
The Role of Extracellular Vesicles from Human Macrophages on Host-Pathogen Interaction


Abstract
:1. Introduction
2. Immunoregulatory Properties of Macrophage-Derived EVs
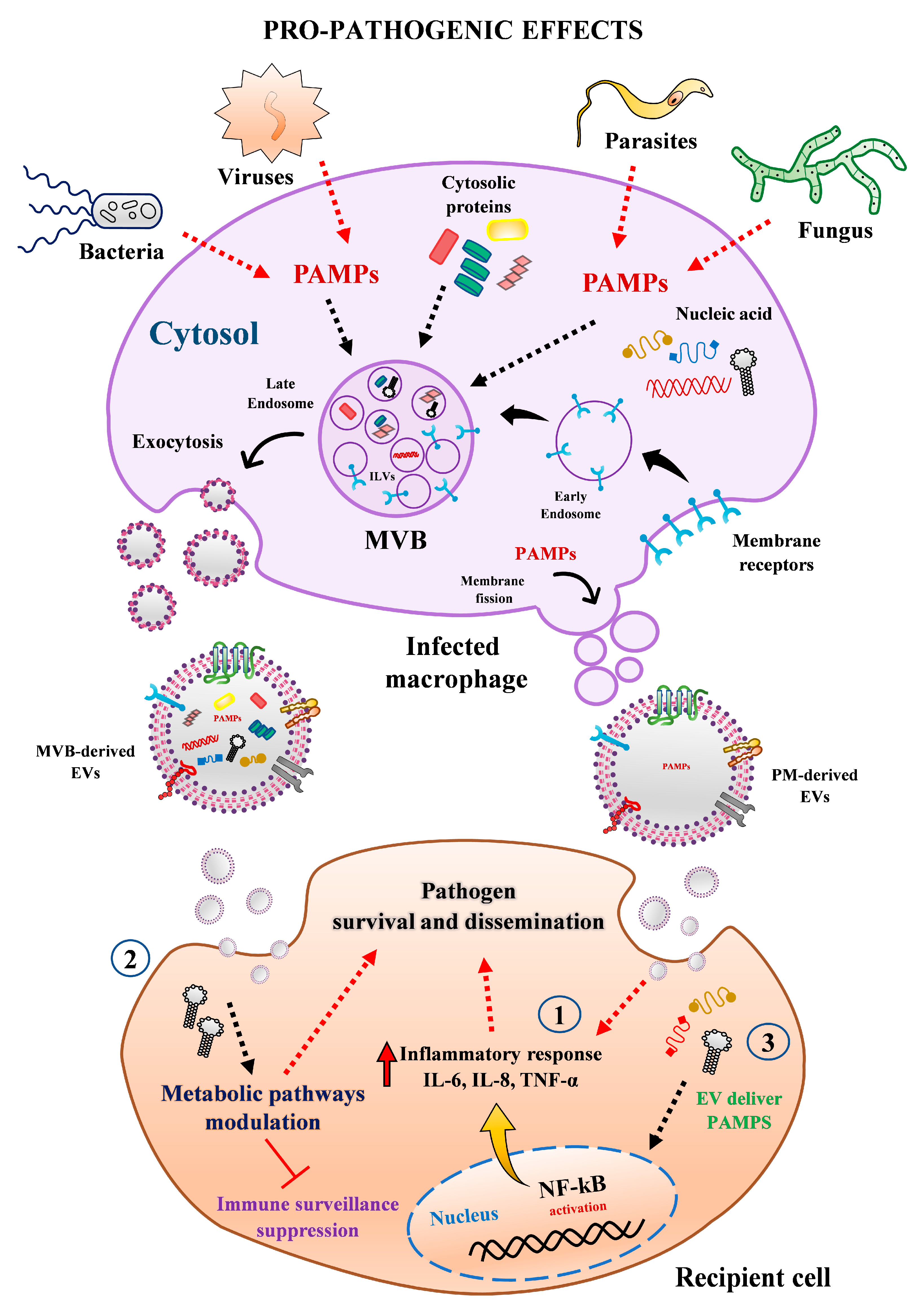
3. Human Macrophage-Derived EVs and Bacterial Infections
3.1. EVs from Macrophages Can Potentiate Bacterial Infections
3.2. Macrophage-EVs Can Contribute to Development of Immune Responses against Bacterial Infections
4. EVs from Human Macrophages and Viral Infections
4.1. Stimulatory Role of Macrophage-Derived EVs during Viral Infections
4.2. Inhibitory Effects of Macrophage-Derived EVs during Viral Infections
5. EVs from Macrophages and Parasitic Infections
6. Macrophage-Derived EVs and Fungal Infections
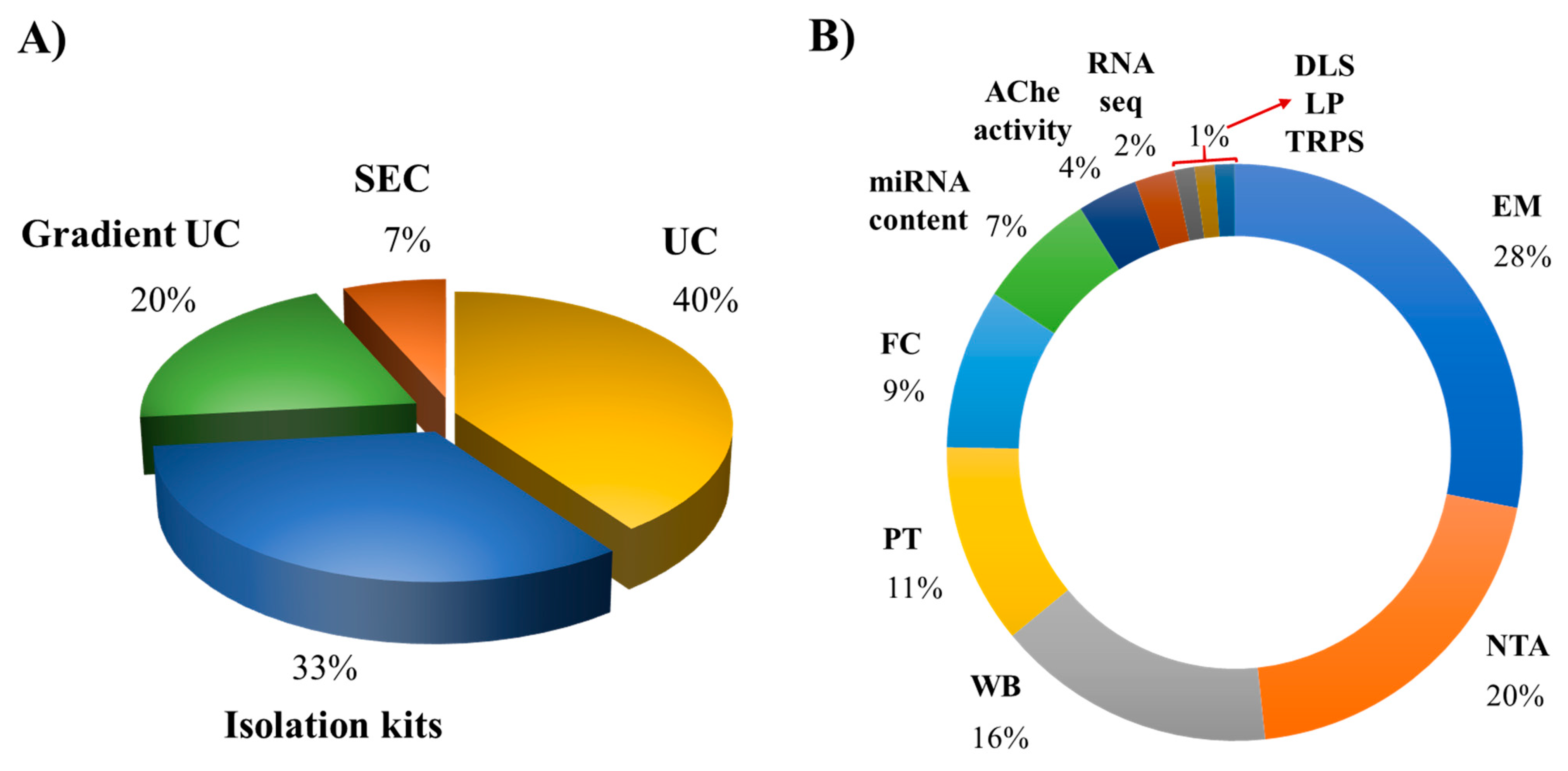
7. Methods for Isolation and Characterization of EVs from Human Macrophages
8. Macrophage-EVs as Diagnostic Markers for Infectious Diseases
9. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7. [Google Scholar] [CrossRef] [Green Version]
- Witwer, K.W.; Théry, C. Extracellular vesicles or exosomes? On primacy, precision, and popularity influencing a choice of nomenclature. J. Extracell. Vesicles 2019, 8. [Google Scholar] [CrossRef] [PubMed]
- Colombo, M.; Raposo, G.; Théry, C. Biogenesis, secretion, and intercellular interactions of exosomes and other extracellular vesicles. Annu. Rev. Cell Dev. Biol. 2014, 30, 255–289. [Google Scholar] [CrossRef] [PubMed]
- Van Niel, G.; D’Angelo, G.; Raposo, G. Shedding light on the cell biology of extracellular vesicles. Nat. Rev. Mol. Cell Biol. 2018, 19, 213–228. [Google Scholar] [CrossRef] [PubMed]
- Yáñez-Mó, M.; Siljander, P.R.-M.; Andreu, Z.; Zavec, A.B.; Borràs, F.E.; Buzas, E.I.; Buzas, K.; Casal, E.; Cappello, F.; Carvalho, J.; et al. Biological Properties of Extracellular Vesicles and their Physiological Functions. J. Extracell. Vesicles 2015, 4, 27066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathieu, M.; Martin-Jaular, L.; Lavieu, G.; Théry, C. Specificities of secretion and uptake of exosomes and other extracellular vesicles for cell-to-cell communication. Nat. Cell Biol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Cocucci, E.; Racchetti, G.; Podini, P.; Meldolesi, J. Enlargeosome Traffic: Exocytosis Triggered by Various Signals Is Followed by Endocytosis, Membrane Shedding or Both. Traffic 2007, 8, 742–757. [Google Scholar] [CrossRef]
- Tkach, M.; Kowal, J.; Théry, C. Why the need and how to approach the functional diversity of extracellular vesicles. Philos. Trans. R. Soc. B Biol. Sci. 2018, 373. [Google Scholar] [CrossRef]
- Johnstone, R.M.; Adam, M.; Hammond, J.R.; Orr, L.; Turbide, C. Vesicle formation during reticulocyte maturation. Association of plasma membrane activities with released vesicles (exosomes). J. Biol. Chem. 1987, 262, 9412–9420. [Google Scholar] [CrossRef]
- Skotland, T.; Hessvik, N.P.; Sandvig, K.; Llorente, A. Exosomal lipid composition and the role of ether lipids and phosphoinositides in exosome biology. J. Lipid Res. 2019, 60, 9–18. [Google Scholar] [CrossRef] [Green Version]
- Marcilla, A.; Jaular, L.M.; Trelis, M.; Menezes-Neto, A.; Osuna, A.; Bernal, D.; Fernandez-Becerra, C.; Almeida, I.; Del Portillo, H.A. Extracellular vesicles in parasitic diseases. J. Extracell. Vesicles 2014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schorey, J.S.; Harding, C.V. Extracellular vesicles and infectious diseases: New complexity to an old story. J. Clin. Investig. 2016, 1181–1189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caobi, A.; Nair, M.; Raymond, A.D. Extracellular Vesicles in the Pathogenesis of Viral Infections in Humans. Viruses 2020, 12, 1200. [Google Scholar] [CrossRef]
- Coakley, G.; Maizels, R.M.; Buck, A.H. Exosomes and Other Extracellular Vesicles: The New Communicators in Parasite Infections. Trends Parasitol. 2015, 477–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuipers, M.E.; Hokke, C.H.; Smits, H.H.; Hoen, E.N.M.N. T Pathogen-Derived Extracellular Vesicle-Associated Molecules That Affect the Host Immune System: An Overview. Front. Microbiol. 2018. [Google Scholar] [CrossRef] [Green Version]
- Rizzo, J.; Rodrigues, M.L.; Janbon, G. Extracellular Vesicles in Fungi: Past, Present, and Future Perspectives. Front. Cell. Infect. Microbiol. 2020, 346. [Google Scholar] [CrossRef]
