
PPAR-α Hypermethylation in the Hippocampus of Mice Exposed to Social Isolation Stress Is Associated with Enhanced Neuroinflammation and Aggressive Behavior



Abstract
:1. Introduction
2. Results
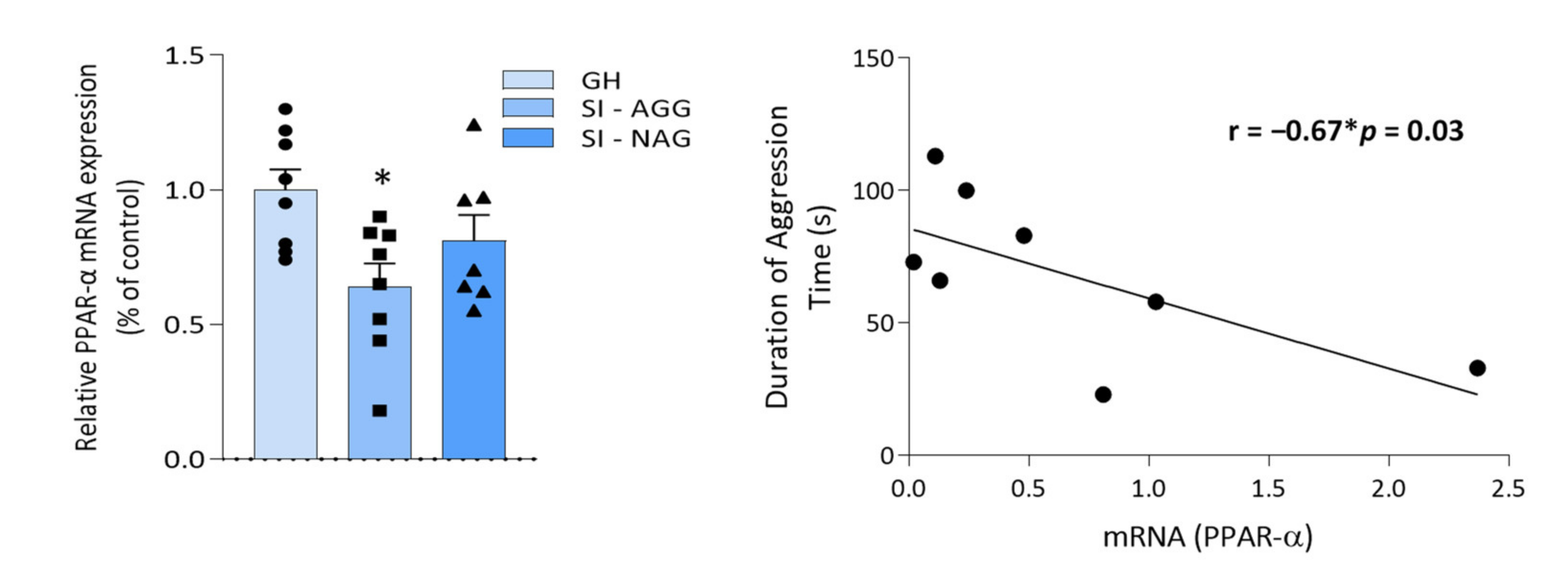
2.1. Decreased PPAR-α Expression in the Hippocampus of SI Mice Correlates with Aggressive Behavior
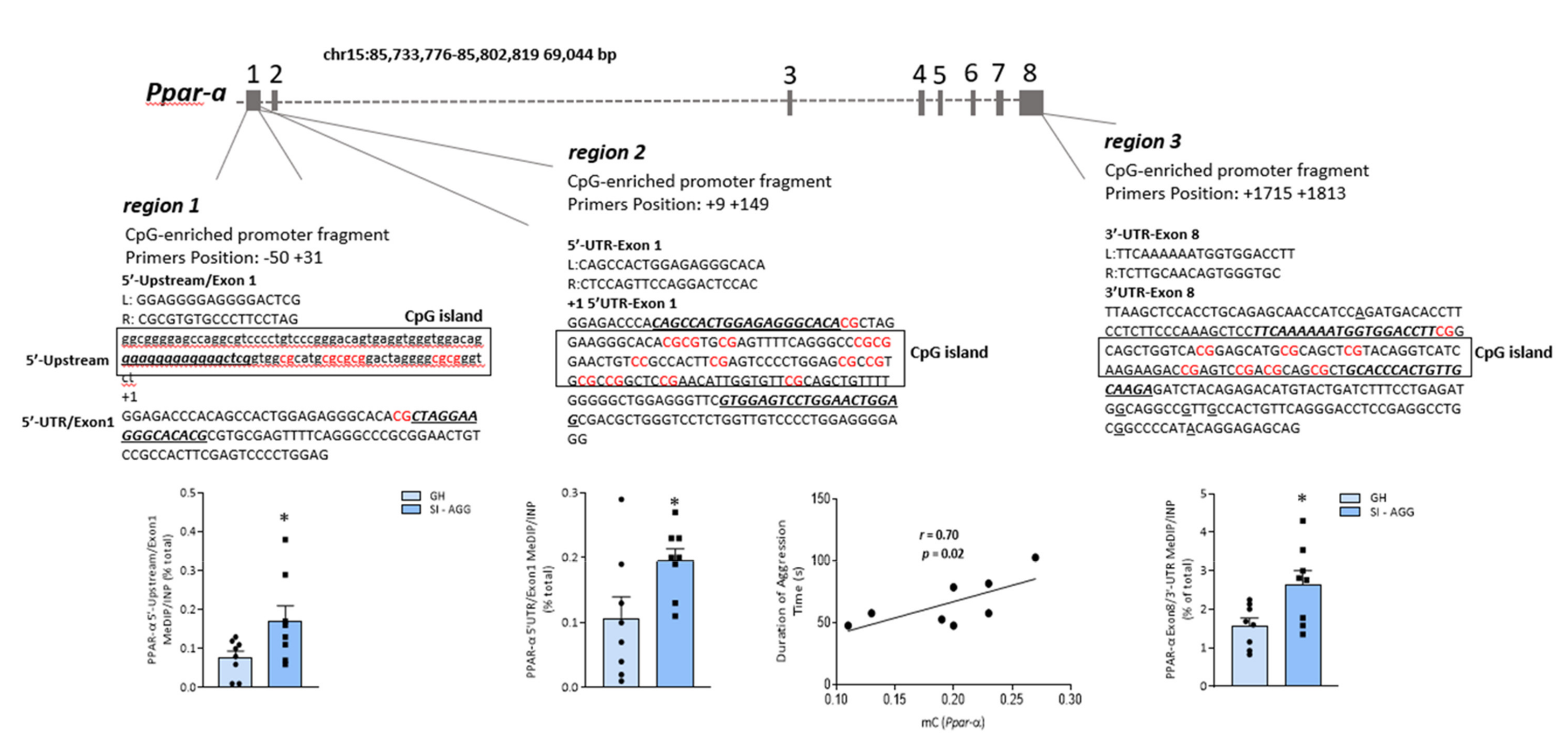
2.2. Social Isolation Stress Induces DNA Methylation of PPAR-α CpG-Rich Promoter Regions
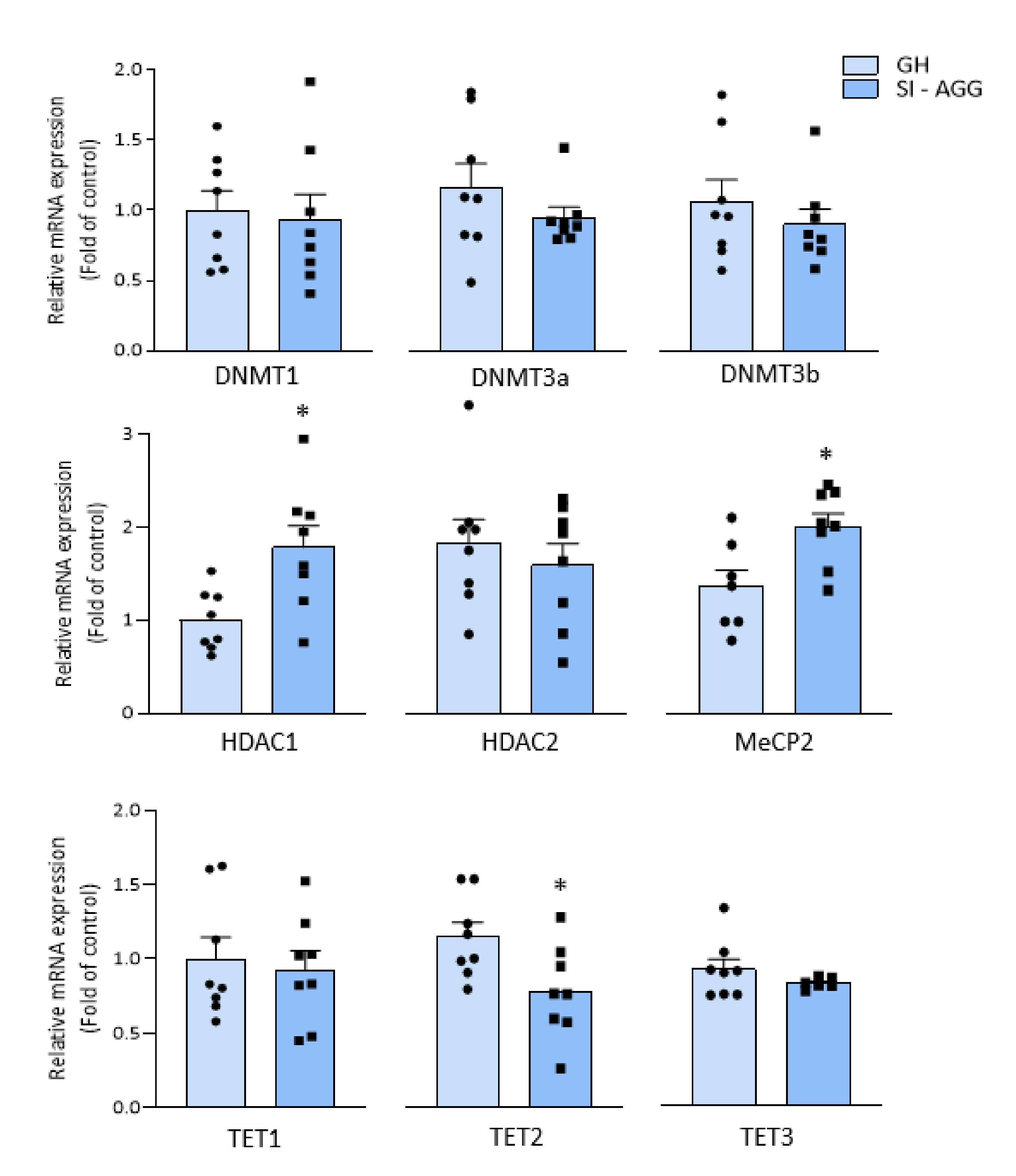
2.3. Altered Expression of Chromatin Remodeling Marks in the Hippocampus of SI Mice
