Endogenous Opioid Signaling in the Mouse Retina Modulates Pupillary Light Reflex


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
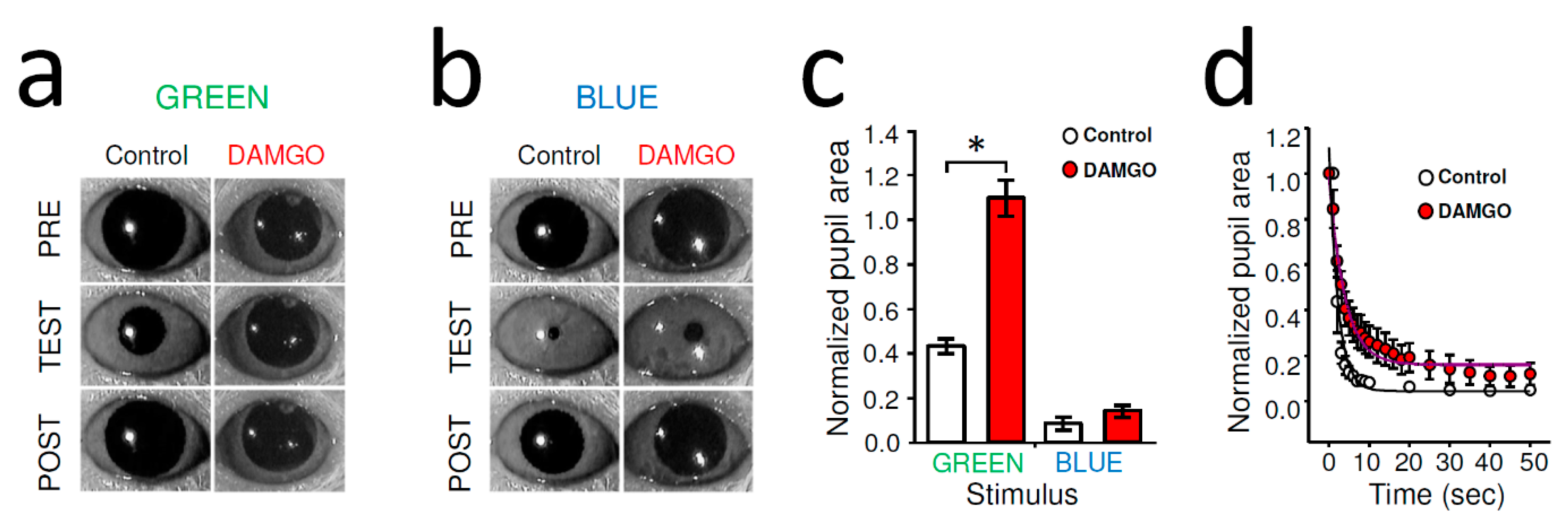
2.1. MOR-Specific Agonist DAMGO Inhibited Dark-Adapted Pupillary Light Reflex (PLR) in Wild-Type (WT) Mice
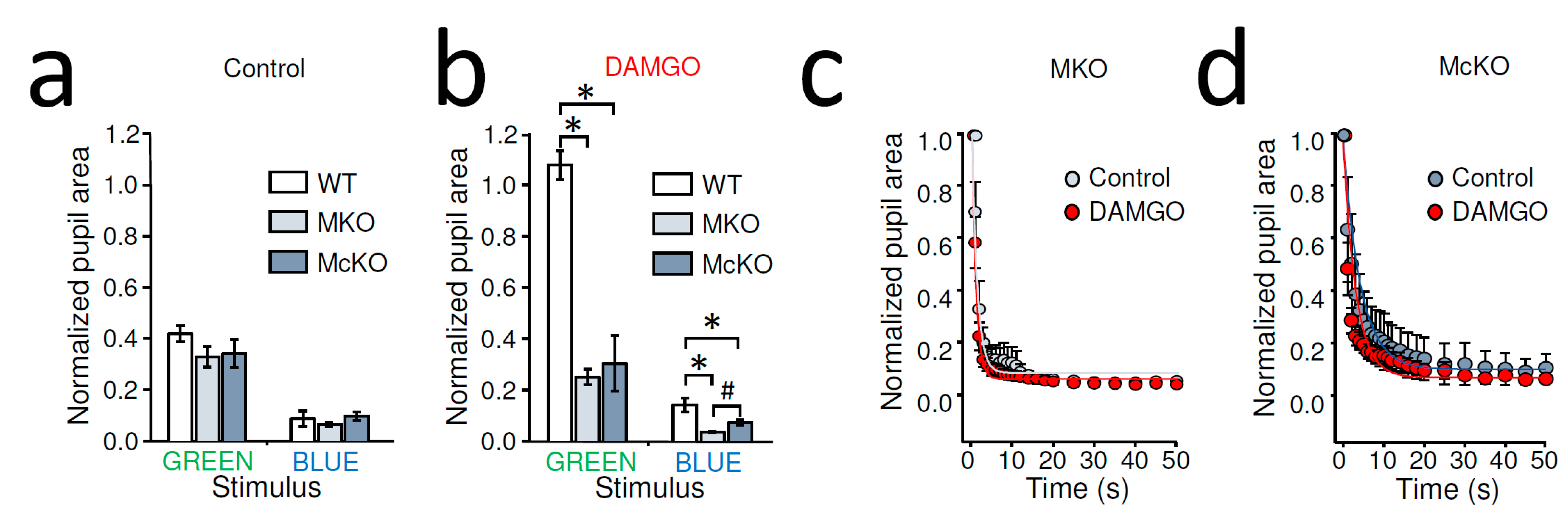
2.2. Dark-Adapted MOR-Deficient Mice Showed Normal PLR, but Intraocular DAMGO Had No Effect on Their PLR
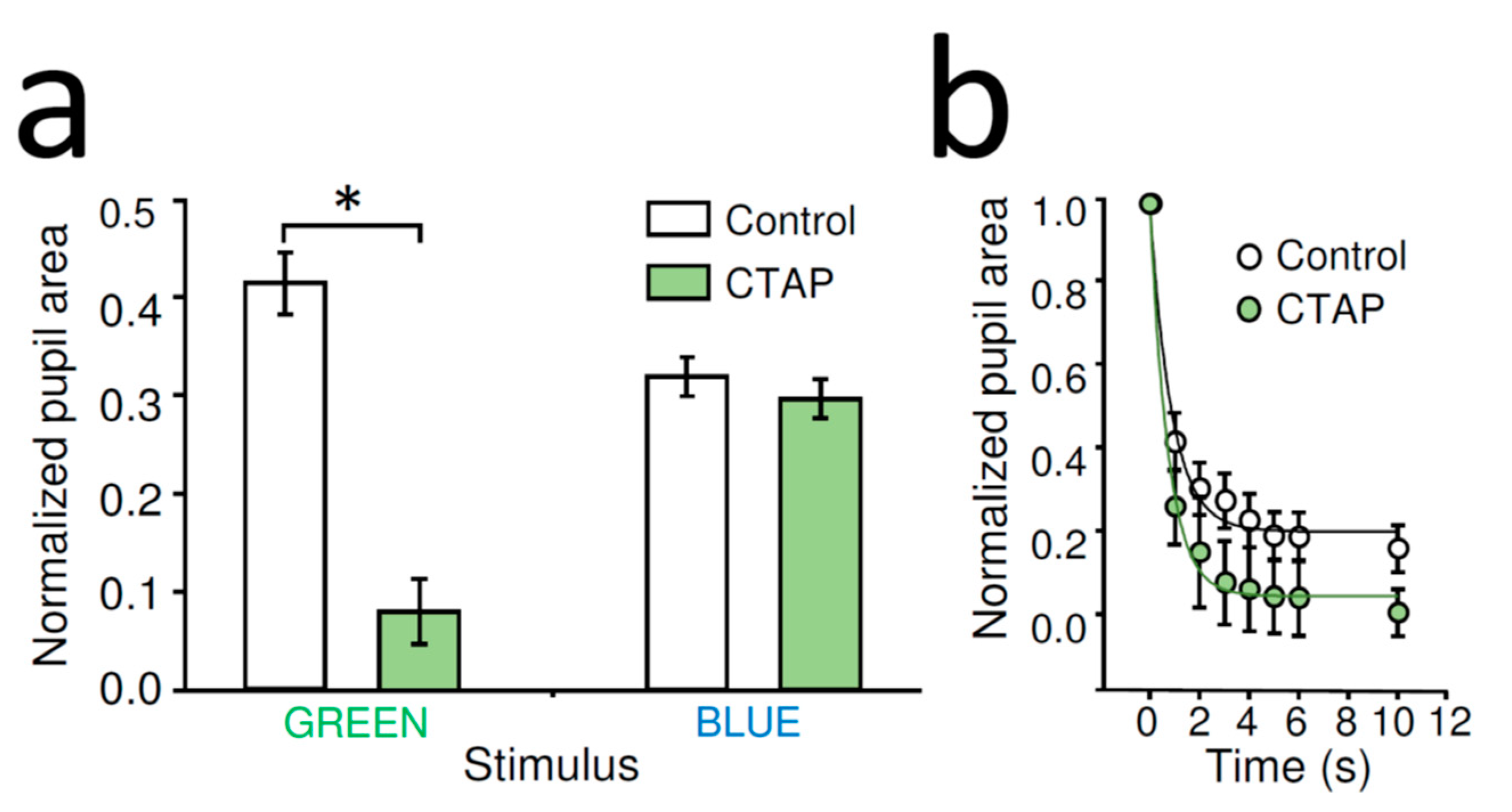
2.3. MOR Selective Antagonist CTAP Increased Dark-Adapted PLR Triggered by Rod-Saturating Green Light in WT Mice
3. Discussion
3.1. Pupillary Light Reflex (PLR) in Mice
3.2. Role for Endogenous Opioid Regulation of the PLR
3.3. Considerations for Human Clinical Practice
4. Materials and Methods
4.1. Animals
4.2. In Vivo Pupillometry
4.3. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| MOR | µ opioid receptor |
| ipRGC | intrinsically photosensitive retinal ganglion cell |
| PLR | pupillary light reflex |
| DAMGO | [D-Ala2, MePhe4, Gly-ol5]-enkephalin |
| CTAP | D-Phe-Cys-Tyr-D-Trp-Arg-Thr-Pen-Thr-NH2 |
| OPN | olivary pretectal nucleus |
| OMN | oculomotor nucleus |
| EWN | Edinger-Westphal nucleus |
| WT | Wild-type mouse |
| MKO | Mice lacking MORs throughout their body |
| McKO | Mice lacking MORs in ipRGCs |
| IOP | Intraocular pressure |
| PUAL | pupillary unrest under ambient light |
References
- Manchikanti, L.; Helm, S., II; Fellow, B.; Janata, J.W.; Pampati, V.; Grider, J.S.; Boswell, M. V Opioid Epidemic in the United States. Pain Physician 2012, 15, ES9–ES38. [Google Scholar] [CrossRef] [PubMed]
- Cobaugh, D.J.; Gainor, C.; Gaston, C.L.; Kwong, T.C.; Magnani, B.; Mcpherson, M.L.; Painter, J.T.; Krenzelok, E.P. The opioid abuse and misuse epidemic: Implications for pharmacists in hospitals and health systems. Am. J. Health-Syst. Pharm. 2014, 71, 1539–1554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poon, S.J.; Greenwood-Ericksen, M.B. The Opioid Prescription Epidemic and the Role of Emergency Medicine. Ann. Emerg. Med. 2014, 64, 490–495. [Google Scholar] [CrossRef] [PubMed]
- Grace, P.M.; Stanford, T.; Gentgall, M.; Rolan, P.E. Utility of saccadic eye movement analysis as an objective biomarker to detect the sedative interaction between opioids and sleep deprivation in opioid-naive and opioid-tolerant populations. J. Psychopharmacol. 2010, 24, 1631–1640. [Google Scholar] [CrossRef] [PubMed]
- Zacny, J.P.; Goldman, R.E. Characterizing the subjective, psychomotor, and physiological effects of oral propoxyphene in non-drug-abusing volunteers. Drug Alcohol Depend. 2004, 73, 133–140. [Google Scholar] [CrossRef]
