
PDF1.5 Enhances Adaptation to Low Nitrogen Levels and Cadmium Stress

 ,
, 
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
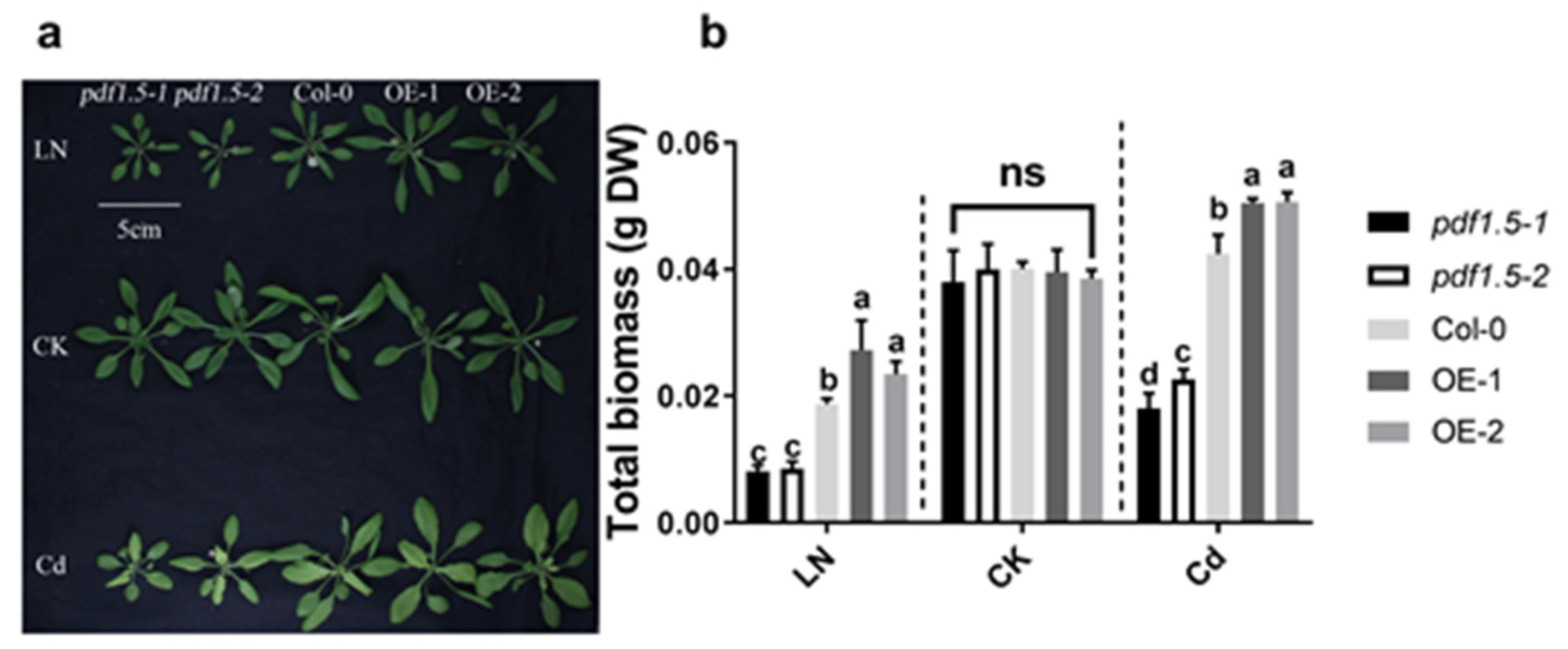
2.1. AtPDF1.5 Enhances Adaptation to LN Levels and Cd Stress
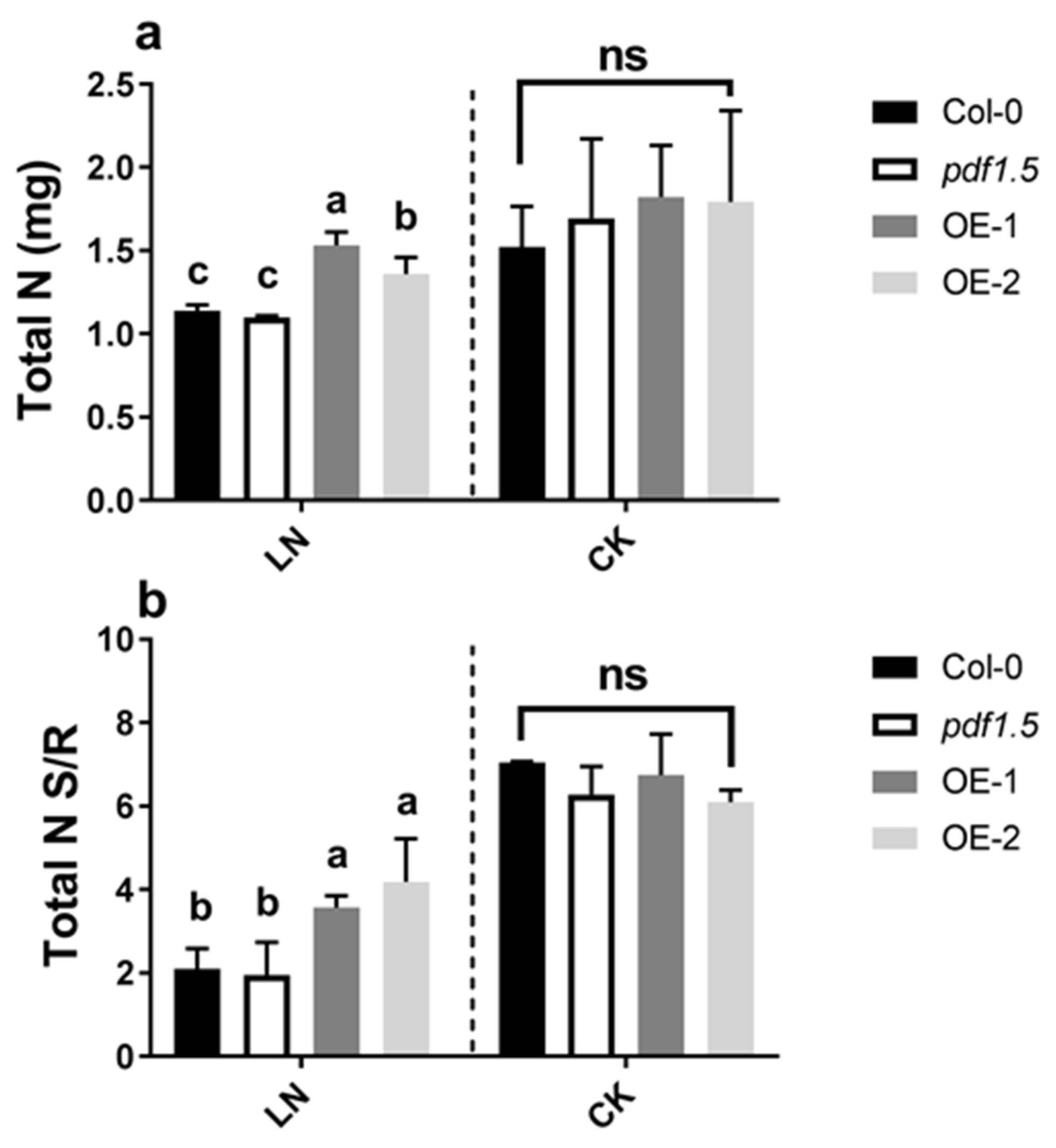
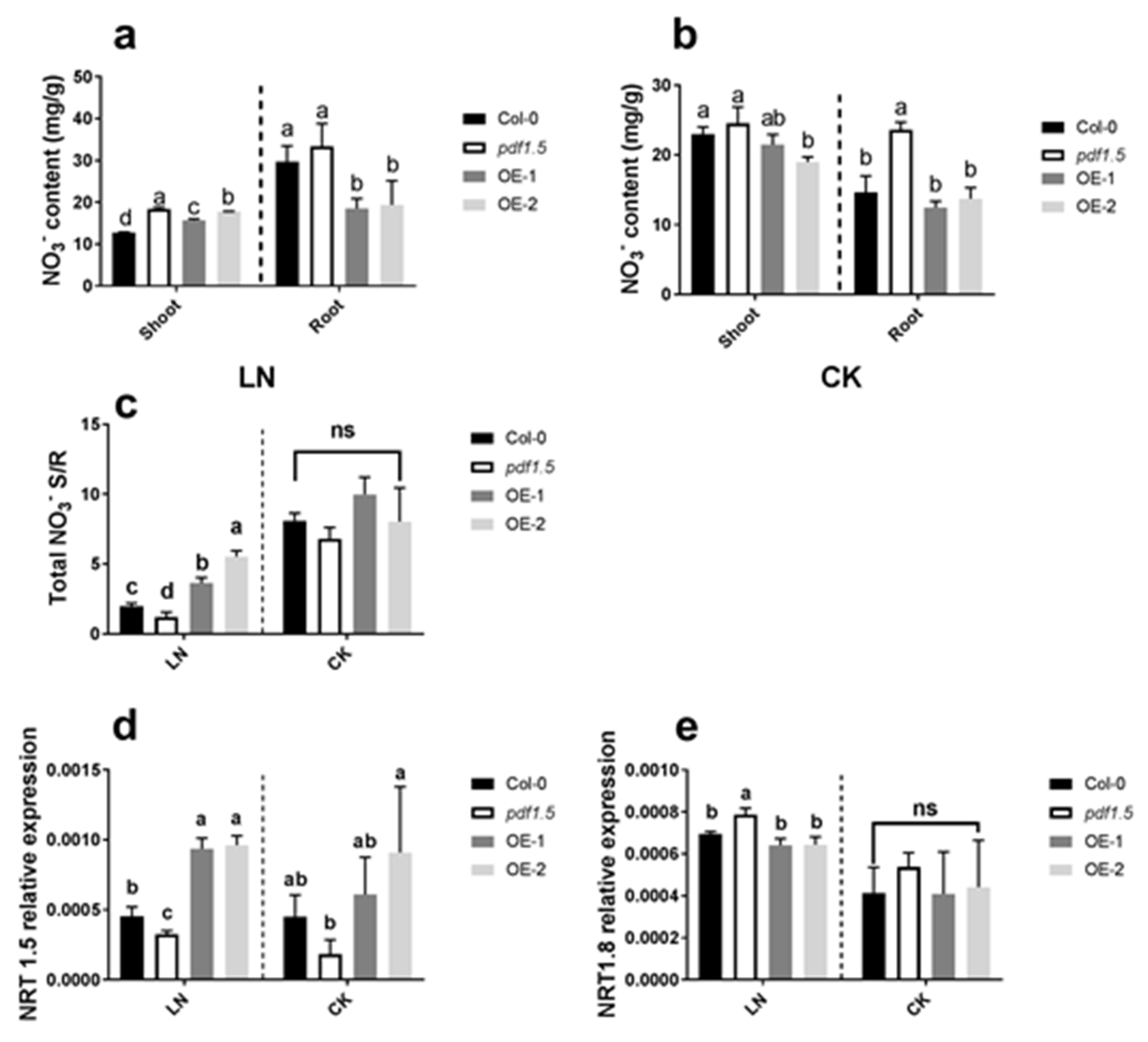
2.2. AtPDF1.5 Influences Nitrogen Uptake and Translocation under LN Conditions
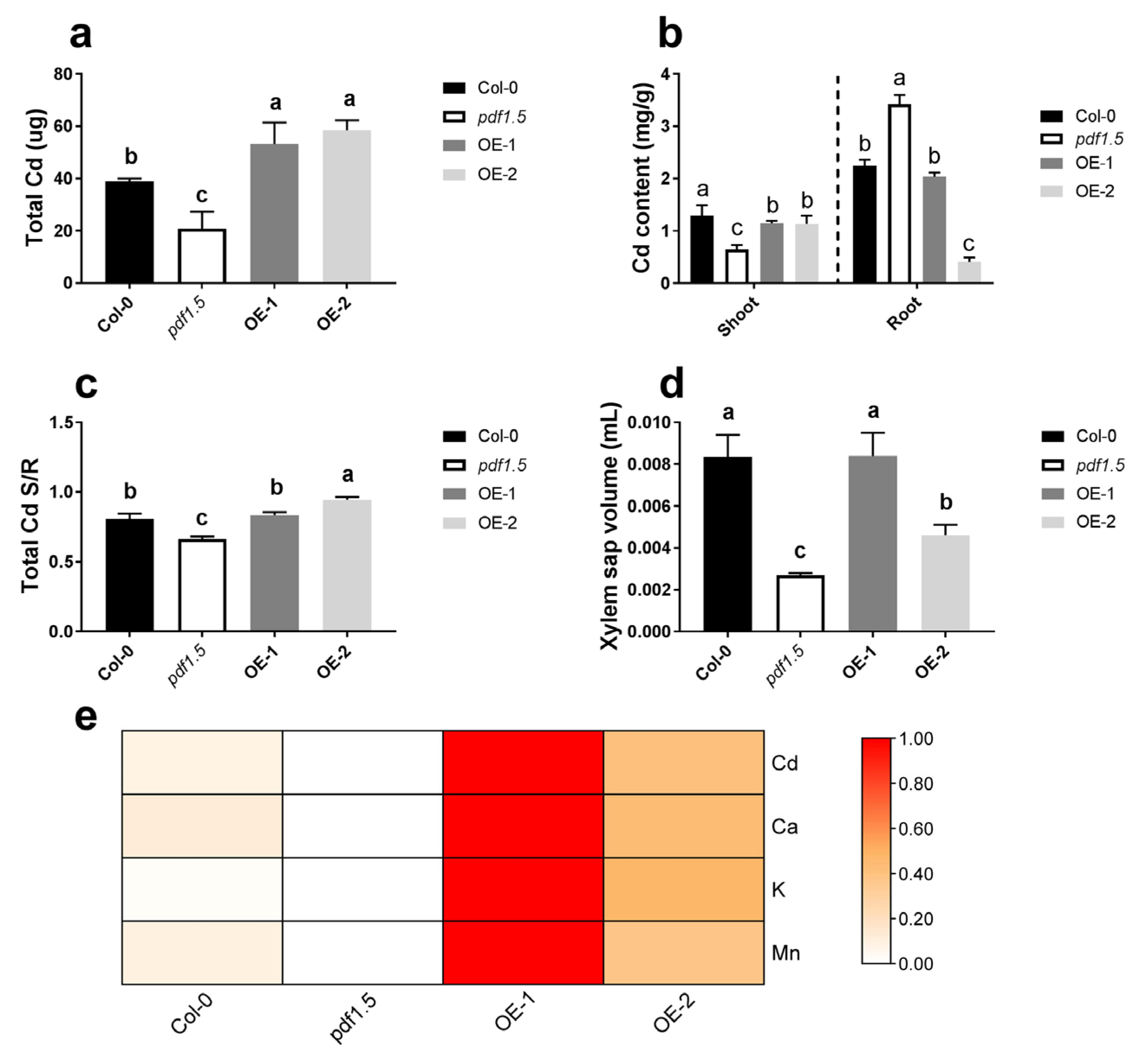
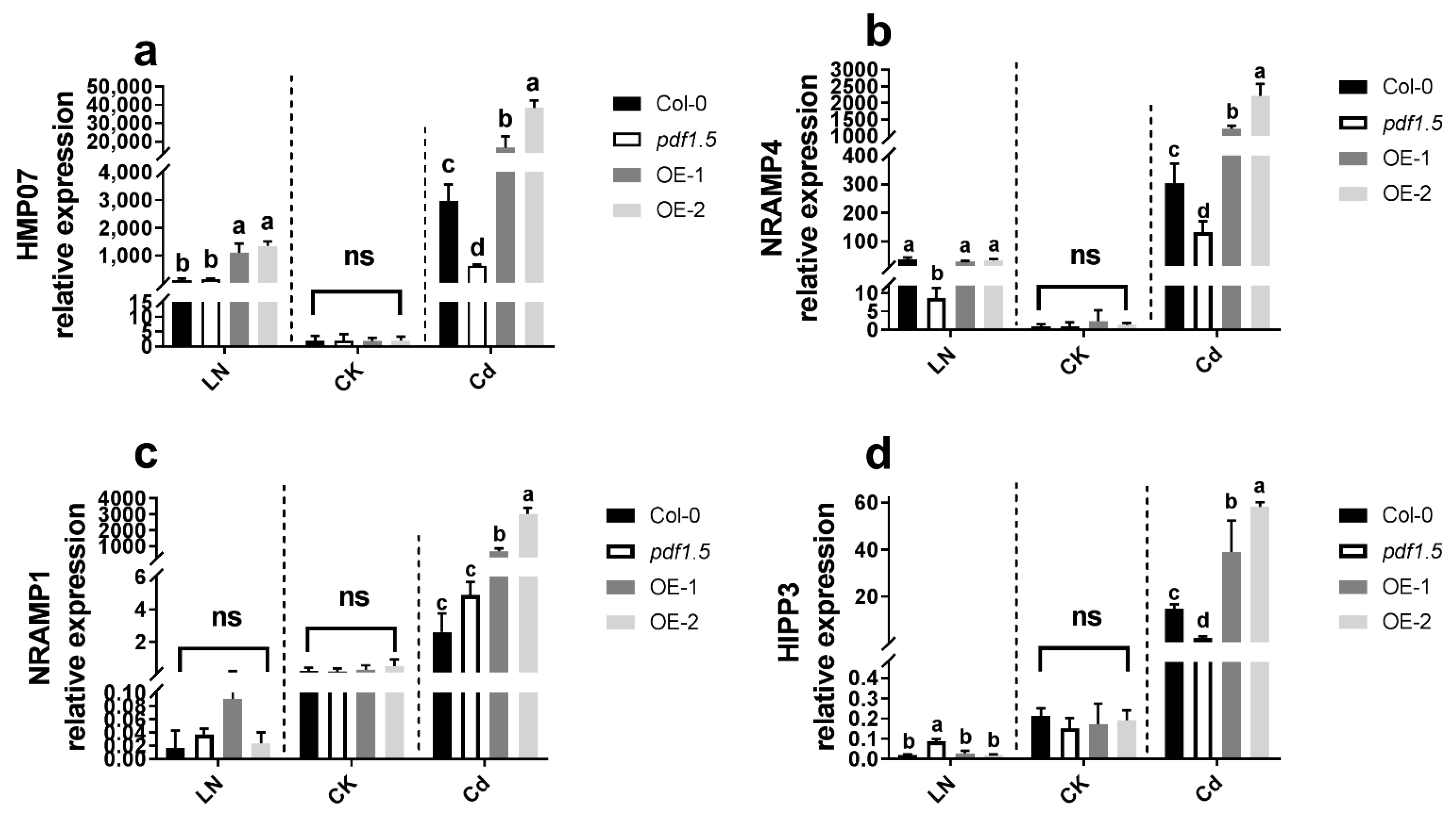
2.3. AtPDF1.5 Enhances Cd Tolerance and Accumulation in A. thaliana
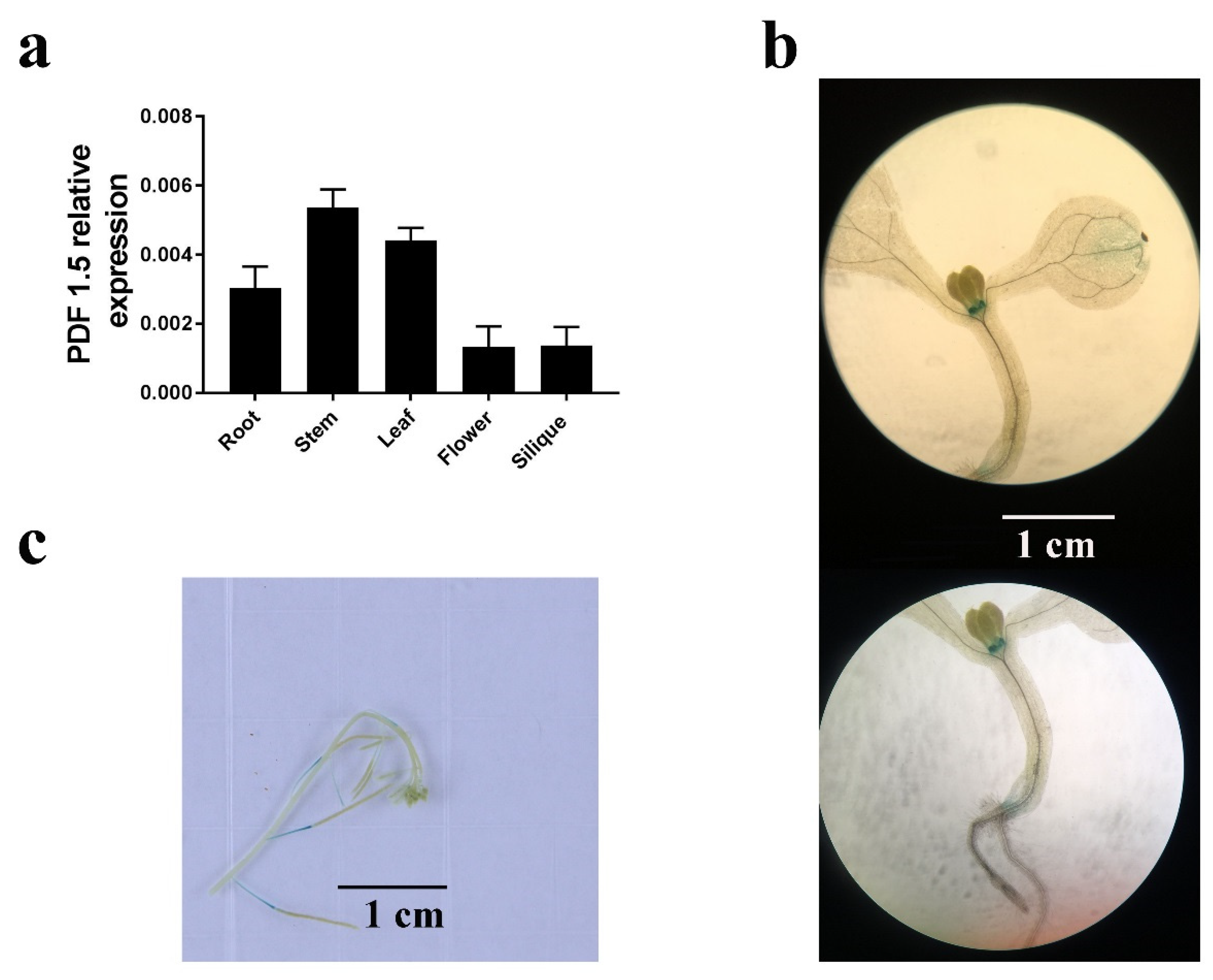
2.4. AtPDF1.5 Expression and Subcellular Localization
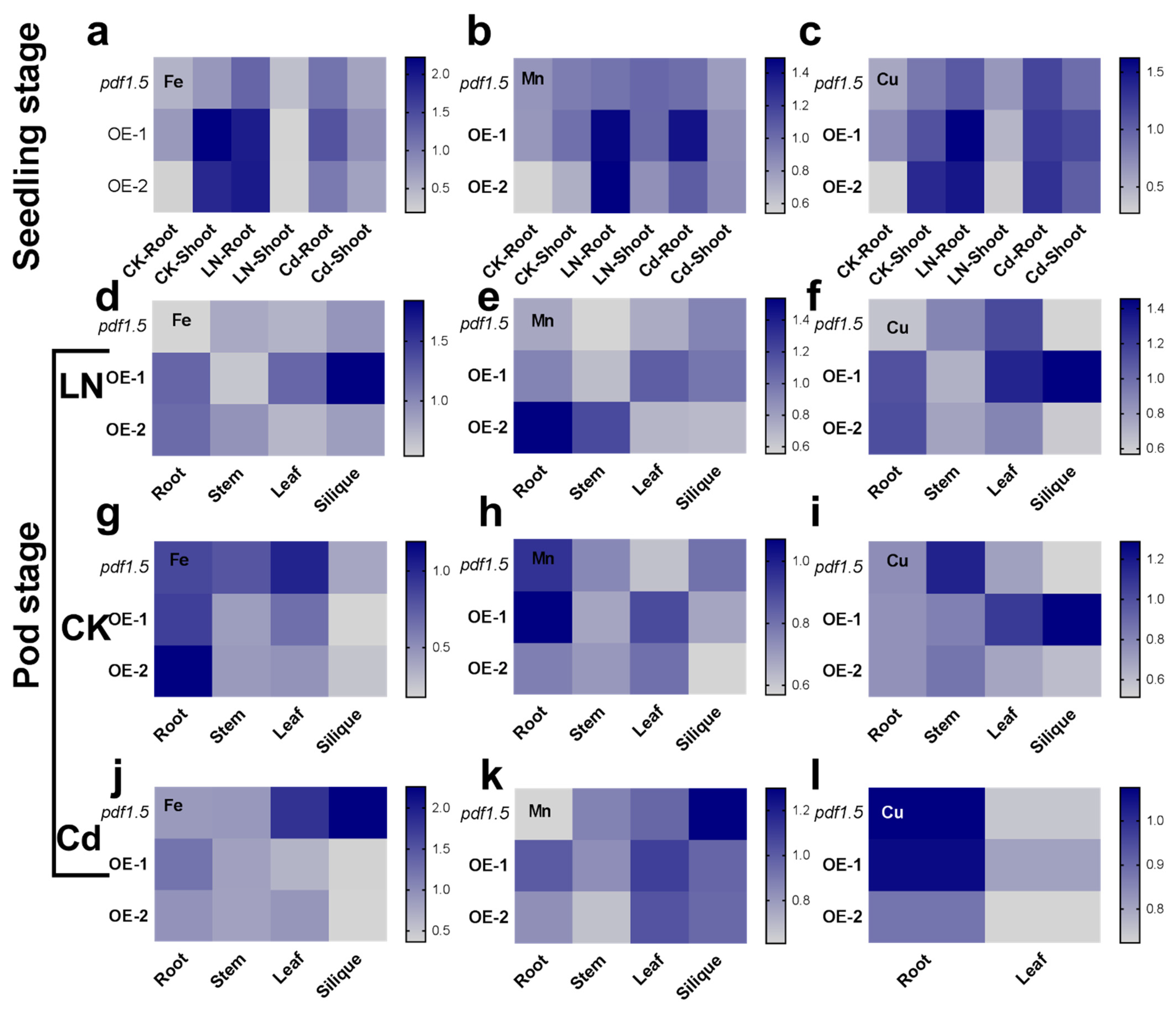
2.5. AtPDF1.5 Is Involved in the Allocation of Multiple Metals in A. thaliana
3. Discussion
3.1. AtPDF1.5 Enhances Adaptation to LN by Regulating AtNRT1.5 and AtNRT1.8 Expression
3.2. AtPDF1.5 Enhances Cd Tolerance by Increasing Cd Transport to Shoots
3.3. AtPDF1.5 Enhances Cd Transport and Affects the Transport of Other Cations
4. Materials and Methods
4.1. Materials and Growth Conditions
4.2. Preparation of DNA Constructs and Plant Transformation
4.3. Expression, β-Glucuronidase (GUS) Histochemical Analyses, and Subcellular Localization
4.4. Nitrogen Concentration Assay
4.5. Xylem Sap Collection and NO3− Concentration Assay
4.6. Cation Concentration Assay
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
References
- Mendez, E.; MORENO, A.; COLILLA, F.; PELAEZ, F.; LIMAS, G.G.; MENDEZ, R.; SORIANO, F.; SALINAS, M.; de HARO, C. Primary structure and inhibition of protein synthesis in eukaryotic cell-free system of a novel thionin, γ-hordothionin, from barley endosperm. Eur. J. Biochem. 1990, 194, 533–539. [Google Scholar] [CrossRef]
- Larsen, B.; Xu, D.; Halkier, B.A.; Nour-Eldin, H.H. Advances in methods for identification and characterization of plant transporter function. J. Exp. Bot. 2017, 68, 4045–4056. [Google Scholar] [CrossRef]
- Wang, J.; Dun, X.; Shi, J.; Wang, X.; Liu, G.; Wang, H. Genetic Dissection of Root Morphological Traits Related to Nitrogen Use Efficiency in Brassica napus L. under Two Contrasting Nitrogen Conditions. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ANDREWS, M. The partitioning of nitrate assimilation between root and shoot of higher plants. Plant. Cell Environ. 1986, 9, 511–519. [Google Scholar] [CrossRef]