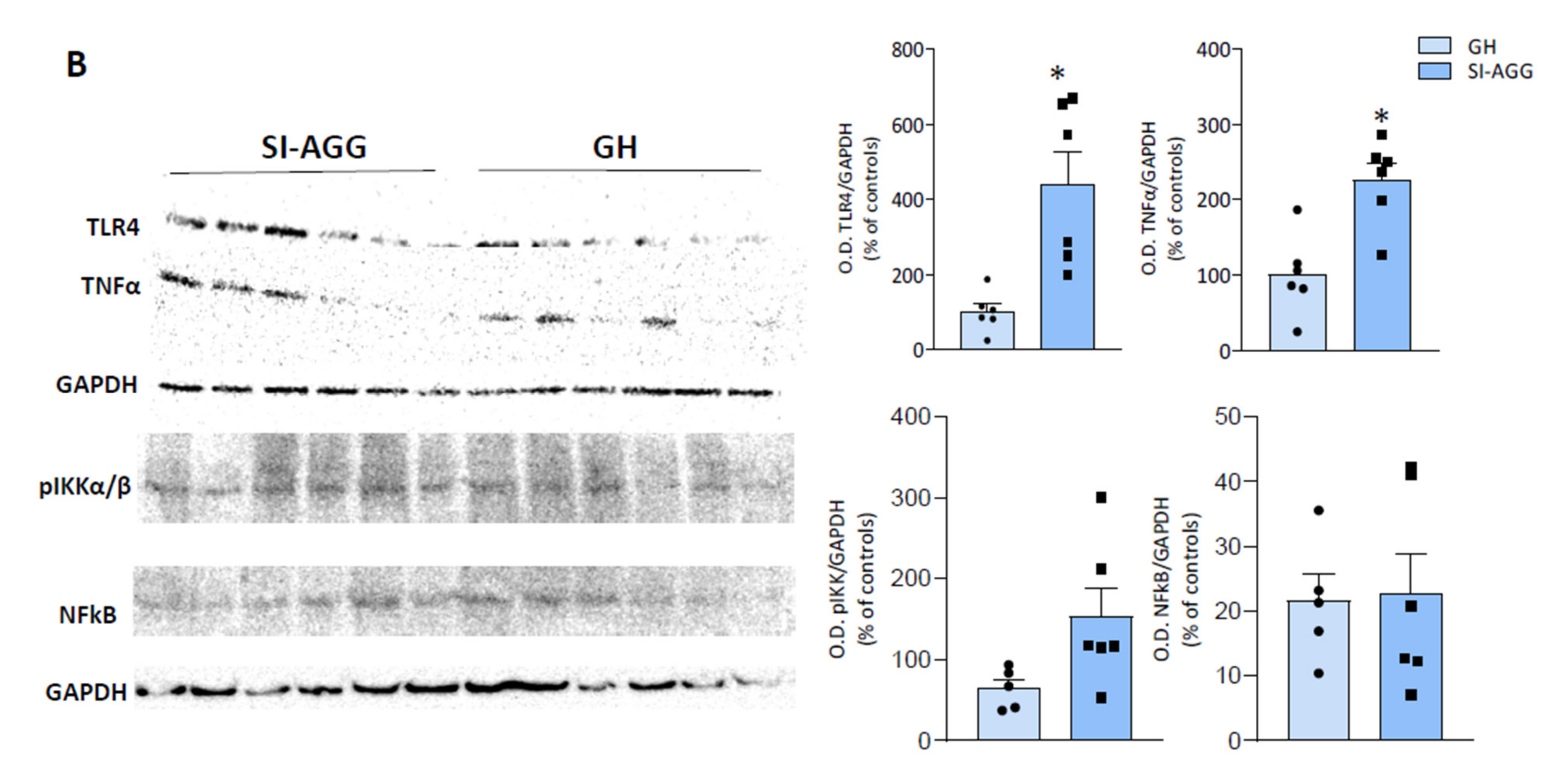
2.4. Increased Expression of Pro-Inflammatory Markers in the Hippocampus of SI Mice Is Associated with Aggression
2.5. Neurosteroid Levels in Aggressive and Non-Aggressive SI Mice
3. Discussion
3.1. Epigenetic Mechanisms Affecting PPAR-α Expression during Social Isolation
3.2. PPAR-α/Neurosteroid Downregulation and Increase in Pro-inflammatory Mediators in SI Mice
3.3. PPAR-α/TLR4/Allo Pathway as a Potential Biomarker and Pharmacological Target in Stress-Induced Behavior
4. Methods and Materials
4.1. Animals
4.2. Behavioral Analysis
4.3. Brain Neurosteroid Measurements
4.4. Real-Time Polymerase Chain Reaction (RT-PCR)
4.5. Western Blotting
4.6. PPAR-α Gene Methylation Analysis
4.7. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sher, L. The impact of the COVID-19 pandemic on suicide rates. QJM 2020, 113, 707–712. [Google Scholar] [CrossRef]
- Lieberman and Olfson in “Meeting the Mental Health Challenge of the COVID-19 Pandemic”. 2020. Available online: https://www.psychiatrictimes.com/coronavirus/meeting-mental-health-challenge-covid19-pandemic (accessed on 18 February 2021).