- Verster, J.C.; Veldhuijzen, D.S.; Volkerts, E.R. Effects of an opioid (oxycodone/paracetamol) and an NSAID (bromfenac) on driving ability, memory functioning, psychomotor performance, pupil size, and mood. Clin. J. Pain 2006, 22, 499–504. [Google Scholar] [CrossRef] [PubMed]
- Murray, R.B.; Adler, M.W.; Korczyn, A.D. The pupillary effects of opioids. Life Sci. 1983, 33, 495–509. [Google Scholar] [CrossRef]
- Pickworth, W.B.; Welch, P.; Henningfield, J.E.; Cone, E.J. Opiate-Induced Pupllary Effects in Humans. Methods Find. Exp. Clin. Pharmacol. 1989, 11, 759–763. [Google Scholar] [PubMed]
- Pickworth, W.B.; Bunker, E.; Welch, P.; Cone, E. Intravenous buprenorphine reduces pupil size and the light reflex in humans. Life Sci. 1991, 49, 129–138. [Google Scholar] [CrossRef]
- Pickworth, W.B.; Sharpe, L.G. Morphine-induced mydriasis and inhibition of pupillary light reflex and fluctuations in the cat. J. Pharmacol. Exp. Ther. 1985, 234, 603–606. [Google Scholar]
- Sharpe, L.G. Separate neural mechanisms mediate sufentanil-induced pupillary responses in the cat. J. Pharmacol. Exp. Ther. 1991, 256, 845–849. [Google Scholar] [PubMed]
- Murray, R.B.; Loughnane, M.H. Infrared video pupillometry: A method used to measure the pupillary effects of drugs in small laboratory animals in real time. J. Neurosci. Methods 1981, 3, 365–375. [Google Scholar] [CrossRef]
- Pickworth, W.B.; Lee, H.; Fudala, P.J. Buprenorphine-induced pupillary effects in human volunteers. Life Sci. 1990, 47, 1269–1277. [Google Scholar] [CrossRef]
- Nisida, I.; Okada, H. The activity of the pupillo-constrictory centers. Jpn. J. Physiol. 1959, 10, 64–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adler, C.H.; Robin, M.; Adler, M.W. Tolerance to morphine-induced mydriasis in the rat pupil. Life Sci. 1981, 28, 2469–2475. [Google Scholar] [CrossRef]
- Lucas, R.J.; Douglas, R.H.; Foster, R.G. Characterization of an ocular photopigment capable of driving pupillary constriction in mice. Nat. Neurosci. 2001, 4, 621–626. [Google Scholar] [CrossRef]
- Lucas, R.J.; Hattar, S.S.; Takao, M.; Berson, D.M.; Foster, R.G.; Yau, K.-W. Diminished Pupillary Light Reflex at High Irradiances in Melanopsin-Knockout Mice. Science 2003, 299, 245–247. [Google Scholar] [CrossRef] [Green Version]
- Berson, D.M.; Dunn, F.A.; Takao, M. Phototransduction by retinal ganglion cells that set the circadian clock. Science 2002, 295, 1070–1073. [Google Scholar] [CrossRef] [Green Version]
- Young, M.J.; Lund, R.D. The anatomical substrates subserving the pupillary light reflex in rats: Origin of the consensual pupillary response. Neuroscience 1994, 62, 481–496. [Google Scholar] [CrossRef]
- Baver, S.B.; Pickard, G.E.; Sollars, P.J.; Pickard, G.E. Two types of melanopsin retinal ganglion cell differentially innervate the hypothalamic suprachiasmatic nucleus and the olivary pretectal nucleus. Eur. J. Neurosci. 2008, 27, 1763–1770. [Google Scholar] [CrossRef]
