
Anti-Inflammatory Effect of Licochalcone A via Regulation of ORAI1 and K+ Channels in T-Lymphocytes

 , ,
, ,  ,
, 
Abstract
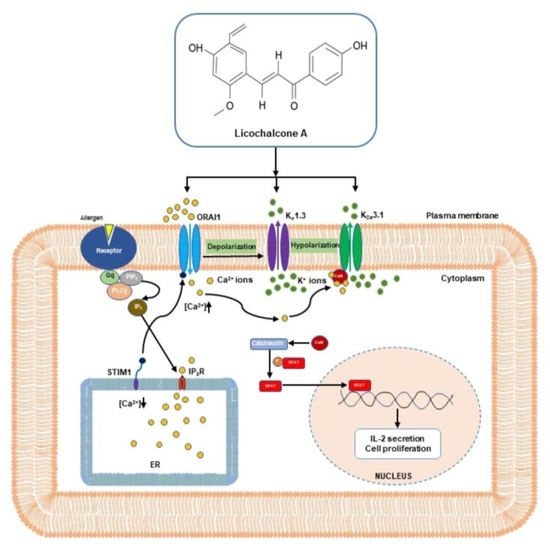
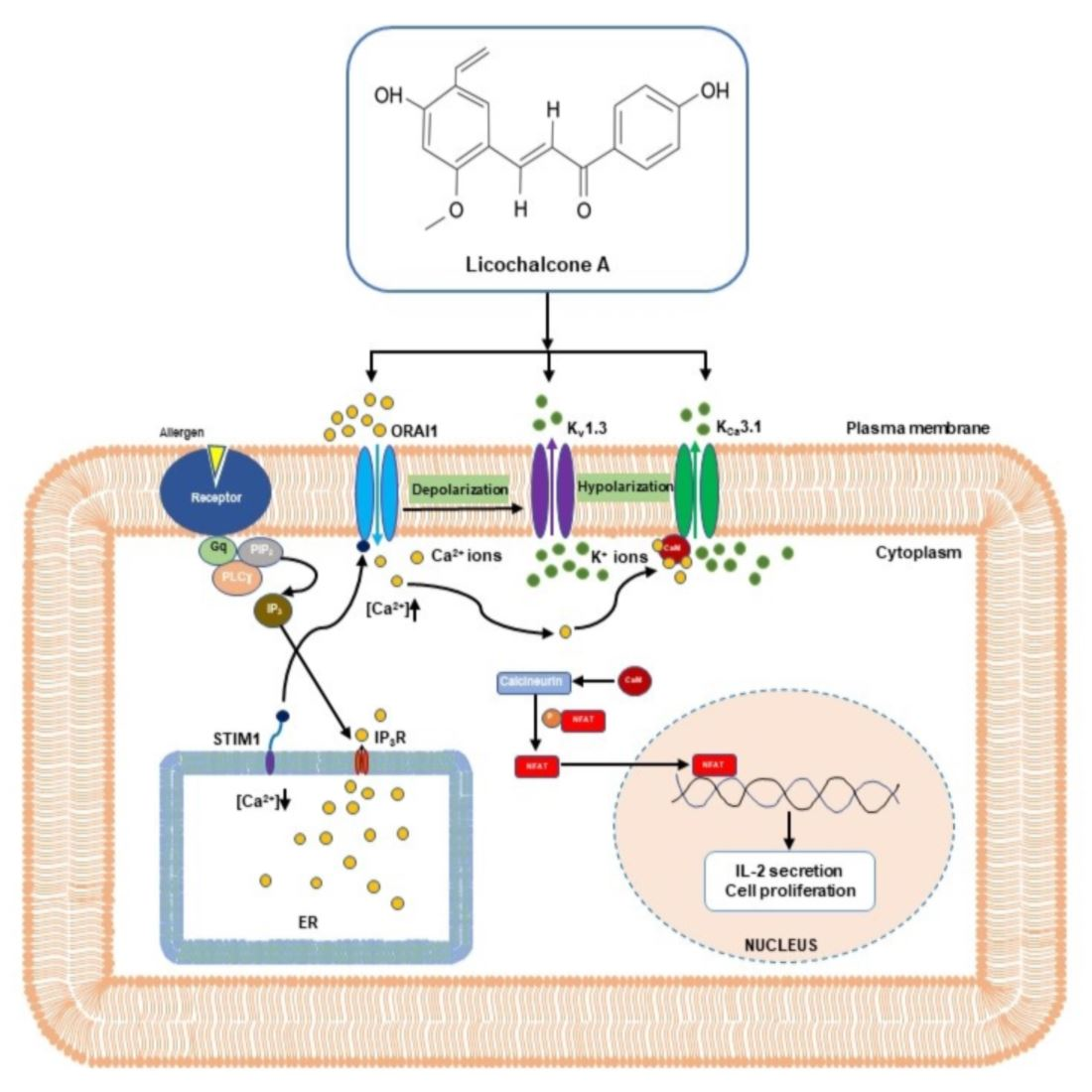
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
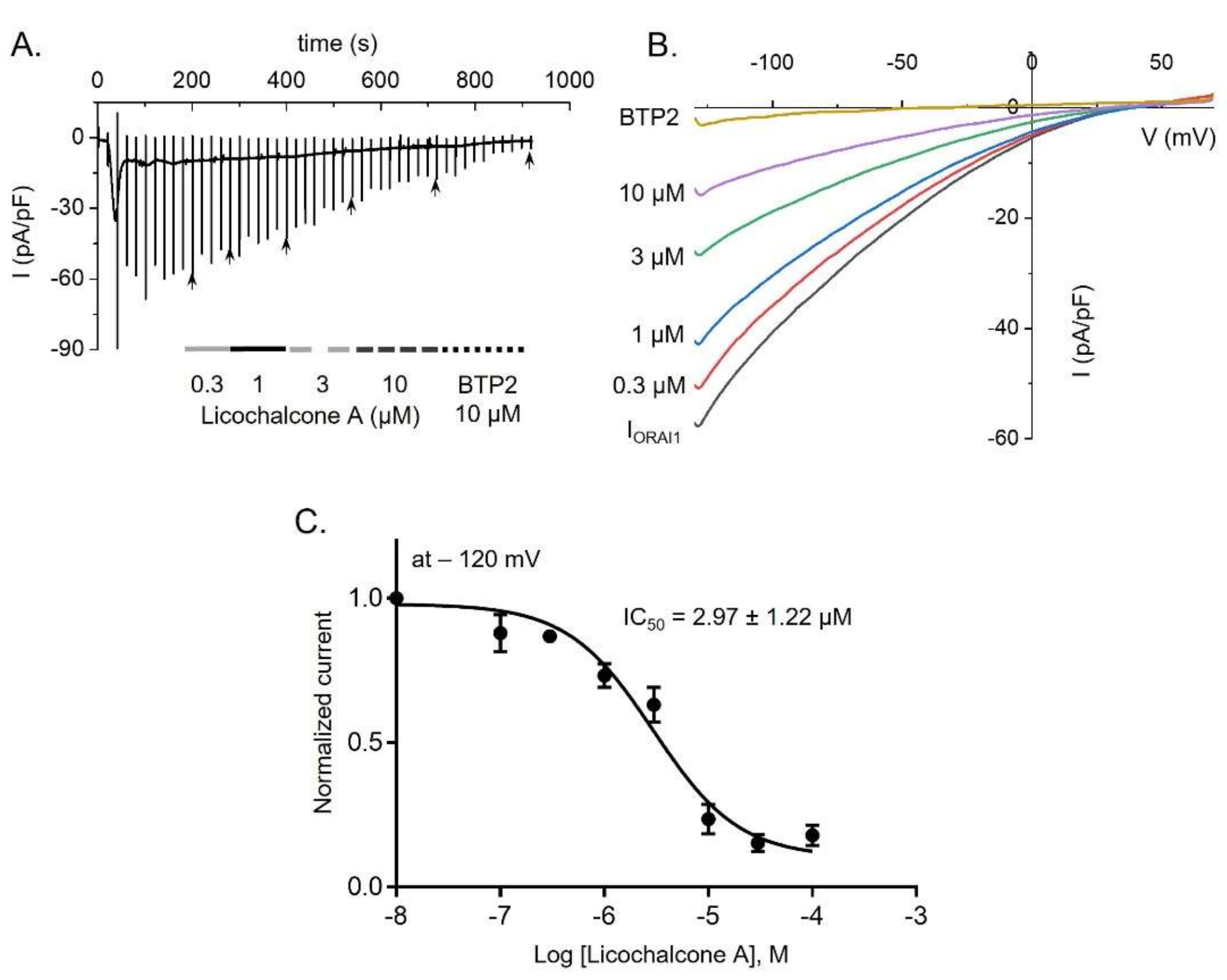
2.1. Licochalcone A Inhibited ORAI1 Currents in HEK293T Cells Coexpressing hORAI1 with hSTIM1
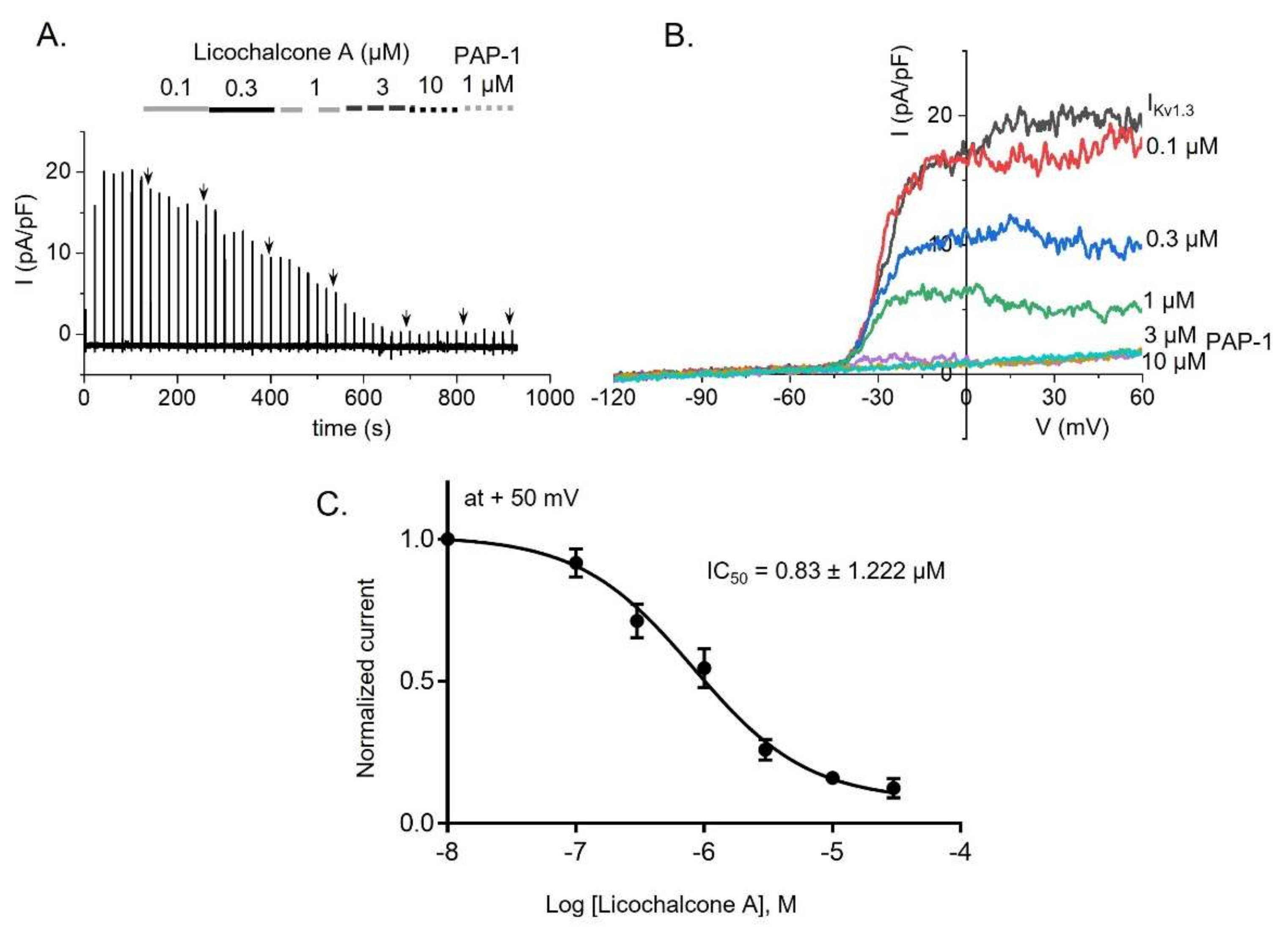
2.2. Inhibitory Effect of Kv1.3 Currents by Licochalcone A in Jurkat T-Cells
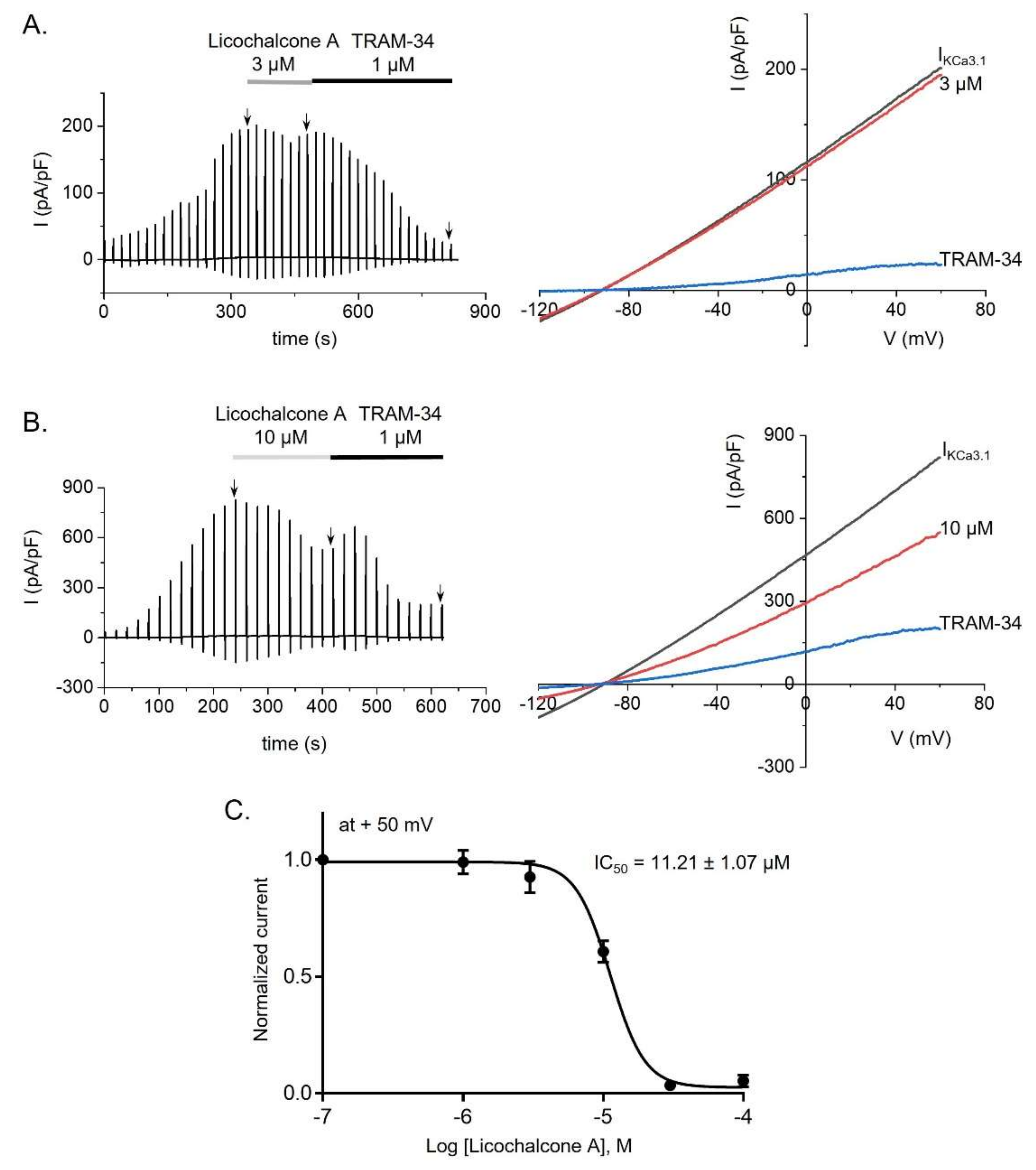
2.3. Licochalcone A Inhibited KCa3.1 Currents in KCa3.1-Overexpressing HEK293 Cells
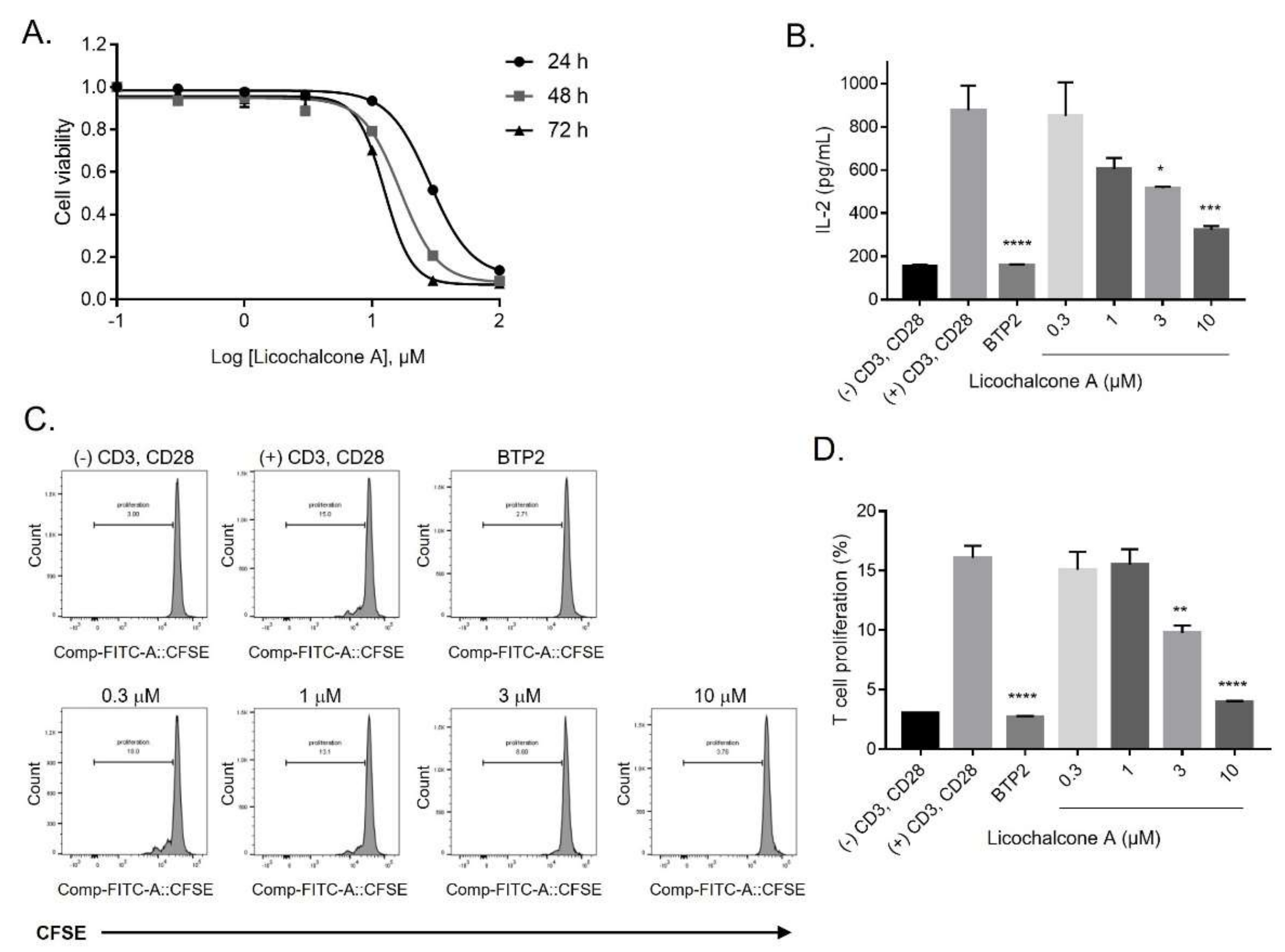
2.4. Inhibition of Release of IL-2 and Proliferation of T-lymphocytes by Licochalcone A
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Cell Line and Cell Culture
4.3. Transient Transfection
4.4. Electrophysiological Experimentation
4.5. Human CD4+ T-Lymphocyte Assay
4.6. Cell Cytotoxicity
4.7. Measurement of IL-2 Secretion
4.8. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Feske, S. Calcium signalling in lymphocyte activation and disease. Nat. Rev. Immunol. 2007, 7, 690–702. [Google Scholar] [CrossRef]
- Vig, M.; Kinet, J.P. Calcium signaling in immune cells. Nat. Immunol. 2009, 10, 21–27. [Google Scholar] [CrossRef] [Green Version]
- Lewis, R.S. Calcium signaling mechanisms in T lymphocytes. Annu. Rev. Immunol. 2001, 19, 497–521. [Google Scholar] [CrossRef]
- Yang, P.-C.; Jafri, M.S. Ca2+ signaling in T lymphocytes: The interplay of the endoplasmic reticulum, mitochondria, membrane potential, and CRAC channels on transcription factor activation. Heliyon 2020, 6, e03526. [Google Scholar] [CrossRef] [PubMed]