- Marel, C.; Mills, K.L.; Teesson, M. Substance use, mental disorders and COVID-19: A volatile mix. Curr. Opin. Psychiatry 2021, 34, 351–356. [Google Scholar] [CrossRef] [PubMed]
- Rice, T.; Sher, L. The Men’s Mental Health Perspective on Adolescent Suicide in the COVID-19 Era. Acta Neuropsychiatr. 2021, 33, 178–181. [Google Scholar] [CrossRef] [PubMed]
- Sher, L. Resilience as a focus of suicide research and prevention. Acta Psychiatr. Scand. 2019, 140, 169–180. [Google Scholar] [CrossRef] [PubMed]
- Calati, R.; Ferrari, C.; Brittner, M.; Oasi, O.; Olié, E.; Carvalho, A.F.; Courtet, P. Suicidal thoughts and behaviors and social isolation: A narrative review of the literature. J. Affect. Disord. 2019, 245, 653–667. [Google Scholar] [CrossRef] [PubMed]
- Horn, S.R.; Feder, A. Understanding Resilience and Preventing and Treating PTSD. Harv. Rev. Psychiatry 2018, 26, 158–174. [Google Scholar] [CrossRef]
- Wu, G.; Feder, A.; Cohen, H.; Kim, J.J.; Calderon, S.; Charney, D.S.; Mathé, A.A. Understanding resilience. Front. Behav. Neurosci. 2013, 7, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osório, C.; Probert, T.; Jones, E.; Young, A.H.; Robbins, I. Adapting to Stress: Understanding the Neurobiology of Resilience. Behav. Med. 2017, 43, 307–322. [Google Scholar] [CrossRef]
- Haglund, M.E.; Nestadt, P.S.; Cooper, N.S.; Southwick, S.M.; Charney, D.S. Psychobiological mechanisms of resilience: Relevance to prevention and treatment of stress-related psychopathology. Dev. Psychopathol. 2007, 19, 889–920. [Google Scholar] [CrossRef]
- Hori, H.; Kim, Y. Inflammation and post-traumatic stress disorder. Psychiatry Clin. Neurosci. 2019, 73, 143–153. [Google Scholar] [CrossRef] [Green Version]
- Köhler-Forsberg, O.; NLydholm, C.; Hjorthøj, C.; Nordentoft, M.; Mors, O.; Benros, M.E. Efficacy of anti-inflammatory treatment on major depressive disorder or depressive symptoms: Meta-analysis of clinical trials. Acta Psychiatr. Scand. 2019, 139, 404–419. [Google Scholar] [CrossRef] [PubMed]
- Moore, L.D.; Le, T.; Fan, G. DNA methylation and its basic function. Neuropsychopharmacology 2013, 38, 23–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinna, G.; Rasmusson, A.M. Ganaxolone improves behavioral deficits in a mouse model of post-traumatic stress disorder. Front. Cell. Neurosci. 2014, 8, 256. [Google Scholar] [CrossRef] [PubMed]
- Locci, A.; Pinna, G. Stimulation of Peroxisome Proliferator-Activated Receptor-α by N-Palmitoylethanolamine Engages Allopregnanolone Biosynthesis to Modulate Emotional Behavior. Biol. Psychiatry 2019, 85, 1036–1045. [Google Scholar] [CrossRef] [PubMed]
- Gvion, Y.; Horresh, N.; Levi-Belz, Y.; Fischel, T.; Treves, I.; Weiser, M.; David, H.S.; Stein-Reizer, O.; Apter, A. Aggression-impulsivity, mental pain, and communication difficulties in medically serious and medically non-serious suicide attempters. Compr. Psychiatry 2014, 55, 40–50. [Google Scholar] [CrossRef] [PubMed]
- Conner, K.R.; Swogger, M.T.; Houston, R.J. A test of the reactive aggression-suicidal behavior hypothesis: Is there a case for proactive aggression? J. Abnorm. Psychol. 2009, 118, 235–240. [Google Scholar] [CrossRef] [Green Version]
- Turecki, G. Dissecting the suicide phenotype: The role of impulsive-aggressive behaviours. J. Psychiatry Neurosci. 2005, 30, 398–408. [Google Scholar]
- Pibiri, F.; Nelson, M.; Guidotti, A.; Costa, E.; Pinna, G. Decreased corticolimbic allopregnanolone expression during social isolation enhances contextual fear: A model relevant for posttraumatic stress disorder. Proc. Natl. Acad. Sci. USA 2008, 105, 5567–5572. [Google Scholar] [CrossRef] [Green Version]
- Locci, A.; Pinna, G. Social isolation as a promising animal model of PTSD comorbid suicide: Neurosteroids and cannabinoids as possible treatment options. Prog. Neuropsychopharmacol. Biol. Psychiatry 2019, 92, 243–259. [Google Scholar] [CrossRef]
