Thrombo-Inflammatory Mechanism

 , , and
, , and 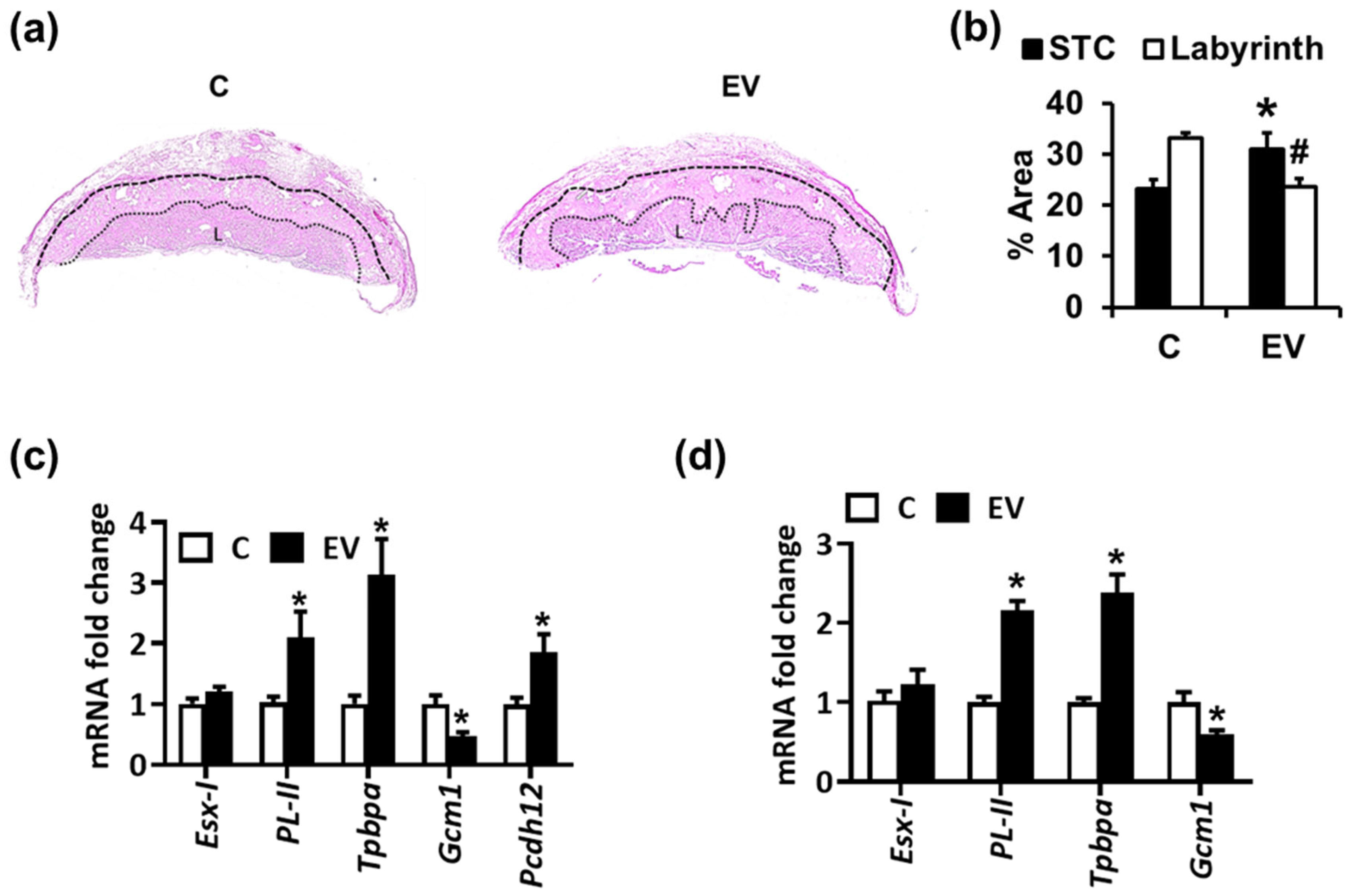{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
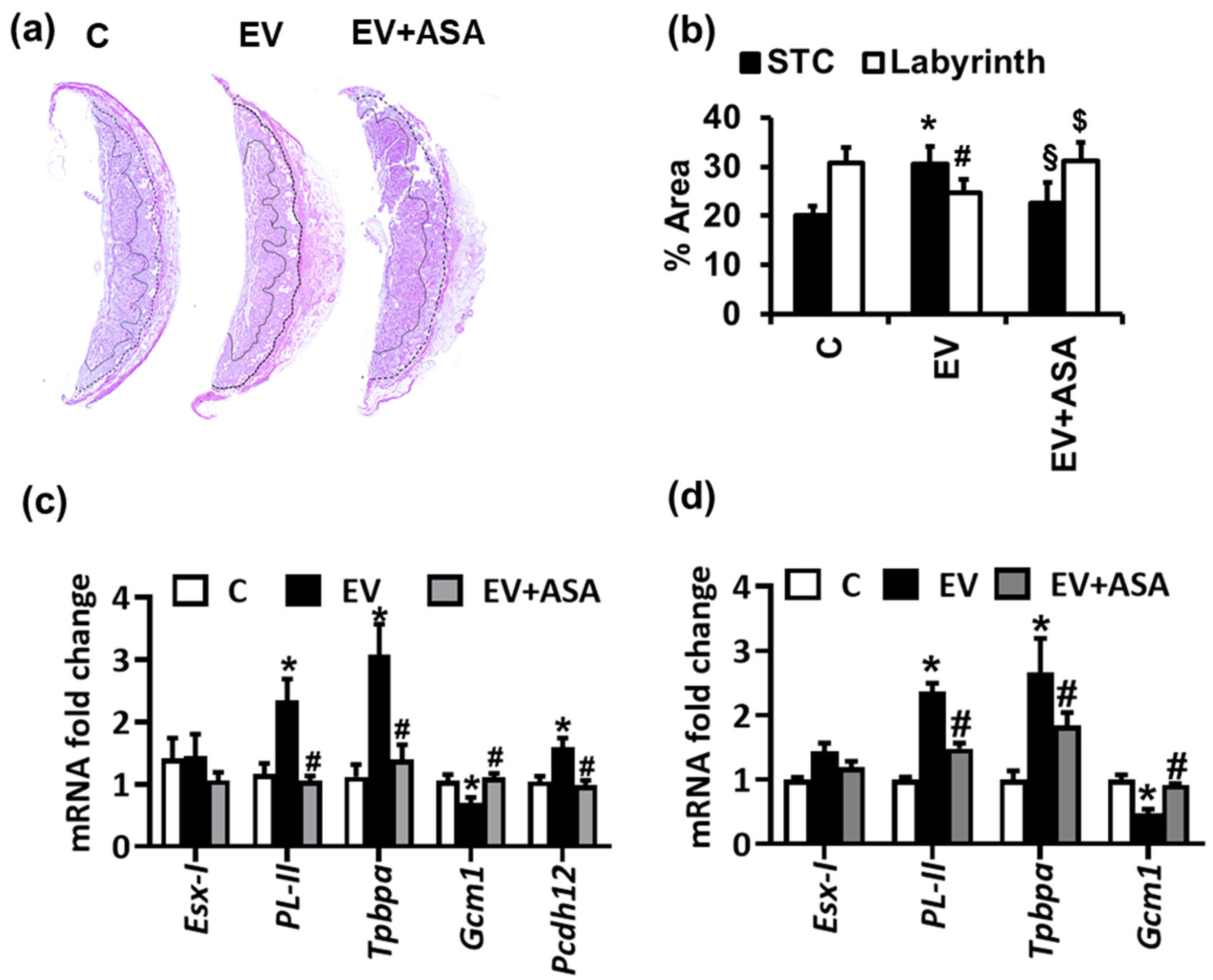
2.1. Extracellular Vesicles and Platelets Alter Trophoblast Morphology, Differentiation, and Glycogen-Rich Cells
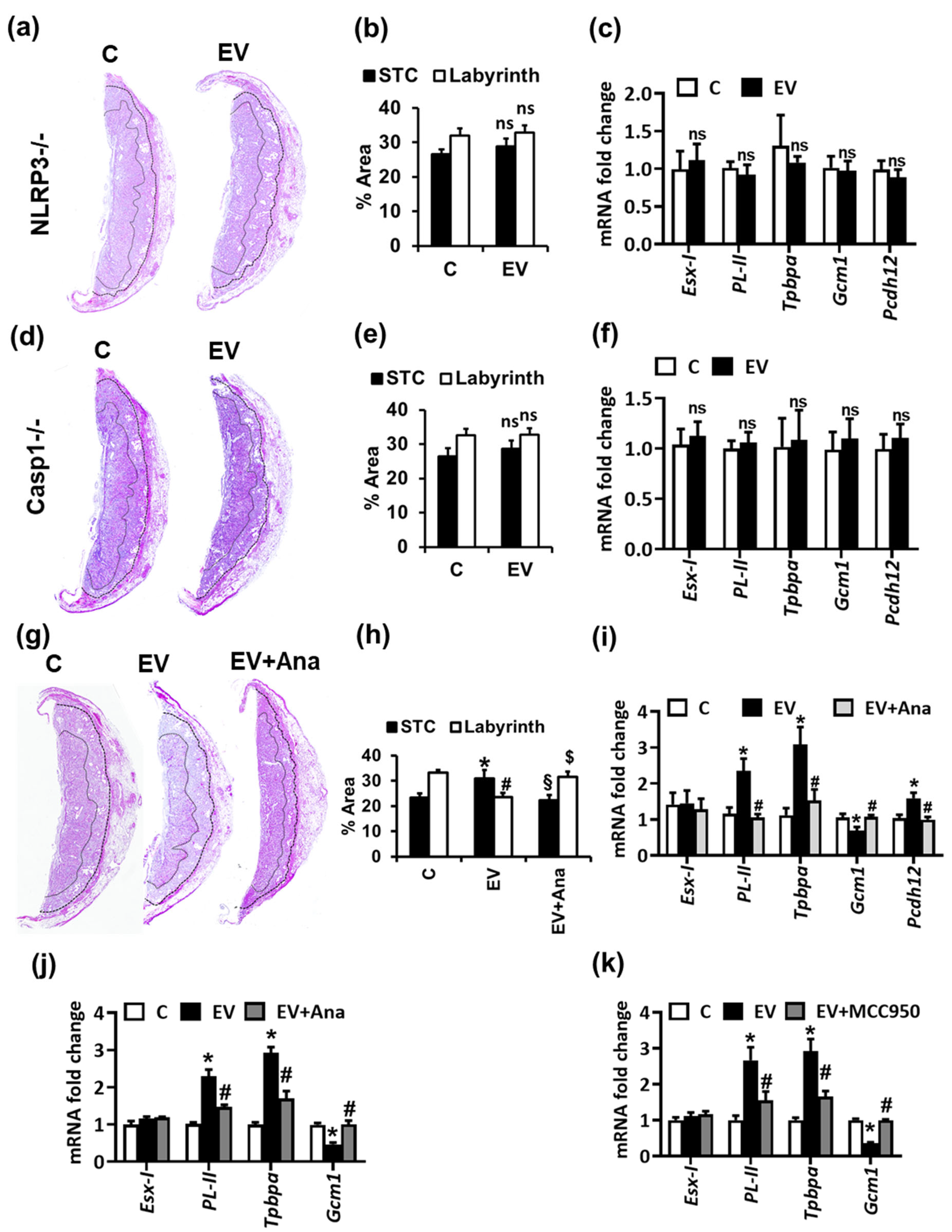
2.2. NLRP3-Mediated Thrombo-Inflammation Causes EV-Induced Altered Trophoblast Differentiation and Glycogen-Rich Cells
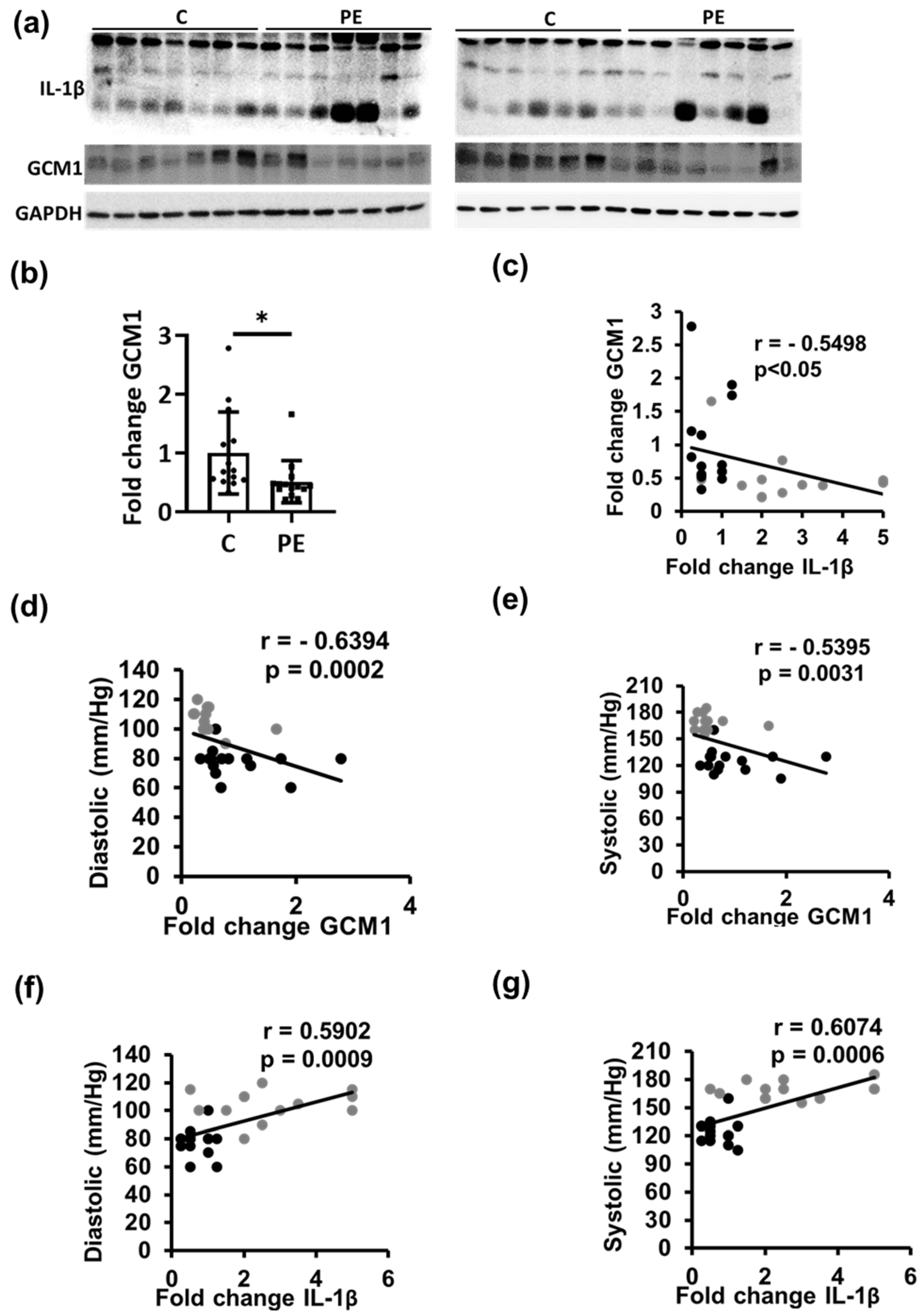
2.3. Trophoblast Differentiation Is Correlated with Placental Thrombo-Inflammation in PE Patients
3. Methods
3.1. Mice
3.2. Generation of Procoagulant EVs
3.3. Cell Culture
3.4. Human Samples
3.5. Immunoblotting
3.6. RNA Isolation and qRT-PCR
3.7. Statistical Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Hemberger, M.; Hanna, C.W.; Dean, W. Mechanisms of early placental development in mouse and humans. Nat. Rev. Genet. 2020, 21, 27–43. [Google Scholar] [CrossRef]
- Woods, L.; Perez-Garcia, V.; Hemberger, M. Regulation of Placental Development and Its Impact on Fetal Growth-New Insights From Mouse Models. Front. Endocrinol. 2018, 9, 570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cross, J.C. How to make a placenta: Mechanisms of trophoblast cell differentiation in mice—A review. Placenta 2005, 26, S3–S9. [Google Scholar] [CrossRef]
- Zhao, C.; Gu, Y.; Zeng, X.; Wang, J. NLRP3 inflammasome regulates Th17 differentiation in rheumatoid arthritis. Clin. Immunol. 2018, 197, 154–160. [Google Scholar] [CrossRef] [PubMed]