- Hattar, S.; Kumar, M.; Park, A.; Tong, P.; Tung, J.; Yau, K.; Berson, D.M. Central Projections of Melanopsin- Expressing Retinal Ganglion Cells in the Mouse. J. Comp. Neurol. 2006, 497, 326–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gooley, J.J.; Lu, J.; Fischer, D.; Saper, C.B. A broad role for melanopsin in nonvisual photoreception. J. Neurosci. 2003, 23, 7093–7106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hattar, S.; Liao, H.W.; Takao, M.; Berson, D.M.; Yau, K.W. Melanopsin-containing retinal ganglion cells: Architecture, projections, and intrinsic photosensitivity. Science 2002, 295, 1065–1070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.K.; Badea, T.C.; Hattar, S. Photoentrainment and pupillary light reflex are mediated by distinct populations of ipRGCs. Nature 2011, 476, 92–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panda, S.; Provencio, I.; Tu, D.C.; Pires, S.S.; Rollag, M.D.; Castrucci, A.M.; Pletcher, M.T.; Sato, T.K.; Wiltshire, T.; Andahazy, M.; et al. Melanopsin Is Required for Non–Image-Forming Photic Responses in Blind Mice. Science 2003, 301, 1488–1490. [Google Scholar] [CrossRef] [Green Version]
- Jones, K.A.; Hatori, M.; Mure, L.S.; Bramley, J.R.; Artymyshyn, R.; Hong, S.-P.; Marzabadi, M.; Zhong, H.; Sprouse, J.; Zhu, Q.; et al. Small-molecule antagonists of melanopsin-mediated phototransduction. Nat. Chem. Biol. 2013, 9, 630–635. [Google Scholar] [CrossRef] [PubMed]
- Hattar, S.; Lucas, R.J.; Mrosovsky, N.; Thompson, S.; Douglas, R.H.; Hankins, M.W.; Lem, J.; Biel, M.; Hofmann, F.; Foster, R.G.; et al. Melanopsin and rod—cone photoreceptive systems account for all major accessory visual functions in mice. Nature 2003, 424, 76–81. [Google Scholar]
- Güler, A.D.; Ecker, J.L.; Lall, G.S.; Haq, S.; Altimus, C.M.; Liao, W.; Barnard, A.R.; Cahill, H.; Badea, T.C.; Zhao, H.; et al. Melanopsin cells are the principal conduits for rod/cone input to non-image forming vision. Nature 2008, 453, 102–105. [Google Scholar] [CrossRef] [Green Version]
- McDougal, D.H.; Gamlin, P.D. The influence of intrinsically-photosensitive retinal ganglion cells on the spectral sensitivity and response dynamics of the human pupillary light reflex. Vis. Res. 2010, 50, 72–87. [Google Scholar] [CrossRef] [Green Version]
- Keenan, W.T.; Rupp, A.C.; Ross, R.A.; Somasundaram, P.; Hiriyanna, S.; Wu, Z.; Badea, T.C.; Robinson, P.R.; Lowell, B.B.; Hattar, S.S. A visual circuit uses complementary mechanisms to support transient and sustained pupil constriction. eLife 2016, 5, e15392. [Google Scholar] [CrossRef]
