
The Germinal Center Milieu in Rheumatoid Arthritis: The Immunological Drummer or Dancer?

 ,
,
Abstract
:1. Introduction
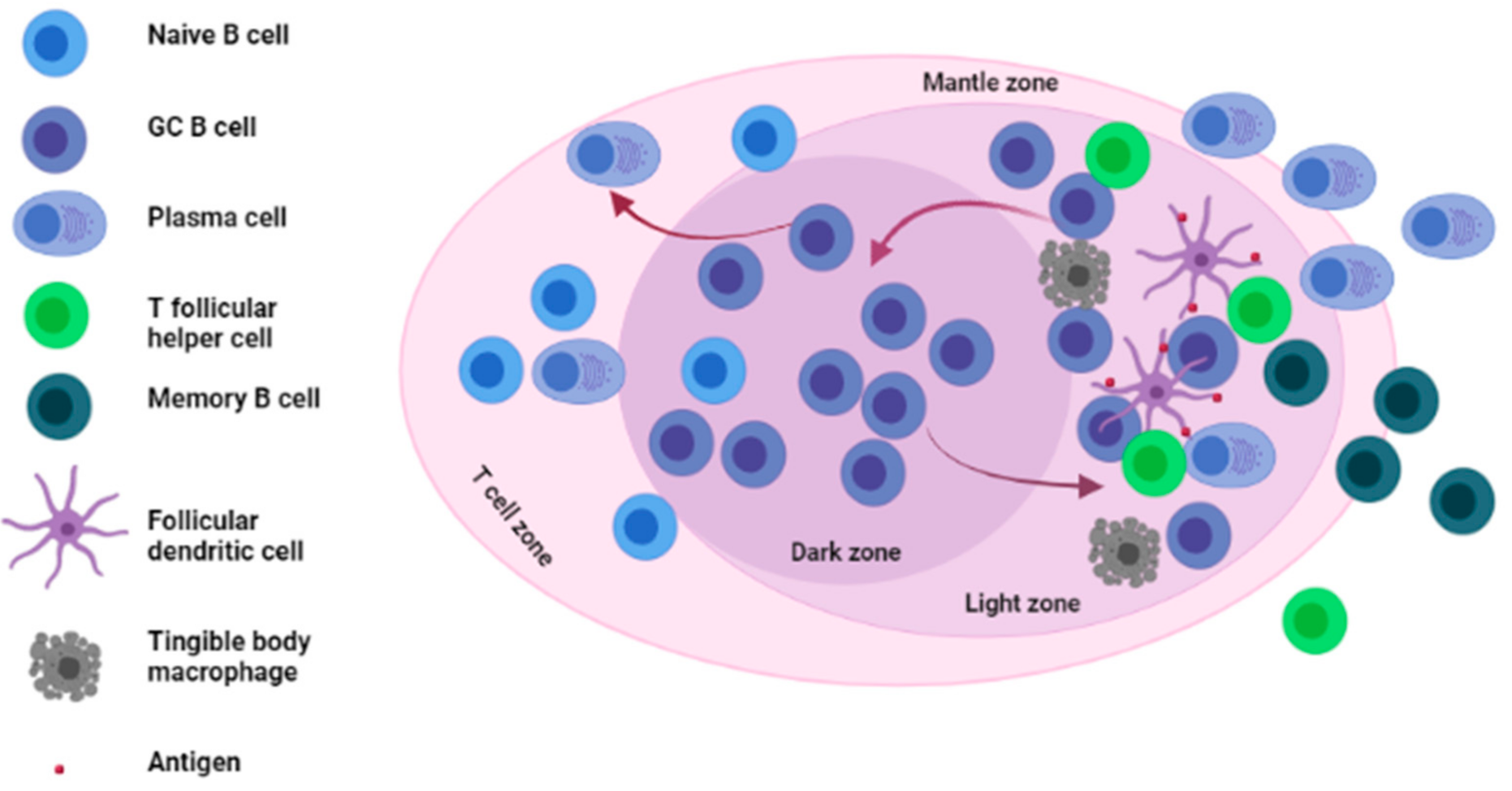
2. Germinal Centers and Adaptive Immune Responses
3. Germinal Centers in Rheumatoid Arthritis
3.1. The Presence of GCs and Ectopic GCs in Tissues of RA Patients and Mouse Models of Arthritis
3.1.1. The Presence of GCs and Ectopic GCs in Tissues of RA Patients
3.1.2. The Presence of GCs and Increased GC Responses in Tissues of RA Mouse Models
3.2. SHM within GCs and the Generation of RA-Related Autoantibodies
3.3. Defective GC-Resident Cellular Subsets and the Generation of Autoantibodies in RA
3.3.1. Defective Tfh Cell Functioning within GCs
3.3.2. Defective Tfr Cell Functioning within GCs
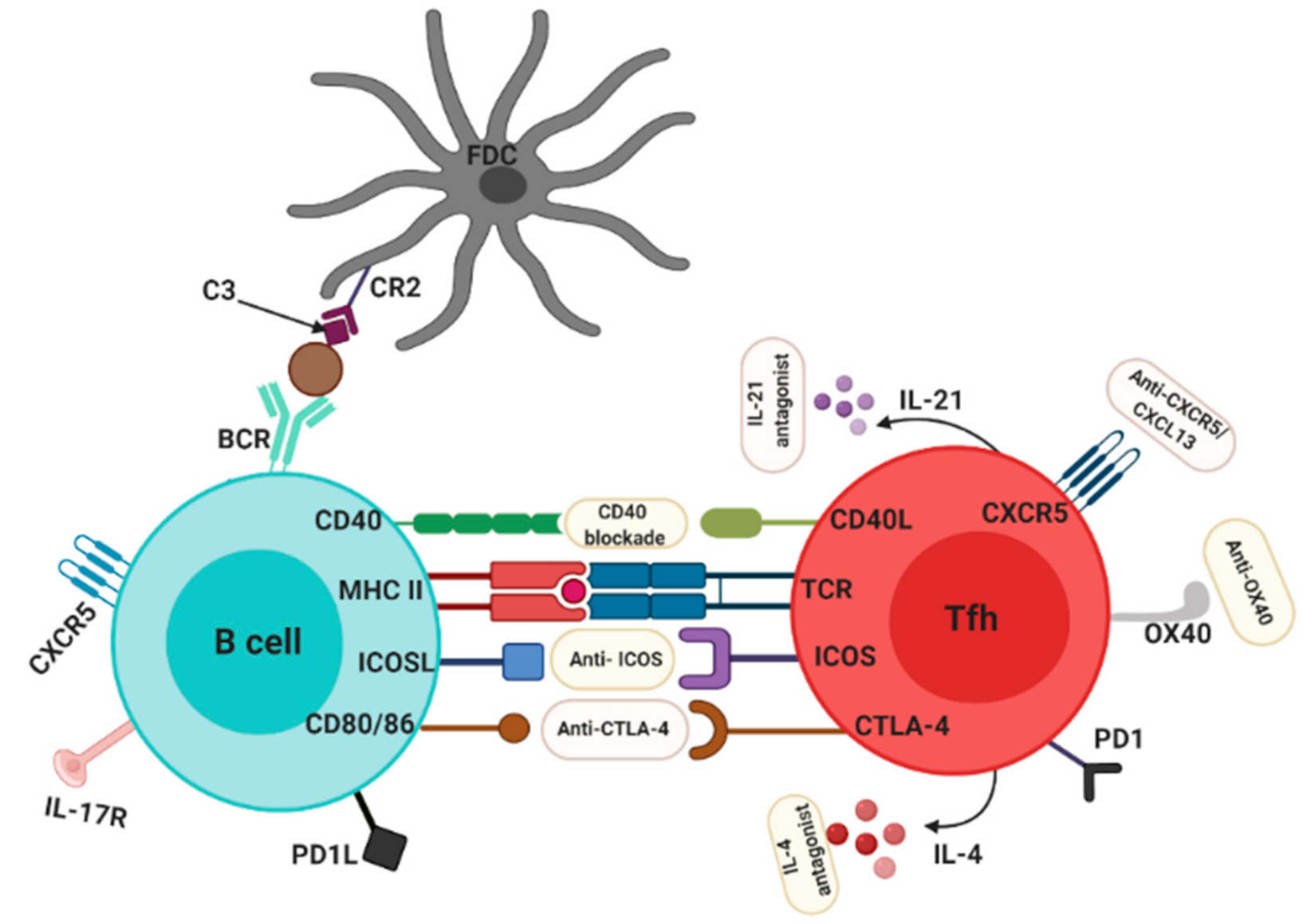
4. Interfering with GC Components as Targets in the Treatment of RA
4.1. Interference with Various GC Processes
4.2. Targeting Tfh Cells for the Treatment of RA
4.3. Tfh-Associated Cytokines as Targets in the Treatment of RA: New Biologics?
4.4. Targeting Tfh Activation and Co-Stimulation Pathways in the Treatment of RA
5. Questions Raised and Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Silman, A.J.; Pearson, J.E. Epidemiology and genetics of rheumatoid arthritis. Arthritis Res. 2002, 4, S265–S272. [Google Scholar] [CrossRef] [Green Version]
- Shapira, Y.; Agmon-Levin, N.; Shoenfeld, Y. Geoepidemiology of autoimmune rheumatic diseases. Nat. Rev. Rheumatol. 2010, 6, 468–476. [Google Scholar] [CrossRef] [PubMed]
- Edwards, J.C.; Szczepanski, L.; Szechinski, J.; Filipowicz-Sosnowska, A.; Emery, P.; Close, D.R.; Stevens, R.M.; Shaw, T. Efficacy of B-cell-targeted therapy with rituximab in patients with rheumatoid arthritis. N. Engl. J. Med. 2004, 350, 2572–2581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerlag, D.M.; Safy, M.; Maijer, K.I.; Tang, M.W.; Tas, S.W.; Starmans-Kool, M.J.F.; van Tubergen, A.; Janssen, M.; de Hair, M.; Hansson, M.; et al. Effects of B-cell directed therapy on the preclinical stage of rheumatoid arthritis: The PRAIRI study. Ann. Rheum. Dis. 2019, 78, 179–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Woude, D.; Rantapaa-Dahlqvist, S.; Ioan-Facsinay, A.; Onnekink, C.; Schwarte, C.M.; Verpoort, K.N.; Drijfhout, J.W.; Huizinga, T.W.; Toes, R.E.; Pruijn, G.J. Epitope spreading of the anti-citrullinated protein antibody response occurs before disease onset and is associated with the disease course of early arthritis. Ann. Rheum. Dis. 2010, 69, 1554–1561. [Google Scholar] [CrossRef]
- Van de Stadt, L.A.; van der Horst, A.R.; de Koning, M.H.; Bos, W.H.; Wolbink, G.J.; van de Stadt, R.J.; Pruijn, G.J.; Dijkmans, B.A.; van Schaardenburg, D.; Hamann, D. The extent of the anti-citrullinated protein antibody repertoire is associated with arthritis development in patients with seropositive arthralgia. Ann. Rheum. Dis. 2011, 70, 128–133. [Google Scholar] [CrossRef] [PubMed]
- Hozumi, N.; Tonegawa, S. Evidence for somatic rearrangement of immunoglobulin genes coding for variable and constant regions. Proc. Natl. Acad. Sci. USA 1976, 73, 3628–3632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wing, J.B.; Tekgüç, M.; Sakaguchi, S. Control of Germinal Center Responses by T-Follicular Regulatory Cells. Front. Immunol. 2018, 9, 1910. [Google Scholar] [CrossRef]
- Xie, M.M.; Dent, A.L. Unexpected Help: Follicular Regulatory T Cells in the Germinal Center. Front. Immunol. 2018, 9, 1536. [Google Scholar] [CrossRef]
- Nieuwenhuis, P.; Opstelten, D. Functional anatomy of germinal centers. Am. J. Anat. 1984, 170, 421–435. [Google Scholar] [CrossRef]
- De Silva, N.S.; Klein, U. Dynamics of B cells in germinal centres. Nat. Rev. Immunol. 2015, 15, 137–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Victora, G.D.; Nussenzweig, M.C. Germinal Centers. Ann. Rev. Immunol. 2012, 30, 429–457. [Google Scholar] [CrossRef] [PubMed]
- Jacob, J.; Kelsoe, G.; Rajewsky, K.; Weiss, U. Intraclonal generation of antibody mutants in germinal centres. Nature 1991, 354, 389–392. [Google Scholar] [CrossRef]
- Brink, R.; Phan, T.G. Self-Reactive B Cells in the Germinal Center Reaction. Annu. Rev. Immunol. 2018, 36, 339–357. [Google Scholar] [CrossRef]
- Tiburzy, B.; Szyska, M.; Iwata, H.; Chrobok, N.; Kulkarni, U.; Hirose, M.; Ludwig, R.J.; Kalies, K.; Westermann, J.; Wong, D.; et al. Persistent Autoantibody-Production by Intermediates between Short-and Long-Lived Plasma Cells in Inflamed Lymph Nodes of Experimental Epidermolysis Bullosa Acquisita. PLoS ONE 2013, 8, e83631. [Google Scholar] [CrossRef] [PubMed]