- Vaeth, M.; Kahlfuss, S.; Feske, S. CRAC Channels and Calcium Signaling in T Cell-Mediated Immunity. Trends Immunol. 2020, 41, 878–901. [Google Scholar] [CrossRef] [PubMed]
- Feske, S.; Wulff, H.; Skolnik, E.Y. Ion Channels in Innate and Adaptive Immunity. Annu. Rev. Immunol. 2015, 33, 291–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hutchings, C.J.; Colussi, P.; Clark, T.G. Ion channels as therapeutic antibody targets. mAbs 2019, 11, 265–296. [Google Scholar] [CrossRef]
- Cahalan, M.D.; Chandy, K.G. The functional network of ion channels in T lymphocytes. Immunol. Rev. 2009, 231, 59–87. [Google Scholar] [CrossRef] [Green Version]
- Lam, J.; Wulff, H. The Lymphocyte Potassium Channels Kv1.3 and KCa3.1 as Targets for Immunosuppression. Drug Dev. Res. 2011, 72, 573–584. [Google Scholar] [CrossRef] [Green Version]
- Fu, Y.; Chen, J.; Li, Y.-J.; Zheng, Y.-F.; Li, P. Antioxidant and anti-inflammatory activities of six flavonoids separated from licorice. Food Chem. 2013, 141, 1063–1071. [Google Scholar] [CrossRef]
- Won, S.-R.; Kim, S.-K.; Kim, Y.-M.; Lee, P.-H.; Ryu, J.-H.; Kim, J.-W.; Rhee, H.-I. Licochalcone A: A lipase inhibitor from the roots of Glycyrrhiza uralensis. Food Res. Int. 2007, 40, 1046–1050. [Google Scholar] [CrossRef]
- Lv, H.; Xiao, Q.; Zhou, J.; Feng, H.; Liu, G.; Ci, X. Licochalcone A Upregulates Nrf2 Antioxidant Pathway and Thereby Alleviates Acetaminophen-Induced Hepatotoxicity. Front. Pharmacol. 2018, 9, 147. [Google Scholar] [CrossRef] [Green Version]