- Gooley, J.J.; Mien, I.H.; Hilaire, M.A.S.; Yeo, S.-C.; Chua, E.C.-P.; van Reen, E.; Hanley, C.J.; Hull, J.T.; Czeisler, C.A.; Lockley, S.W. Melanopsin and Rod-Cone Photoreceptors Play Different Roles in Mediating Pupillary Light Responses during Exposure to Continuous Light in Humans. J. Neurosci. 2012, 32, 14242–14253. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, S.K.; Witkovsky, P.; Roux, M.J.; Low, M.J.; Otero-Corchon, V.; Hentges, S.T.; Vigh, J. β-Endorphin expression in the mouse retina. J. Comp. Neurol. 2010, 3148, 3130–3148. [Google Scholar] [CrossRef] [Green Version]
- Cleymaet, A.M.; Gallagher, S.K.; Tooker, R.E.; Lipin, M.Y.; Renna, J.M.; Sodhi, P.; Berg, D.; Hartwick, A.T.E.; Berson, D.M.; Vigh, J. μ-Opioid Receptor Activation Directly Modulates Intrinsically Photosensitive Retinal Ganglion Cells. Neuroscience 2019, 408, 400–417. [Google Scholar] [CrossRef] [PubMed]
- Wyman, J.; Bultman, S. Postmortem distribution of heroin metabolites in femoral blood, liver, cerebrospinal fluid, and vitreous humor. J. Anal. Toxicol. 2004, 28, 260–263. [Google Scholar] [CrossRef] [PubMed]
- Fernández, P.; Seoane, S.; Vázquez, C.; Tabernero, M.J.; Carro, A.M.; Lorenzo, R.A. Chromatographic determination of drugs of abuse in vitreous humor using solid-phase extraction. J. Appl. Toxicol. 2013, 33, 740–745. [Google Scholar] [CrossRef]
- Busskamp, V.; Duebel, J.; Balya, D.; Fradot, M.; Viney, T.J.; Siegert, S.; Groner, A.C.; Cabuy, E.; Forster, V.; Seeliger, M.; et al. Genetic reactivation of cone photoreceptors restores visual responses in retinitis pigmentosa. Science 2010, 329, 413–417. [Google Scholar] [CrossRef] [Green Version]
- Gamlin, P.D.R.; McDougal, D.H.; Pokorny, J.; Smith, V.C.; Yau, K.W.; Dacey, D.M. Human and macaque pupil responses driven by melanopsin-containing retinal ganglion cells. Vis. Res. 2007, 47, 946–954. [Google Scholar] [CrossRef] [Green Version]
- Adhikari, P.; Zele, A.J.; Feigl, B. The post-illumination pupil response (PIPR). Investig. Ophthalmol. Vis. Sci. 2015, 56, 3838–3849. [Google Scholar] [CrossRef] [Green Version]
- Wong, K.Y.; Dunn, F.A.; Graham, D.M.; Berson, D.M. Synaptic influences on rat ganglion-cell photoreceptors. J. Physiol. 2007, 582, 279–296. [Google Scholar] [CrossRef]
- Tortella, F.C.; Cowan, A.; Adler, M.W. Pupillary effects of leucine and methionine enkephalin in rats after intraperitoneal administration. Peptides 1980, 1, 237–241. [Google Scholar] [CrossRef]
- Gallagher, S.K.; Anglen, J.N.; Mower, J.M.; Vigh, J. Dopaminergic amacrine cells express opioid receptors in the mouse retina. Vis. Neurosci. 2012, 29, 203–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korczyn, A.D.; Eshel, Y.; Keren, O. Enkephalin mydriasis in mice. Eur. J. Pharmacol. 1980, 65, 285–287. [Google Scholar] [CrossRef]