- Scherer, H.U.; Kerkman, P.F.; van der Voort, E.I.H.; Trouw, L.A.; Huizinga, T.W.J.; Toes, R.E.M. ACPA production by circulating B cells isolated from peripheral blood of patients with rheumatoid arthritis. Ann. Rheum. Dis. 2012, 71, A33. [Google Scholar] [CrossRef] [Green Version]
- Vinuesa, C.G.; Sanz, I.; Cook, M.C. Dysregulation of germinal centres in autoimmune disease. Nat. Rev. Immunol. 2009, 9, 845–857. [Google Scholar] [CrossRef]
- Kim, H.-J.; Krenn, V.; Steinhauser, G.; Berek, C. Plasma Cell Development in Synovial Germinal Centers in Patients with Rheumatoid and Reactive Arthritis. J. Immunol. 1999, 162, 3053–3062. [Google Scholar]
- Wagner, U.G.; Kurtin, P.J.; Wahner, A.; Brackertz, M.; Berry, D.J.; Goronzy, J.J.; Weyand, C.M. The role of CD8+ CD40L+ T cells in the formation of germinal centers in rheumatoid synovitis. J. Immunol. 1998, 161, 6390–6397. [Google Scholar]
- Luzina, I.G.; Atamas, S.P.; Storrer, C.E.; daSilva, L.C.; Kelsoe, G.; Papadimitriou, J.C.; Handwerger, B.S. Spontaneous formation of germinal centers in autoimmune mice. J. Leukoc. Biol. 2001, 70, 578–584. [Google Scholar]
- Domeier, P.P.; Schell, S.L.; Rahman, Z.S.M. Spontaneous germinal centers and autoimmunity. Autoimmunity 2017, 50, 4–18. [Google Scholar] [CrossRef] [Green Version]
- Mietzner, B.; Tsuiji, M.; Scheid, J.; Velinzon, K.; Tiller, T.; Abraham, K.; Gonzalez, J.B.; Pascual, V.; Stichweh, D.; Wardemann, H.; et al. Autoreactive IgG memory antibodies in patients with systemic lupus erythematosus arise from nonreactive and polyreactive precursors. Proc. Natl. Acad. Sci. USA 2008, 105, 9727–9732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Zenzo, G.; Di Lullo, G.; Corti, D.; Calabresi, V.; Sinistro, A.; Vanzetta, F.; Didona, B.; Cianchini, G.; Hertl, M.; Eming, R.; et al. Pemphigus autoantibodies generated through somatic mutations target the desmoglein-3 cis-interface. J. Clin. Investig. 2012, 122, 3781–3790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allen, C.D.C.; Cyster, J.G. Follicular dendritic cell networks of primary follicles and germinal centers: Phenotype and function. Semin. Immunol. 2008, 20, 14–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berek, C.; Berger, A.; Apel, M. Maturation of the immune response in germinal centers. Cell 1991, 67, 1121–1129. [Google Scholar] [CrossRef]
- Arulraj, T.; Binder, S.C.; Robert, P.A.; Meyer-Hermann, M. Germinal Centre Shutdown. Front. Immunol. 2021, 12, 2730. [Google Scholar] [CrossRef] [PubMed]
- Young, C.; Brink, R. Germinal centers and autoantibodies. Immunol. Cell Biol. 2020, 98, 480–489. [Google Scholar] [CrossRef]
- Shlomchik, M.J.; Luo, W.; Weisel, F. Linking signaling and selection in the germinal center. Immunol. Rev. 2019, 288, 49–63. [Google Scholar] [CrossRef]
- Chan, T.D.; Gatto, D.; Wood, K.; Camidge, T.; Basten, A.; Brink, R. Antigen Affinity Controls Rapid T-Dependent Antibody Production by Driving the Expansion Rather than the Differentiation or Extrafollicular Migration of Early Plasmablasts. J. Immunol. 2009, 183, 3139–3149. [Google Scholar] [CrossRef] [Green Version]
- Allen, C.D.; Okada, T.; Cyster, J.G. Germinal-center organization and cellular dynamics. Immunity 2007, 27, 190–202. [Google Scholar] [CrossRef] [Green Version]
- Victora, G.D.; Schwickert, T.A.; Fooksman, D.R.; Kamphorst, A.O.; Meyer-Hermann, M.; Dustin, M.L.; Nussenzweig, M.C. Germinal center dynamics revealed by multiphoton microscopy with a photoactivatable fluorescent reporter. Cell 2010, 143, 592–605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gatto, D.; Brink, R. The germinal center reaction. J. Allergy Clin. Immunol. 2010, 126, 898–907. [Google Scholar] [CrossRef] [PubMed]
- Meyer-Hermann, M.; Mohr, E.; Pelletier, N.; Zhang, Y.; Gabriel, D.; Toellner, K.-M. A Theory of Germinal Center B Cell Selection, Division, and Exit. Cell Rep. 2012, 2, 162–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Tech, L.; George, L.A.; Acs, A.; Durrett, R.E.; Hess, H.; Walker, L.S.K.; Tarlinton, D.M.; Fletcher, A.L.; Hauser, A.E.; et al. Plasma cell output from germinal centers is regulated by signals from Tfh and stromal cells. J. Exp. Med. 2018, 215, 1227–1243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oprea, M.; Perelson, A.S. Somatic mutation leads to efficient affinity maturation when centrocytes recycle back to centroblasts. J. Immunol. 1997, 158, 5155–5162. [Google Scholar]
- Tarlinton, D.; Radbruch, A.; Hiepe, F.; Dorner, T. Plasma cell differentiation and survival. Curr. Opin. Immunol. 2008, 20, 162–169. [Google Scholar] [CrossRef] [PubMed]
- Woods, M.; Zou, Y.R.; Davidson, A. Defects in Germinal Center Selection in SLE. Front. Immunol. 2015, 6, 425. [Google Scholar] [CrossRef] [Green Version]
