
Human Pluripotent Stem Cell-Derived Hepatocyte-Like Cells and Organoids for Liver Disease and Therapy

 , and
, and
Abstract
:1. Introduction
2. Development of Hepatic Cells from hPSCs
2.1. Generation of Functional Hepatocyte-Like Cells (HLCs) from hPSCs
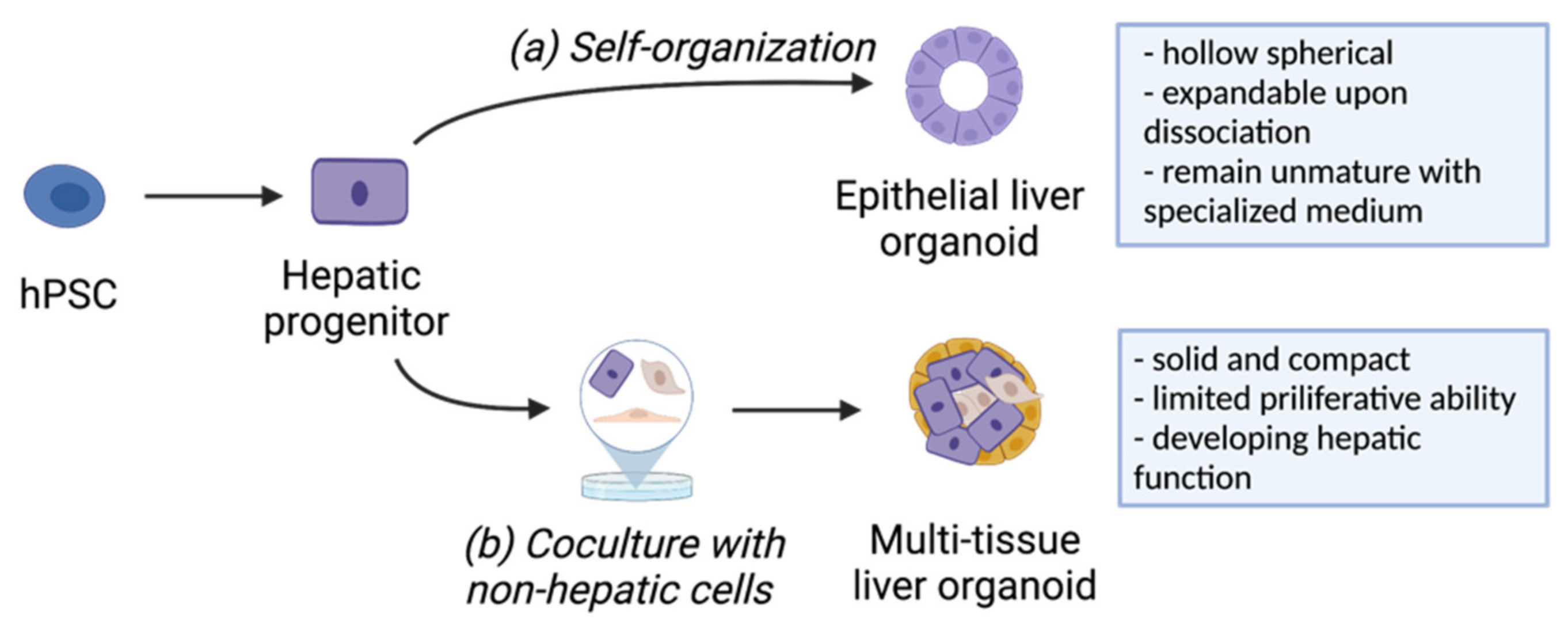
2.2. Development of hPSC-Derived Liver Organoids
2.2.1. Generation of Liver Organoids from hPSCs
2.2.2. Bioengineering Solutions to Generate Robust and Functional Liver Organoids
3. Applications of hPSC-Derived Liver Disease Models
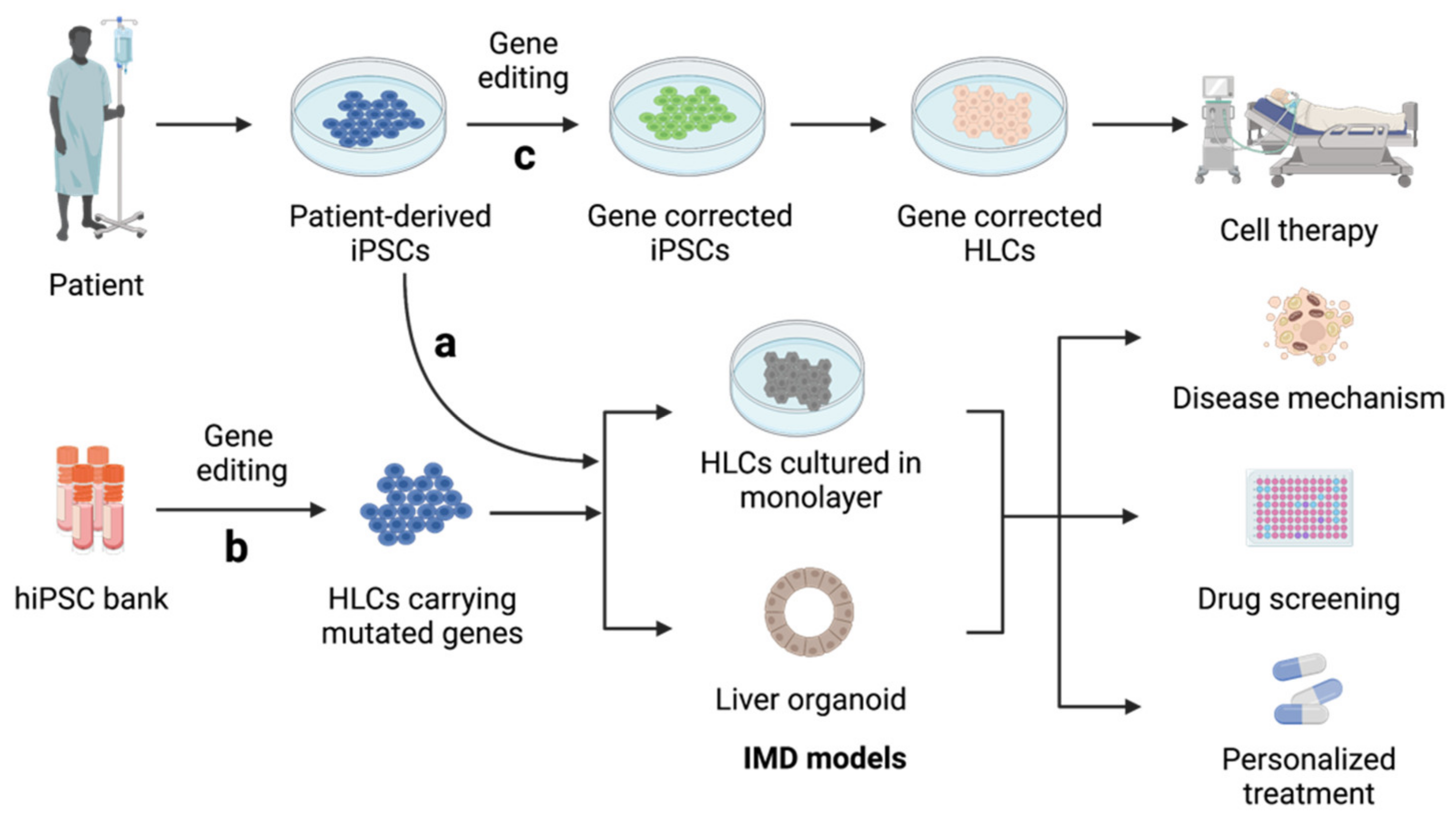
3.1. Inherited Metabolic Disorders of The Liver (IMDs)
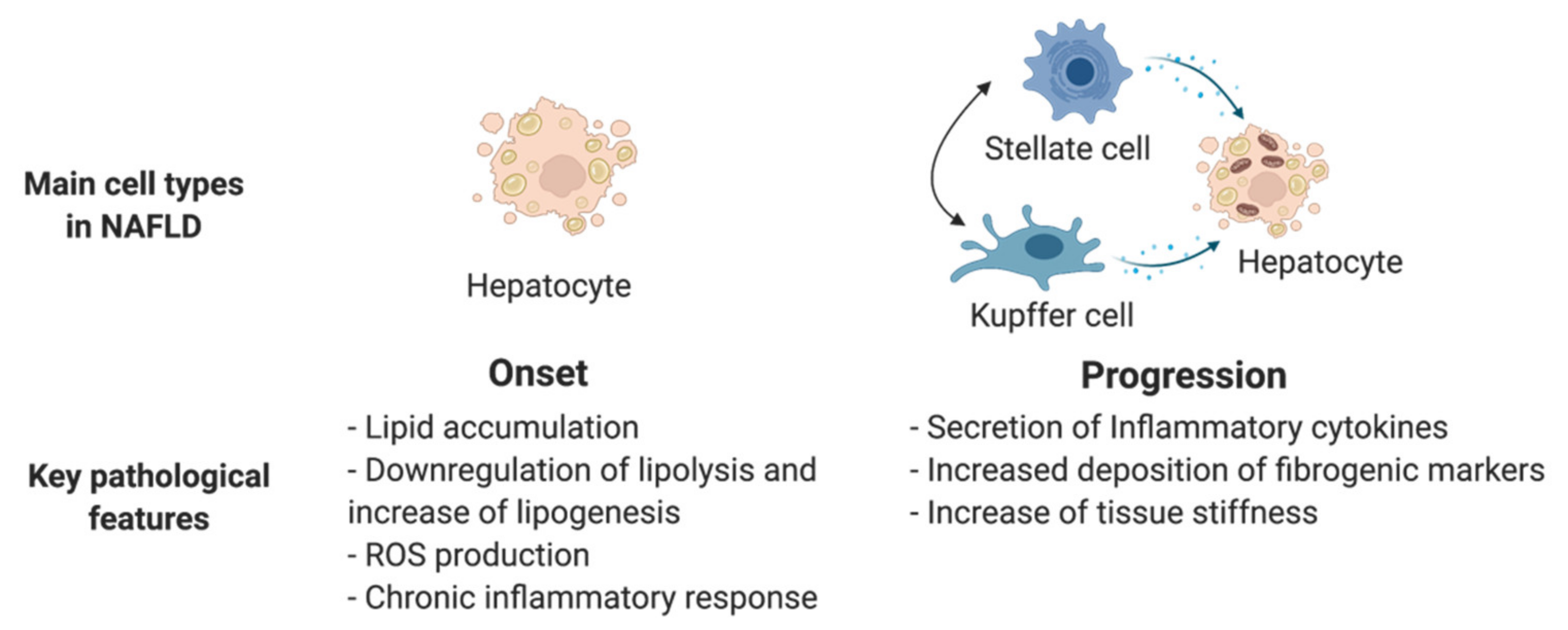
3.2. Non-Alcoholic Fatty Liver Disease (NAFLD)
3.3. Hepatitis B (HBV) and Hepatitis C (HCV) Infection
3.4. Hepatocellular Carcinoma (HCC)
4. Applications of hPSC-Derived Hepatic Lineages in Pharmaceutical Discovery
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Si-Tayeb, K.; Lemaigre, F.P.; Duncan, S.A. Organogenesis and Development of the Liver. Dev. Cell 2010, 18, 175–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stanger, B.Z. Cellular Homeostasis and Repair in the Mammalian Liver. Annu. Rev. Physiol. 2015, 77, 179–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palakkan, A.A.; Hay, D.C.; PR, A.K.; TV, K.; Ross, J.A. Liver Tissue Engineering and Cell Sources: Issues and Challenges. Liver Int. 2013, 33, 666–676. [Google Scholar] [CrossRef] [PubMed]
- Halevy, T.; Urbach, A. Comparing ESC and IPSC—Based Models for Human Genetic Disorders. J. Clin. Med. 2014, 3, 1146–1162. [Google Scholar] [CrossRef] [Green Version]
- Messina, A.; Luce, E.; Hussein, M.; Dubart-Kupperschmitt, A. Pluripotent-Stem-Cell-Derived Hepatic Cells: Hepatocytes and Organoids for Liver Therapy and Regeneration. Cells 2020, 9, 420. [Google Scholar] [CrossRef] [Green Version]
- Zhou, W.; Nelson, E.D.; Rmilah, A.A.A.; Amiot, B.P.; Nyberg, S.L. Stem Cell-Related Studies and Stem Cell-Based Therapies in Liver Diseases. Cell Transpl. 2019, 28, 1116–1122. [Google Scholar] [CrossRef] [Green Version]
- Si-Tayeb, K.; Noto, F.K.; Nagaoka, M.; Li, J.; Battle, M.A.; Duris, C.; North, P.E.; Dalton, S.; Duncan, S.A. Highly Efficient Generation of Human Hepatocyte–like Cells from Induced Pluripotent Stem Cells. Hepatology 2010, 51, 297–305. [Google Scholar] [CrossRef] [Green Version]
- McLean, A.B.; D’Amour, K.A.; Jones, K.L.; Krishnamoorthy, M.; Kulik, M.J.; Reynolds, D.M.; Sheppard, A.M.; Liu, H.; Xu, Y.; Baetge, E.E.; et al. Activin A Efficiently Specifies Definitive Endoderm from Human Embryonic Stem Cells Only When Phosphatidylinositol 3-Kinase Signaling Is Suppressed. Stem Cells 2007, 25, 29–38. [Google Scholar] [CrossRef] [Green Version]
- Hannan, N.R.F.; Segeritz, C.-P.; Touboul, T.; Vallier, L. Production of Hepatocyte-like Cells from Human Pluripotent Stem Cells. Nat. Protoc. 2013, 8, 430–437. [Google Scholar] [CrossRef] [Green Version]
- Touboul, T.; Chen, S.; To, C.C.; Mora-Castilla, S.; Sabatini, K.; Tukey, R.H.; Laurent, L.C. Stage-Specific Regulation of the WNT/β-Catenin Pathway Enhances Differentiation of HESCs into Hepatocytes. J. Hepatol. 2016, 64, 1315–1326. [Google Scholar] [CrossRef] [Green Version]
- Hengstler, J.G.; Brulport, M.; Schormann, W.; Bauer, A.; Hermes, M.; Nussler, A.K.; Fandrich, F.; Ruhnke, M.; Ungefroren, H.; Griffin, L.; et al. Generation of Human Hepatocytes by Stem Cell Technology: Definition of the Hepatocyte. Expert Opin. Drug Met. 2005, 1, 61–74. [Google Scholar] [CrossRef]