- Han, Y.-L.; Song, H.-X.; Liao, Q.; Yu, Y.; Jian, S.-F.; Lepo, J.E.; Liu, Q.; Rong, X.-M.; Tian, C.; Zeng, J.; et al. Nitrogen use efficiency is mediated by vacuolar nitrate sequestration capacity in roots of Brassica napus. Plant Physiol. 2016, 170, 1684–1698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, F.; Yu, G.; Liu, P. Transporter-mediated subcellular distribution in the metabolism and signaling of jasmonates. Front. Plant Sci. 2019, 10, 1–8. [Google Scholar] [CrossRef]
- Tang, Z.; Fan, X.; Li, Q.; Feng, H.; Miller, A.J.; Shen, Q.; Xu, G. Knockdown of a rice stelar nitrate transporter alters long-distance translocation but not root influx. Plant Physiol. 2012, 160, 2052–2063. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.H.; Kuo, H.F.; Canivenc, G.; Lin, C.S.; Lepetit, M.; Hsu, P.K.; Tillard, P.; Lin, H.G.; Wang, Y.Y.; Tsai, C.B.; et al. Mutation of the Arabidopsis NRT1.5 nitrate transporter causes defective root-to-shoot nitrate transport. Plant Cell 2008, 20, 2514–2528. [Google Scholar] [CrossRef] [Green Version]
- Li, J.-Y.; Fu, Y.-L.; Pike, S.M.; Bao, J.; Tian, W.; Zhang, Y.; Chen, C.-Z.; Zhang, Y.; Li, H.-M.; Huang, J.; et al. The Arabidopsis Nitrate Transporter NRT1.8 Functions in Nitrate Removal from the Xylem Sap and Mediates Cadmium Tolerance. Plant Cell Online 2010, 22, 1633–1646. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Gong, Z.; Guo, Y.; Gong, J. Studies on plant responses to environmental change in China: The past and the future. Sci. Sin. 2019, 49, 1457–1478. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.W.; Meng, J.J.; Xing, J.Y.; Yang, S.; Guo, F.; Li, X.G.; Wan, S.B. The K+/H+ antiporter AhNHX1 improved tobacco tolerance to NaCl stress by enhancing K+ retention. J. Plant Biol. 2017, 60, 259–267. [Google Scholar] [CrossRef]
- Luo, Y.; Rimmer, D.L. Zinc-copper interaction affecting plant growth on a metal-contaminated soil. Environ. Pollut. 1995, 88, 79–83. [Google Scholar] [CrossRef]
- Lepp, N.W. Effect of Heavy Metal Pollution on Plants: Effects of Trace Metals on Plant Function; Springer Science & Business Media: Berlin, Germany, 2012; ISBN 9401173397. [Google Scholar]