- Pan, H.L.; Wu, Z.Z.; Zhou, H.Y.; Chen, S.R.; Zhang, H.M.; Li, D.P. Modulation of pain transmission by G-protein-coupled receptors. Pharmacol. Ther. 2008, 117, 141–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Britto, L.R.G.; Hamassaki-Britto, D.E. Enkephalin-immunoreactive ganglion cells in the pigeon retina. Vis. Neurosci. 1992, 9, 389–398. [Google Scholar] [CrossRef]
- Altschuler, R.A.; Mosinger, J.L.; Hoffman, D.W.; Parakkal, M.H. Immunocytochemical localization of enkephalin-like immunoreactivity in the retina of the guinea pig. Proc. Natl. Acad. Sci. USA 1982, 79, 2398–2400. [Google Scholar] [CrossRef] [Green Version]
- Slaughter, M.M.; Mattler, J.A.; Gottlieb, D.I. Opiate binding sites in the chick, rabbit and goldfish retina. Brain Res. 1985, 339, 39–47. [Google Scholar] [CrossRef]
- Howells, R.; Gorth, J.; Hiller, J.; Simon, E. Opiate binding sites in the retina: Properties and distribution. J. Pharmacol. Exp. Ther. 1980, 215, 60–64. [Google Scholar]
- Robin, M.; Kirby, A.; Messner, S.; Geller, E.B.; Adler, M.W. Differentiating opioids by their pupillary effects in the rat. Life Sci. 1985, 36, 1669–1677. [Google Scholar] [CrossRef]
- Kieffer, B.L. Recent advances in molecular recognition and signal transduction of active peptides: Receptors for opioid peptides. Cell. Mol. Neurobiol. 1995, 15, 615–635. [Google Scholar] [CrossRef]
- Do, M.T.H.; Kang, S.H.; Xue, T.; Zhong, H.; Liao, H.; Bergles, D.E.; Yau, K.-W. Photon capture and signalling by melanopsin retinal ganglion cells. Nature 2009, 457, 281–287. [Google Scholar] [CrossRef] [Green Version]
- Morgan, I.G.; Boelen, M.K. A retinal dark-light switch: A review of the evidence. Vis. Neurosci. 1996, 13, 399–409. [Google Scholar] [CrossRef] [PubMed]
- Witkovsky, P. Dopamine and retinal function. Doc. Ophthalmol. 2004, 108, 17–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas Su, Y.Y.; Watt, C.B. Interaction between enkephalin and dopamine in the avian retina. Brain Res. 1987, 423, 63–70. [Google Scholar] [CrossRef]
- Kolbinger, W.; Weiler, R. Modulation of endogenous dopamine release in the turtle retina: Effects of light, calcium, and neurotransmitters. Vis. Neurosci. 1993, 10, 1035–1041. [Google Scholar] [CrossRef] [PubMed]
- Wesche, D.L.; Frederickson, R.C.A. Diurnal differences in opioid peptide levels correlated with nociceptive sensitivity. Life Sci. 1979, 24, 1861–1867. [Google Scholar] [CrossRef]
- Naber, D.; Wirz-Justice, A.; Kafka, M.S. Circadian rhythm in rat brain opiate receptor. Neurosci. Lett. 1981, 21, 45–50. [Google Scholar] [CrossRef]
- Zele, A.J.; Feigl, B.; Smith, S.S.; Markwell, E.L. The circadian response of intrinsically photosensitive retinal ganglion cells. PLoS ONE 2011, 6, e17860. [Google Scholar] [CrossRef] [Green Version]