- Kim, D.; Ryu, J.; Son, M.; Oh, J.; Chung, K.; Lee, S.; Lee, J.; Ahn, J.; Min, J.; Ahn, J.; et al. A Liver-Specific Gene Expression Panel Predicts the Differentiation Status of in Vitro Hepatocyte Models. Hepatology 2017, 66, 1662–1674. [Google Scholar] [CrossRef] [Green Version]
- Ong, J.; Serra, M.P.; Segal, J.; Cujba, A.-M.; Ng, S.S.; Butler, R.; Millar, V.; Hatch, S.; Zimri, S.; Koike, H.; et al. Imaging-Based Screen Identifies Laminin 411 as a Physiologically Relevant Niche Factor with Importance for i-Hep Applications. Stem Cell Rep. 2018, 10, 693–702. [Google Scholar] [CrossRef] [Green Version]
- Hay, D.C.; Zhao, D.; Fletcher, J.; Hewitt, Z.A.; McLean, D.; Urruticoechea-Uriguen, A.; Black, J.R.; Elcombe, C.; Ross, J.A.; Wolf, R.; et al. Efficient Differentiation of Hepatocytes from Human Embryonic Stem Cells Exhibiting Markers Recapitulating Liver Development In Vivo. Stem Cells 2008, 26, 894–902. [Google Scholar] [CrossRef] [Green Version]
- Baxter, M.; Withey, S.; Harrison, S.; Segeritz, C.-P.; Zhang, F.; Atkinson-Dell, R.; Rowe, C.; Gerrard, D.T.; Sison-Young, R.; Jenkins, R.; et al. Phenotypic and Functional Analyses Show Stem Cell-Derived Hepatocyte-like Cells Better Mimic Fetal Rather than Adult Hepatocytes. J. Hepatol. 2015, 62, 581–589. [Google Scholar] [CrossRef]
- Takayama, K.; Hagihara, Y.; Toba, Y.; Sekiguchi, K.; Sakurai, F.; Mizuguchi, H. Enrichment of High-Functioning Human IPS Cell-Derived Hepatocyte-like Cells for Pharmaceutical Research. Biomaterials 2018, 161, 24–32. [Google Scholar] [CrossRef]
- Toba, Y.; Deguchi, S.; Mimura, N.; Sakamoto, A.; Harada, K.; Hirata, K.; Takayama, K.; Mizuguchi, H. Comparison of Commercially Available Media for Hepatic Differentiation and Hepatocyte Maintenance. PLoS ONE 2020, 15, e0229654. [Google Scholar] [CrossRef] [PubMed]
- Matoba, N.; Yamashita, T.; Takayama, K.; Sakurai, F.; Mizuguchi, H. Optimal Human IPS Cell Culture Method for Efficient Hepatic Differentiation. Differentiation 2018, 104, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Mallanna, S.K.; Cayo, M.A.; Twaroski, K.; Gundry, R.L.; Duncan, S.A. Mapping the Cell-Surface N-Glycoproteome of Human Hepatocytes Reveals Markers for Selecting a Homogeneous Population of IPSC-Derived Hepatocytes. Stem Cell Rep. 2016, 7, 543–556. [Google Scholar] [CrossRef] [Green Version]
- Boon, R.; Kumar, M.; Tricot, T.; Elia, I.; Ordovas, L.; Jacobs, F.; One, J.; Smedt, J.D.; Eelen, G.; Bird, M.; et al. Amino Acid Levels Determine Metabolism and CYP450 Function of Hepatocytes and Hepatoma Cell Lines. Nat. Commun. 2020, 11, 1393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thi, V.L.D.; Wu, X.; Belote, R.L.; Andreo, U.; Takacs, C.N.; Fernandez, J.P.; Vale-Silva, L.A.; Prallet, S.; Decker, C.C.; Fu, R.M.; et al. Stem Cell-Derived Polarized Hepatocytes. Nat. Commun. 2020, 11, 1677. [Google Scholar] [CrossRef] [Green Version]
- Török, G.; Erdei, Z.; Lilienberg, J.; Apáti, Á.; Homolya, L. The Importance of Transporters and Cell Polarization for the Evaluation of Human Stem Cell-Derived Hepatic Cells. PLoS ONE 2020, 15, e0227751. [Google Scholar] [CrossRef]
- Kajiwara, M.; Aoi, T.; Okita, K.; Takahashi, R.; Inoue, H.; Takayama, N.; Endo, H.; Eto, K.; Toguchida, J.; Uemoto, S.; et al. Donor-Dependent Variations in Hepatic Differentiation from Human-Induced Pluripotent Stem Cells. Proc. Natl. Acad. Sci. USA 2012, 109, 12538–12543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamanaka, S. Pluripotent Stem Cell-Based Cell Therapy—Promise and Challenges. Cell Stem Cell 2020, 27, 523–531. [Google Scholar] [CrossRef] [PubMed]