- Spencer, N.; Yeruva, L. Role of bacterial infections in extracellular vesicles release and impact on immune response. Biomed. J. 2020. [Google Scholar] [CrossRef]
- Torrecilhas, A.C.; Soares, R.P.; Schenkman, S.; Fernández-Prada, C.; Olivier, M. Extracellular Vesicles in Trypanosomatids: Host Cell Communication. Front. Cell. Infect. Microbiol. 2020, 750. [Google Scholar] [CrossRef]
- Costa, A.O.; Chagas, I.A.R.; de Menezes-Neto, A.; Rêgo, F.D.; Nogueira, P.M.; Torrecilhas, A.C.; Furst, C.; Fux, B.; Soares, R.P. Distinct immunomodulatory properties of extracellular vesicles released by different strains of Acanthamoeba. Cell Biol. Int. 2021, 45, 1060–1071. [Google Scholar] [CrossRef]
- Gordon, S. The macrophage: Past, present and future. Eur. J. Immunol. 2007, 37, S9–S17. [Google Scholar] [CrossRef]
- Gordon, S.; Martinez-Pomares, L. Physiological roles of macrophages. Pflüg. Arch. Eur. J. Physiol. 2017, 469, 365–374. [Google Scholar] [CrossRef] [Green Version]
- Wynn, T.A.; Chawla, A.; Pollard, J.W. Macrophage biology in development, homeostasis and disease. Nature 2013, 496, 445–455. [Google Scholar] [CrossRef]
- Flannagan, R.; Heit, B.; Heinrichs, D.E. Antimicrobial Mechanisms of Macrophages and the Immune Evasion Strategies of Staphylococcus aureus. Pathogens 2015, 4, 826–868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jurberg, A.D.; Cotta-De-Almeida, V.; Temerozo, J.; Savino, W.; Bou-Habib, D.C.; Riederer, I. Neuroendocrine Control of Macrophage Development and Function. Front. Immunol. 2018, 9, 1440. [Google Scholar] [CrossRef]
- Gracia-Hernandez, M.; Sotomayor, E.M.; Villagra, A. Targeting Macrophages as a Therapeutic Option in Coronavirus Disease 2019. Front. Pharmacol. 2020, 11, 577571. [Google Scholar] [CrossRef]
- Torraca, V.; Masud, S.; Spaink, H.; Meijer, A.H. Macrophage-pathogen interactions in infectious diseases: New therapeutic insights from the zebrafish host model. Dis. Model. Mech. 2014, 785–797. [Google Scholar] [CrossRef] [Green Version]
- Atri, C.; Guerfali, F.Z.; Laouini, D. Role of Human Macrophage Polarization in Inflammation during Infectious Diseases. Int. J. Mol. Sci. 2018, 19, 1801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, R.; Rai, A.; Chen, M.; Suwakulsiri, W.; Greening, D.W.; Simpson, R.J. Extracellular vesicles in cancer—Implications for future improvements in cancer care. Nat. Rev. Clin. Oncol. 2018, 617–638. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.-C.; Song, S.-J.; Zhang, Y.; Li, X. Role of Extracellular Vesicles in Autoimmune Pathogenesis. Front. Immunol. 2020, 579043. [Google Scholar] [CrossRef]
- Yin, Z.; Fan, J.; Xu, J.; Wu, F.; Li, Y.; Zhou, M.; Liao, T.; Duan, L.; Wang, S.; Geng, W.; et al. Immunoregulatory Roles of Extracellular Vesicles and Associated Therapeutic Applications in Lung Cancer. Front. Immunol. 2020. [Google Scholar] [CrossRef]
- Kowal, J.; Tkach, M. Dendritic cell extracellular vesicles. Int. Rev. Cell Mol. Biol. 2019, 213–249. [Google Scholar] [CrossRef]
- Federici, C.; Shahaj, E.; Cecchetti, S.; Camerini, S.; Casella, M.; Iessi, E.; Camisaschi, C.; Paolino, G.; Calvieri, S.; Ferro, S.; et al. Natural-killer-derived extracellular vesicles: Immune sensors and interactors. Front. Immunol. 2020, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kormelink, T.G.; Mol, S.; De Jong, E.C.; Wauben, M. The role of extracellular vesicles when innate meets adaptive. Semin. Immunopathol. 2018, 439–452. [Google Scholar] [CrossRef] [Green Version]
- Ismail, N.; Wang, Y.; Dakhlallah, D.; Moldovan, L.; Agarwal, K.; Batte, K.; Shah, P.; Wisler, J.; Eubank, T.D.; Tridandapani, S.; et al. Macrophage microvesicles induce macrophage differentiation and miR-223 transfer. Blood 2013, 121, 984–995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, L.; Wang, Y.; Huang, L. Exosomes from M1-Polarized Macrophages Potentiate the Cancer Vaccine by Creating a Pro-inflammatory Microenvironment in the Lymph Node. Mol. Ther. 2017, 25, 1665–1675. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Zhao, M.; Liu, S.; Guo, J.; Lu, Y.; Cheng, J.; Liu, J. Macrophage-derived extracellular vesicles: Diverse mediators of pathology and therapeutics in multiple diseases. Cell Death Dis. 2020. [Google Scholar] [CrossRef] [PubMed]