- Bonfiglio, V.; Bucolo, C.; Camillieri, G.; Drago, F. Possible involvement of nitric oxide in morphine-induced miosis and reduction of intraocular pressure in rabbits. Eur. J. Pharmacol. 2006, 534, 227–232. [Google Scholar] [CrossRef]
- Wang, Q.; Wing, W.; Yue, S.; Jiang, Z.; Mikoshiba, K.; Offermanns, S.; Yau Correspondence, K.-W.; Xue, T.; Kang, S.H.; Bergles, D.E.; et al. Synergistic Signaling by Light and Acetylcholine in Mouse Iris Sphincter Muscle. Curr. Biol. 2017, 27, 1791–1799.e5. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, T.M.; Rupp, A.C.; Yungher, B.; Cui, Y.; Wess, J.; Park, K.; Hattar, S. A retinal projection to the iris mediates pupil constriction. In Proceedings of the ARVO Annual Meeting 2014—Investigative Ophthalmology & Visual Science, Orlando, FL, USA, 4–8 May 2014; Volume 55, p. 1231. [Google Scholar]
- Semo, M.; Gias, C.; Ahmado, A.; Vugler, A. A role for the ciliary marginal zone in the melanopsin-dependent intrinsic pupillary light reflex. Exp. Eye Res. 2014, 119, 8–18. [Google Scholar] [CrossRef] [Green Version]
- Xue, T.; Do, M.T.H.; Riccio, A.; Jiang, Z.; Hsieh, J.; Wang, H.C.; Merbs, S.L.; Welsbie, D.S.; Yoshioka, T.; Weissgerber, P.; et al. Melanopsin signalling in mammalian iris and retina. Nature 2011, 479, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Rupp, A.; Schmidt, T.; Chew, K.; Yungher, B.; Park, K.; Hattar, S. ipRGCs mediate ipsilateral pupil constriction. In Proceedings of the ARVO Annual Meeting 2013—Investigative Ophthalmology & Visual Science, Seatle, WA, USA, 5–9 May 2013; Volume 54, p. 310. [Google Scholar]
- Bucolo, C.; Leggio, G.M.; Drago, F.; Salomone, S. Dopamine outside the brain: The eye, cardiovascular system and endocrine pancreas. Pharmacol. Ther. 2019, 203, 107392. [Google Scholar] [CrossRef] [PubMed]
- Selley, D.E.; Cao, C.C.; Sexton, T.; Schwegel, J.A.; Martin, T.J.; Childers, S.R. μ Opioid receptor-mediated G-protein activation by heroin metabolites: Evidence for greater efficacy of 6-monoacetylmorphine compared with morphine. Biochem. Pharmacol. 2001, 62, 447–455. [Google Scholar] [CrossRef]
- Lee, C.W.S.; Yan, J.Y.; Chiang, Y.C.; Hung, T.W.; Wang, H.L.; Chiou, L.C.; Ho, I.K. Differential pharmacological actions of methadone and buprenorphine in human embryonic kidney 293 cells coexpressing human μ-opioid and opioid receptor-like 1 receptors. Neurochem. Res. 2011, 36, 2008–2021. [Google Scholar] [CrossRef] [Green Version]
- Saszik, S.M.; Robson, J.G.; Frishman, L.J. The Scotopic Threshold Response of the Dark-Adapted Electroretinogram of the Mouse. J. Physiol. 2002, 543, 899–916. [Google Scholar] [CrossRef] [Green Version]
- Dortch-Carnes, J.; Russell, K.R.M. Morphine-induced reduction of intraocular pressure and pupil diameter: Role of nitric oxide. Pharmacology 2006, 77, 17–24. [Google Scholar] [CrossRef]