- Malkesman, O.; Pine, D.S.; Tragon, T.; Austin, D.R.; Henter, I.D.; Chen, G.; Manji, H.K. Animal models of suicide-trait-related behaviors. Trends Pharmacol. Sci. 2009, 30, 165–173. [Google Scholar] [CrossRef] [Green Version]
- Wahli, W.; Michalik, L. PPARs at the crossroads of lipid signaling and inflammation. Trends Endocrinol. Metab. 2012, 23, 351–363. [Google Scholar] [CrossRef]
- Pineda Torra, I.; Jamshidi, Y.; Flavell, D.M.; Fruchart, J.C.; Staels, B. Characterization of the human PPARalpha promoter: Identification of a functional nuclear receptor response element. Mol. Endocrinol. 2002, 16, 1013–1028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antunes, C.; Sousa, N.; Pinto, L.; Marques, C.J. TET enzymes in neurophysiology and brain function. Neurosci. Biobehav. Rev. 2019, 102, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Nierenberg, A.A.; Ghaznavi, S.A.; Sande Mathias, I.; Ellard, K.K.; Janos, J.A.; Sylvia, L.G. Peroxisome Proliferator-Activated Receptor Gamma Coactivator-1 Alpha as a Novel Target for Bipolar Disorder and Other Neuropsychiatric Disorders. Biol. Psychiatry 2018, 83, 761–769. [Google Scholar] [CrossRef]
- Wójtowicz, S.; Strosznajder, A.K.; Jeżyna, M.; Strosznajder, J.B. The Novel Role of PPAR Alpha in the Brain: Promising Target in Therapy of Alzheimer’s Disease and Other Neurodegenerative Disorders. Neurochem. Res. 2020, 45, 972–988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wada, Y.; Maekawa, M.; Ohnishi, T.; Balan, S.; Matsuoka, S.; Iwamoto, K.; Iwayama, Y.; Ohba, H.; Watanabe, A.; Hisano, Y.; et al. Peroxisome proliferator-activated receptor α as a novel therapeutic target for schizophrenia. EBioMedicine 2020, 62, 103130. [Google Scholar] [CrossRef]
- Matrisciano, F.; Pinna, G. PPAR and functional foods: Rationale for natural neurosteroid-based interventions for postpartum depression. Neurobiol. Stress 2020, 12, 100222. [Google Scholar] [CrossRef] [PubMed]
- Bougarne, N.; Weyers, B.; Desmet, S.J.; Deckers, J.; Ray, D.; Staels, B.; De Bosscher, K. Molecular Actions of PPAR-α in Lipid Metabolism and Inflammation. Endocr. Rev. 2018, 39, 760–802. [Google Scholar] [CrossRef] [Green Version]
- Desvergne, B.; Wahli, W. Peroxisome proliferator-activated receptors: Nuclear control of metabolism. Endocr. Rev. 1999, 20, 649–688. [Google Scholar] [CrossRef] [Green Version]
- Warren, J.S.; Oka, S.I.; Zablocki, D.; Sadoshima, J. Metabolic reprogramming via PPAR-α signaling in cardiac hypertrophy and failure: From metabolomics to epigenetics. Am. J. Physiol. Heart Circ. Physiol. 2017, 313, H584–H596. [Google Scholar] [CrossRef]
- Luo, Y.; Xie, C.; Brocker, C.N.; Fan, J.; Wu, X.; Feng, L.; Wang, Q.; Zhao, J.; Lu, D.; Tandon, M.; et al. Intestinal PPAR-α Protects Against Colon Carcinogenesis via Regulation of Methyltransferases DNMT1 and PRMT6. Gastroenterology 2019, 157, 744–759.e4. [Google Scholar] [CrossRef]
- Kavalali, E.T.; Nelson, E.D.; Monteggia, L.M. Role of MeCP2, DNA methylation, and HDACs in regulating synapse function. J. Neurodev. Disord. 2011, 3, 250–256. [Google Scholar] [CrossRef] [Green Version]
- West, A.C.; Johnstone, R.W. New and emerging HDAC inhibitors for cancer treatment. J. Clin. Investig. 2014, 124, 30–39. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.S.; Parmigiani, R.B.; Marks, P.A. Histone deacetylase inhibitors: Molecular mechanisms of action. Oncogene 2007, 26, 5541–5552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harrison, I.F.; Dexter, D.T. Epigenetic targeting of histone deacetylase: Therapeutic potential in Parkinson’s disease? Pharmacol. Ther. 2013, 140, 34–52. [Google Scholar] [CrossRef] [PubMed]
- Gräff, J.; Tsai, L.H. The potential of HDAC inhibitors as cognitive enhancers. Annu. Rev. Pharmacol. Toxicol. 2013, 53, 311–330. [Google Scholar] [CrossRef] [Green Version]