- Lu, L.; Tian, S.; Liao, H.; Zhang, J.; Yang, X.; Labavitch, J.M.; Chen, W. Analysis of metal element distributions in rice (Oryza sativa L.) seeds and relocation during germination based on X-ray fluorescence imaging of Zn, Fe, K, Ca, and Mn. PLoS ONE 2013, 8, e57360. [Google Scholar] [CrossRef]
- Ramos, O.M.; Magnitskiy, S.; Darghan, A. Effect of nitrogen and potassium on plant height and stem diameter of Jatropha curcas L. in Colombian tropical dry forest. Agron. Colomb. 2019, 37, 203–212. [Google Scholar] [CrossRef] [Green Version]
- Huber, D.M.; Arny, D.C. Interactions of potassium with plant disease. Potassium Agric. 1985, 467–488. [Google Scholar] [CrossRef]
- Riveras, E.; Alvarez, J.M.; Vidal, E.A.; Oses, C.; Vega, A.; Gutiérrez, R.A. The calcium ion is a second messenger in the nitrate signaling pathway of Arabidopsis. Plant Physiol. 2015, 169, 1397–1404. [Google Scholar] [CrossRef] [Green Version]
- McHargue, J.S. The role of manganese in plants. J. Am. Chem. Soc. 1922, 44, 1592–1598. [Google Scholar] [CrossRef] [Green Version]
- ul Hassan, Z.; Ali, S.; Rizwan, M.; Ali, Q.; Haider, M.Z.; Adrees, M.; Hussain, A. Role of Iron in Alleviating Heavy Metal Stress. In Essential Plant Nutrients; Springer: Berlin, Germany, 2017; pp. 335–350. [Google Scholar]
- Tsai, H.H.; Schmidt, W. Mobilization of iron by plant-borne coumarins. Trends Plant Sci. 2017, 22, 538–548. [Google Scholar] [CrossRef] [PubMed]
- Nisha Kevat, N.; Sharma, P.K. Photosynthesis in copper mediated plant is affected due to oxidative damage caused by reactive oxygen species (ros) generation. Int. J. Recent Sci. Res. 2016, 7, 14009–14015. [Google Scholar]
- Makowska, K.; Oleszczuk, S.; Zimny, J. The effect of copper on plant regeneration in barley microspore culture. Czech J. Genet. Plant Breed. 2017, 53, 17–22. [Google Scholar] [CrossRef] [Green Version]
- Dupont, C.L.; Butcher, A.; Valas, R.E.; Bourne, P.E.; Caetano-Anollés, G. History of biological metal utilization inferred through phylogenomic analysis of protein structures. Proc. Natl. Acad. Sci. USA 2010, 107, 10567–10572. [Google Scholar] [CrossRef] [Green Version]