- Bombardieri, M.; Lewis, M.; Pitzalis, C. Ectopic lymphoid neogenesis in rheumatic autoimmune diseases. Nat. Rev. Rheumatol. 2017, 13, 141–154. [Google Scholar] [CrossRef]
- Humby, F.; Bombardieri, M.; Manzo, A.; Kelly, S.; Blades, M.C.; Kirkham, B.; Spencer, J.; Pitzalis, C. Ectopic lymphoid structures support ongoing production of class-switched autoantibodies in rheumatoid synovium. PLoS Med. 2009, 6, e1. [Google Scholar] [CrossRef]
- Takemura, S.; Braun, A.; Crowson, C.; Kurtin, P.J.; Cofield, R.H.; O’Fallon, W.M.; Goronzy, J.J.; Weyand, C.M. Lymphoid neogenesis in rheumatoid synovitis. J. Immunol. 2001, 167, 1072–1080. [Google Scholar] [CrossRef] [Green Version]
- Shipman, W.D.; Dasoveanu, D.C.; Lu, T.T. Tertiary lymphoid organs in systemic autoimmune diseases: Pathogenic or protective? F1000Research 2017, 6, 196. [Google Scholar] [CrossRef] [Green Version]
- Mellors, R.C.; Heimer, R.; Corcos, J.; Korngold, L. Cellular origin of rheumatoid factor. J. Exp. Med. 1959, 110, 875–886. [Google Scholar] [CrossRef]
- Imai, Y.; Sato, T.; Yamakawa, M.; Kasajima, T.; Suda, A.; Watanabe, Y. A morphological and immunohistochemical study of lymphoid germinal centers in synovial and lymph node tissues from rheumatoid arthritis patients with special reference to complement components and their receptors. Acta Pathol. Jpn. 1989, 39, 127–134. [Google Scholar] [CrossRef]
- Randen, I.; Thompson, K.M.; Natvig, J.B.; Førre, O.; Waalen, K. Human monoclonal rheumatoid factors derived from the polyclonal repertoire of rheumatoid synovial tissue: Production and characterization. Clin. Exp. Immunol. 1989, 78, 13–18. [Google Scholar]
- Dahdah, A.; Habir, K.; Nandakumar, K.S.; Saxena, A.; Xu, B.; Holmdahl, R.; Malin, S. Germinal Center B Cells Are Essential for Collagen-Induced Arthritis. Arthritis Rheumatol. 2018, 70, 193–203. [Google Scholar] [CrossRef] [Green Version]
- Louis, C.; Ngo, D.; D’Silva, D.B.; Hansen, J.; Phillipson, L.; Jousset, H.; Novello, P.; Segal, D.; Lawlor, K.E.; Burns, C.J.; et al. Therapeutic Effects of a TANK-Binding Kinase 1 Inhibitor in Germinal Center-Driven Collagen-Induced Arthritis. Arthritis Rheumatol. 2019, 71, 50–62. [Google Scholar] [CrossRef] [Green Version]
- Han, S.; Cao, S.; Bheekha-Escura, R.; Zheng, B. Germinal center reaction in the joints of mice with collagen-induced arthritis: An animal model of lymphocyte activation and differentiation in arthritis joints. Arthritis Rheum. 2001, 44, 1438–1443. [Google Scholar] [CrossRef]
- Hou, L.; Block, K.E.; Huang, H. Artesunate abolishes germinal center B cells and inhibits autoimmune arthritis. PLoS ONE 2014, 9, e104762. [Google Scholar] [CrossRef]
- Wang, J.H.; New, J.S.; Xie, S.; Yang, P.; Wu, Q.; Li, J.; Luo, B.; Ding, Y.; Druey, K.M.; Hsu, H.-C.; et al. Extension of the germinal center stage of B cell development promotes autoantibodies in BXD2 mice. Arthritis Rheum. 2013, 65, 2703–2712. [Google Scholar] [CrossRef] [Green Version]
- Guo, W.; Smith, D.; Aviszus, K.; Detanico, T.; Heiser, R.A.; Wysocki, L.J. Somatic hypermutation as a generator of antinuclear antibodies in a murine model of systemic autoimmunity. J. Exp. Med. 2010, 207, 2225–2237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ray, S.K.; Putterman, C.; Diamond, B. Pathogenic autoantibodies are routinely generated during the response to foreign antigen: A paradigm for autoimmune disease. Proc. Natl. Acad. Sci. USA 1996, 93, 2019–2024. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Yu, Y.; Yue, Y.; Liao, H.; Xie, W.; Thai, J.; Mikuls, T.R.; Thiele, G.M.; Duryee, M.J.; Sayles, H.; et al. Autoantibodies from Single Circulating Plasmablasts React with Citrullinated Antigens and Porphyromonas gingivalis in Rheumatoid Arthritis. Arthritis Rheumatol. 2016, 68, 614–626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.; Hsu, H.C.; Chen, J.; Grizzle, W.E.; Chatham, W.W.; Stockard, C.R.; Wu, Q.; Yang, P.A.; Holers, V.M.; Mountz, J.D. Increased expression of activation-induced cytidine deaminase is associated with anti-CCP and rheumatoid factor in rheumatoid arthritis. Scand. J. Immunol. 2009, 70, 309–316. [Google Scholar] [CrossRef] [Green Version]
- Bugatti, S.; Manzo, A.; Vitolo, B.; Fusetti, C.; Humby, F.; Caporali, R.; Pitzalis, C.; Montecucco, C. B cell distribution and activation-induced cytidine deaminase expression in rheumatoid synovitis: Clinical and bio-molecular correlates. J. Ann. Rheum. Dis. 2011, 70, A55. [Google Scholar] [CrossRef] [Green Version]