- Takeshima, H.; Niwa, T.; Yamashita, S.; Takamura-Enya, T.; Iida, N.; Wakabayashi, M.; Nanjo, S.; Abe, M.; Sugiyama, T.; Kim, Y.-J.; et al. TET repression and increased DNMT activity synergistically induce aberrant DNA methylation. J. Clin. Investig. 2020, 130, 5370–5379. [Google Scholar] [CrossRef] [PubMed]
- Fasolino, M.; Zhou, Z. The Crucial Role of DNA Methylation and MeCP2 in Neuronal Function. Genes 2017, 8, 141. [Google Scholar] [CrossRef] [Green Version]
- Gontier, G.; Iyer, M.; Shea, J.M.; Bieri, G.; Wheatley, E.G.; Ramalho-Santos, M.; Villeda, S.A. Tet2 Rescues Age-Related Regenerative Decline and Enhances Cognitive Function in the Adult Mouse Brain. Cell Rep. 2018, 22, 1974–1981. [Google Scholar] [CrossRef] [Green Version]
- Hull, E.E.; Montgomery, M.R.; Leyva, K.J. HDAC Inhibitors as Epigenetic Regulators of the Immune System: Impacts on Cancer Therapy and Inflammatory Diseases. BioMed Res. Int. 2016, 2016, 8797206. [Google Scholar] [CrossRef] [Green Version]
- Downing, L.E.; Ferguson, B.S.; Rodriguez, K.; Ricketts, M.L. A grape seed procyanidin extract inhibits HDAC activity leading to increased PPAR-α phosphorylation and target-gene expression. Mol. Nutr. Food Res. 2017, 61, 1600347. [Google Scholar] [CrossRef] [Green Version]
- Uzunova, V.; Sheline, Y.; Davis, J.M.; Rasmusson, A.; Uzunov, D.P.; Costa, E.; Guidotti, A. Increase in the cerebrospinal fluid content of neurosteroids in patients with unipolar major depression who are receiving fluoxetine or fluvoxamine. Proc. Natl. Acad. Sci. USA 1998, 95, 3239–3244. [Google Scholar] [CrossRef] [Green Version]
- Agís-Balboa, R.C.; Guidotti, A.; Pinna, G. 5α-reductase type I expression is downregulated in the prefrontal cortex/Brodmann’s area 9 (BA9) of depressed patients. Psychopharmacology 2014, 231, 3569–3580. [Google Scholar] [CrossRef]
- Rasmusson, A.M.; Pinna, G.; Paliwal, P.; Weisman, D.; Gottschalk, C.; Charney, D.; Krystal, J.; Guidotti, A. Decreased cerebrospinal fluid allopregnanolone levels in women with posttraumatic stress disorder. Biol. Psychiatry 2006, 60, 704–713. [Google Scholar] [CrossRef] [PubMed]
- Meltzer-Brody, S.; Kanes, S. Allopregnanolone in postpartum depression: Role in pathophysiology and treatment. Neurobiol. Stress 2020, 12, 100212. [Google Scholar] [CrossRef] [PubMed]
- Maguire, J.L.; Stell, B.M.; Rafizadeh, M.; Mody, I. Ovarian cycle-linked changes in GABA(A) receptors mediating tonic inhibition alter seizure susceptibility and anxiety. Nat. Neurosci. 2005, 8, 797–804. [Google Scholar] [CrossRef] [PubMed]
- Paul, S.M.; Pinna, G.; Guidotti, A. Allopregnanolone: From molecular pathophysiology to therapeutics. A historical perspective. Neurobiol. Stress 2020, 12, 100215. [Google Scholar] [CrossRef] [PubMed]
- Zorumski, C.F.; Paul, S.M.; Covey, D.F.; Mennerick, S. Neurosteroids as novel antidepressants and anxiolytics: GABA-A receptors and beyond. Neurobiol. Stress 2019, 11, 100196. [Google Scholar] [CrossRef]
- Walton, N.; Maguire, J. Allopregnanolone-based treatments for postpartum depression: Why/how do they work? Neurobiol. Stress 2019, 11, 100198. [Google Scholar] [CrossRef]
- Kawasaki, T.; Kawai, T. Toll-like receptor signaling pathways. Front. Immunol. 2014, 5, 461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawai, T.; Akira, S. The role of pattern-recognition receptors in innate immunity: Update on Toll-like receptors. Nat. Immunol. 2010, 11, 373–384. [Google Scholar] [CrossRef]
- Yilmaz, C.; Karali, K.; Fodelianaki, G.; Gravanis, A.; Chavakis, T.; Charalampopoulos, I.; Alexaki, V.I. Neurosteroids as regulators of neuroinflammation. Front. Neuroendocrinol. 2019, 55, 100788. [Google Scholar] [CrossRef]
- Balan, I.; Beattie, M.C.; O’Buckley, T.K.; Aurelian, L.; Morrow, A.L. Endogenous Neurosteroid (3α,5α)3-Hydroxypregnan-20-one Inhibits Toll-like-4 Receptor Activation and Pro-inflammatory Signaling in Macrophages and Brain. Sci. Rep. 2019, 9, 1220. [Google Scholar] [CrossRef]