- Huffman, D.L.; O’Halloran, T.V. Function, Structure, and Mechanism of Intracellular Copper Trafficking Proteins. Annu. Rev. Biochem. 2001, 70, 677–701. [Google Scholar] [CrossRef]
- Zlobin, I.E.; Kholodova, V.P.; Rakhmankulova, Z.F.; Kuznetsov, V.V. Brassica napus responses to short-term excessive copper treatment with decrease of photosynthetic pigments, differential expression of heavy metal homeostasis genes including activation of gene NRAMP4 involved in photosystem II stabilization. Photosynth. Res. 2015, 125, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Oomen, R.J.F.J.; Wu, J.; Lelièvre, F.; Blanchet, S.; Richaud, P.; Barbier-Brygoo, H.; Aarts, M.G.M.; Thomine, S. Functional characterization of NRAMP3 and NRAMP4 from the metal hyperaccumulator Thlaspi caerulescens. New Phytol. 2009, 181, 637–650. [Google Scholar] [CrossRef] [PubMed]
- Agorio, A.; Giraudat, J.; Bianchi, M.W.; Marion, J.; Espagne, C.; Castaings, L.; Lelièvre, F.; Curie, C.; Thomine, S.; Merlot, S. Phosphatidylinositol 3-phosphate-binding protein AtPH1 controls the localization of the metal transporter NRAMP1 in Arabidopsis. Proc. Natl. Acad. Sci. USA 2017, 114, E3354–E3363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.M.; Wang, Y.M.; Yang, H.L.; Zeng, Q.Y.; Liu, Y.J. NRAMP1 promotes iron uptake at the late stage of iron deficiency in poplars. Tree Physiol. 2019, 39, 1235–1250. [Google Scholar] [CrossRef]
- De Abreu-Neto, J.B.; Turchetto-Zolet, A.C.; De Oliveira, L.F.V.; Bodanese Zanettini, M.H.; Margis-Pinheiro, M. Heavy metal-associated isoprenylated plant protein (HIPP): Characterization of a family of proteins exclusive to plants. FEBS J. 2013, 280, 1604–1616. [Google Scholar] [CrossRef] [PubMed]
- Sato, M.; Mitra, R.M.; Coller, J.; Wang, D.; Spivey, N.W.; Dewdney, J.; Denoux, C.; Glazebrook, J.; Katagiri, F. A high-performance, small-scale microarray for expression profiling of many samples in Arabidopsis-pathogen studies. Plant J. 2007, 49, 565–577. [Google Scholar] [CrossRef]
- Zschiesche, W.; Barth, O.; Daniel, K.; Böhme, S.; Rausche, J.; Humbeck, K. The zinc-binding nuclear protein HIPP3 acts as an upstream regulator of the salicylate-dependent plant immunity pathway and of flowering time in Arabidopsis thaliana. New Phytol. 2015, 207, 1084–1096. [Google Scholar] [CrossRef]