- Tyrkalska, S.D.; Perez-Oliva, A.B.; Rodriguez-Ruiz, L.; Martinez-Morcillo, F.J.; Alcaraz-Perez, F.; Martinez-Navarro, F.J.; Lachaud, C.; Ahmed, N.; Schroeder, T.; Pardo-Sanchez, I.; et al. Inflammasome Regulates Hematopoiesis through Cleavage of the Master Erythroid Transcription Factor GATA1. Immunity 2019, 51, 50–63.e55. [Google Scholar] [CrossRef]
- Schror, K. Aspirin and platelets: The antiplatelet action of aspirin and its role in thrombosis treatment and prophylaxis. Semin. Thromb. Hemost. 1997, 23, 349–356. [Google Scholar] [CrossRef]
- Rolnik, D.L.; Wright, D.; Poon, L.C.; O’Gorman, N.; Syngelaki, A.; de Paco Matallana, C.; Akolekar, R.; Cicero, S.; Janga, D.; Singh, M.; et al. Aspirin versus Placebo in Pregnancies at High Risk for Preterm Preeclampsia. N. Engl. J. Med. 2017, 377, 613–622. [Google Scholar] [CrossRef] [PubMed]
- Kohli, S.; Ranjan, S.; Hoffmann, J.; Kashif, M.; Daniel, E.A.; Al-Dabet, M.M.; Bock, F.; Nazir, S.; Huebner, H.; Mertens, P.R.; et al. Maternal extracellular vesicles and platelets promote preeclampsia through inflammasome activation in embryonic trophoblast. Blood 2016, 128, 2153–2164. [Google Scholar] [CrossRef]
- Morel, O.; Toti, F.; Hugel, B.; Bakouboula, B.; Camoin-Jau, L.; Dignat-George, F.; Freyssinet, J.M. Procoagulant microparticles: Disrupting the vascular homeostasis equation? Arterioscler. Thromb. Vasc. Biol. 2006, 26, 2594–2604. [Google Scholar] [CrossRef]