- Giri, P.K.; Schorey, J.S. Exosomes Derived from M. bovis BCG Infected Macrophages Activate Antigen-Specific CD4+ and CD8+ T Cells in vitro and in vivo. PLoS ONE 2008, 3, e2461. [Google Scholar] [CrossRef] [Green Version]
- Robbins, P.D.; Morelli, A.E. Regulation of immune responses by extracellular vesicles. Nat. Rev. Immunol. 2014, 195–208. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, M.; Fan, J.; Lyon, C.; Wan, M.; Hu, T. Role of Extracellular Vesicles in Viral and Bacterial Infections: Pathogenesis, Diagnostics, and Therapeutics. Theranostics 2018, 2709–2721. [Google Scholar] [CrossRef]
- Volgers, C.; Benedikter, B.J.; Grauls, G.E.; Savelkoul, P.H.M.; Stassen, F.R.M. Immunomodulatory role for membrane vesicles released by THP-1 macrophages and respiratory pathogens during macrophage infection. BMC Microbiol. 2017, 17, 216. [Google Scholar] [CrossRef] [Green Version]
- Carrière, J.; Bretin, A.; Darfeuille-Michaud, A.; Barnich, N.; Nguyen, H.T.T. Exosomes Released from Cells Infected with Crohn’s Disease–Associated Adherent-Invasive Escherichia coli Activate Host Innate Immune Responses and Enhance Bacterial Intracellular Replication. Inflamm. Bowel Dis. 2016, 22, 516–528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alipoor, S.D.; Mortaz, E.; Tabarsi, P.; Farnia, P.; Mirsaeidi, M.; Garssen, J.; Movassaghi, M.; Adcock, I.M. Bovis Bacillus Calmette–Guerin (BCG) infection induces exosomal miRNA release by human macrophages. J. Transl. Med. 2017, 15. [Google Scholar] [CrossRef] [Green Version]
- Xu, B.-F.; Wang, Q.-Q.; Zhang, J.-P.; Hu, W.-L.; Zhang, R.-L. Treponema pallidum induces the activation of endothelial cells via macrophage-derived exosomes. Arch. Dermatol. Res. 2019, 311, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Xu, B.; Zhang, J.; Kou, C.; Liu, J.; Wang, Q.; Zhang, R. Exosomal miR-146a-5p from Treponema pallidum-stimulated macrophages reduces endothelial cells permeability and monocyte transendothelial migration by targeting JAM-C. Exp. Cell Res. 2020, 388. [Google Scholar] [CrossRef]
- Diaz, G.; Wolfe, L.M.; Kruh-Garcia, N.; Dobos, K.M. Changes in the Membrane-Associated Proteins of Exosomes Released from Human Macrophages after Mycobacterium tuberculosis Infection. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef] [Green Version]
- Bhatnagar, S.; Shinagawa, K.; Castellino, F.J.; Schorey, J.S. Exosomes released from macrophages infected with intracellular pathogens stimulate a proinflammatory response in vitro and in vivo. Blood 2007, 110, 3234–3244. [Google Scholar] [CrossRef] [Green Version]
- Walters, S.B.; Kieckbusch, J.; Nagalingam, G.; Swain, A.; Latham, S.L.; Grau, G.E.R.; Britton, W.J.; Combes, V.; Saunders, B.M. Microparticles from Mycobacteria-Infected Macrophages Promote Inflammation and Cellular Migration. J. Immunol. 2012, 190, 669–677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hare, N.J.; Chan, B.; Chan, E.; Kaufman, K.L.; Britton, W.J.; Saunders, B. Microparticles released fromMycobacterium tuberculosis-infected human macrophages contain increased levels of the type I interferon inducible proteins including ISG15. Proteomics 2015, 15, 3020–3029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.-J.; Chen, C.; Xie, P.-F.; Pan, Y.; Tan, Y.-H.; Tang, L.-J. Proteomic analysis and immune properties of exosomes released by macrophages infected with Mycobacterium avium. Microbes Infect. 2014, 16, 283–291. [Google Scholar] [CrossRef]
- Wang, J.; Yao, Y.; Xiong, J.; Wu, J.; Tang, X.; Li, G. Evaluation of the Inflammatory Response in Macrophages Stimulated with Exosomes Secreted by Mycobacterium avium-Infected Macrophages. BioMed Res. Int. 2015, 2015, 658421. [Google Scholar] [CrossRef] [Green Version]
- Pawar, K.; Shigematsu, M.; Sharbati, S.; Kirino, Y. Infection-induced 5′-half molecules of tRNAHisGUG activate Toll-like receptor 7. PLoS Biol. 2020, 18, e3000982. [Google Scholar] [CrossRef]
- Jung, A.L.; Herkt, C.E.; Schulz, C.; Bolte, K.; Seidel, K.; Scheller, N.; Sittka-Stark, A.; Bertrams, W.; Schmeck, B. Legionella pneumophila infection activates bystander cells differentially by bacterial and host cell vesicles. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [Green Version]