- Shahzad, Z.; Ranwez, V.; Fizames, C.; Marquès, L.; Le Martret, B.; Alassimone, J.; Godé, C.; Lacombe, E.; Castillo, T.; Saumitou-Laprade, P.; et al. Plant Defensin type 1 (PDF1): Protein promiscuity and expression variation within the Arabidopsis genus shed light on zinc tolerance acquisition in Arabidopsis halleri. New Phytol. 2013, 200, 820–833. [Google Scholar] [CrossRef]
- Zai, W.S.; Ye, S.G.; Miao, L.X.; Xiong, Z.L.; Zhang, H.-L.; Ma, Y.-R.; Li, Y.-L.; Chen, Y.-B. Genome-wide identification and expression analysis of defensin genes in tomato (Solanum lycopersicum). J. Agric. Biotechnol. 2015, 23, 1588–1596. [Google Scholar] [CrossRef]
- Wang, C.; Yao, J.; Du, X.; Zhang, Y.; Sun, Y.; Rollins, J.A.; Mou, Z. The Arabidopsis Mediator Complex Subunit16 Is a Key Component of Basal Resistance against the Necrotrophic Fungal Pathogen Sclerotinia sclerotiorum. Plant Physiol. 2015, 169, 856–872. [Google Scholar] [CrossRef] [Green Version]
- Koornneef, A.; Rindermann, K.; Gatz, C.; Pieterse, C.M.J. Histone modifications do not play a major role in salicylate-mediated suppression of jasmonate-induced PDF1.2 gene expression. Commun. Integr. Biol. 2008, 1, 143–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomma, B.P.H.J.; Broekaert, W.F. Tissue-specific expression of plant defensin genes PDF2.1 and PDF2.2 in Arabidopsis thaliana. Plant Physiol. Biochem. 1998, 36, 533–537. [Google Scholar] [CrossRef]
- Yao, J.; Luo, J.; Xiao, Y.; Zhang, Z. The plant defensin gene AtPDF2.1 mediates ammonium metabolism by regulating glutamine synthetase activity in Arabidopsis thaliana. BMC Plant Biol. 2019, 19, 557. [Google Scholar] [CrossRef]
- Luo, J.S.; Gu, T.; Yang, Y.; Zhang, Z. A non-secreted plant defensin AtPDF2.6 conferred cadmium tolerance via its chelation in Arabidopsis. Plant Mol. Biol. 2019, 100, 561–569. [Google Scholar] [CrossRef]
- Luo, J.S.; Yang, Y.; Gu, T.; Wu, Z.; Zhang, Z. The Arabidopsis defensin gene AtPDF2.5 mediates cadmium tolerance and accumulation. Plant Cell Environ. 2019, 42, 2681–2695. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Luo, J.; Han, Y.; Hua, Y.; Guan, C.; Zhang, Z. Low Nitrogen Enhances Nitrogen Use Efficiency by Triggering NO3− Uptake and Its Long-Distance Translocation. J. Agric. Food Chem. 2019, 67, 6736–6747. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.I.R.; Iqbal, N.; Masood, A.; Khan, N.A. Variation in Salt Tolerance of Wheat Cultivars: Role of Glycinebetaine and Ethylene. Pedosphere 2012, 22, 746–754. [Google Scholar] [CrossRef]