- Sabouri, Z.; Schofield, P.; Horikawa, K.; Spierings, E.; Kipling, D.; Randall, K.L.; Langley, D.; Roome, B.; Vazquez-Lombardi, R.; Rouet, R.; et al. Redemption of autoantibodies on anergic B cells by variable-region glycosylation and mutation away from self-reactivity. Proc. Natl. Acad. Sci. USA 2014, 111, E2567–E2575. [Google Scholar] [CrossRef] [Green Version]
- Reed, J.H.; Jackson, J.; Christ, D.; Goodnow, C.C. Clonal redemption of autoantibodies by somatic hypermutation away from self-reactivity during human immunization. J. Exp. Med. 2016, 213, 1255–1265. [Google Scholar] [CrossRef] [Green Version]
- Weber, J.P.; Fuhrmann, F.; Feist, R.K.; Lahmann, A.; Al Baz, M.S.; Gentz, L.-J.; Vu Van, D.; Mages, H.W.; Haftmann, C.; Riedel, R.; et al. ICOS maintains the T follicular helper cell phenotype by down-regulating Krüppel-like factor 2. J. Exp. Med. 2015, 212, 217–233. [Google Scholar] [CrossRef] [Green Version]
- Meyer-Hermann, M.E.; Maini, P.K.; Iber, D. An analysis of B cell selection mechanisms in germinal centers. Math. Med. Biol. A J. IMA 2006, 23, 255–277. [Google Scholar] [CrossRef]
- Cao, G.; Chi, S.; Wang, X.; Sun, J.; Zhang, Y. CD4+CXCR5+PD-1+ T Follicular Helper Cells Play a Pivotal Role in the Development of Rheumatoid Arthritis. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2019, 25, 3032–3040. [Google Scholar] [CrossRef] [PubMed]
- Chevalier, N.; Macia, L.; Tan, J.K.; Mason, L.J.; Robert, R.; Thorburn, A.N.; Wong, C.H.; Tsai, L.M.; Bourne, K.; Brink, R.; et al. The Role of Follicular Helper T Cell Molecules and Environmental Influences in Autoantibody Production and Progression to Inflammatory Arthritis in Mice. Arthritis Rheumatol. 2016, 68, 1026–1038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moschovakis, G.L.; Bubke, A.; Friedrichsen, M.; Falk, C.S.; Feederle, R.; Forster, R. T cell specific Cxcr5 deficiency prevents rheumatoid arthritis. Sci. Rep. 2017, 7, 8933. [Google Scholar] [CrossRef] [PubMed]
- Jang, E.; Cho, S.H.; Park, H.; Paik, D.J.; Kim, J.M.; Youn, J. A positive feedback loop of IL-21 signaling provoked by homeostatic CD4+CD25- T cell expansion is essential for the development of arthritis in autoimmune K/BxN mice. J. Immunol. 2009, 182, 4649–4656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morita, R.; Schmitt, N.; Bentebibel, S.-E.; Ranganathan, R.; Bourdery, L.; Zurawski, G.; Foucat, E.; Dullaers, M.; Oh, S.; Sabzghabaei, N.; et al. Human blood CXCR5(+)CD4(+) T cells are counterparts of T follicular cells and contain specific subsets that differentially support antibody secretion. Immunity 2011, 34, 108–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crotty, S. Follicular helper CD4 T cells (TFH). Annu. Rev. Immunol. 2011, 29, 621–663. [Google Scholar] [CrossRef]
- Arroyo-Villa, I.; Bautista-Caro, M.-B.; Balsa, A.; Aguado-Acín, P.; Bonilla-Hernán, M.-G.; Plasencia, C.; Villalba, A.; Nuño, L.; Puig-Kröger, A.; Martín-Mola, E.; et al. Constitutively altered frequencies of circulating follicullar helper T cell counterparts and their subsets in rheumatoid arthritis. Arthritis Res. Ther. 2014, 16, 500. [Google Scholar] [CrossRef] [Green Version]
- Zhang, N.; Zhao, P.; Shrestha, A.; Zhang, L.; Qu, Z.; Liu, M.; Zhang, S.; Jiang, Y. A higher frequency of CD4+CXCR5+ T follicular helper cells in adult patients with minimal change disease. BioMed. Res. Int. 2014, 2014, 836157. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Shan, Y.; Jiang, Z.; Feng, J.; Li, C.; Ma, L.; Jiang, Y. High frequencies of activated B cells and T follicular helper cells are correlated with disease activity in patients with new-onset rheumatoid arthritis. Clin. Exp. Immunol. 2013, 174, 212–220. [Google Scholar] [CrossRef]
- Niu, Q.; Huang, Z.C.; Wu, X.J.; Jin, Y.X.; An, Y.F.; Li, Y.M.; Xu, H.; Yang, B.; Wang, L.L. Enhanced IL-6/phosphorylated STAT3 signaling is related to the imbalance of circulating T follicular helper/T follicular regulatory cells in patients with rheumatoid arthritis. Arthritis Res. Ther. 2018, 20, 200. [Google Scholar] [CrossRef] [Green Version]
- Zhou, H.; Hu, B.; Zhaopeng, Z.; Liu, J.; Zhong, Q.; Fan, Y.; Li, L. Elevated circulating T cell subsets and cytokines expression in patients with rheumatoid arthritis. Clin. Rheumatol. 2019, 38, 1831–1839. [Google Scholar] [CrossRef]
- Nakayamada, S.; Kubo, S.; Yoshikawa, M.; Miyazaki, Y.; Yunoue, N.; Iwata, S.; Miyagawa, I.; Hirata, S.; Nakano, K.; Saito, K.; et al. Differential effects of biological DMARDs on peripheral immune cell phenotypes in patients with rheumatoid arthritis. Rheumatology 2018, 57, 164–174. [Google Scholar] [CrossRef] [Green Version]
- Su, R.; Wang, Y.; Hu, F.; Li, B.; Guo, Q.; Zheng, X.; Liu, Y.; Gao, C.; Li, X.; Wang, C. Altered Distribution of Circulating T Follicular Helper-Like Cell Subsets in Rheumatoid Arthritis Patients. Front. Med. 2021, 8, 1024. [Google Scholar] [CrossRef]