- Hui, W.W.; Hercik, K.; Belsare, S.; Alugubelly, N.; Clapp, B.; Rinaldi, C.; Edelmann, M.J. Salmonella enterica Serovar Typhimurium Alters the Extracellular Proteome of Macrophages and Leads to the Production of Proinflammatory Exosomes. Infect. Immun. 2018, 86. [Google Scholar] [CrossRef] [Green Version]
- Kadiu, I.; Narayanasamy, P.; Dash, P.K.; Zhang, W.; Gendelman, H.E. Biochemical and Biologic Characterization of Exosomes and Microvesicles as Facilitators of HIV-1 Infection in Macrophages. J. Immunol. 2012, 189, 744–754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arenaccio, C.; Anticoli, S.; Manfredi, F.; Chiozzini, C.; Olivetta, E.; Federico, M. Latent HIV-1 is activated by exosomes from cells infected with either replication-competent or defective HIV-1. Retrovirology 2015, 12, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernard, M.A.; Zhao, H.; Yue, S.C.; Anandaiah, A.; Koziel, H.; Tachado, S.D. Novel HIV-1 MiRNAs Stimulate TNFα Release in Human Macrophages via TLR8 Signaling Pathway. PLoS ONE 2014, 9, e106006. [Google Scholar] [CrossRef]
- Roth, W.W.; Huang, M.B.; Konadu, K.A.; Powell, M.D.; Bond, V.C. Micro RNA in Exosomes from HIV-Infected Macrophages. Int. J. Environ. Res. Public Health 2016, 13, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, Z.; Petree, J.R.; Lee, F.E.-H.; Fan, X.; Salaita, K.; Guidot, D.M.; Sadikot, R.T. Macrophages exposed to HIV viral protein disrupt lung epithelial cell integrity and mitochondrial bioenergetics via exosomal microRNA shuttling. Cell Death Dis. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Sharma, H.; Chinnappan, M.; Agarwal, S.; Dalvi, P.; Gunewardena, S.; O’Brien-Ladner, A.; Dhillon, N.K. Macrophage-derived extracellular vesicles mediate smooth muscle hyperplasia: Role of altered miRNA cargo in response to HIV infection and substance abuse. FASEB J. 2018, 32, 5174–5185. [Google Scholar] [CrossRef]
- Cantres-Rosario, Y.M.; Ortiz-Rodríguez, S.C.; Santos-Figueroa, A.G.; Plaud, M.; Negron, K.; Cotto, B.; Langford, D.; Melendez, L.M. HIV Infection Induces Extracellular Cathepsin B Uptake and Damage to Neurons. Sci. Rep. 2019, 9. [Google Scholar] [CrossRef]
- Okamoto, M.; Fukushima, Y.; Kouwaki, T.; Daito, T.; Kohara, M.; Kida, H.; Oshiumi, H. MicroRNA-451a in extracellular, blood-resident vesicles attenuates macrophage and dendritic cell responses to influenza whole-virus vaccine. J. Biol. Chem. 2018, 293, 18585–18600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Velandia-Romero, M.L.; Calderón-Peláez, M.A.; Balbás-Tepedino, A.; Márquez-Ortiz, R.A.; Madroñero, L.J.; Prieto, A.B.; Castellanos, J.E. Extracellular vesicles of U937 macrophage cell line infected with DENV-2 induce activation in endothelial cells EA.hy926. PLoS ONE 2020, 15, e0227030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, Z.; Qiao, Y.; Li, X.; Chen, J.; Ding, J.; Bai, L.; Shen, F.; Shi, B.; Liu, J.; Peng, L.; et al. Exosomes Exploit the Virus Entry Machinery and Pathway to Transmit Alpha Interferon-Induced Antiviral Activity. J. Virol. 2018, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Wang, X.; Sun, L.; Zhou, L.; Ma, T.; Song, L.; Wu, J.; Li, J.; Ho, W. Toll-like receptor 3-activated macrophages confer anti-HCV activity to hepatocytes through exosomes. FASEB J. 2016, 30, 4132–4140. [Google Scholar] [CrossRef] [Green Version]
- Cai, C.; Koch, B.; Morikawa, K.; Suda, G.; Sakamoto, N.; Rueschenbaum, S.; Akhras, S.; Dietz, J.; Hildt, E.; Zeuzem, S.; et al. Macrophage-Derived Extracellular Vesicles Induce Long-Lasting Immunity Against Hepatitis C Virus which is Blunted by Polyunsaturated Fatty Acids. Front. Immunol. 2018, 9, 723. [Google Scholar] [CrossRef]
- Cestari, I.; Ansa-Addo, E.; Deolindo, P.; Inal, J.; Ramirez, M.I. Trypanosoma cruziImmune Evasion Mediated by Host Cell-Derived Microvesicles. J. Immunol. 2012, 188, 1942–1952. [Google Scholar] [CrossRef] [Green Version]
- Cronemberger-Andrade, A.; Xander, P.; Soares, R.P.; Pessoa, N.L.; Campos, M.A.; Ellis, C.C.; Grajeda, B.; Ofir-Birin, Y.; Almeida, I.C.; Regev-Rudzki, N.; et al. Trypanosoma cruzi-Infected Human Macrophages Shed Proinflammatory Extracellular Vesicles That Enhance Host-Cell Invasion via Toll-Like Receptor 2. Front. Cell. Infect. Microbiol. 2020, 10, 99. [Google Scholar] [CrossRef]