- Nielsen, T.; Kristensen, A.F.; Pedersen, S.; Christiansen, G.; Kristensen, S.R. Investigation of procoagulant activity in extracellular vesicles isolated by differential ultracentrifugation. J. Extracell. Vesicles 2018, 7, 1454777. [Google Scholar] [CrossRef] [Green Version]
- Baczyk, D.; Drewlo, S.; Proctor, L.; Dunk, C.; Lye, S.; Kingdom, J. Glial cell missing-1 transcription factor is required for the differentiation of the human trophoblast. Cell Death Differ. 2009, 16, 719–727. [Google Scholar] [CrossRef]
- Bainbridge, S.A.; Minhas, A.; Whiteley, K.J.; Qu, D.; Sled, J.G.; Kingdom, J.C.; Adamson, S.L. Effects of reduced Gcm1 expression on trophoblast morphology, fetoplacental vascularity, and pregnancy outcomes in mice. Hypertension 2012, 59, 732–739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shahzad, K.; Bock, F.; Dong, W.; Wang, H.; Kopf, S.; Kohli, S.; Al-Dabet, M.M.; Ranjan, S.; Wolter, J.; Wacker, C.; et al. Nlrp3-inflammasome activation in non-myeloid-derived cells aggravates diabetic nephropathy. Kidney Int. 2015, 87, 74–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohli, S.; Singh, K.K.; Gupta, A.; Markmeyer, P.; Lochmann, F.; Gupta, D.; Rana, R.; Elwakiel, A.; Huebner, H.; Ruebner, M.; et al. Placental thrombo-inflammation impairs embryonic survival by reducing placental thrombomodulin expression. Blood 2020, 137, 977–982. [Google Scholar] [CrossRef]
- Wolter, J.; Schild, L.; Bock, F.; Hellwig, A.; Gadi, I.; Al-Dabet, M.M.; Ranjan, S.; Ronicke, R.; Nawroth, P.P.; Petersen, K.U.; et al. Thrombomodulin-dependent protein C activation is required for mitochondrial function and myelination in the central nervous system. J. Thromb. Haemost. JTH 2016, 14, 2212–2226. [Google Scholar] [CrossRef]