- Tsukiyama, R.; Katsura, H.; Tokuriki, N.; Kobayashi, M. Antibacterial activity of licochalcone A against spore-forming bacteria. Antimicrob. Agents Chemother 2002, 46, 1226–1230. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.C.; Liu, C.Y.; Shen, S.C.; Chen, L.C.; Yeh, K.W.; Liu, S.H.; Liou, C.J. Protective Effects of Licochalcone A Improve Airway Hyper-Responsiveness and Oxidative Stress in a Mouse Model of Asthma. Cells 2019, 8, 617. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.K.; Shin, E.K.; Park, J.H.; Kim, Y.H.; Park, J.H. Antitumor and antimetastatic effects of licochalcone A in mouse models. J. Mol. Med. 2010, 88, 829–838. [Google Scholar] [CrossRef] [PubMed]
- Jia, T.; Qiao, J.; Guan, D.; Chen, T. Anti-Inflammatory Effects of Licochalcone A on IL-1β-Stimulated Human Osteoarthritis Chondrocytes. Inflammation 2017, 40, 1894–1902. [Google Scholar] [CrossRef]
- Kolbe, L.; Immeyer, J.; Batzer, J.; Wensorra, U.; tom Dieck, K.; Mundt, C.; Wolber, R.; Stäb, F.; Schönrock, U.; Ceilley, R.I.; et al. Anti-inflammatory efficacy of Licochalcone A: Correlation of clinical potency and in vitro effects. Arch. Derm. Res. 2006, 298, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Chu, X.; Ci, X.; Wei, M.; Yang, X.; Cao, Q.; Guan, M.; Li, H.; Deng, Y.; Feng, H.; Deng, X. Licochalcone A Inhibits Lipopolysaccharide-Induced Inflammatory Response In Vitro and In Vivo. J. Agric. Food Chem. 2012, 60, 3947–3954. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Liu, B.; Yin, Y.; Kan, X.; Gong, Q.; Li, Y.; Cao, Y.; Wang, J.; Xu, D.; Ma, H.; et al. Licochalcone A Protects the Blood–Milk Barrier Integrity and Relieves the Inflammatory Response in LPS-Induced Mastitis. Front. Immunol. 