- Cypryk, W.; Öhman, T.; Eskelinen, E.-L.; Matikainen, S.; Nyman, T.A. Quantitative Proteomics of Extracellular Vesicles Released from Human Monocyte-Derived Macrophages upon β-Glucan Stimulation. J. Proteome Res. 2014, 13, 2468–2477. [Google Scholar] [CrossRef]
- Reales-Calderón, J.A.; Vaz, C.; Monteoliva, L.; Molero, G.; Gil, C. Candida albicans Modifies the Protein Composition and Size Distribution of THP-1 Macrophage-Derived Extracellular Vesicles. J. Proteome Res. 2017, 16, 87–105. [Google Scholar] [CrossRef]
- Anand, P.K.; Anand, E.; Bleck, C.; Anes, E.; Griffiths, G. Exosomal Hsp70 Induces a Pro-Inflammatory Response to Foreign Particles Including Mycobacteria. PLoS ONE 2010, 5, e10136. [Google Scholar] [CrossRef]
- Floyd, K.; Raviglione, M.C.; Glaziou, P. Global Epidemiology of Tuberculosis. Semin. Respir. Crit. Care Med. 2018, 39, 271–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, D.R.; Mello, F.C.D.Q.; Migliori, G.B. Tuberculosis series 2020. J. Bras. Pneumol. 2020, 46. [Google Scholar] [CrossRef]
- O’Neill, S.K.; Cao, Y.; Hamel, K.M.; Doodes, P.D.; Hutas, G.; Finnegan, A. Expression of CD80/86 on B cells is essential for autoreactive T cell activation and the development of arthritis. J. Immunol. 2007, 179, 5109–5116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dias, M.V.S.; Costa, C.S.; DaSilva, L.L.P. The Ambiguous Roles of Extracellular Vesicles in HIV Replication and Pathogenesis. Front. Microbiol. 2018. [Google Scholar] [CrossRef] [Green Version]
- Pérez, P.S.; Romaniuk, M.A.; Duette, G.A.; Zhao, Z.; Huang, Y.; Martin-Jaular, L.; Witwer, K.W.; Théry, C.; Ostrowski, M. Extracellular vesicles and chronic inflammation during HIV infection. J. Extracell. Vesicles 2019, 8, 1687275. [Google Scholar] [CrossRef] [PubMed]
- Urbanelli, L.; Buratta, S.; Tancini, B.; Sagini, K.; Delo, F.; Porcellati, S.; Emiliani, C. The Role of Extracellular Vesicles in Viral Infection and Transmission. Vaccines 2019, 7, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ipinmoroti, A.O.; Matthews, Q.L. Extracellular Vesicles: Roles in Human Viral Infections, Immune-Diagnostic, and Therapeutic Applications. Pathogens 2020, 9, 1056. [Google Scholar] [CrossRef]
- Giannessi, F.; Aiello, A.; Franchi, F.; Percario, Z.A.; Affabris, E. The Role of Extracellular Vesicles as Allies of HIV, HCV and SARS Viruses. Viruses 2020, 12, 571. [Google Scholar] [CrossRef]
- Martins, S.D.T.; Alves, L.R. Extracellular Vesicles in Viral Infections: Two Sides of the Same Coin? Front. Cell. Infect. Microbiol. 2020, 10, 593170. [Google Scholar] [CrossRef]
- Jiang, Y.; Cai, X.; Yao, J.; Guo, H.; Yin, L.; Leung, W.; Xu, C. Role of Extracellular Vesicles in Influenza Virus Infection. Front. Cell. Infect. Microbiol. 2020, 366. [Google Scholar] [CrossRef]
- Votteler, J.; Sundquist, W.I. Virus Budding and the ESCRT Pathway. Cell Host Microbe 2013, 232–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadeghipour, S.; Mathias, R.A. Herpesviruses hijack host exosomes for viral pathogenesis. Semin. Cell Dev. Biol. 2017, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Van Der Grein, S.G.; Defourny, K.A.Y.; Slot, E.F.J.; Hoen, E.N.M.N. T Intricate relationships between naked viruses and extracellular vesicles in the crosstalk between pathogen and host. Semin. Immunopathol. 2018, 491–504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konadu, K.A.; Chu, J.; Huang, M.B.; Amancha, P.K.; Armstrong, W.S.; Powell, M.D.; Villinger, F.; Bond, V.C. Association of Cytokines with Exosomes in the Plasma of HIV-1–Seropositive Individuals. J. Infect. Dis. 2015, 211, 1712–1716. [Google Scholar] [CrossRef]
- Luron, L.; Saliba, D.; Blazek, K.; Lanfrancotti, A.; Udalova, I.A. FOXO3 as a new IKK-ε-controlled check-point of regulation of IFN-β expression. Eur. J. Immunol. 2012, 42, 1030–1037. [Google Scholar] [CrossRef] [PubMed]
- Rosenberger, C.M.; Podyminogin, R.L.; Navarro, G.; Zhao, G.-W.; Askovich, P.S.; Weiss, M.; Aderem, A. miR-451 Regulates Dendritic Cell Cytokine Responses to Influenza Infection. J. Immunol. 2012, 189, 5965–5975. [Google Scholar] [CrossRef] [Green Version]