- Tanaka, S.; Kunath, T.; Hadjantonakis, A.K.; Nagy, A.; Rossant, J. Promotion of trophoblast stem cell proliferation by FGF4. Science 1998, 282, 2072–2075. [Google Scholar] [CrossRef]
- Kashif, M.; Hellwig, A.; Hashemolhosseini, S.; Kumar, V.; Bock, F.; Wang, H.; Shahzad, K.; Ranjan, S.; Wolter, J.; Madhusudhan, T.; et al. Nuclear factor erythroid-derived 2 (Nfe2) regulates JunD DNA-binding activity via acetylation: A novel mechanism regulating trophoblast differentiation. J. Biol. Chem. 2012, 287, 5400–5411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohli, S.; Hoffmann, J.; Lochmann, F.; Markmeyer, P.; Huebner, H.; Fahlbusch, F.B.; Al-Dabet, M.M.; Gadi, I.; Manoharan, J.; Lottge, M.; et al. p45 NF-E2 regulates syncytiotrophoblast differentiation by post-translational GCM1 modifications in human intrauterine growth restriction. Cell Death Dis. 2017, 8, e2730. [Google Scholar] [CrossRef]
- Huppertz, B. Trophoblast differentiation, fetal growth restriction and preeclampsia. Pregnancy Hypertens. 2011, 1, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Rondina, M.T. The Era of Thromboinflammation: Platelets Are Dynamic Sensors and Effector Cells During Infectious Diseases. Front. Immunol. 2019, 10, 2204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossant, J.; Cross, J.C. Placental development: Lessons from mouse mutants. Nat. Rev. Genet. 2001, 2, 538–548. [Google Scholar] [CrossRef]