- Brown, R.L.; Kazan, K.; McGrath, K.C.; Maclean, D.J.; Manners, J.M. A role for the GCC-box in jasmonate-mediated activation of the PDF1.2 gene of Arabidopsis. Plant Physiol. 2003, 132, 1020–1032. [Google Scholar] [CrossRef] [Green Version]
- Larsen, P.B.; Cancel, J.D. A recessive mutation in the RUB1-conjugating enzyme, RCE1, reveals a requirement for RUB modification for control of ethylene biosynthesis and proper induction of basic chitinase and PDF1.2 in Arabidopsis. Plant J. 2004, 38, 626–638. [Google Scholar] [CrossRef]
- Luo, J.S.; Huang, J.; Zeng, D.L.; Peng, J.S.; Zhang, G.B.; Ma, H.L.; Guan, Y.; Yi, H.Y.; Fu, Y.L.; Han, B.; et al. A defensin-like protein drives cadmium efflux and allocation in rice. Nat. Commun. 2018, 9, 645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tabata, R.; Sumida, K.; Yoshii, T.; Ohyama, K.; Shinohara, H.; Matsubayashi, Y. Perception of root-derived peptides by shoot LRR-RKs mediates systemic N-demand signaling. Science 2014, 346, 343–346. [Google Scholar] [CrossRef]
- Ohkubo, Y.; Tanaka, M.; Tabata, R.; Ogawa-Ohnishi, M.; Matsubayashi, Y. Shoot-to-root mobile polypeptides involved in systemic regulation of nitrogen acquisition. Nat. Plants 2017, 3, 17029. [Google Scholar] [CrossRef]
- Takahashi, F.; Suzuki, T.; Osakabe, Y.; Betsuyaku, S.; Kondo, Y.; Dohmae, N.; Fukuda, H.; Yamaguchi-Shinozaki, K.; Shinozaki, K. A small peptide modulates stomatal control via abscisic acid in long-distance signaling. Nature 2018, 556, 235–238. [Google Scholar] [CrossRef]
- Chen, C.-Z.; Lv, X.-F.; Li, J.-Y.; Yi, H.-Y.; Gong, J.-M. Arabidopsis NRT1.5 Is Another Essential Component in the Regulation of Nitrate Reallocation and Stress Tolerance. Plant Physiol. 2012, 159, 1582–1590. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.B.; Yi, H.Y.; Gong, J.M. The Arabidopsis Ethylene/Jasmonic acid-NRT signaling module coordinates nitrate reallocation and the trade-off between growth and environmental adaptation. Plant Cell 2014, 26, 3984–3998. [Google Scholar] [CrossRef] [Green Version]