2019, 10, 287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanifuji, S.; Aizu-Yokota, E.; Funakoshi-Tago, M.; Sonoda, Y.; Inoue, H.; Kasahara, T. Licochalcones suppress degranulation by decreasing the intracellular Ca2+ level and tyrosine phosphorylation of ERK in RBL-2H3 cells. Int. Immunopharmacol. 2010, 10, 769–776. [Google Scholar] [CrossRef]
- Cui, Y.; Ao, M.; Hu, J.; Yu, L. Anti-Inflammatory Activity of Licochalcone A Isolated from Glycyrrhiza inflata. Z. Für. Nat. C 2008, 63, 361–365. [Google Scholar] [CrossRef] [Green Version]
- Chu, X.; Jiang, L.; Wei, M.; Yang, X.; Guan, M.; Xie, X.; Wei, J.; Liu, D.; Wang, D. Attenuation of allergic airway inflammation in a murine model of asthma by Licochalcone A. Immunopharmacol. Immunotoxicol. 2013, 35, 653–661. [Google Scholar] [CrossRef]
- Chiu, Y.J.; Lee, C.M.; Lin, T.H.; Lin, H.Y.; Lee, S.Y.; Mesri, M.; Chang, K.H.; Lin, J.Y.; Lee-Chen, G.J.; Chen, C.M. Chinese Herbal Medicine Glycyrrhiza inflata Reduces Aβ Aggregation and Exerts Neuroprotection through Anti-Oxidation and Anti-Inflammation. Am. J. Chin. Med. 2018, 12, 1–25. [Google Scholar] [CrossRef]
- Nicolaou, S.A.; Neumeier, L.; Steckly, A.; Kucher, V.; Takimoto, K.; Conforti, L. Localization of Kv1.3 Channels in the Immunological Synapse Modulates the Calcium Response to Antigen Stimulation in T Lymphocytes. J. Immunol. 