- Hassani, K.; Olivier, M. Immunomodulatory Impact of Leishmania-Induced Macrophage Exosomes: A Comparative Proteomic and Functional Analysis. PLoS Negl. Trop. Dis. 2013, 7, e2158. [Google Scholar] [CrossRef]
- Ghosh, J.; Bose, M.; Roy, S.; Bhattacharyya, S.N. Leishmania donovani Targets Dicer1 to Downregulate miR-122, Lower Serum Cholesterol, and Facilitate Murine Liver Infection. Cell Host Microbe 2013, 13, 277–288. [Google Scholar] [CrossRef] [Green Version]
- Cronemberger-Andrade, A.; Aragão-França, L.; De Araujo, C.F.; Rocha, V.J.; Silva, M.D.C.B.; Figueiras, C.P.; Oliveira, P.R.; De Freitas, L.A.R.; Veras, P.S.T.; Pontes-De-Carvalho, L. Extracellular Vesicles from Leishmania-Infected Macrophages Confer an Anti-infection Cytokine-Production Profile to Naïve Macrophages. PLoS Negl. Trop. Dis. 2014, 8, e3161. [Google Scholar] [CrossRef] [Green Version]
- Gioseffi, A.; Hamerly, T.; Van, K.; Zhang, N.; Dinglasan, R.R.; Yates, P.A.; Kima, P.E. Leishmania-infected macrophages release extracellular vesicles that can promote lesion development. Life Sci. Alliance 2020, 3. [Google Scholar] [CrossRef]
- Nieuwland, R.; Falcón-Pérez, J.M.; Théry, C.; Witwer, K.W. Rigor and standardization of extracellular vesicle research: Paving the road towards robustness. J. Extracell. Vesicles 2020, 10. [Google Scholar] [CrossRef]
- Royo, F.; Théry, C.; Falcón-Pérez, J.M.; Nieuwland, R.; Witwer, K.W. Methods for Separation and Characterization of Extracellular Vesicles: Results of a Worldwide Survey Performed by the ISEV Rigor and Standardization Subcommittee. Cells 2020, 9, 1955. [Google Scholar] [CrossRef]
- Chiriacò, M.S.; Bianco, M.; Nigro, A.; Primiceri, E.; Ferrara, F.; Romano, A.; Quattrini, A.; Furlan, R.; Arima, V.; Maruccio, G. Lab-on-Chip for Exosomes and Microvesicles Detection and Characterization. Sensors 2018, 18, 3175. [Google Scholar] [CrossRef] [Green Version]
- Hartjes, T.A.; Mytnyk, S.; Jenster, G.W.; Van Steijn, V.; Van Royen, M.E. Extracellular Vesicle Quantification and Characterization: Common Methods and Emerging Approaches. Bioengineering 2019, 6, 7. [Google Scholar] [CrossRef] [Green Version]
- Sidhom, K.; Obi, P.O.; Saleem, A. A Review of Exosomal Isolation Methods: Is Size Exclusion Chromatography the Best Option? Int. J. Mol. Sci. 2020, 21, 6466. [Google Scholar] [CrossRef] [PubMed]
- Arteaga-Blanco, L.A.; Mojoli, A.; Monteiro, R.Q.; Sandim, V.; Menna-Barreto, R.F.S.; Pereira-Dutra, F.S.; Bozza, P.T.; Resende, R.D.O.; Bou-Habib, D.C. Characterization and internalization of small extracellular vesicles released by human primary macrophages derived from circulating monocytes. PLoS ONE 2020, 15, e0237795. [Google Scholar] [CrossRef] [PubMed]
- Stremersch, S.; De Smedt, S.C.; Raemdonck, K. Therapeutic and diagnostic applications of extracellular vesicles. J. Control. Release 2016, 244, 167–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sahoo, S.; Adamiak, M.; Mathiyalagan, P.; Kenneweg, F.; Kafert-Kasting, S.; Thum, T. Therapeutic and Diagnostic Translation of Extracellular Vesicles in Cardiovascular Diseases. Circulation 2021, 143, 1426–1449. [Google Scholar] [CrossRef]
- Ka, M.B.; Daumas, A.; Textoris, J.; Mege, J.-L. Phenotypic Diversity and Emerging New Tools to Study Macrophage Activation in Bacterial Infectious Diseases. Front. Immunol. 2014. [Google Scholar] [CrossRef] [Green Version]
- Merad, M.; Martin, J.C. Author Correction: Pathological inflammation in patients with COVID-19: A key role for monocytes and macrophages. Nat. Rev. Immunol. 2020, 20, 355–362. [Google Scholar] [CrossRef] [PubMed]
- Moin, A.S.M.; Sathyapalan, T.; Diboun, I.; Atkin, S.L.; Butler, A.E. Identification of macrophage activation-related biomarkers in obese type 2 diabetes that may be indicative of enhanced respiratory risk in COVID-19. Sci. Rep. 2021, 11. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Yang, M.; Ericsson, A.C. Function of Macrophages in Disease: Current Understanding on Molecular Mechanisms. Front. Immunol. 2021, 620510. [Google Scholar] [CrossRef]