- Dortch-Carnes, J.; Russell, K. Morphine-stimulated nitric oxide release in rabbit aqueous humor. Exp. Eye Res. 2007, 84, 185–190. [Google Scholar] [CrossRef] [Green Version]
- Stagni, E.; Bucolo, C.; Motterlini, R.; Drago, F. Morphine-induced ocular hypotension is modulated by nitric oxide and carbon monoxide: Role of μ 3 receptors. J. Ocul. Pharmacol. Ther. 2010, 26, 31–36. [Google Scholar] [CrossRef]
- Wang, D.; Potter, D.E. Ocular action of an opioid peptide, DPDPE. J. Ocul. Pharmacol. Ther. 1996, 12, 131–139. [Google Scholar] [CrossRef]
- Russell, K.R.M.; Wang, D.R.; Potter, D.E. Modulation of ocular hydrodynamics and iris function by bremazocine, a kappa opioid receptor agonist. Exp. Eye Res. 2000, 70, 675–682. [Google Scholar] [CrossRef]
- Cadet, P.; Mantione, K.J.; Stefano, G.B. Molecular Identification and Functional Expression of μ 3, a Novel Alternatively Spliced Variant of the Human μ Opiate Receptor Gene. J. Immunol. 2003, 170, 5118–5123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.C.; Moura, A.L.; Raza, A.S.; Rhee, D.W.; Kardon, R.H.; Hood, D.C. Toward a clinical protocol for assessing rod, cone, and melanopsin contributions to the human pupil response. Investig. Ophthalmol. Vis. Sci. 2011, 52, 6624–6635. [Google Scholar] [CrossRef] [PubMed]
- Yeh, C.Y.; Koehl, K.L.; Harman, C.D.; Iwabe, S.; Guzman, J.M.; Petersen-Jones, S.M.; Kardon, R.H.; Komáromy, A.M. Assessment of rod, cone, and intrinsically photosensitive retinal ganglion cell contributions to the canine chromatic pupillary response. Investig. Ophthalmol. Vis. Sci. 2017, 58, 65–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rukmini, A.V.; Milea, D.; Baskaran, M.; How, A.C.; Perera, S.A.; Aung, T.; Gooley, J.J. Pupillary Responses to High-Irradiance Blue Light Correlate with Glaucoma Severity. Ophthalmology 2015, 122, 1777–1785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laurenzo, S.A.; Kardon, R.; Ledolter, J.; Poolman, P.; Schumacher, A.M.; Potash, J.B.; Full, J.M.; Rice, O.; Ketcham, A.; Starkey, C.; et al. Pupillary response abnormalities in depressive disorders. Psychiatry Res. 2016, 246, 492–499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neice, A.E.; Behrends, M.; Bokoch, M.P.; Seligman, K.M.; Conrad, N.M.; Larson, M.D. Prediction of Opioid Analgesic Efficacy by Measurement of Pupillary Unrest. Anesth. Analg. 2017, 124, 915–921. [Google Scholar] [CrossRef]
- Kirkpatrick, S.L.; Bryant, C.D. Behavioral architecture of opioid reward and aversion in C57BL/6 substrains. Front. Behav. Neurosci. 2015, 8, 450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weibel, R.; Reiss, D.; Karchewski, L.; Gardon, O.; Matifas, A.; Filliol, D.; Becker, J.A.J.; Wood, J.N.; Kieffer, B.L.; Gaveriaux-Ruff, C. Mu Opioid Receptors on Primary Afferent Nav1.8 Neurons Contribute to Opiate-Induced Analgesia: Insight from Conditional Knockout Mice. PLoS ONE 2013, 8, e74706. [Google Scholar] [CrossRef] [Green Version]