2009, 183, 6296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szabò, I.; Bock, J.; Jekle, A.; Soddemann, M.; Adams, C.; Lang, F.; Zoratti, M.; Gulbins, E. A Novel Potassium Channel in Lymphocyte Mitochondria. J. Biol. Chem. 2005, 280, 12790–12798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srivastava, S.; Di, L.; Zhdanova, O.; Li, Z.; Vardhana, S.; Wan, Q.; Yan, Y.; Varma, R.; Backer, J.; Wulff, H.; et al. The class II phosphatidylinositol 3 kinase C2beta is required for the activation of the K+ channel KCa3.1 and CD4+ T-cells. Mol. Biol. Cell. 2009, 20, 3783–3791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiang, E.Y.; Li, T.; Jeet, S.; Peng, I.; Zhang, J.; Lee, W.P.; DeVoss, J.; Caplazi, P.; Chen, J.; Warming, S.; et al. Potassium channels Kv1.3 and KCa3.1 cooperatively and compensatorily regulate antigen-specific memory T cell functions. Nat. Commun. 2017, 8, 14644. [Google Scholar] [CrossRef]
- Kaufmann, U.; Shaw, P.J.; Kozhaya, L.; Subramanian, R.; Gaida, K.; Unutmaz, D.; McBride, H.J.; Feske, S. Selective ORAI1 Inhibition Ameliorates Autoimmune Central Nervous System Inflammation by Suppressing Effector but Not Regulatory T Cell Function. J. Immunol. 2016, 196, 573–585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Watanabe, S.; Shudou, M.; Kuno, M.; Miura, H.; Maeyama, K. Upregulation of store-operated Ca2+ entry in the naïve CD4+ T cells with aberrant cytokine releasing in active rheumatoid arthritis. Immunol. Cell Biol. 2014, 92, 752–760. [Google Scholar] [CrossRef]
- Koch Hansen, L.; Sevelsted-Møller, L.; Rabjerg, M.; Larsen, D.; Hansen, T.P.; Klinge, L.; Wulff, H.; Knudsen, T.; Kjeldsen, J.; Köhler, R. Expression of T-cell KV1.3 potassium channel correlates with pro-inflammatory cytokines and disease activity in ulcerative colitis. J. Crohns. Colitis. 2014, 8, 1378–1391. [Google Scholar] [CrossRef]