- Lidofsky, A.; Holmes, J.A.; Feeney, E.R.; Kruger, A.J.; Salloum, S.; Zheng, H.; Seguin, I.S.; Altinbas, A.; Masia, R.; Corey, K.E.; et al. Macrophage Activation Marker Soluble CD163 Is a Dynamic Marker of Liver Fibrogenesis in Human Immunodeficiency Virus/Hepatitis C Virus Coinfection. J. Infect. Dis. 2018, 218, 1394–1403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Relster, M.M.; Gaini, S.; Møller, H.J.; Johansen, I.S.; Pedersen, C. The macrophage activation marker sMR as a diagnostic and prognostic marker in patients with acute infectious disease with or without sepsis. Scand. J. Clin. Lab. Investig. 2018, 78, 180–186. [Google Scholar] [CrossRef]
- Fu, Y.; Yang, X.; Chen, H.; Lu, Y. Diagnostic value of miR-145 and its regulatory role in macrophage immune response in tuberculosis. Genet. Mol. Biol. 2020, 43. [Google Scholar] [CrossRef]
- Hadjadj, J.; Yatim, N.; Barnabei, L.; Corneau, A.; Boussier, J.; Smith, N.; Péré, H.; Charbit, B.; Bondet, V.; Chenevier-Gobeaux, C.; et al. Impaired type I interferon activity and inflammatory responses in severe COVID-19 patients. Science 2020, 369, 718–724. [Google Scholar] [CrossRef] [PubMed]
- György, B.; Hung, M.; Breakefield, X.O.; Leonard, J.N. Therapeutic Applications of Extracellular Vesicles: Clinical Promise and Open Questions. Annu. Rev. Pharmacol. Toxicol. 2015, 55, 439–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crenshaw, B.J.; Sims, B.; Matthews, Q.L. Biological Function of Exosomes as Diagnostic Markers and Therapeutic Delivery Vehicles in Carcinogenesis and Infectious Diseases. Nanomedicines 2019. [Google Scholar] [CrossRef] [Green Version]
- Rontogianni, S.; Synadaki, E.; Li, B.; Liefaard, M.C.; Lips, E.H.; Wesseling, J.; Wu, W.; Altelaar, M. Proteomic profiling of extracellular vesicles allows for human breast cancer subtyping. Commun. Biol. 2019, 2. [Google Scholar] [CrossRef] [Green Version]
- Weber, B.; Franz, N.; Marzi, I.; Henrich, D.; Leppik, L. Extracellular vesicles as mediators and markers of acute organ injury: Current concepts. Eur. J. Trauma Emerg. Surg. 2021. [Google Scholar] [CrossRef]
- Vlassov, A.V.; Magdaleno, S.; Setterquist, R.; Conrad, R. Exosomes: Current knowledge of their composition, biological functions, and diagnostic and therapeutic potentials. Biochim. Biophys. Acta Gen. Subj. 2012, 940–948. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pathogen | EV Effect on Pathogenesis | Source of EVs | Outcome in Recipient Cells | References |
|---|---|---|---|---|
| BACTERIA | ||||
| Haemophilus influenzae, Moraxella catarrhalis, Streptococcus pneumoniae, and Pseudomonas aeruginosa | Dual role | THP-1 | Trigger and/or dampen immune responses against bacterial infection | [40] |
| Escherichia coli | Dual role | THP-1 | Favor or inhibit bacterial replication | [41] |
| Mycobacterium bovis Bovis Bacillus Calmette–Guerin (BCG) | Enhancement | Primary monocyte-derived macrophage (MDM) | Increase bacterial replication and survival | [42] |
| Treponema pallidum | Enhancement | THP-1 | Affect adhesion and permeability of human endothelial cells | [43] |
| Enhancement | THP-1 | Increase evasion of host immune response | [44] | |
| M. tuberculosis (M. tb) | Enhancement | THP-1 | Induce changes in EV protein composition to increased bacterial infection | [45] |
| Mycobacterium tuberculosis, M. bovis BCG, Salmonella typhimurium | Inhibition | THP-1 | Enhance host immune response | [46] |
| M. tuberculosis (M. tb) | Inhibition | THP-1 | Promote inflammation, intercellular communication, and cell migration to contain bacterial infection | [47] |
| Inhibition | THP-1 | Induce type I IFN response against M. tb infection | [48] | |
| Mycobacterium avium sp. paratuberculosis | Inhibition | THP-1 | Enhance host immune response | [49] |
| M. avium | Inhibition | THP-1 | Enhance host immune response | [50] |
| M. bovis BCG | Inhibition | THP-1 | Increase host innate immune response via endosomal TLR-7 activation | [51] |
| Legionella pneumophila | Inhibition | THP-1 or MDM | Stimulate immune response to control bacterial infection | [52] |
| Salmonella enterica serovar Typhimurium | Inhibition | THP-1 | Enhance the release of pro-inflammatory mediators to control bacterial infection | [53] |
| VIRUSES | ||||
| HIV-1 | Enhancement | MDM | Increase viral infection | [54] |
| Enhancement | U937 or MDM | Increase HIV-1 reactivation in latent cellular reservoirs | [55] | |
| Enhancement | THP-1 or alveolar macrophages | Increase chronic immune activation through HIV vmiR88 and vmiR99 | [56] | |