- Du, C.; Feng, Y.; Qiu, D.; Xu, Y.; Pang, M.; Cai, N.; Xiang, A.P.; Zhang, Q. Highly Efficient and Expedited Hepatic Differentiation from Human Pluripotent Stem Cells by Pure Small-Molecule Cocktails. Stem Cell Res. Ther. 2018, 9, 58. [Google Scholar] [CrossRef] [PubMed]
- Meseguer-Ripolles, J.; Lucendo-Villarin, B.; Wang, Y.; Hay, D.C. Semi-Automated Production of Hepatocyte Like Cells from Pluripotent Stem Cells. J. Vis. Exp. Jove 2018, 57995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siller, R.; Greenhough, S.; Naumovska, E.; Sullivan, G.J. Small-Molecule-Driven Hepatocyte Differentiation of Human Pluripotent Stem Cells. Stem Cell Rep. 2015, 4, 939–952. [Google Scholar] [CrossRef] [Green Version]
- Shan, J.; Schwartz, R.E.; Ross, N.T.; Logan, D.J.; Thomas, D.; Duncan, S.A.; North, T.E.; Goessling, W.; Carpenter, A.E.; Bhatia, S.N. Identification of Small Molecules for Human Hepatocyte Expansion and IPS Differentiation. Nat. Chem. Biol. 2013, 9, 514–520. [Google Scholar] [CrossRef] [Green Version]
- Kamiya, A.; Kinoshita, T.; Miyajima, A. Oncostatin M and Hepatocyte Growth Factor Induce Hepatic Maturation via Distinct Signaling Pathways. FEBS Lett. 2001, 492, 90–94. [Google Scholar] [CrossRef] [Green Version]
- Fatehullah, A.; Tan, S.H.; Barker, N. Organoids as an in Vitro Model of Human Development and Disease. Nat. Cell Biol. 2016, 18, 246–254. [Google Scholar] [CrossRef] [Green Version]
- Marsee, A.; Roos, F.J.M.; Verstegen, M.M.A.; Consortium, H.O.; Marsee, A.; Roos, F.; Verstegen, M.; Clevers, H.; Vallier, L.; Takebe, T.; et al. Building Consensus on Definition and Nomenclature of Hepatic, Pancreatic, and Biliary Organoids. Cell Stem Cell 2021, 28, 816–832. [Google Scholar] [CrossRef]
- Wang, S.; Wang, X.; Tan, Z.; Su, Y.; Liu, J.; Chang, M.; Yan, F.; Chen, J.; Chen, T.; Li, C.; et al. Human ESC-Derived Expandable Hepatic Organoids Enable Therapeutic Liver Repopulation and Pathophysiological Modeling of Alcoholic Liver Injury. Cell Res. 2019, 29, 1009–1026. [Google Scholar] [CrossRef]
- Akbari, S.; Sevinç, G.G.; Ersoy, N.; Basak, O.; Kaplan, K.; Sevinç, K.; Ozel, E.; Sengun, B.; Enustun, E.; Ozcimen, B.; et al. Robust, Long-Term Culture of Endoderm-Derived Hepatic Organoids for Disease Modeling. Stem Cell Rep. 2019, 13, 627–641. [Google Scholar] [CrossRef] [Green Version]
- Ramli, M.N.B.; Lim, Y.S.; Koe, C.T.; Demircioglu, D.; Tng, W.; Gonzales, K.A.U.; Tan, C.P.; Szczerbinska, I.; Liang, H.; Soe, E.L.; et al. Human Pluripotent Stem Cell-Derived Organoids as Models of Liver Disease. Gastroenterology 2020, 159, 1471–1486. [Google Scholar] [CrossRef]
- Guan, Y.; Xu, D.; Garfin, P.M.; Ehmer, U.; Hurwitz, M.; Enns, G.; Michie, S.; Wu, M.; Zheng, M.; Nishimura, T.; et al. Human Hepatic Organoids for the Analysis of Human Genetic Diseases. JCI Insight 2017, 2, e94954. [Google Scholar] [CrossRef] [Green Version]
- Nie, Y.-Z.; Zheng, Y.-W.; Ogawa, M.; Miyagi, E.; Taniguchi, H. Human Liver Organoids Generated with Single Donor-Derived Multiple Cells Rescue Mice from Acute Liver Failure. Stem Cell Res. Ther. 2018, 9, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goulart, E.; de Caires-Junior, L.C.; Telles-Silva, K.A.; Araujo, B.H.S.; Kobayashi, G.S.; Musso, C.M.; Assoni, A.F.; Oliveira, D.; Caldini, E.; Gerstenhaber, J.A.; et al. Adult and IPS-Derived Non-Parenchymal Cells Regulate Liver Organoid Development through Differential Modulation of Wnt and TGF-β. Stem Cell Res. Ther. 2019, 10, 258. [Google Scholar] [CrossRef] [PubMed]