- Cross, J.C.; Hemberger, M.; Lu, Y.; Nozaki, T.; Whiteley, K.; Masutani, M.; Adamson, S.L. Trophoblast functions, angiogenesis and remodeling of the maternal vasculature in the placenta. Mol. Cell. Endocrinol. 2002, 187, 207–212. [Google Scholar] [CrossRef]
- Akison, L.K.; Nitert, M.D.; Clifton, V.L.; Moritz, K.M.; Simmons, D.G. Review: Alterations in placental glycogen deposition in complicated pregnancies: Current preclinical and clinical evidence. Placenta 2017, 54, 52–58. [Google Scholar] [CrossRef] [Green Version]
- Motta-Mejia, C.; Kandzija, N.; Zhang, W.; Mhlomi, V.; Cerdeira, A.S.; Burdujan, A.; Tannetta, D.; Dragovic, R.; Sargent, I.L.; Redman, C.W.; et al. Placental Vesicles Carry Active Endothelial Nitric Oxide Synthase and Their Activity is Reduced in Preeclampsia. Hypertension 2017, 70, 372–381. [Google Scholar] [CrossRef] [PubMed]
- Forstner, D.; Maninger, S.; Nonn, O.; Guettler, J.; Moser, G.; Leitinger, G.; Pritz, E.; Strunk, D.; Schallmoser, K.; Marsche, G.; et al. Platelet-derived factors impair placental chorionic gonadotropin beta-subunit synthesis. J. Mol. Med. 2020, 98, 193–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Sivasubramaniyam, T.; Yinon, Y.; Tagliaferro, A.; Ray, J.; Nevo, O.; Post, M.; Caniggia, I. Aberrant TGFbeta Signaling Contributes to Altered Trophoblast Differentiation in Preeclampsia. Endocrinology 2016, 157, 883–899. [Google Scholar] [CrossRef] [Green Version]
- Yusuf, K.; Smith, S.D.; Levy, R.; Schaiff, W.T.; Wyatt, S.M.; Sadovsky, Y.; Nelson, D.M. Thromboxane A(2) limits differentiation and enhances apoptosis of cultured human trophoblasts. Pediatric Res. 2001, 50, 203–209. [Google Scholar] [CrossRef] [Green Version]