- Gojon, A. Nitrogen nutrition in plants: Rapid progress and new challenges. J. Exp. Bot. 2017, 68, 2457–2462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masclaux-Daubresse, C.; Daniel-Vedele, F.; Dechorgnat, J.; Chardon, F.; Gaufichon, L.; Suzuki, A. Nitrogen uptake, assimilation and remobilization in plants: Challenges for sustainable and productive agriculture. Ann. Bot. 2010, 105, 1141–1157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eren, E.; Argüello, J.M. Arabidopsis HMA2, a divalent heavy metal-transporting P(IB)-type ATPase, is involved in cytoplasmic Zn2+ homeostasis. Plant Physiol. 2004, 136, 3712–3723. [Google Scholar] [CrossRef] [Green Version]
- Korenkov, V.; Hirschi, K.; Crutchfield, J.D.; Wagner, G.J. Enhancing tonoplast Cd/H antiport activity increases Cd, Zn, and Mn tolerance, and impacts root/shoot Cd partitioning in Nicotiana tabacum L. Planta 2007, 226, 1379–1387. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Wen, Y.; Yang, W.; Zhang, X.; Xia, M.; Zhou, N.; Xiong, Y.; Zhou, Z. The mechanism transformation of ramie biochar’s cadmium adsorption by aging. Bioresour. Technol. 2021, 330, 124947. [Google Scholar] [CrossRef]
- Gravot, A.; Lieutaud, A.; Verret, F.; Auroy, P.; Vavasseur, A.; Richaud, P. AtHMA3, a plant P1B-ATPase, functions as a Cd/Pb transporter in yeast. FEBS Lett. 2004, 561, 22–28. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Song, H.; Guan, C.; Zhang, Z. Boron alleviates cadmium toxicity in Brassica napus by promoting the chelation of cadmium onto the root cell wall components. Sci. Total Environ. 2020, 728, 138833. [Google Scholar] [CrossRef]
- Page, V.; Feller, U. Heavy metals in crop plants: Transport and redistribution processes on the whole plant level. Agronomy 2015, 5, 447–463. [Google Scholar] [CrossRef] [Green Version]
- Stotz, H.U.; Thomson, J.; Wang, Y. Plant defensins: Defense, development and application. Plant Signal. Behav. 2009, 4, 1010–1012. [Google Scholar] [CrossRef] [Green Version]
- Penninckx, I.A.M.A.; Eggermont, K.; Terras, F.R.G.; Thomma, B.P.H.J.; De Samblanx, G.W.; Buchala, A.; Metraux, J.-P.; Manners, J.M.; Broekaert, W.F. Pathogen-Induced Systemic Activation of a Plant Defensin Gene in Arabidopsis Follows a Salicylic Acid-Independent Pathway. Plant Cell 2007, 8, 2309. [Google Scholar] [CrossRef] [Green Version]