- Schepers, A.; Li, C.; Chhabra, A.; Seney, B.T.; Bhatia, S. Engineering a Perfusable 3D Human Liver Platform from IPS Cells. Lab Chip 2016, 16, 2644–2653. [Google Scholar] [CrossRef] [PubMed]
- Takebe, T.; Sekine, K.; Enomura, M.; Koike, H.; Kimura, M.; Ogaeri, T.; Zhang, R.-R.; Ueno, Y.; Zheng, Y.-W.; Koike, N.; et al. Vascularized and Functional Human Liver from an IPSC-Derived Organ Bud Transplant. Nature 2013, 499, 481–484. [Google Scholar] [CrossRef] [PubMed]
- Camp, J.G.; Sekine, K.; Gerber, T.; Loeffler-Wirth, H.; Binder, H.; Gac, M.; Kanton, S.; Kageyama, J.; Damm, G.; Seehofer, D.; et al. Multilineage Communication Regulates Human Liver Bud Development from Pluripotency. Nature 2017, 546, 533–538. [Google Scholar] [CrossRef]
- Takebe, T.; Sekine, K.; Kimura, M.; Yoshizawa, E.; Ayano, S.; Koido, M.; Funayama, S.; Nakanishi, N.; Hisai, T.; Kobayashi, T.; et al. Massive and Reproducible Production of Liver Buds Entirely from Human Pluripotent Stem Cells. Cell Rep. 2017, 21, 2661–2670. [Google Scholar] [CrossRef] [Green Version]
- Guye, P.; Ebrahimkhani, M.R.; Kipniss, N.; Velazquez, J.J.; Schoenfeld, E.; Kiani, S.; Griffith, L.G.; Weiss, R. Genetically Engineering Self-Organization of Human Pluripotent Stem Cells into a Liver Bud-like Tissue Using Gata6. Nat. Commun. 2016, 7, 10243. [Google Scholar] [CrossRef] [Green Version]
- Ouchi, R.; Togo, S.; Kimura, M.; Shinozawa, T.; Koido, M.; Koike, H.; Thompson, W.; Karns, R.A.; Mayhew, C.N.; McGrath, P.S.; et al. Modeling Steatohepatitis in Humans with Pluripotent Stem Cell-Derived Organoids. Cell Metab. 2019, 30, 374–384. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, H.; Deng, P.; Chen, W.; Guo, Y.; Tao, T.; Qin, J. In Situ Differentiation and Generation of Functional Liver Organoids from Human IPSCs in a 3D Perfusable Chip System. Lab Chip 2018, 18, 3606–3616. [Google Scholar] [CrossRef] [PubMed]
- Yin, F.; Zhang, X.; Wang, L.; Wang, Y.; Zhu, Y.; Li, Z.; Tao, T.; Chen, W.; Yu, H.; Qin, J. HiPSC-Derived Multi-Organoids-on-Chip System for Safety Assessment of Antidepressant Drugs. Lab Chip 2020, 21, 571–581. [Google Scholar] [CrossRef] [PubMed]
- Ng, S.S.; Saeb-Parsy, K.; Blackford, S.J.I.; Segal, J.M.; Serra, M.P.; Horcas-Lopez, M.; No, D.Y.; Mastoridis, S.; Jassem, W.; Frank, C.W.; et al. Human IPS Derived Progenitors Bioengineered into Liver Organoids Using an Inverted Colloidal Crystal Poly (Ethylene Glycol) Scaffold. Biomaterials 2018, 182, 299–311. [Google Scholar] [CrossRef] [PubMed]
- Cayo, M.A.; Mallanna, S.K.; Furio, F.D.; Jing, R.; Tolliver, L.B.; Bures, M.; Urick, A.; Noto, F.K.; Pashos, E.E.; Greseth, M.D.; et al. A Drug Screen Using Human IPSC-Derived Hepatocyte-like Cells Reveals Cardiac Glycosides as a Potential Treatment for Hypercholesterolemia. Cell Stem Cell 2017, 20, 478–489. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Wang, Y.; Zhou, T.; Wong, L.-Y.; Tian, X.-Y.; Hong, X.; Lai, W.-H.; Au, K.-W.; Wei, R.; Liu, Y.; et al. Generation of Human Liver Chimeric Mice with Hepatocytes from Familial Hypercholesterolemia Induced Pluripotent Stem Cells. Stem Cell Rep. 2017, 8, 605–618. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.; Duan, L.; He, X.; Li, S.; Wu, Y.; Xiang, G.; Bao, F.; Yang, L.; Shi, H.; Gao, M.; et al. A Combined Model of Human IPSC-Derived Liver Organoids and Hepatocytes Reveals Ferroptosis in DGUOK Mutant MtDNA Depletion Syndrome. Adv. Sci. 2021, 8, 2004680. [Google Scholar] [CrossRef] [PubMed]