| Enhancement | MDM | Increase viral infection | [57] | |
| Enhancement | THP-1 or MDM | Affect epithelial barrier integrity and mitochondrial activity which contribute to immune dysfunctions against viral infection | [58] | |
| Enhancement | MDM | Promote hyperproliferation of pulmonary arterial smooth muscle cells | [59] | |
| Enhancement | MDM | Induce neuronal apoptosis and dysfunction through cathepsin B | [60] | |
| H1N1 | Enhancement | MDM or THP-1 | Affect innate immune response in macrophages and DC against viral infection | [61] |
| DENV | Inhibition | U937 | Induce activation of endothelial cells and promote endothelial barrier changes to increase proinflammatory response during infection | [62] |
| HBV | Inhibition | THP-1 | Induce anti-HBV activity | [63] |
| HCV | Inhibition | MDM | Induce anti-HCV activity | [64] |
| Inhibition | MDM or THP-1 | Induce anti-HCV activity | [65] | |
| PARASITES | ||||
| Toxoplasma gondii | Inhibition | THP-1 | Enhance host immune response | [46] |
| Trypanosoma cruzi | Enhancement | THP-1 | Inhibit innate immune response | [66] |
| Undefined | THP-1 | Undefined | [67] | |
| FUNGI | ||||
| Fungal cell components: β-glucans | Inhibition | MDM | Activate innate immune system via Dectin-1 receptor | [68] |
| Candida albicans | Inhibition | THP-1 | Enhance host immune response | [69] |
| Source of EVs | Isolation Methods | Characterization Methods | References |
|---|---|---|---|
| BACTERIA | |||
| THP-1 | SEC | EM, FC, and TRPS | [40] |
| THP-1 | Isolation kit | EM and WB | [41] |
| MDM | Isolation kit | EM, NTA, and miRNA content | [42] |
| THP-1 | Isolation kit | EM, NTA, and WB | [43] |
| THP-1 | Isolation kit | EM, NTA, miRNA content, and WB | [44] |
| THP-1 | Isolation kit | EM, NTA, PT, and WB | [45] |
| THP-1 | UC and sucrose density gradient separation | EM, FC, and WB | [46] |
| THP-1 | UC | EM, FC, and PT | [47] |
| THP-1 | UC | PT | [48] |
| THP-1 | UC | EM, FC, and PT | [49] |
| THP-1 | Isolation kit | EM | [50] |
| MDM | UC | EM, NTA, RNA-seq, and WB | [51] |
| THP-1 | UC | EM, NTA, and WB | [52] |
| THP-1 | UC and iodixanol density gradient separation | EM, NTA, PT, and WB | [53] |
| VIRUSES | |||
| MDM | UC and sucrose density gradient separation | EM, LP, and PT | [54] |
| U937 or MDM | UC and iodixanol density gradient separation | AChE activity and FC | [55] |
| THP-1 or primary human alveolar macrophages | Isolation kit | miRNA content and RNA-seq | [56] |
| MDM | UC and iodixanol density gradient separation | AChE activity, miRNA content, and WB | [57] |
| THP-1 or MDM | UC | EM, NTA, and WB | [58] |
| MDM | UC | EM, NTA, and WB | [59] |
| MDM | Isolation kit | NTA and WB | [60] |
| MDM or THP-1 | Isolation kit | NTA and miRNA content | [61] |
| U937 | UC and sucrose density gradient separation | AChE activity, EM, miRNA content, PT, and WB | [62] |
| THP-1 | UC | EM, FC, and WB | [63] |
| THP-1 | Isolation kit | EM and WB | [64] |
| MDM or THP-1 | UC | NTA, EM, FC, and WB | [65] |
| PARASITES | |||
| THP-1 | UC and sucrose density gradient separation | EM, FC, and WB | [46] |
| THP-1 | UC | EM and FC | [66] |
| THP-1 | SEC | EM, NTA, PT, and WB | [67] |
| FUNGI | |||
| MDM | UC | EM, NTA, PT, and WB | [68] |
| THP-1 | UC | DLS, EM, and PT | [69] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arteaga-Blanco, L.A.; Bou-Habib, D.C. The Role of Extracellular Vesicles from Human Macrophages on Host-Pathogen Interaction. Int. J. Mol. Sci. 2021, 22, 10262. https://doi.org/10.3390/ijms221910262
Arteaga-Blanco LA, Bou-Habib DC. The Role of Extracellular Vesicles from Human Macrophages on Host-Pathogen Interaction. International Journal of Molecular Sciences. 2021; 22(19):10262. https://doi.org/10.3390/ijms221910262
Chicago/Turabian StyleArteaga-Blanco, Luis A., and Dumith Chequer Bou-Habib. 2021. "The Role of Extracellular Vesicles from Human Macrophages on Host-Pathogen Interaction" International Journal of Molecular Sciences 22, no. 19: 10262. https://doi.org/10.3390/ijms221910262
APA StyleArteaga-Blanco, L. A., & Bou-Habib, D. C. (2021). The Role of Extracellular Vesicles from Human Macrophages on Host-Pathogen Interaction. International Journal of Molecular Sciences, 22(19), 10262. https://doi.org/10.3390/ijms221910262