- Balan, I.; Aurelian, L.; Schleicher, R.; Boero, G.; O’Buckley, T.; Morrow, A.L. Neurosteroid allopregnanolone (3α,5α-THP) inhibits inflammatory signals induced by activated MyD88-dependent toll-like receptors. Transl. Psychiatry 2021, 11, 145. [Google Scholar] [CrossRef]
- Franklin, T.C.; Xu, C.; Duman, R.S. Depression and sterile inflammation: Essential role of danger associated molecular patterns. Brain Behav. Immun. 2018, 72, 2–13. [Google Scholar] [CrossRef]
- Chistyakov, D.V.; Aleshin, S.E.; Astakhova, A.A.; Sergeeva, M.G.; Reiser, G. Regulation of peroxisome proliferator-activated receptors (PPAR) α and -γ of rat brain astrocytes in the course of activation by toll-like receptor agonists. J. Neurochem. 2015, 134, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Guidotti, A.; Dong, E.; Matsumoto, K.; Pinna, G.; Rasmusson, A.M.; Costa, E. The socially-isolated mouse: A model to study the putative role of allopregnanolone and 5alpha-dihydroprogesterone in psychiatric disorders. Brain Res. Brain Res. Rev. 2001, 37, 110–115. [Google Scholar] [CrossRef]
- Sasso, O.; Russo, R.; Vitiello, S.; Raso, G.M.; D’Agostino, G.; Iacono, A.; La Rana, G.; Vallée, M.; Cuzzocrea, S.; Piazza, P.V.; et al. Implication of allopregnanolone in the antinociceptive effect of N-palmitoylethanolamide in acute or persistent pain. Pain 2012, 153, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Berger, J.; Moller, D.E. The mechanisms of action of PPARs. Annu. Rev. Med. 2002, 53, 409–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radley, J.J.; Kabbaj, M.; Jacobson, L.; Heydendael, W.; Yehuda, R.; Herman, J.P. Stress risk factors and stress-related pathology: Neuroplasticity, epigenetics and endophenotypes. Stress 2011, 14, 481–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGowan, P.O.; Sasaki, A.; D’Alessio, A.C.; Dymov, S.; Labonté, B.; Szyf, M.; Turecki, G.; Meaney, M.J. Epigenetic regulation of the glucocorticoid receptor in human brain associates with childhood abuse. Nat. Neurosci. 2009, 12, 342–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merali, Z.; Du, L.; Hrdina, P.; Palkovits, M.; Faludi, G.; Poulter, M.; Anisman, H. Dysregulation in the suicide brain: mRNA expression of corticotropin-releasing hormone receptors and GABA(A) receptor subunits in frontal cortical brain region. J. Neurosci. 2004, 24, 1478–1485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aspesi, D.; Pinna, G. Could a blood test for PTSD and depression be on the horizon? Expert Rev. Proteom. 2018, 15, 983–1006. [Google Scholar] [CrossRef] [PubMed]
- Li, T.T.; Tan, T.B.; Hou, H.Q.; Zhao, X.Y. Changes in peroxisome proliferator-activated receptor alpha target gene expression in peripheral blood mononuclear cells associated with non-alcoholic fatty liver disease. Lipids Health Dis. 2018, 17, 256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Locci, A.; Geoffroy, P.; Miesch, M.; Mensah-Nyagan, A.G.; Pinna, G. Social Isolation in Early versus Late Adolescent Mice Is Associated with Persistent Behavioral Deficits That Can Be Improved by Neurosteroid-Based Treatment. Front. Cell. Neurosci. 2017, 11, 208. [Google Scholar] [CrossRef] [Green Version]
- Akhter, N.; Madhoun, A.; Arefanian, H.; Wilson, A.; Kochumon, S.; Thomas, R.; Shenouda, S.; Al-Mulla, F.; Ahmad, R.; Sindhu, S. Oxidative Stress Induces Expression of the Toll-Like Receptors (TLRs) 2 and 4 in the Human Peripheral Blood Mononuclear Cells: Implications for Metabolic Inflammation. Cell. Physiol. Biochem. 2019, 53, 1–18. [Google Scholar] [CrossRef]
- Lo Verme, J.; Fu, J.; Astarita, G.; La Rana, G.; Russo, R.; Calignano, A.; Piomelli, D. The nuclear receptor peroxisome proliferator-activated receptor-alpha mediates the anti-inflammatory actions of palmitoylethanolamide. Mol. Pharmacol. 2005, 67, 15–19. [Google Scholar] [CrossRef]