- Beeton, C.; Wulff, H.; Barbaria, J.; Clot-Faybesse, O.; Pennington, M.; Bernard, D.; Cahalan, M.D.; Chandy, K.G.; Béraud, E. Selective blockade of T lymphocyte K+ channels ameliorates experimental autoimmune encephalomyelitis, a model for multiple sclerosis. Proc. Natl. Acad. Sci. 2001, 98, 13942. [Google Scholar] [CrossRef] [Green Version]
- Kimura, Y.; Okuda, H.; Okuda, T.; Arichi, S. Effects of chalcones isolated from licorice roots on leukotriene biosynthesis in human polymorphonuclear neutrophls. Phytother. Res. 1988, 2, 140–145. [Google Scholar] [CrossRef]
- Price, M.; Lee, S.C.; Deutsch, C. Charybdotoxin inhibits proliferation and interleukin 2 production in human peripheral blood lymphocytes. Proc. Natl. Acad. Sci. USA 1989, 86, 10171–10175. [Google Scholar] [CrossRef] [Green Version]
- Tang, Z.-H.; Chen, X.; Wang, Z.-Y.; Chai, K.; Wang, Y.-F.; Xu, X.-H.; Wang, X.-W.; Lu, J.-H.; Wang, Y.-T.; Chen, X.-P.; et al. Induction of C/EBP homologous protein-mediated apoptosis and autophagy by licochalcone A in non-small cell lung cancer cells. Sci. Rep. 2016, 6, 26241. [Google Scholar] [CrossRef] [Green Version]
- Lin, R.-C.; Yang, S.-F.; Chiou, H.-L.; Hsieh, S.-C.; Wen, S.-H.; Lu, K.-H.; Hsieh, Y.-H. Licochalcone A-Induced Apoptosis Through the Activation of p38MAPK Pathway Mediated Mitochondrial Pathways of Apoptosis in Human Osteosarcoma Cells In Vitro and In Vivo. Cells 2019, 8, 1441. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Xing, Y.; Li, M.; Zhang, Z.; Wang, J.; Ri, M.; Jin, C.; Xu, G.; Piao, L.; Jin, H.; et al. Licochalcone A inhibits proliferation and promotes apoptosis of colon cancer cell by targeting programmed cell death-ligand 1 via the NF-κB and Ras/Raf/MEK pathways. J. Ethnopharmacol. 2021, 273, 113989. [Google Scholar] [CrossRef]
- Hao, Y.; Zhang, C.; Sun, Y.; Xu, H. Licochalcone A inhibits cell proliferation, migration, and invasion through regulating the PI3K/AKT signaling pathway in oral squamous cell carcinoma. Onco. Targets 2019, 12, 4427–4435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, W.; Yuan, X.; Yu, L.; Gao, C.; Sun, X.; Wang, D.; Zheng, Q. Licochalcone A-induced human gastric cancer BGC-823 cells apoptosis by regulating ROS-mediated MAPKs and PI3K/AKT signaling pathways. Sci. Rep. 2015, 5, 10336. [Google Scholar] [CrossRef] [PubMed]
- Shen, T.-S.; Hsu, Y.-K.; Huang, Y.-F.; Chen, H.-Y.; Hsieh, C.-P.; Chen, C.-L. Licochalcone A Suppresses the Proliferation of Osteosarcoma Cells through Autophagy and ATM-Chk2 Activation. Molecules 2019, 24, 2435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, W.-C.; Su, H.-H.; Fang, L.-W.; Wu, S.-J.; Liou, C.-J. Licochalcone A Inhibits Cellular Motility by Suppressing E-cadherin and MAPK Signaling in Breast Cancer. Cells 2019, 8, 218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, X.; Tian, L.; Wang, L.; Li, W.; Xu, Q.; Xiao, X. Antitumor effects and the underlying mechanism of licochalcone A combined with 5-fluorouracil in gastric cancer cells. Oncol. Lett. 2017, 13, 1695–1701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, P.T.L.; Kim, H.J.; Kim, W.K.; Nam, J.H. Flos magnoliae constituent fargesin has an anti-allergic effect via ORAI1 channel inhibition. Korean J. Physiol. Pharmacol. 