- Jing, R.; Corbett, J.L.; Cai, J.; Beeson, G.C.; Beeson, C.C.; Chan, S.S.; Dimmock, D.P.; Lazcares, L.; Geurts, A.M.; Lemasters, J.J.; et al. A Screen Using IPSC-Derived Hepatocytes Reveals NAD(+) as a Potential Treatment for MtDNA Depletion Syndrome. Cell Rep. 2018, 25, 1469–1484. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Kim, S.-B.; Ryu, J.L.; Hong, H.; Chang, J.-H.; Yoo, T.-J.; Jin, X.; Park, H.-J.; Han, C.; Lee, B.H.; et al. Human Embryonic Stem Cell-Derived Wilson’s Disease Model for Screening Drug Efficacy. Cells 2020, 9, 872. [Google Scholar] [CrossRef] [Green Version]
- Overeem, A.W.; Klappe, K.; Parisi, S.; Klöters-Planchy, P.; Mataković, L.; du Espina, M.T.; Drouin, C.A.; Weiss, K.H.; van IJzendoorn, S.C.D. Pluripotent Stem Cell-Derived Bile Canaliculi-Forming Hepatocytes to Study Genetic Liver Diseases Involving Hepatocyte Polarity. J. Hepatol. 2019, 71, 344–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estève, J.; Blouin, J.-M.; Lalanne, M.; Azzi-Martin, L.; Dubus, P.; Bidet, A.; Harambat, J.; Llanas, B.; Moranvillier, I.; Bedel, A.; et al. Generation of Induced Pluripotent Stem Cells-Derived Hepatocyte-like Cells for Ex Vivo Gene Therapy of Primary Hyperoxaluria Type 1. Stem Cell Res. 2019, 38, 101467. [Google Scholar] [CrossRef] [PubMed]
- Tsunoda, T.; Kakinuma, S.; Miyoshi, M.; Kamiya, A.; Kaneko, S.; Sato, A.; Tsuchiya, J.; Nitta, S.; Kawai-Kitahata, F.; Murakawa, M.; et al. Loss of Fibrocystin Promotes Interleukin-8-Dependent Proliferation and CTGF Production of Biliary Epithelium. J. Hepatol. 2019, 71, 143–152. [Google Scholar] [CrossRef]
- Zabulica, M.; Jakobsson, T.; Ravaioli, F.; Vosough, M.; Gramignoli, R.; Ellis, E.; Rooyackers, O.; Strom, S.C. Gene Editing Correction of a Urea Cycle Defect in Organoid Stem Cell Derived Hepatocyte-like Cells. Int. J. Mol. Sci. 2021, 22, 1217. [Google Scholar] [CrossRef]
- Sinton, M.C.; Meseguer-Ripolles, J.; Lucendo-Villarin, B.; Wernig-Zorc, S.; Thomson, J.P.; Carter, R.N.; Lyall, M.J.; Walker, P.D.; Thakker, A.; Meehan, R.R.; et al. A Human Pluripotent Stem Cell Model for the Analysis of Metabolic Dysfunction in Hepatic Steatosis. Iscience 2021, 24, 101931. [Google Scholar] [CrossRef]
- Mun, S.J.; Ryu, J.-S.; Lee, M.-O.; Son, Y.S.; Oh, S.J.; Cho, H.-S.; Son, M.-Y.; Kim, D.-S.; Kim, S.J.; Yoo, H.J.; et al. Generation of Expandable Human Pluripotent Stem Cell-Derived Hepatocyte-like Liver Organoids. J. Hepatol. 2019, 71, 970–985. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, H.; Deng, P.; Tao, T.; Liu, H.; Wu, S.; Chen, W.; Qin, J. Modeling Human Nonalcoholic Fatty Liver Disease (NAFLD) with an Organoids-on-a-Chip System. ACS Biomater. Sci. Eng. 2020, 6, 5734–5743. [Google Scholar] [CrossRef]
- De l’Hortet, A.C.; Takeishi, K.; Guzman-Lepe, J.; Morita, K.; Achreja, A.; Popovic, B.; Wang, Y.; Handa, K.; Mittal, A.; Meurs, N.; et al. Generation of Human Fatty Livers Using Custom-Engineered Induced Pluripotent Stem Cells with Modifiable SIRT1 Metabolism. Cell Metab. 2019, 30, 385–401.e9. [Google Scholar] [CrossRef]
- Xia, Y.; Carpentier, A.; Cheng, X.; Block, P.D.; Zhao, Y.; Zhang, Z.; Protzer, U.; Liang, T.J. Human Stem Cell-Derived Hepatocytes as a Model for Hepatitis B Virus Infection, Spreading and Virus-Host Interactions. J. Hepatol. 2017, 66, 494–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nie, Y.-Z.; Zheng, Y.-W.; Miyakawa, K.; Murata, S.; Zhang, R.-R.; Sekine, K.; Ueno, Y.; Takebe, T.; Wakita, T.; Ryo, A.; et al. Recapitulation of Hepatitis B Virus–Host Interactions in Liver Organoids from Human Induced Pluripotent Stem Cells. Ebiomedicine 2018, 35, 114–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, L.; Liu, X.; Zhang, L.; Li, X.; Zhang, Y.; Wu, K.; Chen, Y.; Cao, J.; Hou, W.; Zhang, J.; et al. A Chimeric Humanized Mouse Model by Engrafting the Human Induced Pluripotent Stem Cell-Derived Hepatocyte-Like Cell for the Chronic Hepatitis B Virus Infection. Front. Microbiol. 2018, 9, 908. [Google Scholar] [CrossRef]