- Esmaeili, M.A.; Yadav, S.; Gupta, R.K.; Waggoner, G.R.; Deloach, A.; Calingasan, N.Y.; Beal, M.F.; Kiaei, M. Preferential PPAR-α activation reduces neuroinflammation, and blocks neurodegeneration in vivo. Hum. Mol. Genet. 2016, 25, 317–327. [Google Scholar] [CrossRef] [Green Version]
- Beggiato, S.; Tomasini, M.C.; Ferraro, L. Palmitoylethanolamide (PEA) as a Potential Therapeutic Agent in Alzheimer’s Disease. Front. Pharmacol. 2019, 10, 821. [Google Scholar] [CrossRef] [Green Version]
- Pinna, G.; Dong, E.; Matsumoto, K.; Costa, E.; Guidotti, A. In socially isolated mice, the reversal of brain allopregnanolone down-regulation mediates the anti-aggressive action of fluoxetine. Proc. Natl. Acad. Sci. USA 2003, 100, 2035–2040. [Google Scholar] [CrossRef] [Green Version]
- Matrisciano, F.; Tueting, P.; Dalal, I.; Kadriu, B.; Grayson, D.R.; Davis, J.M.; Nicoletti, F.; Guidotti, A. Epigenetic modifications of GABAergic interneurons are associated with the schizophrenia-like phenotype induced by prenatal stress in mice. Neuropharmacology 2013, 68, 184–194. [Google Scholar] [CrossRef] [Green Version]
- Gatta, E.; Guidotti, A.; Saudagar, V.; Pinna, G. Epigenetic Regulation of GABAergic Neurotransmission and Neurosteroid Biosynthesis in Alcohol Use Disorder. Int. J. Neuropsychopharmacol. 2021, 24, 130–141. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward | Reverse |
|---|---|---|
| mRNA expression | ||
| β-Actin | 5′-TGTGATGGTGGGAATGGGTCAGAA | 5′-TGTGGTGCCAGATCTTCTCCATGT |
| Ppar-alpha | 5′-CACAGACACCCTCTCTCCAG | 5′-AGCCCTTACAGCCTTCACAT |
| Epigenetic Marks | ||
| HDAC1 | 5′-CACAGACACCCTCTCTCCAG | 5′-AGCCCTTACAGCCTTCACAT |
| HDAC2 | 5′-GCTTGCCATCCTCGAATTAC | 5′-CCCTCAAGTCTCCTGTTCCA |
| MeCP2 | 5′-GGTTGTCTCCACTGCTACTTAC | 5′-GCTAACTTGGGTGCTGATCT |
| TET1 | 5′-TTGCCCAGACCATAAGGAAC | 5′-GTGGTGACACTCATGGCATC |
| TET2 | 5′-GTTCTCAACGAGCAGGAAGG | 5′-TGAGATGCGGTACTCTGCAC |
| TET3 | 5′-TCCGGATTGAGAAGGTCATC | 5′- CCAGGCCAGGATCAAGATAA |
| DNMT1 | 5′-CCACCACCAAGCTGGTCTAT | 5′-TGCCACCAAACTTCACCATA |
| DNMT3a DNMT3b Toll-Like Receptors | 5′-ACCAGGCCACCTACAACAAG 5′-ACTTGGTGATTGGTGGAAGC | 5′-TGCTTGTTCTGCACTTCCAC 5′-CCAGAAGAATGGACGGTTGT |
| TLR4 | 5′-GGCAGCAGGTGGAATTGTAT | 5′-AGGATTCGAGGCTTTTCCAT |
| Inflammatory marks | ||
| TNFα | 5′- ACGGCATGGATCTCAAAGAC | 5′-GTGGGTGAGGAGCACGTAGT |
| MCP-1 | 5′-CCCAATGAGTAGGCTGGAGA | 5′-TCTGGACCCATTCCTTCTTG |
| NFkB1 | 5′- CACCTAGCTGCCAAAGAAGG | 5′ GCAGGCTATTGCTCATCACA |
| NFkB2 | 5′- GATCTCCCGAATGGACAAGA | 5′ GAACCGAACCTCAATGTCGT |
| Ppar-alpha gene | ||
| (−50 to +31 bp) | 5′- GGAGGGGAGGGGACTCG | 5′- CGCGTGTGCCCTTCCTAG |
| (+9 +149 bp) | 5′- CAGCCACTGGAGAGGGCACA | 5′- CTCCAGTTCCAGGACTCCAC |
| (+1715 +1813 bp) | 5′- TTCAAAAAATGGTGGACCTT | 5′- TCTTGCAACAGTGGGTGC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matrisciano, F.; Pinna, G. PPAR-α Hypermethylation in the Hippocampus of Mice Exposed to Social Isolation Stress Is Associated with Enhanced Neuroinflammation and Aggressive Behavior. Int. J. Mol. Sci. 2021, 22, 10678. https://doi.org/10.3390/ijms221910678
Matrisciano F, Pinna G. PPAR-α Hypermethylation in the Hippocampus of Mice Exposed to Social Isolation Stress Is Associated with Enhanced Neuroinflammation and Aggressive Behavior. International Journal of Molecular Sciences. 2021; 22(19):10678. https://doi.org/10.3390/ijms221910678
Chicago/Turabian StyleMatrisciano, Francesco, and Graziano Pinna. 2021. "PPAR-α Hypermethylation in the Hippocampus of Mice Exposed to Social Isolation Stress Is Associated with Enhanced Neuroinflammation and Aggressive Behavior" International Journal of Molecular Sciences 22, no. 19: 10678. https://doi.org/10.3390/ijms221910678
APA StyleMatrisciano, F., & Pinna, G. (2021). PPAR-α Hypermethylation in the Hippocampus of Mice Exposed to Social Isolation Stress Is Associated with Enhanced Neuroinflammation and Aggressive Behavior. International Journal of Molecular Sciences, 22(19), 10678. https://doi.org/10.3390/ijms221910678