2021, 25, 251–258. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Park, S.; Shin, H.Y.; Nam, Y.R.; Lam Hong, P.T.; Chin, Y.W.; Nam, J.H.; Kim, W.K. Inhibitory effects of α-Mangostin on T cell cytokine secretion via ORAI1 calcium channel and K+ channels inhibition. PeerJ 2021, 9, e10973. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Phan, H.T.L.; Kim, H.J.; Jo, S.; Kim, W.K.; Namkung, W.; Nam, J.H. Anti-Inflammatory Effect of Licochalcone A via Regulation of ORAI1 and K+ Channels in T-Lymphocytes. Int. J. Mol. Sci. 2021, 22, 10847. https://doi.org/10.3390/ijms221910847
Phan HTL, Kim HJ, Jo S, Kim WK, Namkung W, Nam JH. Anti-Inflammatory Effect of Licochalcone A via Regulation of ORAI1 and K+ Channels in T-Lymphocytes. International Journal of Molecular Sciences. 2021; 22(19):10847. https://doi.org/10.3390/ijms221910847
Chicago/Turabian StylePhan, Hong T. L., Hyun J. Kim, Sungwoo Jo, Woo K. Kim, Wan Namkung, and Joo H. Nam. 2021. "Anti-Inflammatory Effect of Licochalcone A via Regulation of ORAI1 and K+ Channels in T-Lymphocytes" International Journal of Molecular Sciences 22, no. 19: 10847. https://doi.org/10.3390/ijms221910847
APA StylePhan, H. T. L., Kim, H. J., Jo, S., Kim, W. K., Namkung, W., & Nam, J. H. (2021). Anti-Inflammatory Effect of Licochalcone A via Regulation of ORAI1 and K+ Channels in T-Lymphocytes. International Journal of Molecular Sciences, 22(19), 10847. https://doi.org/10.3390/ijms221910847