- Carpentier, A.; Tesfaye, A.; Chu, V.; Nimgaonkar, I.; Zhang, F.; Lee, S.B.; Thorgeirsson, S.S.; Feinstone, S.M.; Liang, T.J. Engrafted Human Stem Cell–Derived Hepatocytes Establish an Infectious HCV Murine Model. J. Clin. Investig. 2014, 124, 4953–4964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira, C.R.; Cassiman, D.; Blau, N. Clinical and Biochemical Footprints of Inherited Metabolic Diseases. II. Metabolic Liver Diseases. Mol. Genet. Metab. 2019, 127, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Rashid, S.T.; Corbineau, S.; Hannan, N.; Marciniak, S.J.; Miranda, E.; Alexander, G.; Huang-Doran, I.; Griffin, J.; Ahrlund-Richter, L.; Skepper, J.; et al. Modeling Inherited Metabolic Disorders of the Liver Using Human Induced Pluripotent Stem Cells. J. Clin. Investig. 2010, 120, 3127–3136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estève, J.; Blouin, J.-M.; Lalanne, M.; Azzi-Martin, L.; Dubus, P.; Bidet, A.; Harambat, J.; Llanas, B.; Moranvillier, I.; Bedel, A.; et al. Targeted Gene Therapy in Human-Induced Pluripotent Stem Cells from a Patient with Primary Hyperoxaluria Type 1 Using CRISPR/Cas9 Technology. Biochem. Biophys. Res. Commun. 2019, 517, 677–683. [Google Scholar] [CrossRef] [PubMed]
- Morishige, S.; Mizuno, S.; Ozawa, H.; Nakamura, T.; Mazahery, A.; Nomura, K.; Seki, R.; Mouri, F.; Osaki, K.; Yamamura, K.; et al. CRISPR/Cas9-Mediated Gene Correction in Hemophilia B Patient-Derived IPSCs. Int. J. Hematol. 2020, 111, 225–233. [Google Scholar] [CrossRef]
- Yusa, K.; Rashid, S.T.; Strick-Marchand, H.; Varela, I.; Liu, P.-Q.; Paschon, D.E.; Miranda, E.; Ordóñez, A.; Hannan, N.R.F.; Rouhani, F.J.; et al. Targeted Gene Correction of A1-Antitrypsin Deficiency in Induced Pluripotent Stem Cells. Nature 2011, 478, 391–394. [Google Scholar] [CrossRef] [Green Version]
- Fernando, D.H.; Forbes, J.M.; Angus, P.W.; Herath, C.B. Development and Progression of Non-Alcoholic Fatty Liver Disease: The Role of Advanced Glycation End Products. Int. J. Mol. Sci. 2019, 20, 5037. [Google Scholar] [CrossRef] [Green Version]
- DeWeerdt, S. Disease Progression: Divergent Paths. Nature 2017, 551, S92–S93. [Google Scholar] [CrossRef] [Green Version]
- Parafati, M.; Kirby, R.J.; Khorasanizadeh, S.; Rastinejad, F.; Malany, S. A Nonalcoholic Fatty Liver Disease Model in Human Induced Pluripotent Stem Cell-Derived Hepatocytes, Created by Endoplasmic Reticulum Stress-Induced Steatosis. Dis. Model. Mech. 2018, 11, dmm033530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakurai, F.; Mitani, S.; Yamamoto, T.; Takayama, K.; Tachibana, M.; Watashi, K.; Wakita, T.; Iijima, S.; Tanaka, Y.; Mizuguchi, H. Human Induced-Pluripotent Stem Cell-Derived Hepatocyte-like Cells as an in Vitro Model of Human Hepatitis B Virus Infection. Sci. Rep. 2017, 7, 45698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Wang, Y.; Shen, F.; Wu, M.; Li, Y.; Fang, Z.; Ye, J.; Wang, L.; Gao, L.; Yuan, Z.; et al. Identification of Retinoic Acid Receptor Agonists as Potent Hepatitis B Virus Inhibitors via a Drug Repurposing Screen. Antimicrob. Agents Chemother. 2018, 62, e00465-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Wang, C.; Liu, Z.; Zou, G.; Li, J.; Lu, M. Host Genetic Determinants of Hepatitis B Virus Infection. Front. Genet. 2019, 10, 696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torresi, J.; Tran, B.M.; Christiansen, D.; Earnest-Silveira, L.; Schwab, R.H.M.; Vincan, E. HBV-Related Hepatocarcinogenesis: The Role of Signalling Pathways and Innovative Ex Vivo Research Models. BMC Cancer 2019, 19, 707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alter, H.J.; Chisari, F.V. Is Elimination of Hepatitis B and C a Pipe Dream or Reality? Gastroenterology 2019, 156, 294–296. [Google Scholar] [CrossRef]