- Wang, X.; Yang, C.; Xu, F.; Qi, L.; Wang, J.; Yang, P. Imbalance of circulating Tfr/Tfh ratio in patients with rheumatoid arthritis. Clin. Exp. Med. 2019, 19, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Macdonald, A.J.; Cerosaletti, K.; Chen, J.; Nguyen, T.-S.; Posso, S.; Marchesini, G.; Abashian, M.; Martin, A.P.; De Rosa, D.; Samanta, T.; et al. OP0264 Relative Frequencies of Circulating T Follicular Helper and T Follicular Regulatory Cells in Autoimmune Patients and Healthy Control Donors and The Effect of Disease Modulating Therapy. J. Ann. Rheum. Dis. 2016, 75, 158. [Google Scholar] [CrossRef]
- Liu, C.; Wang, D.; Lu, S.; Xu, Q.; Zhao, L.; Zhao, J.; Song, Y.; Wang, H. Increased Circulating Follicular Treg Cells Are Associated with Lower Levels of Autoantibodies in Patients with Rheumatoid Arthritis in Stable Remission. Arthritis Rheumatol. 2018, 70, 711–721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sage, P.T.; Sharpe, A.H. T follicular regulatory cells in the regulation of B cell responses. Trends Immunol. 2015, 36, 410–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, Y.; Tong, J.; Wang, S. Are Follicular Regulatory T Cells Involved in Autoimmune Diseases? Front. Immunol. 2017, 8, 1790. [Google Scholar] [CrossRef] [Green Version]
- Linterman, M.A.; Pierson, W.; Lee, S.K.; Kallies, A.; Kawamoto, S.; Rayner, T.F.; Srivastava, M.; Divekar, D.P.; Beaton, L.; Hogan, J.J.; et al. Foxp3+ follicular regulatory T cells control the germinal center response. Nat. Med. 2011, 17, 975–982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, Y.; Tanaka, S.; Chu, F.; Nurieva, R.I.; Martinez, G.J.; Rawal, S.; Wang, Y.H.; Lim, H.; Reynolds, J.M.; Zhou, X.H.; et al. Follicular regulatory T cells expressing Foxp3 and Bcl-6 suppress germinal center reactions. Nat. Med. 2011, 17, 983–988. [Google Scholar] [CrossRef]
- Fonseca, V.R.; Agua-Doce, A.; Maceiras, A.R.; Pierson, W.; Ribeiro, F.; Romao, V.C.; Pires, A.R.; da Silva, S.L.; Fonseca, J.E.; Sousa, A.E.; et al. Human blood Tfr cells are indicators of ongoing humoral activity not fully licensed with suppressive function. Sci. Immunol. 2017, 2. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Zou, L.; Liu, Y.-C. T follicular helper cells, T follicular regulatory cells and autoimmunity. Int. Immunol. 2016, 28, 173–179. [Google Scholar] [CrossRef] [Green Version]
- Laidlaw, B.J.; Lu, Y.; Amezquita, R.A.; Weinstein, J.S.; Vander Heiden, J.A.; Gupta, N.T.; Kleinstein, S.H.; Kaech, S.M.; Craft, J. Interleukin-10 from CD4(+) follicular regulatory T cells promotes the germinal center response. Sci. Immunol. 2017, 2. [Google Scholar] [CrossRef] [Green Version]
- Ding, Y.; Li, J.; Yang, P.; Luo, B.; Wu, Q.; Zajac, A.J.; Wildner, O.; Hsu, H.-C.; Mountz, J.D. Interleukin-21 Promotes Germinal Center Reaction by Skewing the Follicular Regulatory T Cell to Follicular Helper T Cell Balance in Autoimmune BXD2 Mice. Arthritis Rheumatol. 2014, 66, 2601–2612. [Google Scholar] [CrossRef]
- Fonseca, V.R.; Romao, V.C.; Agua-Doce, A.; Santos, M.; Lopez-Presa, D.; Ferreira, A.C.; Fonseca, J.E.; Graca, L. The Ratio of Blood T Follicular Regulatory Cells to T Follicular Helper Cells Marks Ectopic Lymphoid Structure Formation While Activated Follicular Helper T Cells Indicate Disease Activity in Primary Sjogren’s Syndrome. Arthritis Rheumatol. 2018, 70, 774–784. [Google Scholar] [CrossRef]
- Shan, Y.; Qi, C.; Zhao, J.; Liu, Y.; Gao, H.; Zhao, D.; Ding, F.; Wang, J.; Jiang, Y. Higher frequency of peripheral blood follicular regulatory T cells in patients with new onset ankylosing spondylitis. Clin. Exp. Pharmacol. Physiol. 2015, 42, 154–161. [Google Scholar] [CrossRef]
- Fu, W.; Liu, X.; Lin, X.; Feng, H.; Sun, L.; Li, S.; Chen, H.; Tang, H.; Lu, L.; Jin, W.; et al. Deficiency in T follicular regulatory cells promotes autoimmunity. J. Exp. Med. 2018, 215, 815–825. [Google Scholar] [CrossRef] [Green Version]
- Miyabe, C.; Miyabe, Y.; Strle, K.; Kim, N.D.; Stone, J.H.; Luster, A.D.; Unizony, S. An expanded population of pathogenic regulatory T cells in giant cell arteritis is abrogated by IL-6 blockade therapy. Ann. Rheum. Dis. 2017, 76, 898–905. [Google Scholar] [CrossRef]
- Xu, X.; Shi, Y.; Cai, Y.; Zhang, Q.; Yang, F.; Chen, H.; Gu, Y.; Zhang, M.; Yu, L.; Yang, T. Inhibition of increased circulating Tfh cell by anti-CD20 monoclonal antibody in patients with type 1 diabetes. PLoS ONE 2013, 8, e79858. [Google Scholar] [CrossRef] [Green Version]