- Nicola, C.; Chirpac, A.; Lala, P.K.; Chakraborty, C. Roles of Rho guanosine 5′-triphosphatase A, Rho kinases, and extracellular signal regulated kinase (1/2) in prostaglandin E2-mediated migration of first-trimester human extravillous trophoblast. Endocrinology 2008, 149, 1243–1251. [Google Scholar] [CrossRef]
- Sato, Y.; Fujiwara, H.; Zeng, B.X.; Higuchi, T.; Yoshioka, S.; Fujii, S. Platelet-derived soluble factors induce human extravillous trophoblast migration and differentiation: Platelets are a possible regulator of trophoblast infiltration into maternal spiral arteries. Blood 2005, 106, 428–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujiwara, H.; Matsumoto, H.; Sato, Y.; Horie, A.; Ono, M.; Nakamura, M.; Mizumoto, Y.; Kagami, K.; Fujiwara, T.; Hattori, A.; et al. Factors Regulating Human Extravillous Trophoblast Invasion: Chemokine-peptidase and CD9-integrin Systems. Curr. Pharm. Biotechnol. 2018, 19, 764–770. [Google Scholar] [CrossRef]
- Phillips, R.J.; Fortier, M.A.; Lopez Bernal, A. Prostaglandin pathway gene expression in human placenta, amnion and choriodecidua is differentially affected by preterm and term labour and by uterine inflammation. BMC Pregnancy Childbirth 2014, 14, 241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomez-Lopez, N.; Motomura, K.; Miller, D.; Garcia-Flores, V.; Galaz, J.; Romero, R. Inflammasomes: Their Role in Normal and Complicated Pregnancies. J. Immunol. 