- Wang, H.; Calvisi, D.; Chen, X. Organoids for the Study of Liver Cancer. Semin. Liver Dis. 2021, 41, 19–27. [Google Scholar] [CrossRef]
- Afify, S.M.; Calle, A.S.; Hassan, G.; Kumon, K.; Nawara, H.M.; Zahra, M.H.; Mansour, H.M.; Khayrani, A.C.; Alam, M.J.; Du, J.; et al. A Novel Model of Liver Cancer Stem Cells Developed from Induced Pluripotent Stem Cells. Br. J. Cancer 2020, 122, 1378–1390. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Yan, Q.; Sun, Y.; Nam, Y.; Hu, L.; Loong, J.H.; Ouyang, Q.; Zhang, Y.; Li, H.-L.; Kong, F.-E.; et al. A Hepatocyte Differentiation Model Reveals Two Subtypes of Liver Cancer with Different Oncofetal Properties and Therapeutic Targets. Proc. Natl. Acad. Sci. USA 2020, 117, 6103–6113. [Google Scholar] [CrossRef]
- Ostapowicz, G.; Fontana, R.J.; Schiødt, F.V.; Larson, A.; Davern, T.J.; Han, S.H.B.; McCashland, T.M.; Shakil, A.O.; Hay, J.E.; Hynan, L.; et al. Results of a Prospective Study of Acute Liver Failure at 17 Tertiary Care Centers in the United States. Ann. Intern. Med. 2002, 137, 947. [Google Scholar] [CrossRef]
- Choudhury, Y.; Toh, Y.C.; Xing, J.; Qu, Y.; Poh, J.; Li, H.; Tan, H.S.; Kanesvaran, R.; Yu, H.; Tan, M.-H. Patient-Specific Hepatocyte-like Cells Derived from Induced Pluripotent Stem Cells Model Pazopanib-Mediated Hepatotoxicity. Sci. Rep. 2017, 7, 41238. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.-T.; Lamprecht, M.P.; Duncan, S.A. Using Human Induced Pluripotent Stem Cell-Derived Hepatocyte-like Cells for Drug Discovery. J. Vis. Exp. 2018, e57194. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.; Kim, H.; Park, J.Y.; Kim, G.; Han, J.; Chung, S.; Yang, J.H.; Jeon, J.S.; Woo, D.-H.; Han, C.; et al. Generation of Uniform Liver Spheroids from Human Pluripotent Stem Cells for Imaging-Based Drug Toxicity Analysis. Biomaterials 2021, 269, 120529. [Google Scholar] [CrossRef] [PubMed]
- Shinozawa, T.; Kimura, M.; Cai, Y.; Saiki, N.; Yoneyama, Y.; Ouchi, R.; Koike, H.; Maezawa, M.; Zhang, R.-R.; Dunn, A.; et al. High-Fidelity Drug-Induced Liver Injury Screen Using Human Pluripotent Stem Cell–Derived Organoids. Gastroenterology 2021, 160, 831–846.e10. [Google Scholar] [CrossRef] [PubMed]
- Bell, C.C.; Lauschke, V.M.; Vorrink, S.U.; Palmgren, H.; Duffin, R.; Andersson, T.B.; Ingelman-Sundberg, M. Transcriptional, Functional and Mechanistic Comparisons of Stem Cell-Derived Hepatocytes, HepaRG Cells and 3D Human Hepatocyte Spheroids as Predictive in Vitro Systems for Drug-Induced Liver Injury. Drug Metab. Dispos. 2017, 45, 419–429. [Google Scholar] [CrossRef] [Green Version]
- Clinical Study of HAES Transplantation in Patients with Neonatal Onset Urea Cycle Disorder. Available online: https://rctportal.niph.go.jp/en/detail?trial_id=JMA-IIA00412 (accessed on 22 September 2021).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Differentiation Stages | Growth Factors | Target Signaling Pathways | Alternative Small Molecules |
|---|---|---|---|
| Endoderm induction | activin A wnt3a | Activin/Nodal signaling pathway Wnt/β-catenin signaling pathway | CHIR99021 [9]; NaB [14]; IWR-1 [10]; |