- Verstappen, G.M.; Meiners, P.M.; Corneth, O.B.J.; Visser, A.; Arends, S.; Abdulahad, W.H.; Hendriks, R.W.; Vissink, A.; Kroese, F.G.M.; Bootsma, H. Attenuation of Follicular Helper T Cell-Dependent B Cell Hyperactivity by Abatacept Treatment in Primary Sjogren’s Syndrome. Arthritis Rheumatol. 2017, 69, 1850–1861. [Google Scholar] [CrossRef]
- Edner, N.M.; Heuts, F.; Thomas, N.; Wang, C.J.; Petersone, L.; Kenefeck, R.; Kogimtzis, A.; Ovcinnikovs, V.; Ross, E.M.; Ntavli, E.; et al. Follicular helper T cell profiles predict response to costimulation blockade in type 1 diabetes. Nat. Immunol. 2020, 21, 1244–1255. [Google Scholar] [CrossRef]
- Glatigny, S.; Höllbacher, B.; Motley, S.J.; Tan, C.; Hundhausen, C.; Buckner, J.H.; Smilek, D.; Khoury, S.J.; Ding, L.; Qin, T.; et al. Abatacept Targets T Follicular Helper and Regulatory T Cells, Disrupting Molecular Pathways That Regulate Their Proliferation and Maintenance. J. Immunol. 2019, 202, ji1801425. [Google Scholar] [CrossRef] [Green Version]
- Schmiel, S.E.; Kalekar, L.A.; Zhang, N.; Blankespoor, T.W.; Robinson, L.J.; Mueller, D.L. Adenosine 2a receptor signals block autoimmune arthritis by inhibiting pathogenic germinal center T follicular helper cells. Arthritis Rheumatol. 2018, 71, 773–783. [Google Scholar] [CrossRef]
- Klimatcheva, E.; Pandina, T.; Reilly, C.; Torno, S.; Bussler, H.; Scrivens, M.; Jonason, A.; Mallow, C.; Doherty, M.; Paris, M.; et al. CXCL13 antibody for the treatment of autoimmune disorders. BMC Immunol. 2015, 16, 6. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Shen, W.; Kong, K.; Liu, Z. Interleukin-21 Induces T-cell Activation and Proinflammatory Cytokine Secretion in Rheumatoid Arthritis. Scand. J. Imunol. 2006, 64, 515–522. [Google Scholar] [CrossRef]
- Young, D.A.; Hegen, M.; Ma, H.L.; Whitters, M.J.; Albert, L.M.; Lowe, L.; Senices, M.; Wu, P.W.; Sibley, B.; Leathurby, Y.; et al. Blockade of the interleukin-21/interleukin-21 receptor pathway ameliorates disease in animal models of rheumatoid arthritis. Arthritis Rheum. 2007, 56, 1152–1163. [Google Scholar] [CrossRef] [PubMed]
- Roeleveld, D.M.; Marijnissen, R.J.; Walgreen, B.; Helsen, M.M.; van den Bersselaar, L.; van de Loo, F.A.; van Lent, P.L.; van der Kraan, P.M.; van den Berg, W.B.; Koenders, M.I. Higher efficacy of anti-IL-6/IL-21 combination therapy compared to monotherapy in the induction phase of Th17-driven experimental arthritis. PLoS ONE 2017, 12, e0171757. [Google Scholar] [CrossRef] [PubMed]
- Andersson, A.K.; Feldmann, M.; Brennan, F.M. Neutralizing IL-21 and IL-15 inhibits pro-inflammatory cytokine production in rheumatoid arthritis. Scand. J. Immunol. 2008, 68, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Kwok, S.K.; Cho, M.L.; Park, M.K.; Oh, H.J.; Park, J.S.; Her, Y.M.; Lee, S.Y.; Youn, J.; Ju, J.H.; Park, K.S.; et al. Interleukin-21 promotes osteoclastogenesis in humans with rheumatoid arthritis and in mice with collagen-induced arthritis. Arthritis Rheum. 2012, 64, 740–751. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; de Leur, K.; Hendriks, R.W.; van der Laan, L.J.W.; Shi, Y.; Wang, L.; Baan, C.C. T Follicular Helper Cells as a New Target for Immunosuppressive Therapies. Front. Immunol. 2017, 8, 1510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daoussis, D.; Andonopoulos, A.P.; Liossis, S.-N.C. Targeting CD40L: A promising therapeutic approach. Clin. Diagn. Lab. Immunol. 2004, 11, 635–641. [Google Scholar] [CrossRef] [Green Version]
- Aruffo, A.; Farrington, M.; Hollenbaugh, D.; Li, X.; Milatovich, A.; Nonoyama, S.; Bajorath, J.; Grosmaire, L.S.; Stenkamp, R.; Neubauer, M.; et al. The CD40 ligand, gp39, is defective in activated T cells from patients with X-linked hyper-IgM syndrome. Cell 1993, 72, 291–300. [Google Scholar] [CrossRef]
- Berner, B.; Wolf, G.; Hummel, K.M.; Muller, G.A.; Reuss-Borst, M.A. Increased expression of CD40 ligand (CD154) on CD4+ T cells as a marker of disease activity in rheumatoid arthritis. Ann. Rheum. Dis. 2000, 59, 190–195. [Google Scholar] [CrossRef] [Green Version]
- MacDonald, K.P.; Nishioka, Y.; Lipsky, P.E.; Thomas, R. Functional CD40 ligand is expressed by T cells in rheumatoid arthritis. J. Clin. Investig. 1997, 100, 2404–2414. [Google Scholar] [CrossRef] [Green Version]
- Kyburz, D.; Corr, M.; Brinson, D.C.; Von Damm, A.; Tighe, H.; Carson, D.A. Human rheumatoid factor production is dependent on CD40 signaling and autoantigen. J. Immunol. 1999, 163, 3116–3122. [Google Scholar]
- Tellander, A.C.; Michaelsson, E.; Brunmark, C.; Andersson, M. Potent adjuvant effect by anti-CD40 in collagen-induced arthritis. Enhanced disease is accompanied by increased production of collagen type-II reactive IgG2a and IFN-gamma. J. Autoimmun. 2000, 14, 295–302. [Google Scholar] [CrossRef]