2019, 203, 2757–2769. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Markmeyer, P.; Lochmann, F.; Singh, K.K.; Gupta, A.; Younis, R.; Shahzad, K.; Biemann, R.; Huebner, H.; Ruebner, M.; Isermann, B.; et al. Procoagulant Extracellular Vesicles Alter Trophoblast Differentiation in Mice by a Thrombo-Inflammatory Mechanism. Int. J. Mol. Sci. 2021, 22, 9873. https://doi.org/10.3390/ijms22189873
Markmeyer P, Lochmann F, Singh KK, Gupta A, Younis R, Shahzad K, Biemann R, Huebner H, Ruebner M, Isermann B, et al. Procoagulant Extracellular Vesicles Alter Trophoblast Differentiation in Mice by a Thrombo-Inflammatory Mechanism. International Journal of Molecular Sciences. 2021; 22(18):9873. https://doi.org/10.3390/ijms22189873
Chicago/Turabian StyleMarkmeyer, Paulina, Franziska Lochmann, Kunal Kumar Singh, Anubhuti Gupta, Ruaa Younis, Khurrum Shahzad, Ronald Biemann, Hanna Huebner, Matthias Ruebner, Berend Isermann, and et al. 2021. "Procoagulant Extracellular Vesicles Alter Trophoblast Differentiation in Mice by a Thrombo-Inflammatory Mechanism" International Journal of Molecular Sciences 22, no. 18: 9873. https://doi.org/10.3390/ijms22189873
APA StyleMarkmeyer, P., Lochmann, F., Singh, K. K., Gupta, A., Younis, R., Shahzad, K., Biemann, R., Huebner, H., Ruebner, M., Isermann, B., & Kohli, S. (2021). Procoagulant Extracellular Vesicles Alter Trophoblast Differentiation in Mice by a Thrombo-Inflammatory Mechanism. International Journal of Molecular Sciences, 22(18), 9873. https://doi.org/10.3390/ijms22189873