- Severino, A.L.; Mittal, N.; Hakimian, J.K.; Velarde, N.; Minasyan, A.; Albert, R.; Torres, C.; Romaneschi, N.; Johnston, C.; Tiwari, S.; et al. μ-Opioid Receptors on Distinct Neuronal Populations Mediate Different Aspects of Opioid Reward-Related Behaviors. eNeuro 2020, 7. [Google Scholar] [CrossRef]
- Mohan, K.; Harper, M.M.; Kecova, H.; Ye, E.-A.; Lazic, T.; Sakaguchi, D.S.; Kardon, R.H.; Grozdanic, S.D. Characterization of structure and function of the mouse retina using pattern electroretinography, pupil light reflex, and optical coherence tomography. Vet. Ophthalmol. 2012, 15, 94–104. [Google Scholar] [CrossRef]
- Krizaj, D.; Hawlina, M. Cellular basis for rod-cone interactions in the outer retina. Zdrav. Vestn. 2002, 71, II-137. [Google Scholar]
- Kostic, C.; Crippa, S.V.; Martin, C.; Kardon, R.H.; Biel, M.; Arsenijevic, Y.; Kawasaki, A. Determination of Rod and Cone Influence to the Early and Late Dynamic of the Pupillary Light Response. Investig. Ophthalmol. Vis. Sci. 2016, 57, 2501–2508. [Google Scholar] [CrossRef] [PubMed]
- Mojumder, D.K.; Qian, Y.; Wensel, T.G. Two R7 Regulator of G-Protein Signaling Proteins Shape Retinal Bipolar Cell Signaling. J. Neurosci. 2009, 29, 7753–7765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rappoport, D.; Morzaev, D.; Weiss, S.; Vieyra, M.; Nicholson, J.D.; Leiba, H.; Goldenberg-Cohen, N. Effect of intravitreal injection of bevacizumab on optic nerve head leakage and retinal ganglion cell survival in a mouse model of optic nerve crush. Investig. Ophthalmol. Vis. Sci. 2013, 54, 8160–8171. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cleymaet, A.M.; Berezin, C.-T.; Vigh, J. Endogenous Opioid Signaling in the Mouse Retina Modulates Pupillary Light Reflex. Int. J. Mol. Sci. 2021, 22, 554. https://doi.org/10.3390/ijms22020554
Cleymaet AM, Berezin C-T, Vigh J. Endogenous Opioid Signaling in the Mouse Retina Modulates Pupillary Light Reflex. International Journal of Molecular Sciences. 2021; 22(2):554. https://doi.org/10.3390/ijms22020554
Chicago/Turabian StyleCleymaet, Allison M., Casey-Tyler Berezin, and Jozsef Vigh. 2021. "Endogenous Opioid Signaling in the Mouse Retina Modulates Pupillary Light Reflex" International Journal of Molecular Sciences 22, no. 2: 554. https://doi.org/10.3390/ijms22020554
APA StyleCleymaet, A. M., Berezin, C. -T., & Vigh, J. (2021). Endogenous Opioid Signaling in the Mouse Retina Modulates Pupillary Light Reflex. International Journal of Molecular Sciences, 22(2), 554. https://doi.org/10.3390/ijms22020554