| Hepatic specification | FGF4 BMP2 | MAPK signaling pathway PI3K signaling pathway TGF-β signaling pathway | A83–01 [25]; DMSO [27]; |
| Hepatic maturation | OSM HGF | OSM/gp130 signaling pathway STAT3-independent HGF signaling pathway | FH1 + FPH1 [28]; Dex [29]; Dihexa [27]; |
| Diseases | Modeling Strategies | Applications |
|---|---|---|
| Familial hypercholesterolemia | HLCs from patient-derived iPSCs | High-throughput drug screening [47]; Preclinical drug efficacy evaluation [48] |
| mtDNA depletion syndrome | HLCs and liver organoids from patient-derived iPSCs and gene-corrected counterparts | Disease mechanism investigation [49] |
| HLCs from genetically engineered iPSCs (DGUOK deficient) | High-throughput drug screening [50] | |
| Wilson’s disease | HLCs from genetically engineered hPSCs (mutations in the ATP7B gene) | Preclinical drug efficacy evaluation [51]; Disease features modeling [52] |
| Primary hyperoxaluria type 1 | HLCs from genetically corrected patient-derived iPSCs | Therapeutical gene correction [53] |
| Congenital hepatic fibrosis | HLCs from genetically engineered iPSCs (PKHD1 knockout) | Disease mechanism investigation [54] |
| Urea cycle disorders | Liver organoids from patient-derived iPSCs andthe gene-corrected counterpart | Therapeutical gene correction [55] |
| Liver steatosis | HLCs cultured in monolayers | Disease mechanism investigation [56] |
| Liver organoids | Anti-steatosis drug screening [57]; Disease features modeling [34] | |
| Multicellular liver organoids from patient-derived iPSCs | Modeling the progressive features of steatohepatitis [43] | |
| Liver organoids-on-a-chip system | Modeling the features of fatty liver using bioengineered systems [58] | |
| Biofabricated human fatty liver tissue with SIRT1 knock down hiPSC-derived HLCs | Investigating the effects of a specific gene [59] | |
| HBV infection | HLCs infected with HBV | Novel antiviral agent identification [60] |
| Liver organoids infected with HBV | Recapitulating virus life cycle and hepatic dysfunction in 3D organoids [61] | |
| Chimeric mice engrafted with hiPSC-HLCs | Antivirals evaluation in vitro and in vivo [62] | |
| HCV infection | Chimeric mice engrafted with hPSC-derived hepatic lineages | Long-term infection of multiple HCV genotypes [63] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Yang, X.; Plummer, R.; Hayashi, Y.; Deng, X.-S.; Nie, Y.-Z.; Taniguchi, H. Human Pluripotent Stem Cell-Derived Hepatocyte-Like Cells and Organoids for Liver Disease and Therapy. Int. J. Mol. Sci. 2021, 22, 10471. https://doi.org/10.3390/ijms221910471
Li Y, Yang X, Plummer R, Hayashi Y, Deng X-S, Nie Y-Z, Taniguchi H. Human Pluripotent Stem Cell-Derived Hepatocyte-Like Cells and Organoids for Liver Disease and Therapy. International Journal of Molecular Sciences. 2021; 22(19):10471. https://doi.org/10.3390/ijms221910471
Chicago/Turabian StyleLi, Yang, Xia Yang, Richie Plummer, Yoshihito Hayashi, Xiao-Shan Deng, Yun-Zhong Nie, and Hideki Taniguchi. 2021. "Human Pluripotent Stem Cell-Derived Hepatocyte-Like Cells and Organoids for Liver Disease and Therapy" International Journal of Molecular Sciences 22, no. 19: 10471. https://doi.org/10.3390/ijms221910471
APA StyleLi, Y., Yang, X., Plummer, R., Hayashi, Y., Deng, X.-S., Nie, Y.-Z., & Taniguchi, H. (2021). Human Pluripotent Stem Cell-Derived Hepatocyte-Like Cells and Organoids for Liver Disease and Therapy. International Journal of Molecular Sciences, 22(19), 10471. https://doi.org/10.3390/ijms221910471