- Kyburz, D.; Carson, D.A.; Corr, M. The role of CD40 ligand and tumor necrosis factor alpha signaling in the transgenic K/BxN mouse model of rheumatoid arthritis. Arthritis Rheum. 2000, 43, 2571–2577. [Google Scholar] [CrossRef] [Green Version]
- Visvanathan, S.; Daniluk, S.; Ptaszyński, R.; Müller-Ladner, U.; Ramanujam, M.; Rosenstock, B.; Eleftheraki, A.G.; Vinisko, R.; Petříková, A.; Kellner, H.; et al. Effects of BI 655064, an antagonistic anti-CD40 antibody, on clinical and biomarker parameters in patients with active rheumatoid arthritis: A randomised, double-blind, placebo-controlled, phase IIa study. J. Ann. Rheum. Dis. 2019, 78, 754–760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karnell, J.L.; Albulescu, M.; Drabic, S.; Wang, L.; Moate, R.; Baca, M.; Oganesyan, V.; Gunsior, M.; Thisted, T.; Yan, L.; et al. A CD40L-targeting protein reduces autoantibodies and improves disease activity in patients with autoimmunity. J. Sci. Transl. Med. 2019, 11, eaar6584. [Google Scholar] [CrossRef] [PubMed]
- Croft, M. Control of immunity by the TNFR-related molecule OX40 (CD134). Ann. Rev. Immunol. 2010, 28, 57–78. [Google Scholar] [CrossRef] [Green Version]
- Kurata, I.; Matsumoto, I.; Ohyama, A.; Osada, A.; Ebe, H.; Kawaguchi, H.; Kaneko, S.; Kondo, Y.; Tsuboi, H.; Tomioka, A.; et al. Potential involvement of OX40 in the regulation of autoantibody sialylation in arthritis. Ann. Rheum. Dis. 2019, 78, 1488–1496. [Google Scholar] [CrossRef] [PubMed]
- Tahiliani, V.; Hutchinson, T.E.; Abboud, G.; Croft, M.; Salek-Ardakani, S. OX40 Cooperates with ICOS To Amplify Follicular Th Cell Development and Germinal Center Reactions during Infection. J. Immunol. 2017, 198, 218–228. [Google Scholar] [CrossRef]
- Gwyer Findlay, E.; Danks, L.; Madden, J.; Cavanagh, M.M.; McNamee, K.; McCann, F.; Snelgrove, R.J.; Shaw, S.; Feldmann, M.; Taylor, P.C.; et al. OX40L blockade is therapeutic in arthritis, despite promoting osteoclastogenesis. Proc. Natl. Acad. Sci. USA 2014, 111, 2289–2294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giacomelli, R.; Passacantando, A.; Perricone, R.; Parzanese, I.; Rascente, M.; Minisola, G.; Tonietti, G. T lymphocytes in the synovial fluid of patients with active rheumatoid arthritis display CD134-OX40 surface antigen. Clin. Exp. Rheumatol. 2001, 19, 317–320. [Google Scholar] [PubMed]
- Laustsen, J.K.; Rasmussen, T.K.; Stengaard-Pedersen, K.; Hørslev-Petersen, K.; Hetland, M.L.; Østergaard, M.; Junker, P.; Hvid, M.; Deleuran, B. Soluble OX40L is associated with presence of autoantibodies in early rheumatoid arthritis. Arthritis Res. Ther. 2014, 16, 474. [Google Scholar] [CrossRef] [Green Version]
- Yoshioka, T.; Nakajima, A.; Akiba, H.; Ishiwata, T.; Asano, G.; Yoshino, S.; Yagita, H.; Okumura, K. Contribution of OX40/OX40 ligand interaction to the pathogenesis of rheumatoid arthritis. Eur. J. Immunol. 2000, 30, 2815–2823. [Google Scholar] [CrossRef]
- Horai, R.; Nakajima, A.; Habiro, K.; Kotani, M.; Nakae, S.; Matsuki, T.; Nambu, A.; Saijo, S.; Kotaki, H.; Sudo, K.; et al. TNF-alpha is crucial for the development of autoimmune arthritis in IL-1 receptor antagonist-deficient mice. J. Clin. Investig. 2004, 114, 1603–1611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Platt, A.M.; Gibson, V.B.; Patakas, A.; Benson, R.A.; Nadler, S.G.; Brewer, J.M.; McInnes, I.B.; Garside, P. Abatacept Limits Breach of Self-Tolerance in a Murine Model of Arthritis via Effects on the Generation of T Follicular Helper Cells. J. Immunol. 2010, 185, 1558–1567. [Google Scholar] [CrossRef] [Green Version]
- Linterman, M.A.; Rigby, R.J.; Wong, R.; Silva, D.; Withers, D.; Anderson, G.; Verma, N.K.; Brink, R.; Hutloff, A.; Goodnow, C.C.; et al. Roquin differentiates the specialized functions of duplicated T cell costimulatory receptor genes CD28 and ICOS. Immunity 2009, 30, 228–241. [Google Scholar] [CrossRef] [Green Version]
- Bossaller, L.; Burger, J.; Draeger, R.; Grimbacher, B.; Knoth, R.; Plebani, A.; Durandy, A.; Baumann, U.; Schlesier, M.; Welcher, A.A.; et al. ICOS Deficiency Is Associated with a Severe Reduction of CXCR5+CD4 Germinal Center Th Cells. J. Immunol. 2006, 177, 4927–4932. [Google Scholar] [CrossRef] [Green Version]
- Cheng, L.E.; Amoura, Z.; Cheah, B.; Hiepe, F.; Sullivan, B.A.; Zhou, L.; Arnold, G.E.; Tsuji, W.H.; Merrill, J.T.; Chung, J.B. Brief Report: A Randomized, Double-Blind, Parallel-Group, Placebo-Controlled, Multiple-Dose Study to Evaluate AMG 557 in Patients with Systemic Lupus Erythematosus and Active Lupus Arthritis. Arthritis Rheumatol. 2018, 70, 1071–1076. [Google Scholar] [CrossRef] [PubMed]