- da Silva Conceição, A.; Broekaert, W.F. Plant defensins. Pathog. Proteins Plants 1999, 247–260. [Google Scholar] [CrossRef]
- Nguyen, N.N.T.; Ranwez, V.; Vile, D.; Soulié, M.-C.; Dellagi, A.; Expert, D.; Gosti, F. Evolutionary tinkering of the expression of PDF1s suggests their joint effect on zinc tolerance and the response to pathogen attack. Front. Plant Sci. 2014, 5, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Y.; Liu, Q.; Gu, J.; Gong, J.; Guan, C.; Lepo, J.E.; Rong, X.; Song, H.; Zhang, Z. V-ATPase and V-PPase at the Tonoplast Affect NO3− Content in Brassica napus by Controlling Distribution of NO3− between the Cytoplasm and Vacuole. J. Plant Growth Regul. 2015, 34, 22–34. [Google Scholar] [CrossRef]
- Gong, J.-M.; Lee, D.A.; Schroeder, J.I. Long-distance root-to-shoot transport of phytochelatins and cadmium in Arabidopsis. Proc. Natl. Acad. Sci. USA 2003, 100, 10118–10123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braver, S.L.; MacKinnon, D.P.; Page, M. Levine’s Guide to SPSS for Analysis of Variance; Psychology Press: New York, NY, USA, 2003; ISBN 113567440X. [Google Scholar]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Z.; Liu, D.; Yue, N.; Song, H.; Luo, J.; Zhang, Z. PDF1.5 Enhances Adaptation to Low Nitrogen Levels and Cadmium Stress. Int. J. Mol. Sci. 2021, 22, 10455. https://doi.org/10.3390/ijms221910455
Wu Z, Liu D, Yue N, Song H, Luo J, Zhang Z. PDF1.5 Enhances Adaptation to Low Nitrogen Levels and Cadmium Stress. International Journal of Molecular Sciences. 2021; 22(19):10455. https://doi.org/10.3390/ijms221910455
Chicago/Turabian StyleWu, Zhimin, Dong Liu, Ningyan Yue, Haixing Song, Jinsong Luo, and Zhenhua Zhang. 2021. "PDF1.5 Enhances Adaptation to Low Nitrogen Levels and Cadmium Stress" International Journal of Molecular Sciences 22, no. 19: 10455. https://doi.org/10.3390/ijms221910455
APA StyleWu, Z., Liu, D., Yue, N., Song, H., Luo, J., & Zhang, Z. (2021). PDF1.5 Enhances Adaptation to Low Nitrogen Levels and Cadmium Stress. International Journal of Molecular Sciences, 22(19), 10455. https://doi.org/10.3390/ijms221910455