- Iwai, H.; Kozono, Y.; Hirose, S.; Akiba, H.; Yagita, H.; Okumura, K.; Kohsaka, H.; Miyasaka, N.; Azuma, M. Amelioration of collagen-induced arthritis by blockade of inducible costimulator-B7 homologous protein costimulation. J. Immunol. 2002, 169, 4332–4339. [Google Scholar] [CrossRef]
- Dwyer, R.; Kovaleva, M.; Zhang, J.; Steven, J.; Cummins, E.; Luxenberg, D.; Darmanin-Sheehan, A.; Carvalho, M.F.; Whitters, M.; Saunders, K.; et al. Anti-ICOSL New Antigen Receptor Domains Inhibit T Cell Proliferation and Reduce the Development of Inflammation in the Collagen-Induced Mouse Model of Rheumatoid Arthritis. J. Immunol. Res. 2018, 2018, 13. [Google Scholar] [CrossRef]
- Frey, O.; Meisel, J.; Hutloff, A.; Bonhagen, K.; Bruns, L.; Kroczek, R.A.; Morawietz, L.; Kamradt, T. Inducible costimulator (ICOS) blockade inhibits accumulation of polyfunctional T helper 1/T helper 17 cells and mitigates autoimmune arthritis. J. Ann. Rheum. Dis. 2010, 69, 1495–1501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holers, V.M.; Demoruelle, M.K.; Kuhn, K.A.; Buckner, J.H.; Robinson, W.H.; Okamoto, Y.; Norris, J.M.; Deane, K.D. Rheumatoid arthritis and the mucosal origins hypothesis: Protection turns to destruction. Nat. Rev. Rheumatol. 2018, 14, 542–557. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zou, Q.; Zeng, B.; Fang, Y.; Wei, H. Analysis of fecal Lactobacillus community structure in patients with early rheumatoid arthritis. Curr. Microbiol. 2013, 67, 170–176. [Google Scholar] [CrossRef]
- Vaahtovuo, J.; Munukka, E.; Korkeamaki, M.; Luukkainen, R.; Toivanen, P. Fecal microbiota in early rheumatoid arthritis. J. Rheumatol. 2008, 35, 1500–1505. [Google Scholar]
- Chen, H.; Zhang, Y.; Ye, A.Y.; Du, Z.; Xu, M.; Lee, C.S.; Hwang, J.K.; Kyritsis, N.; Ba, Z.; Neuberg, D.; et al. BCR selection and affinity maturation in Peyer’s patch germinal centres. Nature 2020, 582, 421–425. [Google Scholar] [CrossRef] [PubMed]
- Gerlag, D.M.; Raza, K.; van Baarsen, L.G.M.; Brouwer, E.; Buckley, C.D.; Burmester, G.R.; Gabay, C.; Catrina, A.I.; Cope, A.P.; Cornelis, F.; et al. EULAR recommendations for terminology and research in individuals at risk of rheumatoid arthritis: Report from the Study Group for Risk Factors for Rheumatoid Arthritis. J. Ann. Rheum. Dis. 2012, 71, 638–641. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Tfh Cells in RA | Findings |
|---|---|
| Arroyo-Villa et al. [65] | RA patients with active disease have a higher frequency of Tfh cells and a higher Tfh/Tfr ratio resulting from lower Tfr frequencies |
| Zhang et al. [66] | Increased frequencies of Tfh cells and IL-21 in RA patients which correlates positively with DAS28 |
| Wang et al. [67], Zhou et al. [69] | Increased frequencies of Tfh cells in newly diagnosed RA patients correlating with activated B cells and DAS28 |
| Niu et al. [68] | Elevated frequencies of Tfh cells, IL-21 and PD-1 in RA patients with active disease |
| Nakayamada et al. [70] | Higher proportions of Tfh cells in RA patients with active disease and, treatment with Abatacept reduces Tfh cell levels. |
| Su et al. [71] | Elevated frequencies of Tfh cells in RA patients compared to HCs |
| Tfr cells in RA | |
| Su et al. [71] | Decreased frequencies of Tfr cells in RA patients compared to HCs |
| Wang et al. [72] | Increased levels of Tfr cells in RA patients compared to HCs |
| Macdonald et al. [73] | Elevated percentages of Tfr cells in RA patients |
| Liu et al. [74] | Higher frequencies of Tfr and Tfr/Tfh cell ratio in RA patients with stable remission |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anang, D.C.; Balzaretti, G.; van Kampen, A.; de Vries, N.; Klarenbeek, P.L. The Germinal Center Milieu in Rheumatoid Arthritis: The Immunological Drummer or Dancer? Int. J. Mol. Sci. 2021, 22, 10514. https://doi.org/10.3390/ijms221910514
Anang DC, Balzaretti G, van Kampen A, de Vries N, Klarenbeek PL. The Germinal Center Milieu in Rheumatoid Arthritis: The Immunological Drummer or Dancer? International Journal of Molecular Sciences. 2021; 22(19):10514. https://doi.org/10.3390/ijms221910514
Chicago/Turabian StyleAnang, Dornatien C., Giulia Balzaretti, Antoine van Kampen, Niek de Vries, and Paul L. Klarenbeek. 2021. "The Germinal Center Milieu in Rheumatoid Arthritis: The Immunological Drummer or Dancer?" International Journal of Molecular Sciences 22, no. 19: 10514. https://doi.org/10.3390/ijms221910514
APA StyleAnang, D. C., Balzaretti, G., van Kampen, A., de Vries, N., & Klarenbeek, P. L. (2021). The Germinal Center Milieu in Rheumatoid Arthritis: The Immunological Drummer or Dancer? International Journal of Molecular Sciences, 22(19), 10514. https://doi.org/10.3390/ijms221910514